
The Rhizobacterium Pseudomonas alcaligenes AVO110 Induces the Expression of Biofilm-Related Genes in Response to Rosellinia necatrix Exudates

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Media, and Growth Conditions
2.2. Construction of Bacterial Strains and Plasmids
2.3. Complete Sequencing of the P. alcaligenes AVO110 Genome
2.4. Bioinformatics Methods
2.5. RNA Techniques
2.6. Biofilm Assays
3. Results
3.1. Sequencing and General Features of P. alcaligenes AVO110 Genome
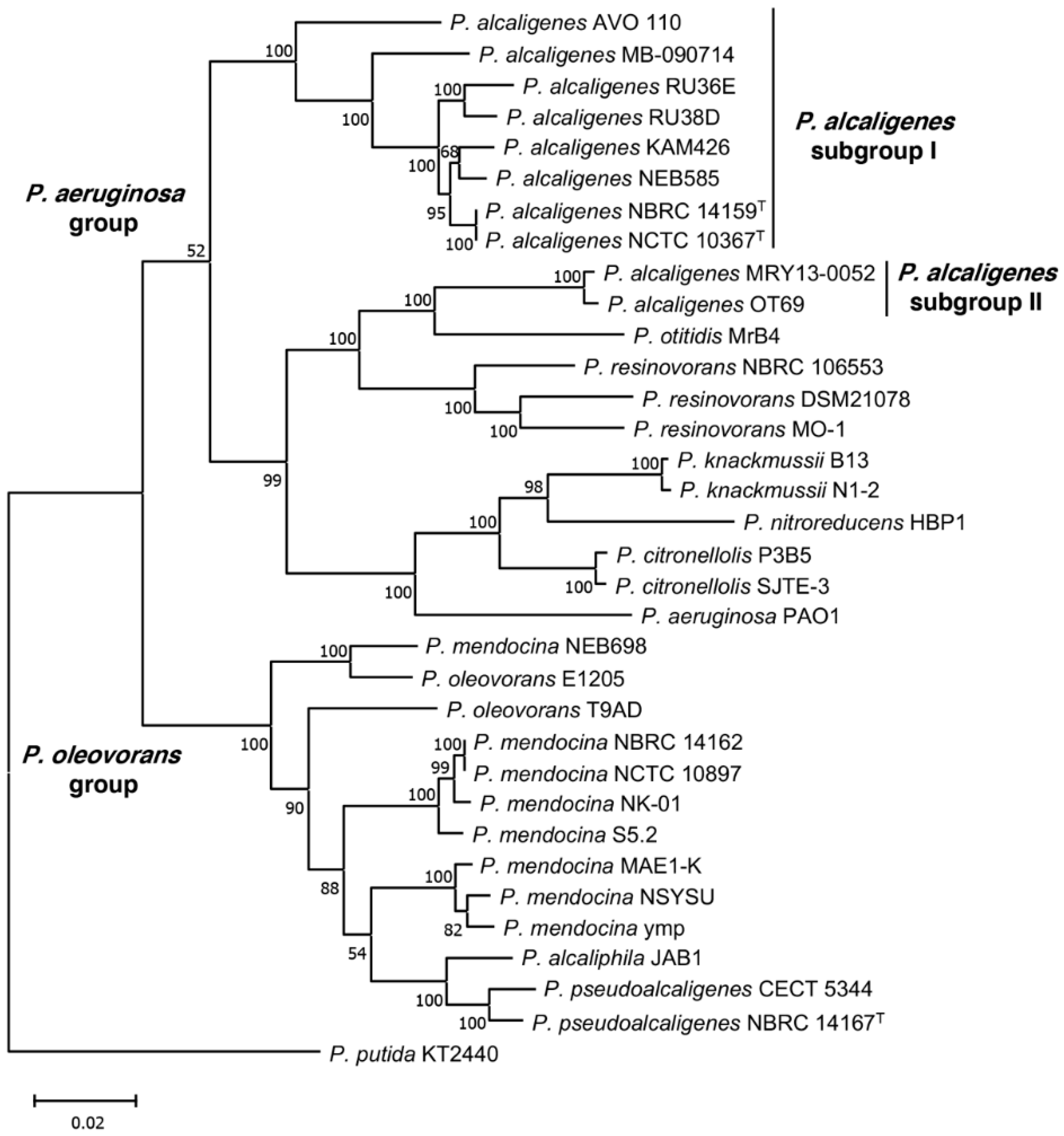
3.2. Phylogenetic Analysis of P. alcaligenes and P. pseudoalcaligenes Strains

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain a | Subgroup b | Genome Size (Mb) | G+C Content (%) | Coding Sequences | Scaffolds (Number) | Coverage | Source of Isolation | Year of Isolation | Accession Number c | Reference or Sequence Source/Year |
|---|---|---|---|---|---|---|---|---|---|---|
| Pseudomonas alcaligenes | ||||||||||
| NCTC 10367 T | I | 5.02 | 64.5 | 4594 | 2 | 100× | Swimming pool water | 1961 | UGUP00000000 | [63] |
| NBRC 14159 T | I | 4.82 | 64.8 | 4445 | 122 | 131× | Swimming pool water | 1961 | BATI00000000 | [63] |
| NEB 585 | I | 4.41 | 65.5 | 4072 | 1 | 382× | Water sample | 1989 | CP014784 | R. D. Morgan/2016 |
| AVO110 | I | 4.97 | 64.9 | 4406 | 3 | 100× | Avocado rhizosphere | 2005 | LZEU00000000 | This study |
| MB-090714 | I | 4.01 | 66.5 | 3815 | 16 | 92.7× | Water (lake) | 2009 | QJRX00000000 | M. Batrich/2018 |
| Bin_52_1 | I | 3.68 | 64.6 | 3700 | 325 | 978.2× | Water purification facility | 2017 | SSFO00000000 | B.W. Stamps/2018 |
| KAM 426 | I | 4.68 | 64.8 | 4363 | 1 | 93× | Wastewater | 2020 | AP024354 | M. Suzuki/2021 |
| RU38D | I | 4.25 | 65.1 | 3920 | 10 | 262× | Water (duckweeds) | NA | FTNW00000000 | S. Lebeis /2016 |
| RU36E | I | 4.63 | 64.1 | 4246 | 23 | 221× | Water (duckweeds) | NA | FTMP00000000 | S. Lebeis /2016 |
| OT 69 | II | 7.03 | 66.0 | 6379 | 223 | 200× | Oyster tissue | 2013 | ATCP00000000 | [64] |
| MRY13-0052 | II | 6.88 | 65.8 | 6129 | 237 | 16.1× | Blood infection | 2013 | BATO00000000 | [65] |
| Pseudomonas pseudoalcaligenes | ||||||||||
| NBRC 14167 T | NAP | 4.70 | 62.2 | 4200 | 204 | 131× | Sinus drainage | 1982 | BDAJ00000000 | D. Wibberg/2013 |
| CECT 5344 | NAP | 4.69 | 62.3 | 4082 | 1 | 40× | Water (river) | 2005 | HG916826 | A. Hosoyama/2016 |
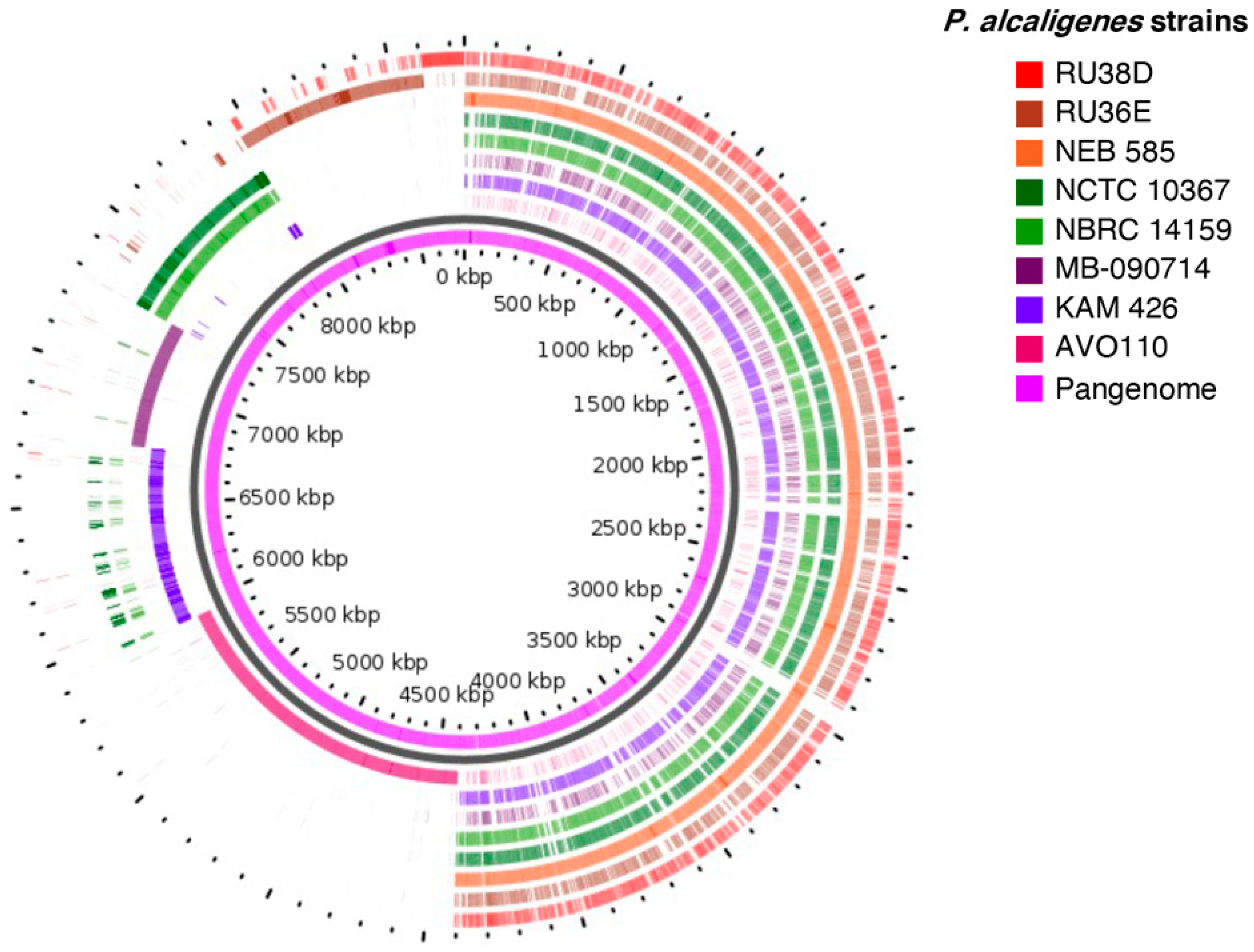
3.3. Pan-Genome and Core Genome of P. alcaligenes
3.4. Identification of P. alcaligenes AVO110 Genes Transcriptionally Regulated by R. necatrix Exudates
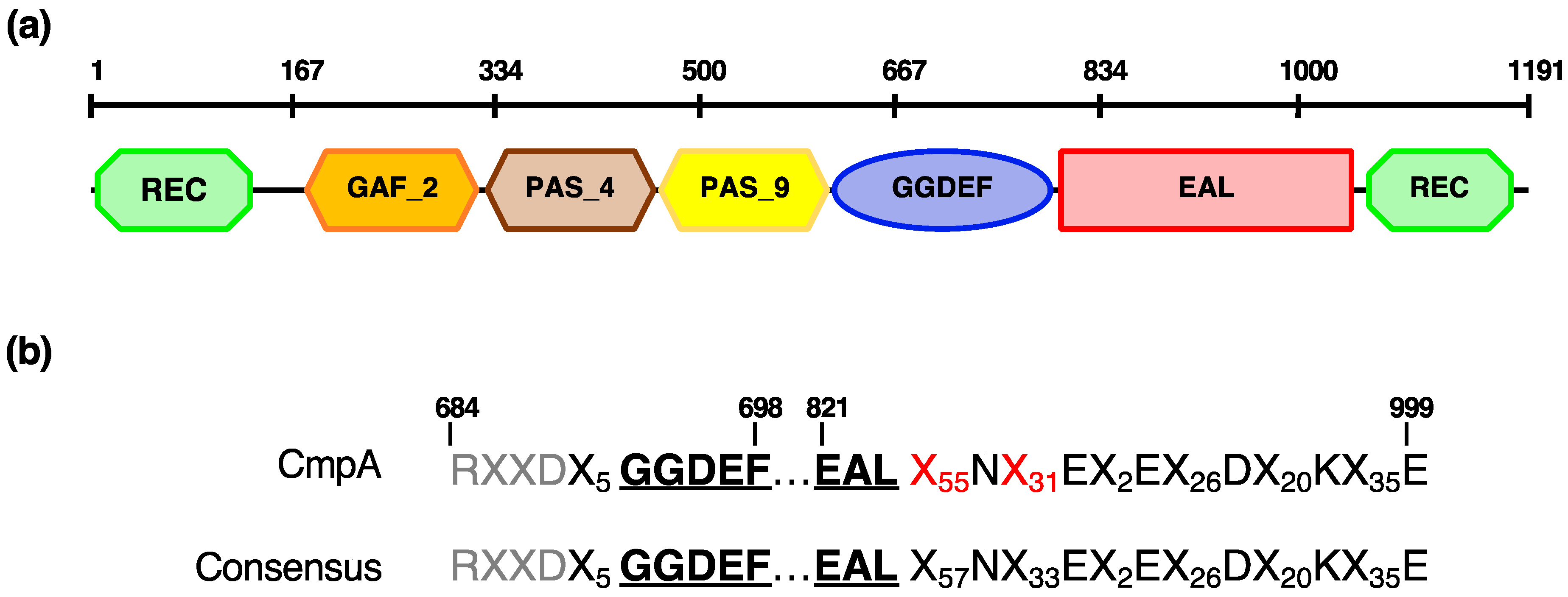
3.5. CmpA, a Pseudomonas spp.-Specific Multidomain Protein
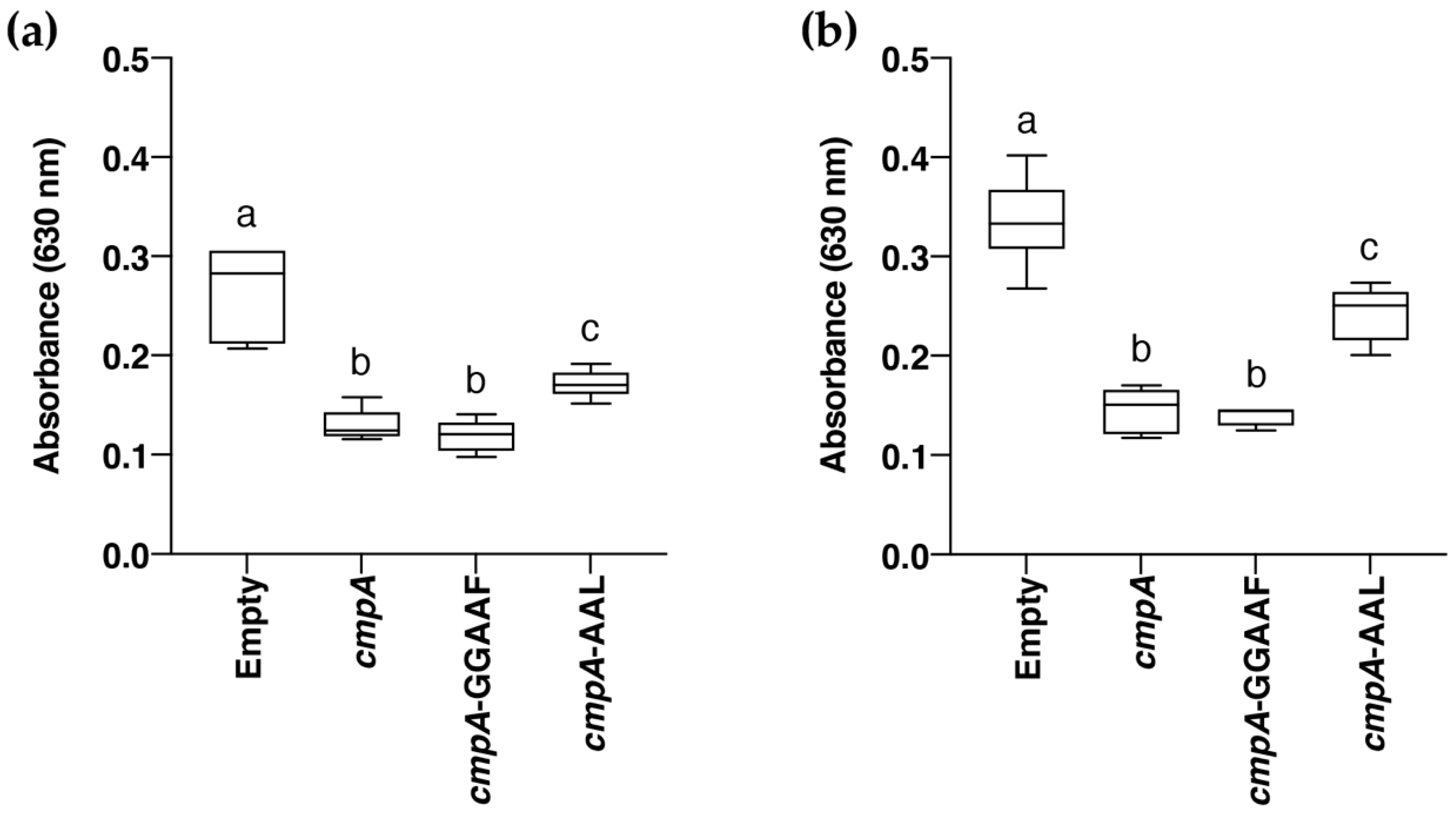
3.6. Role of CmpA in Biofilm Formation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pliego, C.; de Weert, S.; Lamers, G.; de Vicente, A.; Bloemberg, G.; Cazorla, F.M.; Ramos, C. Two similar enhanced root-colonizing Pseudomonas strains differ largely in their colonization strategies of avocado roots and Rosellinia necatrix hyphae. Environ. Microbiol. 2008, 10, 3295–3304. [Google Scholar] [CrossRef]
- Calderón, C.E.; de Vicente, A.; Cazorla, F.M. Role of 2-hexyl, 5-propyl resorcinol production by Pseudomonas chlororaphis PCL1606 in the multitrophic interactions in the avocado rhizosphere during the biocontrol process. FEMS Microbiol. Ecol. 2014, 89, 20–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubey, R.K.; Tripathi, V.; Dubey, P.K.; Singh, H.B.; Abhilash, P.C. Exploring rhizospheric interactions for agricultural sustainability: The need of integrative research on multi-trophic interactions. J. Clean. Prod. 2016, 115, 362–365. [Google Scholar] [CrossRef]
- Tamborindeguy, C.; Huot, O.B.; Ibanez, F.; Levy, J. The influence of bacteria on multitrophic interactions among plants, psyllids, and pathogen. Insect Sci. 2017, 24, 961–974. [Google Scholar] [CrossRef] [PubMed]
- Rossmann, M.; Pérez-Jaramillo, J.E.; Kavamura, V.N.; Chiaramonte, J.B.; Dumack, K.; Fiore-Donno, A.M.; Mendes, L.W.; Ferreira, M.M.C.; Bonkowski, M.; Raaijmakers, J.M.; et al. Multitrophic interactions in the rhizosphere microbiome of wheat: From bacteria and fungi to protists. FEMS Microbiol. Ecol. 2020, 96. [Google Scholar] [CrossRef] [PubMed]
- Braga, R.M.; Dourado, M.N.; Araújo, W.L. Microbial interactions: Ecology in a molecular perspective. Braz. J. Microbiol. 2016, 47, 86–98. [Google Scholar] [CrossRef] [Green Version]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moënne-Loccoz, Y. The rhizosphere: A playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 2009, 321, 341–361. [Google Scholar] [CrossRef] [Green Version]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef]
- Berendsen, R.L.; Pieterse, C.M.J.; Bakker, P.A.H.M. The rhizosphere microbiome and plant health. Trends Plant Sci. 2012, 17, 478–486. [Google Scholar] [CrossRef]
- Hu, L.; Robert, C.A.M.; Cadot, S.; Zhang, X.; Ye, M.; Li, B.; Manzo, D.; Chervet, N.; Steinger, T.; Van Der Heijden, M.G.A.; et al. Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Zhalnina, K.; Louie, K.B.; Hao, Z.; Mansoori, N.; Da Rocha, U.N.; Shi, S.; Cho, H.; Karaoz, U.; Loqué, D.; Bowen, B.P.; et al. Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly. Nat. Microbiol. 2018, 3, 470–480. [Google Scholar] [CrossRef] [Green Version]
- Leveau, J.H.J.; Preston, G.M. Bacterial mycophagy: Definition and diagnosis of a unique bacterial–fungal interaction. New Phytol. 2008, 177, 859–876. [Google Scholar] [CrossRef]
- De Boer, W.; Folman, L.B.; Summerbell, R.C.; Boddy, L. Living in a fungal world: Impact of fungi on soil bacterial niche development. FEMS Microbiol. Rev. 2005, 29, 795–811. [Google Scholar] [CrossRef] [Green Version]
- Höppener-Ogawa, S.; Leveau, J.H.J.; Van Veen, J.A.; De Boer, W. Mycophagous growth of Collimonas bacteria in natural soils, impact on fungal biomass turnover and interactions with mycophagous Trichoderma Fungi. ISME J. 2009, 3, 190–198. [Google Scholar] [CrossRef] [Green Version]
- Leveau, J.H.J.; Uroz, S.; de Boer, W. The bacterial genus Collimonas: Mycophagy, weathering and other adaptive solutions to life in oligotrophic soil environments. Environ. Microbiol. 2010, 12, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Mela, F.; Fritsche, K.; De Boer, W.; Van Veen, J.A.; De Graaff, L.H.; Van Den Berg, M.; Leveau, J.H.J. Dual transcriptional profiling of a bacterial/fungal confrontation: Collimonas fungivorans versus Aspergillus niger. ISME J. 2011, 5, 1494–1504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mela, F.; Fritsche, K.; De Boer, W.; Van den Berg, M.; Van Veen, J.A.; Maharaj, N.N.; Leveau, J.H.J. Comparative genomics of bacteria from the genus Collimonas: Linking (dis)similarities in gene content to phenotypic variation and conservation. Environ. Microbiol. Rep. 2012, 4, 424–432. [Google Scholar] [CrossRef] [Green Version]
- Garbeva, P.; Hordijk, C.; Gerards, S.; de Boer, W. Volatiles produced by the mycophagous soil bacterium Collimonas. FEMS Microbiol. Ecol. 2014, 87, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Ballhausen, M.B.; Van Veen, J.A.; Hundscheid, M.P.J.; De Boer, W. Methods for baiting and enriching fungus-feeding (Mycophagous) rhizosphere bacteria. Front. Microbiol. 2015, 6, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, C.; Schmidt, R.; de Jager, V.; Krzyzanowska, D.; Jongedijk, E.; Cankar, K.; Beekwilder, J.; van Veen, A.; de Boer, W.; van Veen, J.A.; et al. Exploring the genomic traits of fungus-feeding bacterial genus Collimonas. BMC Genom. 2015, 16, 1103. [Google Scholar] [CrossRef] [PubMed]
- Rudnick, M.B.; van Veen, J.A.; de Boer, W. Baiting of rhizosphere bacteria with hyphae of common soil fungi reveals a diverse group of potentially mycophagous secondary consumers. Soil Biol. Biochem. 2015, 88, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Pliego, C.; Cazorla, F.M.; González-Sánchez, M.Á.; Pérez-Jiménez, R.M.; de Vicente, A.; Ramos, C. Selection for biocontrol bacteria antagonistic toward Rosellinia necatrix by enrichment of competitive avocado root tip colonizers. Res. Microbiol. 2007, 158, 463–470. [Google Scholar] [CrossRef] [PubMed]
- De Weert, S.; Vermeiren, H.; Mulders, I.H.M.; Kuiper, I.; Hendrickx, N.; Bloemberg, G.V.; Vanderleyden, J.; De Mot, R.; Lugtenberg, B.J.J. Flagella-driven chemotaxis towards exudate components is an important trait for tomato root colonization by Pseudomonas fluorescens. Mol. Plant-Microbe Interact. 2002, 15, 1173–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Granero, F.; Rivilla, R.; Martín, M. Rhizosphere selection of highly motile phenotypic variants of Pseudomonas fluorescens with enhanced competitive colonization ability. Appl. Environ. Microbiol. 2006, 72, 3429–3434. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.W.; Cooksey, D.A. Genes expressed in Pseudomonas putida during colonization of a plant-pathogenic fungus. Appl. Environ. Microbiol. 2000, 66, 2764–2772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, S.J.; Yang, C.H.; Cooksey, D.A. Pseudomonas putida 06909 genes expressed during colonization on mycelial surfaces and phenotypic characterization of mutants. J. Appl. Microbiol. 2007, 103, 120–132. [Google Scholar] [CrossRef]
- Barret, M.; Frey-Klett, P.; Boutin, M.; Guillerm-Erckelboudt, A.Y.; Martin, F.; Guillot, L.; Sarniguet, A. The plant pathogenic fungus Gaeumannomyces graminis var. tritici improves bacterial growth and triggers early gene regulations in the biocontrol strain Pseudomonas fluorescens Pf29Arp. New Phytol. 2009, 181, 435–447. [Google Scholar] [CrossRef]
- Pliego, C.; Crespo-Gómez, J.I.; Pintado, A.; Pérez-Martínez, I.; de Vicente, A.; Cazorla, F.M.; Ramos, C. Response of the biocontrol agent Pseudomonas pseudoalcaligenes AVO110 to Rosellinia necatrix exudate. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef] [Green Version]
- Holden, D.W.; Hensel, M. 7.3 Signature Tagged Mutagenesis. Methods Microbiol. 1998, 27, 359–370. [Google Scholar] [CrossRef]
- Stapper, A.P.; Narasimhan, G.; Ohman, D.E.; Barakat, J.; Hentzer, M.; Molin, S.; Kharazmi, A.; Høiby, N.; Mathee, K. Alginate production affects Pseudomonas aeruginosa biofilm development and architecture, but is not essential for biofilm formation. J. Med. Microbiol. 2004, 53, 679–690. [Google Scholar] [CrossRef]
- Matilla, M.A.; Travieso, M.L.; Ramos, J.L.; Ramos-González, M.I. Cyclic diguanylate turnover mediated by the sole GGDEF/EAL response regulator in Pseudomonas putida: Its role in the rhizosphere and an analysis of its target processes. Environ. Microbiol. 2011, 13, 1745–1766. [Google Scholar] [CrossRef]
- Muriel, C.; Arrebola, E.; Redondo-Nieto, M.; Martínez-Granero, F.; Jalvo, B.; Pfeilmeier, S.; Blanco-Romero, E.; Baena, I.; Malone, J.G.; Rivilla, R.; et al. AmrZ is a major determinant of c-di-GMP levels in Pseudomonas fluorescens F113. Sci. Rep. 2018, 8, 1979. [Google Scholar] [CrossRef] [Green Version]
- Simm, R.; Morr, M.; Kader, A.; Nimtz, M.; Römling, U. GGDEF and EAL domains inversely regulate cyclic di-GMP levels and transition from sessibility to motility. Mol. Microbiol. 2004, 53, 1123–1134. [Google Scholar] [CrossRef]
- Romling, U.; Galperin, M.Y.; Gomelsky, M. Cyclic di-GMP: The First 25 Years of a Universal Bacterial Second Messenger. Microbiol. Mol. Biol. Rev. 2013, 77, 1–52. [Google Scholar] [CrossRef] [Green Version]
- Jenal, U.; Reinders, A.; Lori, C. Cyclic di-GMP: Second messenger extraordinaire. Nat. Rev. Microbiol. 2017, 15, 271–284. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Gil, M.; Ramos, C. Role of cyclic di-GMP in the bacterial virulence and evasion of the plant immunity. Curr. Issues Mol. Biol. 2018, 25, 199–221. [Google Scholar] [CrossRef]
- Ross, P.; Weinhouse, H.; Aloni, Y.; Michaeli, D.; Weinberger-Ohana, P.; Mayer, R.; Braun, S.; De Vroom, E.; Van Der Marel, G.A.; Van Boom, J.H.; et al. Regulation of cellulose synthesis in Acetobacter xylinum by cyclic diguanylic acid. Nature 1987, 325, 279–281. [Google Scholar] [CrossRef] [PubMed]
- Ryan, R.P.; Fouhy, Y.; Lucey, J.F.; Crossman, L.C.; Spiro, S.; He, Y.W.; Zhang, L.H.; Heeb, S.; Cámara, M.; Williams, P.; et al. Cell-cell signaling in Xanthomonas campestris involves an HD-GYP domain protein that functions in cyclic di-GMP turnover. Proc. Natl. Acad. Sci. USA 2006, 103, 6712–6717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sondermann, H.; Shikuma, N.J.; Yildiz, F.H. You’ve come a long way: c-di-GMP signaling. Curr. Opin. Microbiol. 2012, 15, 140–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertani, G. Studies on lysogenesis I. J. Bacteriol. 1951, 62, 293–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D. Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 1983, 166, 557–580. [Google Scholar] [CrossRef]
- Lugtenberg, B.J.J.; Dekkers, L.C. What makes Pseudomonas bacteria rhizosphere competent? Environ. Microbiol. 1999, 1, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Franklin, F.C.H.; Bagdasarian, M.; Bagdasarian, M.M.; Timmis, K.N. Molecular and functional analysis of the TOL plasmid pWWO from Pseudomonas putida and cloning of genes for the entire regulated aromatic ring meta cleavage pathway. Proc. Natl. Acad. Sci. USA 1981, 78, 7458–7462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, B.R.; Marinus, M.G. The dam and dcm strains of Escherichia coli—A review. Gene 1994, 143, 1–12. [Google Scholar] [CrossRef]
- Macho, A.P.; Ruiz-Albert, J.; Tornero, P.; Beuzón, C.R. Identification of new type III effectors and analysis of the plant response by competitive index. Mol. Plant Pathol. 2009, 10, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Martin, I.; Thwaites, R.; Mansfield, J.W.; Beuzón, C.R. Negative regulation of the Hrp type III secretion system in Pseudomonas syringae pv. phaseolicola. Mol. Plant-Microbe Interact. 2010, 23, 682–701. [Google Scholar] [CrossRef] [Green Version]
- Russell, D.W.; Sambrook, J. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Pérez-Martínez, I.; Rodriguez-Moreno, L.; Matas, I.M.; Ramos, C. Strain selection and improvement of gene transfer for genetic manipulation of Pseudomonas savastanoi isolated from olive knots. Res. Microbiol. 2007, 158, 60–69. [Google Scholar] [CrossRef]
- Li, R.; Li, Y.; Kristiansen, K.; Wang, J. SOAP: Short oligonucleotide alignment program. Bioinformatics 2008, 24, 713–714. [Google Scholar] [CrossRef] [Green Version]
- Chaudhari, N.M.; Gupta, V.K.; Dutta, C. BPGA- an ultra-fast pan-genome analysis pipeline. Nat. Publ. Gr. 2016, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Tettelin, H.; Riley, D.; Cattuto, C.; Medini, D. Comparative genomics: The bacterial pan-genome. Curr. Opin. Microbiol. 2008, 11, 472–477. [Google Scholar] [CrossRef]
- Page, A.J.; Cummins, C.A.; Hunt, M.; Wong, V.K.; Reuter, S.; Holden, M.T.G.; Fookes, M.; Falush, D.; Keane, J.A.; Parkhill, J. Roary: Rapid large-scale prokaryote pan genome analysis. Bioinformatics 2015, 31, 3691–3693. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Petkau, A.; Stuart-Edwards, M.; Stothard, P.; van Domselaar, G. Interactive microbial genome visualization with GView. Bioinformatics 2010, 26, 3125–3126. [Google Scholar] [CrossRef]
- Casimiro-Soriguer, C.S.; Muñoz-Mérida, A.; Pérez-Pulido, A.J. Sma3s: A universal tool for easy functional annotation of proteomes and transcriptomes. Proteomics 2017, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas, P.; Felipe, A.; Michán, C.; Gallegos, M.-T. Induction of Pseudomonas syringae pv. tomato DC3000 MexAB-OprM multidrug efflux pump by flavonoids is mediated by the repressor PmeR. Mol. Plant. Microbe. Interact. 2011, 24, 1207–1219. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- López-Sánchez, A.; Jiménez-Fernández, A.; Calero, P.; Gallego, L.D.; Govantes, F. New methods for the isolation and characterization of biofilm-persistent mutants in Pseudomonas putida. Environ. Microbiol. Rep. 2013, 5, 679–685. [Google Scholar] [CrossRef] [PubMed]
- Belda, E.; van Heck, R.G.A.; José Lopez-Sanchez, M.; Cruveiller, S.; Barbe, V.; Fraser, C.; Klenk, H.-P.; Petersen, J.; Morgat, A.; Nikel, P.I.; et al. The revisited genome of Pseudomonas putida KT2440 enlightens its value as a robust metabolic chassis. Environ. Microbiol. 2016, 18, 3403–3424. [Google Scholar] [CrossRef]
- Mulet, M.; Lalucat, J.; García-Valdés, E. DNA sequence-based analysis of the Pseudomonas species. Environ. Microbiol. 2010, 12, 1513–1530. [Google Scholar] [CrossRef] [Green Version]
- Lalucat, J.; Mulet, M.; Gomila, M.; García-Valdés, E. Genomics in Bacterial Taxonomy: Impact on the Genus Pseudomonas. Genes 2020, 11, 139. [Google Scholar] [CrossRef] [Green Version]
- Hugh, R.; Ikari, P. The proposed neotype strain of Pseudomonas alcaligenes Monias 1928: Request for an Opinion. Int. Bull. Bacteriol. Nomencl. Taxon. 1964, 14, 103–107. [Google Scholar] [CrossRef]
- Chauhan, A.; Green, S.; Pathak, A.; Thomas, J.; Venkatramanan, R. Whole-genome sequences of five oyster-associated bacteria show potential for crude oil hydrocarbon degradation. Genome Announc. 2013, 1, 61–62. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.; Suzuki, S.; Matsui, M.; Hiraki, Y.; Kawano, F.; Shibayama, K. Genome Sequence of a Strain of the Human Pathogenic Bacterium Pseudomonas alcaligenes That Caused Bloodstream Infection. Genome Announc. 2013, 1, e00919-13. [Google Scholar] [CrossRef] [Green Version]
- Galperin, M.Y. Bacterial signal transduction network in a genomic perspective. Environ. Microbiol. 2004, 6, 552–567. [Google Scholar] [CrossRef]
- Galperin, M.Y. Structural classification of bacterial response regulators: Diversity of output domains and domain combinations. J. Bacteriol. 2006, 188, 4169–4182. [Google Scholar] [CrossRef] [Green Version]
- Cardinali-Rezende, J.; Alexandrino, P.M.R.; Nahat, R.A.T.P.D.S.; Sant’Ana, D.P.V.; Silva, L.F.; Gomez, J.G.C.; Taciro, M.K. Draft Genome Sequence of Pseudomonas sp. Strain LFM046, a Producer of Medium-Chain-Length Polyhydroxyalkanoate. Genome Announc. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Tsang, H.L.; Huang, J.L.; Lin, Y.H.; Huang, K.F.; Lu, P.L.; Lin, G.H.; Khine, A.A.; Hu, A.; Chen, H.P. Borneol dehydrogenase from Pseudomonas sp. strain TCU-HL1 catalyzes the oxidation of (+)-borneol and its isomers to camphor. Appl. Environ. Microbiol. 2016, 82, 6378–6385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busquets, A.; Peña, A.; Gomila, M.; Mulet, M.; Mayol, J.; García-Valdés, E.; Bennasar, A.; Huntemann, M.; Han, J.; Chen, I.-M.; et al. Draft Genome Sequence of Pseudomonas azotifigens Strain DSM 17556T (6H33bT), a Nitrogen Fixer Strain Isolated from a Compost Pile. Genome Announc. 2013, 1, e00893-1. [Google Scholar] [CrossRef] [Green Version]
- O’Mahony, M.M.; Dobson, A.D.W.; Barnes, J.D.; Singleton, I. The use of ozone in the remediation of polycyclic aromatic hydrocarbon contaminated soil. Chemosphere 2006, 63, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Garrity, G.M.; Bell, J.A.; Lilburn, T. Pseudomonadales Orla-Jensen 1921, 270AL. In Bergey’s Manual® of Systematic Bacteriology; Springer: Boston, MA, USA, 2005; pp. 323–442. [Google Scholar]
- Miyazaki, K.; Hase, E.; Maruya, T. Complete Genome Sequence of Pseudomonas otitidis Strain MrB4, Isolated from Lake Biwa in Japan. Microbiol. Resour. Announc. 2020, 9, e00148-20. [Google Scholar] [CrossRef] [Green Version]
- Pathak, A.; Stothard, P.; Chauhan, A. Comparative Genomic Analysis of Three Pseudomonas Species Isolated from the Eastern Oyster (Crassostrea virginica) Tissues, Mantle Fluid, and the Overlying Estuarine Water Column. Microorganisms 2021, 9, 490. [Google Scholar] [CrossRef]
- Janßen, H.J.; Steinbüchel, A. Fatty acid synthesis in Escherichia coli and its applications towards the production of fatty acid based biofuels. Biotechnol. Biofuels 2014, 7, 1–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varivarn, K.; Champa, L.A.; Silby, M.W.; Robleto, E.A. Colonization strategies of Pseudomonas fluorescens Pf0-1: Activation of soil-specific genes important for diverse and specific environments. BMC Microbiol. 2013, 13, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Giddens, S.R.; Jackson, R.W.; Moon, C.D.; Jacobs, M.A.; Zhang, X.X.; Gehrig, S.M.; Rainey, P.B. Mutational activation of niche-specific genes provides insight into regulatory networks and bacterial function in a complex environment. Proc. Natl. Acad. Sci. USA 2007, 104, 18247–18252. [Google Scholar] [CrossRef] [Green Version]
- Blanco-Romero, E.; Redondo-Nieto, M.; Martínez-Granero, F.; Garrido-Sanz, D.; Ramos-González, M.I.; Martín, M.; Rivilla, R. Genome-wide analysis of the FleQ direct regulon in Pseudomonas fluorescens F113 and Pseudomonas putida KT2440. Sci. Rep. 2018, 8, 13145. [Google Scholar] [CrossRef]
- Craig, L.; Forest, K.T.; Maier, B. Type IV pili: Dynamics, biophysics and functional consequences. Nat. Rev. Microbiol. 2019, 17, 429–440. [Google Scholar] [CrossRef]
- Hernández, S.B.; Cava, F. Environmental roles of microbial amino acid racemases. Environ. Microbiol. 2016, 18, 1673–1685. [Google Scholar] [CrossRef]
- López-Sánchez, A.; Leal-Morales, A.; Jiménez-Díaz, L.; Platero, A.I.; Bardallo-Pérez, J.; Díaz-Romero, A.; Acemel, R.D.; Illán, J.M.; Jiménez-López, J.; Govantes, F. Biofilm formation-defective mutants in Pseudomonas putida. FEMS Microbiol. Lett. 2016, 363. [Google Scholar] [CrossRef]
- Chao, L.; Rakshe, S.; Leff, M.; Spormanna, A.M. PdeB, a cyclic di-GMP-specific phosphodiesterase that regulates Shewanella oneidensis MR-1 motility and biofilm formation. J. Bacteriol. 2013, 195, 3827–3833. [Google Scholar] [CrossRef] [Green Version]




| Strain/Plasmid | Relevant Characteristics | Reference or Source |
|---|---|---|
| Pseudomonas alcaligenes | ||
| AVO110 | Wild-type (NfR) | [1,22] |
| AVO110 ∆cmpA | cmpA deletion mutant (NfR KmR) | This work |
| Pseudomonas putida | ||
| KT2440 | Wild-type (NfR) | [43] |
| Escherichia coli | ||
| DH5α | F−, ϕ80dlacZΔM15, Δ(lacZYA-argF)U169, deoR, recA1 endA1, hsdR17(rK− mK+), phoA, supE44, λ−, thi-1, gyrA96, relA1 | [41] |
| GM2929 | F−, ara-14, leuB6, thi-1, tonA31, lacY1, tsx-78, galK2, galT22, glnV44, hisG4, rpsL136, xyl-5, mtl-1, dam13::Tn9, dcm-6, mcrB1, hsdR2, mcrA, recF143 (SpR CmR) | [44] |
| Plasmids | ||
| pGEM-T Easy vector | Cloning vector, orif1(-) and lacZ (ApR) | Promega, Madison, WI, USA |
| pBluescript SK II | Cloning vector, orif1(-), oripUC, Plac, lacZ’ (ApR) | Agilent Technologies, USA |
| pAMEX | Expression vector, PnptII (KmR) | [45] |
| pGEM-T-KmFRT-BamHI | Contains nptII gene from pKD4 (ApR KmR) | [46] |
| pGEM-T-cmpA | Contains P. alcaligenes AVO110 cmpA ORF and its ribosomal binding site | This work |
| pBSKII-ΔcmpA-Km | pBluescript SK II derivative used for marker exchange mutagenesis of P. alcaligenes AVO110 cmpA gene (ApR KmR) | This work |
| pAMEX-cmpA | pAMEX derivative carrying wild-type cmpA gene from P. alcaligenes AVO110 (KmR) | This work |
| pAMEX-cmpA-GGAAF | pAMEX-cmpA derivative carrying mutated cmpA-GGAAF allele (KmR) | This work |
| pAMEX-cmpA-AAL | pAMEX-cmpA derivative carrying mutated cmpA-AAL allele (KmR) | This work |
| Species | Subgroup a | Strains | Hard Core Genome b | Soft Core Genome b | Hard Pan-Genome b | Soft Pan- Genome b | Heaps’ Law (Gamma) |
|---|---|---|---|---|---|---|---|
| P. alcaligenes | I | 8 | 974 | 984 | 11,778 | 11,553 | 0.5030 |
| II | 2 | 5503 | 6319 | 6890 | 6319 | NAP | |
| I + II | 10 | 402 | 405 | 17,284 | 17,064 | 0.5817 | |
| P. pseudoalcaligenes | NAP | 2 | 3301 | 4009 | 4872 | 4009 | NAP |
| Transcription | Accession a (A9179_No.) | Product | Fold Change b |
|---|---|---|---|
| Upregulated | RS00870 | Chemotaxis-specific protein-glutamate methyltransferase CheB | 149.92 ± 55.77 |
| RS08440 | Long-chain acyl-CoA synthetase/AMP-binding protein FadD | 389.19 ± 46.96 | |
| RS08445 | Long-chain fatty acid-CoA ligase/iron-containing redox enzyme family protein | 373.48 ± 68.63 | |
| RS08450 | Short-chain dehydrogenase/ SDR family oxidoreductase | 317.69 ± 94.41 | |
| RS08455 | Tetratricopeptide motif repeat protein | 107.12 ± 22.51 | |
| RS08460 | DNA-binding response regulator | 38.27 ± 10.57 | |
| RS09465 | Pilus assembly protein PilA (type IVb pili) | 50.76 ± 11.58 | |
| RS09470 | Prepilin peptidase (type IVb pili) | 29.27 ± 6.44 | |
| RS09515 | Flp pilus assembly complex ATPase component TadA (type IVb pili) | 39.55 ± 4.21 | |
| RS09580 * | cmpA (GGDEF/EAL domain-containing protein) | 32.27 ± 14,61 | |
| RS10675 | DUF411 domain-containing protein | 94.81 ± 28.25 | |
| RS10680 | OprD family outer membrane porin | 935.76 ± 253.68 | |
| RS21175 | Hypothetical protein | 192.12 ± 85.68 | |
| Downregulated | RS00240 | Glutamine synthetase | 0.17 ± 0.02 |
| RS10035 | Potassium-transporting ATPase subunit KdpA | 0.50 ± 0.04 | |
| RS19970 | MBL fold metallohydrolase (hydrolysis of beta-lactam antibiotics) | 0.39 ± 0.01 | |
| RS21875 | Alanine racemase (peptidoglycan biosynthesis) | 0.17 ± 0.02 | |
| RS22500 | Glutamine synthetase | 0.50 ± 0.06 |
| Strain | Query Cover | Identity | Accession Number a | Source | Reference or Sequence Source/Year |
|---|---|---|---|---|---|
| Pseudomonas sp. LFM046 | 100% | 78.34% | WP_044875132 | Sugarcane soil | [68] |
| Pseudomonas sp. F(2018) | 100% | 76.91% | WP_171016323 | Alpine spring water | O. Jousson/2018 |
| Pseudomonas sp. PDM15 | 100% | 76.24% | WP_192398170 | Soil | K. Gowda/2020 |
| Pseudomonas sp. 30_B | 100% | 75.99% | WP_207883400 | Rice rhizosphere | V. Venturi/2021 |
| Pseudomonas sp. TCU-HL1 | 99% | 75.97% | WP_069081885 | Soil | [69] |
| Pseudomonas sp. ML96 b | 99% | 75.32% | WP_043307322 | Lake water | X. Li/2014 |
| Pseudomonas indica PIC105 | 99% | 71.52% | WP_084333623 | Olea europaea rhizosphere | C. Gomez-Lama/2017 |
| Pseudomonas sp. LAM-KW06 b | 99% | 70.94% | WP_172149757 | Soil | D. Kong/2020 |
| Pseudomonas azotifigens DSM 17556 | 99% | 63.30% | WP_028239826 | Hyperthermal compost material | [70] |
| Pseudomonas stutzeri NT0128 b | 99% | 62.51% | WP_052679321 | Wheat rhizosphere | N. Tovi/2015 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pintado, A.; Pérez-Martínez, I.; Aragón, I.M.; Gutiérrez-Barranquero, J.A.; de Vicente, A.; Cazorla, F.M.; Ramos, C. The Rhizobacterium Pseudomonas alcaligenes AVO110 Induces the Expression of Biofilm-Related Genes in Response to Rosellinia necatrix Exudates. Microorganisms 2021, 9, 1388. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071388
Pintado A, Pérez-Martínez I, Aragón IM, Gutiérrez-Barranquero JA, de Vicente A, Cazorla FM, Ramos C. The Rhizobacterium Pseudomonas alcaligenes AVO110 Induces the Expression of Biofilm-Related Genes in Response to Rosellinia necatrix Exudates. Microorganisms. 2021; 9(7):1388. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071388
Chicago/Turabian StylePintado, Adrián, Isabel Pérez-Martínez, Isabel M. Aragón, José Antonio Gutiérrez-Barranquero, Antonio de Vicente, Francisco M. Cazorla, and Cayo Ramos. 2021. "The Rhizobacterium Pseudomonas alcaligenes AVO110 Induces the Expression of Biofilm-Related Genes in Response to Rosellinia necatrix Exudates" Microorganisms 9, no. 7: 1388. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071388