
Analysis and Reconstitution of the Menaquinone Biosynthesis Pathway in Lactiplantibacillus plantarum and Lentilactibacillus buchneri

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids, Primers, and Culture Conditions
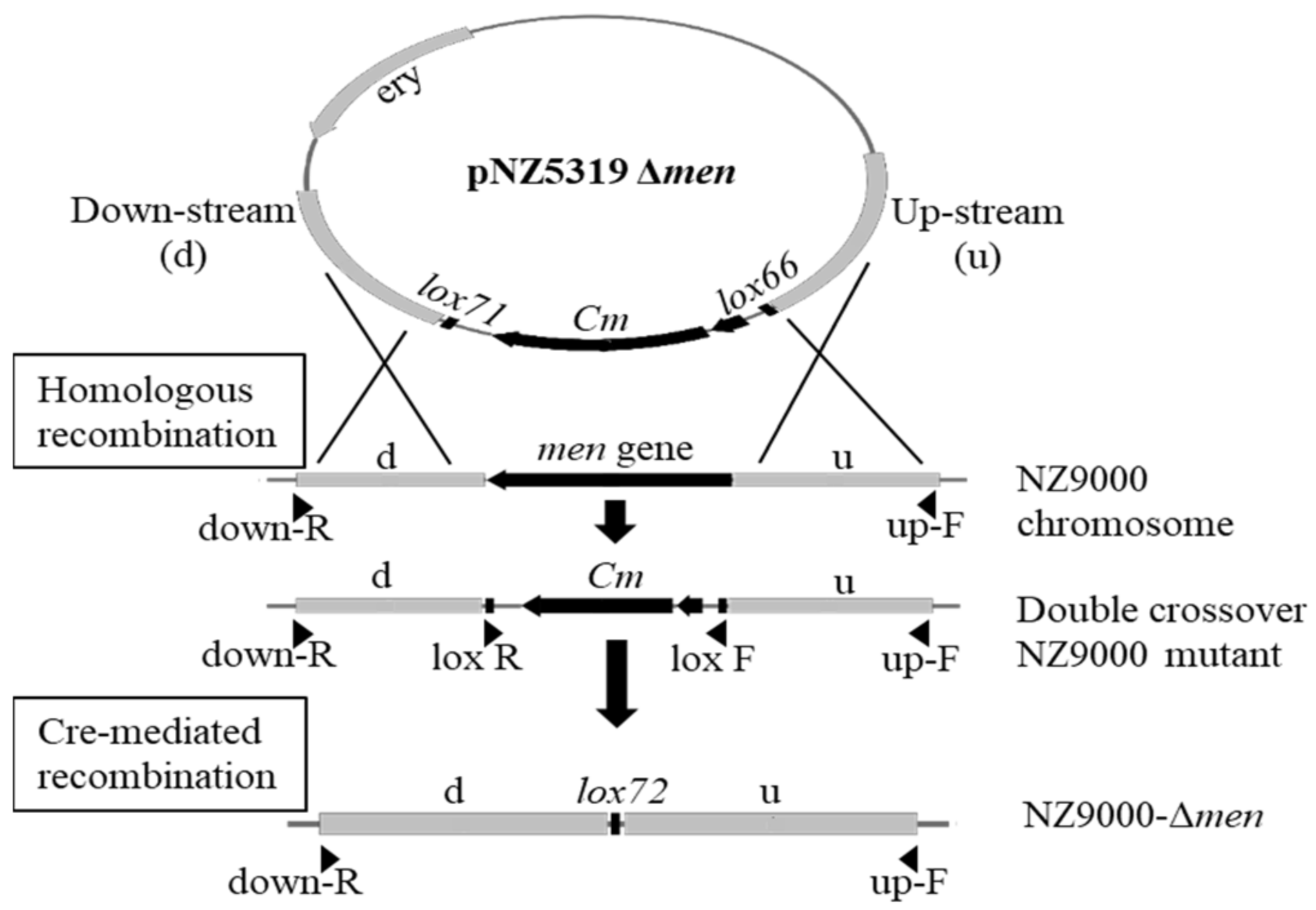
2.2. Knock-Out Vector Constructions
2.3. L. lactis NZ9000 Gene Replacements
2.4. Construction of Expression Vectors for Menaquinone Biosynthesis Genes
2.5. Complementation of Deleted Menaquinone Biosynthesis Genes and Menaquinone Extraction
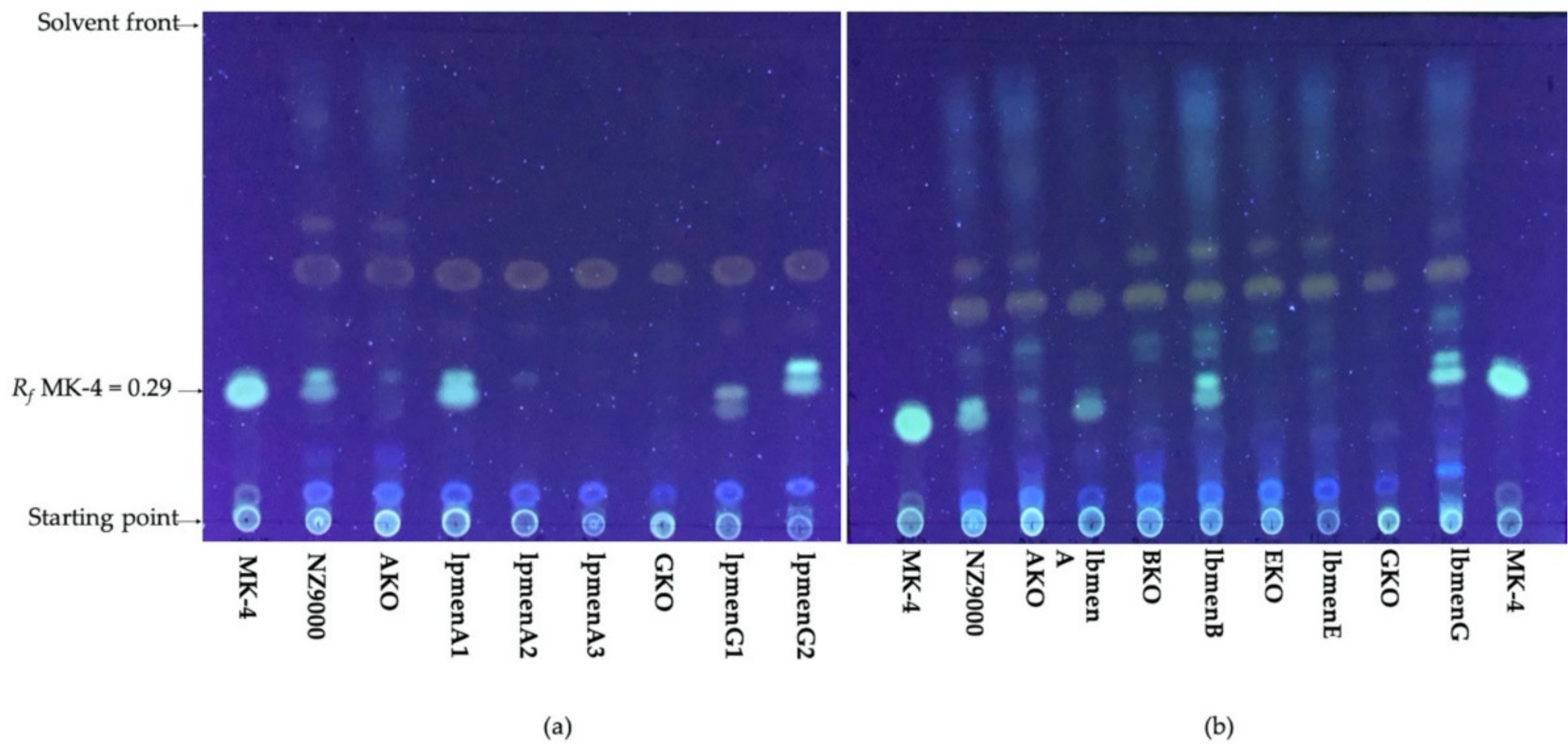
2.6. Thin-Layer Chromatography (TLC)
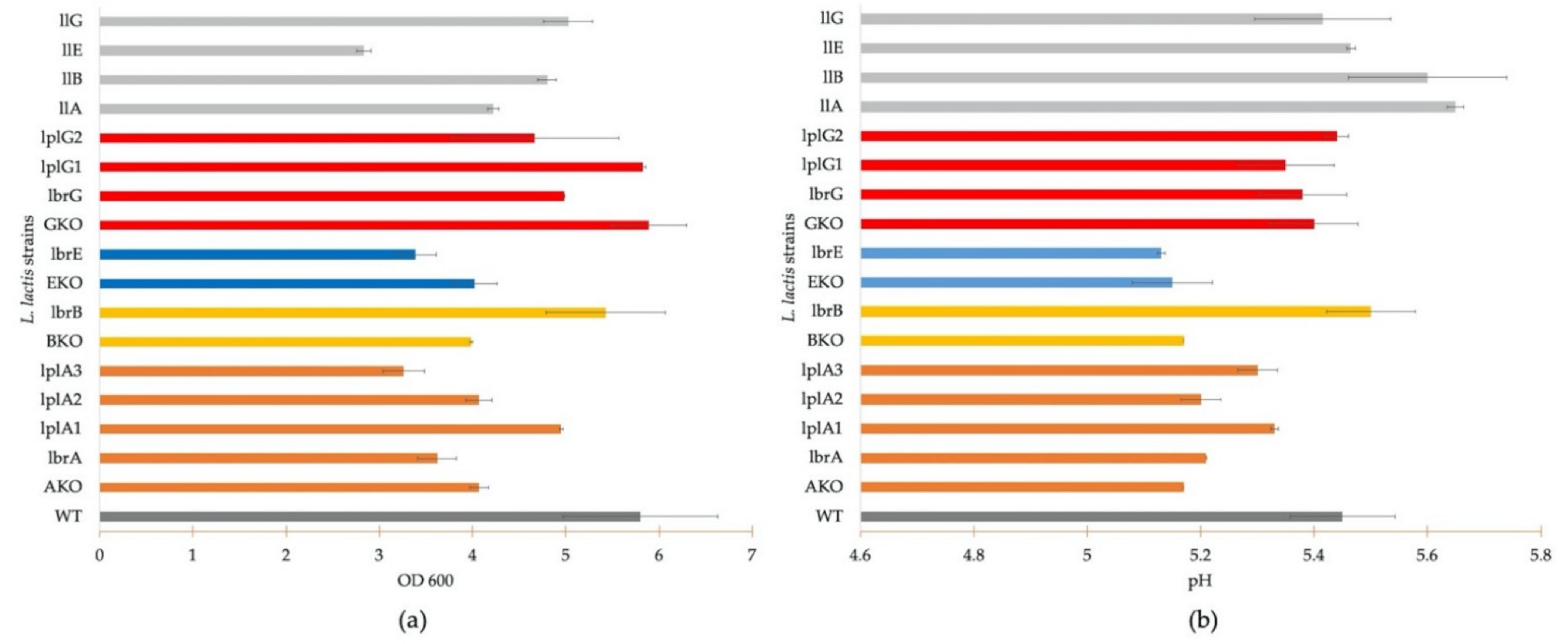
2.7. Growth Profile Analysis
2.8. Construction of Expression Vectors for Menaquinone Biosynthesis
2.9. Analysis of Aerobic Phenotype of Lactipl. plantarum variants
3. Results
3.1. Construction of Menaquinone Knockout Vectors
3.2. Expression of Menaquinone Genes from Lactipl. plantarum WCFS1
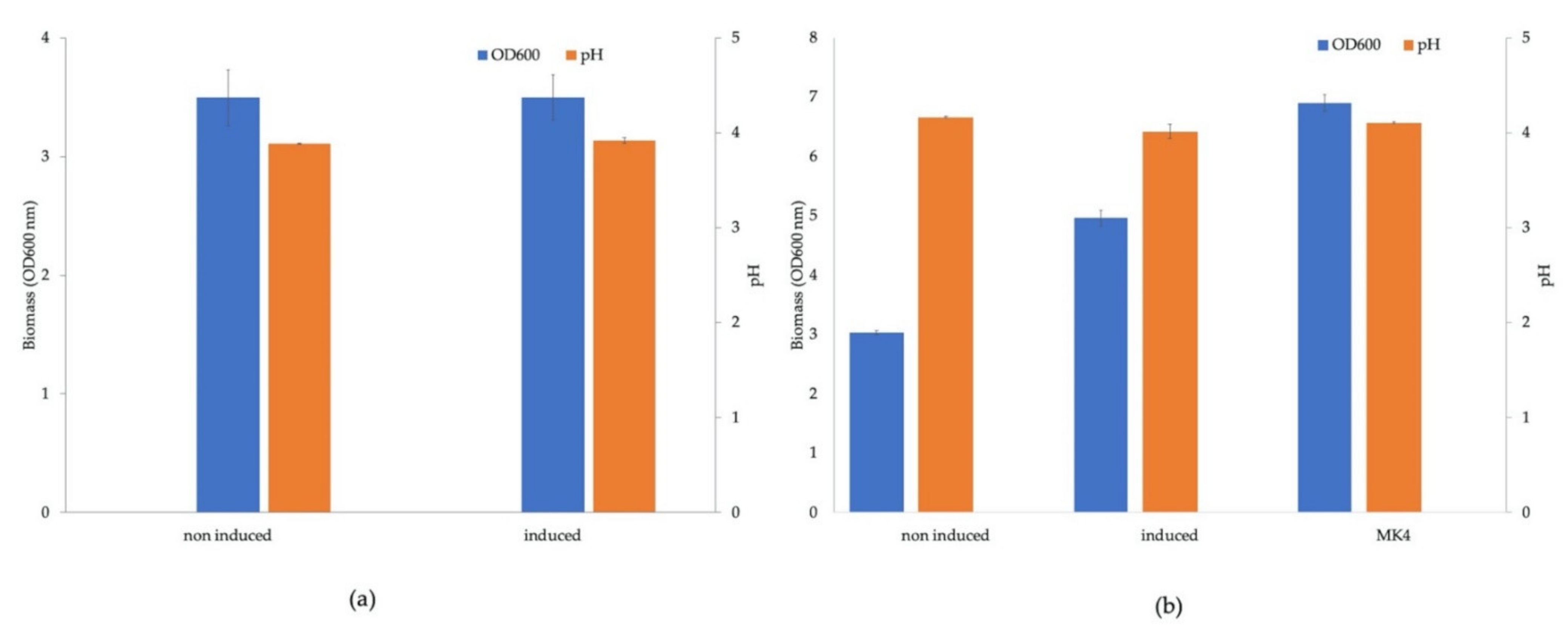
3.3. Growth Profile Analysis
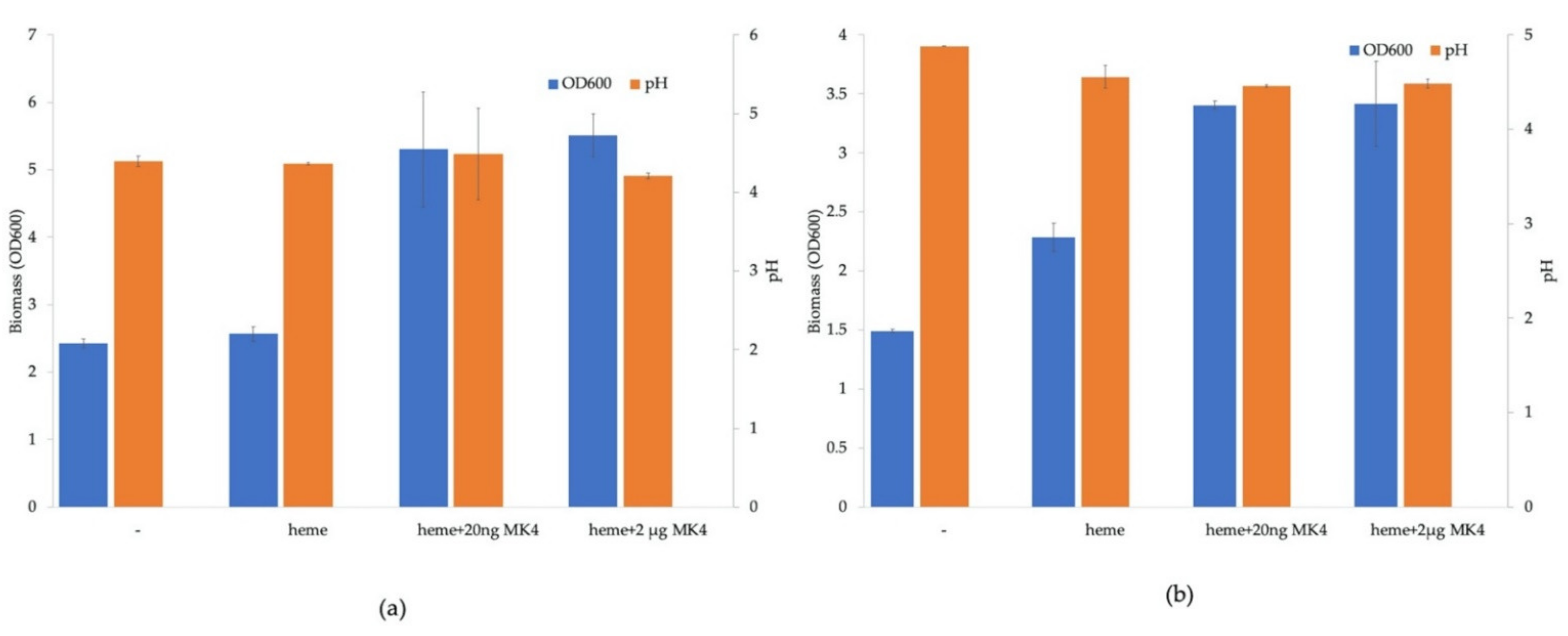
3.4. Respiratory Phenotype of Lent. buchneri DSM 20057 and Lactipl. plantarum WCFS1 and Variant Strains
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Brooijmans, R.; Smit, B.; Santos, F.; van Riel, J.; de Vos, W.M.; Hugenholtz, J. Heme and menaquinone induced electron transport in lactic acid bacteria. Microb. Cell Fact. 2009, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaudu, P.; Vido, K.; Cesselin, B.; Kulakauskas, S.; Tremblay, J.; Rezaïki, L.; Lamberet, G.; Sourice, S.; Duwat, P.; Gruss, A. Respiration capacity and consequences in Lactococcus lactis. Antonie Van Leeuwenhoek 2002, 82, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Lechardeur, D.; Cesselin, B.N.D.; Fernandez, A.; Lamberet, G.; Garrigues, C.; Pedersen, M.; Gaudu, P.; Gruss, A. Using heme as an energy boost for lactic acid bacteria. Curr. Opin. Biotechnol. 2010, 22, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Zotta, T.; Ricciardi, A.; Ianniello, R.G.; Storti, L.V.; Glibota, N.A.; Parente, E. Aerobic and respirative growth of heterofermentative lactic acid bacteria: A screening study. Food Microbiol. 2018, 76, 117–127. [Google Scholar] [CrossRef]
- Reale, A.; Ianniello, R.G.; Ciocia, F.; di Renzo, T.; Boscaino, F.; Ricciardi, A.; Coppola, R.; Parente, E.; Zotta, T.; McSweeney, P.L.H. Effect of respirative and catalase-positive Lactobacillus casei adjuncts on the production and quality of Cheddar-type cheese. Int. Dairy J. 2016, 63, 78–87. [Google Scholar] [CrossRef]
- Reale, A.; di Renzo, T.; Zotta, T.; Preziuso, M.; Boscaino, F.; Ianniello, R.; Storti, L.V.; Tremonte, P.; Coppola, R. Effect of respirative cultures of Lactobacillus casei on model sourdough fermentation. LWT Food Sci. Technol. 2016, 73, 622–629. [Google Scholar] [CrossRef]
- Pedersen, M.B.; Gaudu, P.; Lechardeur, D.; Petit, M.-A.; Gruss, A. Aerobic respiration metabolism in lactic acid bacteria and uses in biotechnology. Annu. Rev. Food Sci. Technol. 2012, 3, 37–58. [Google Scholar] [CrossRef]
- Zotta, T.; Parente, E.; Ricciardi, A. Aerobic metabolism in the genus Lactobacillus: Impact on stress response and potential applications in the food industry. J. Appl. Microbiol. 2017, 122, 857–869. [Google Scholar] [CrossRef] [Green Version]
- Kurosu, M.; Begari, E. Vitamin K2 in electron transport system: Are enzymes involved in vitamin K2 biosynthesis promising drug targets? Molecules 2010, 15, 1531–1553. [Google Scholar] [CrossRef] [Green Version]
- Brooijmans, R. Electron Transport Chains of Lactic Acid Bacteria; Wageningen University: Wageningen, The Netherlands, 2008. [Google Scholar]
- Cesselin, B.; Derré-Bobillot, A.; Fernandez, A.; Lamberet, G.; Lechardeur, D.; Yamamoto, Y.; Pedersen, M.B.; Garrigues, C.; Gaudu, P. Respiration, a strategy to avoid oxidative stress in Lactococcus lactis, is regulated by the heme status. Jpn. J. Lact. Acid Bact. 2010, 21, 10–15. [Google Scholar] [CrossRef]
- Malarme, K.; Malarme, K.; Weissenbach, J.; Weissenbach, J.; Ehrlich, S.D.; Ehrlich, S.D.; Sorokin, A.; Sorokin, A. The complete genome sequence of the lactic acid bacterium Lactococcus lactis ssp. lactis IL1403. Genome Res. 2001, 11, 731–753. [Google Scholar] [CrossRef]
- Wegmann, U.; Zomer, A.; Buist, G.; Shearman, C.; Canchaya, C.; Ventura, M.; Goesmann, A.; Gasson, M.J.; Kuipers, O.P.; van Sinderen, D.; et al. Complete genome sequence of the prototype lactic acid bacterium Lactococcus lactis subsp cremoris MG1363. J. Bacteriol. 2007, 189, 3256–3270. [Google Scholar] [CrossRef] [Green Version]
- Brooijmans, R.; de Vos, W.M.; Hugenholtz, J. Lactobacillus plantarum WCFS1 electron transport chains. Appl. Environ. Microbiol. 2009, 75, 3580–3585. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Fritsch, F.E.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Kuipers, O.P.; de Ruyter, P.G.G.; Kleerebezem, M.; de Vos, W.M. Quorum sensing-controlled gene expression in lactic acid bacteria. J. Biotechnol. 1998, 64, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Lambert, J.M.; Bongers, R.S.; Kleerebezem, M. Cre-lox-based system for multiple gene deletions and selectable-marker removal in Lactobacillus plantarum. Appl. Environ. Microbiol. 2007, 73, 1126–1135. [Google Scholar] [CrossRef] [Green Version]
- Geertsma, E.R.; Poolman, B. High-throughput cloning and expression in Lactococcus lactis. Nat. Methods 2007, 4, 705–707. [Google Scholar] [CrossRef] [Green Version]
- Holo, H.; Nes, I.F. High-frequency transformation, by electroporation, of Lactococcus lactis subsp cremoris grown with glycine in osmotically stabilized media. Appl. Environ. Microbiol. 1989, 55, 3119–3123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicoloff, H.; Hubert, J.C.; Bringel, F. In Lactobacillus plantarum, carbamoyl phosphate is synthesized by two carbamoyl-phosphate synthetases (CPS): Carbon dioxide differentiates the arginine-repressed from the pyrimidine-regulated CPS. J. Bacteriol. 2000, 182, 3416–3422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, H.; Zhao, G.; Liu, H.; Wang, H.; Ni, W.; Wang, P.; Zheng, Z. A simple and efficient method for the extraction and separation of menaquinone homologs from wet biomass of Flavobacterium. Bioprocess Biosyst. Eng. 2018, 41, 107–113. [Google Scholar] [CrossRef]
- Myers, J.M.; Myers, C.R. Role of the tetraheme cytochrome CymA in anaerobic electron transport in cells of Shewanella putrefaciens MR-1 with normal levels of menaquinone. J. Bacteriol. 2000, 182, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Morishita, T.; Tamura, N.; Makino, T.; Kudo, S. Production of menaquinones by lactic acid bacteria. J. Dairy Sci. 1999, 82, 1897–1903. [Google Scholar] [CrossRef]
- Zheng, L.; Baumann, U.; Reymond, J.-L. An efficient one-step site-directed and site-saturation mutagenesis protocol. Nucleic Acids Res. 2004, 32, e115. [Google Scholar] [CrossRef] [Green Version]
- Sørvig, E.; Mathiesen, G.; Naterstad, K.; Eijsink, V.G.H.; Axelsson, L. High-level, inducible gene expression in Lactobacillus sakei and Lactobacillus plantarum using versatile expression vectors. Microbiology 2005, 151, 2439–2449. [Google Scholar] [CrossRef] [Green Version]
- Dairi, T. An alternative menaquinone biosynthetic pathway operating in microorganisms: An attractive target for drug discovery to pathogenic Helicobacter and Chlamydia strains. J. Antibiot. 2009, 62, 347–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakeman, C.A.; Hammer, N.D.; Stauff, D.L.; Attia, A.S.; Anzaldi, L.L.; Dikalov, S.I.; Calcutt, M.W.; Skaar, E.P. Menaquinone biosynthesis potentiates haem toxicity in Staphylococcus aureus. Mol. Microbiol. 2012, 86, 1376–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezaïki, L.; Lamberet, G.; Derré, A.; Gruss, A.; Gaudu, P. Lactococcus lactis produces short-chain quinones that cross-feed Group B Streptococcus to activate respiration growth. Mol. Microbiol. 2008, 67, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Matschiner, J.T.; Amelotti, J.M. Characterization of vitamin K from bovine liver. J. Lipid Res. 1968, 9, 176–179. [Google Scholar] [CrossRef]
- Dunphy, P.J.; Phillips, P.G.; Brodie, A.F. Separation and identification of menaquinones iron microorganisms. J. Lipid Res. 1971, 12, 442–449. [Google Scholar] [CrossRef]
- Collin, M.D.; Shah, H.; Minnikin, D.E. A note on the separation of natural mixtures of bacterial menaquinones using reverse phase thin-layer chromatography. J. Appl. Bacteriol. 1980, 48, 277–282. [Google Scholar] [CrossRef]
- Suvarna, K.; Stevenson, D.; Meganathan, R.; Hudspeth, M.E.S. Menaquinone (vitamin k 2) biosynthesis: Localization and characterization of the mena gene from Escherichia coli. J. Bacteriol. 1998, 180, 2782–2787. [Google Scholar] [CrossRef] [Green Version]
- Ramsey, M.; Hartke, A.; Huycke, M. The physiology and metabolism of enterococci. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection; Gilmore, M.S., Clewell, D.B., Ike, Y., Shankar, N., Eds.; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014; pp. 581–635. [Google Scholar]
- Huycke, M.M.; Moore, D.; Joyce, W.; Wise, P.; Shepard, L.; Kotake, Y.; Gilmore, M.S. Extracellular superoxide production by Enterococcus faecalis requires demethylmenaquinone and is attenuated by functional terminal quinol oxidases. Mol. Microbiol. 2001, 42, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Winstedt, L.; Frankenberg, L.; Hederstedt, L. Enterococcus faecalis V583 contains a cytochrome bd-type respiratory oxidase. J. Bacteriol. 2000, 182, 3863–3866. [Google Scholar] [CrossRef] [Green Version]
- Baureder, M.; Hederstedt, L. Genes important for catalase activity in Enterococcus faecalis. PLoS ONE 2012, 7, e36725. [Google Scholar] [CrossRef] [Green Version]
- Garrigues, C.; Loubiere, P.; Lindley, N.D.; Durand, G. Control of the shift from homolactic acid to mixed-acid fermentation in Lactococcus lactis: Predominant role of the NADH/NAD ratio. J. Bacteriol. 1997, 179, 5282–5287. [Google Scholar] [CrossRef] [Green Version]
- Koebmann, B.; Blank, L.M.; Solem, C.; Petranovic, D.; Nielsen, L.K.; Jensen, P.R. Increased biomass yield of Lactococcus lactis during energetically limited growth and respiratory conditions. Biotechnol. Appl. Biochem. 2008, 50, 25–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, Y.; Gaudu, P.; Gruss, A. Oxidative stress and oxygen metabolism in lactic acid bacteria. In Lactic Acid Bacteria and Bifidobacteria Current Progress in Advanced Research; Sonomoto, A.Y.K., Ed.; Caister Academic Press: Norfolk, UK, 2011; pp. 91–102. [Google Scholar]
- Mazzeo, M.F.; Cacace, G.; Peluso, A.; Zotta, T.; Muscariello, L.; Vastano, V.; Parente, E.; Siciliano, R.A. Effect of inactivation of ccpA and aerobic growth in Lactobacillus plantarum: A proteomic perspective. J. Proteomics 2012, 75, 4050–4061. [Google Scholar] [CrossRef]
- Eikmeyer, F.G.; Heinl, S.; Marx, H.; Pühler, A.; Grabherr, R.; Schlüter, A. Identification of oxygen-responsive transcripts in the silage inoculant Lactobacillus buchneri CD034 by RNA sequencing. PLoS ONE 2015, 10, e0134149. [Google Scholar] [CrossRef]
- Ricciardi, A.; Ianniello, R.G.; Parente, E.; Zotta, T. Factors affecting gene expression and activity of heme- and manganese-dependent catalases in Lactobacillus casei strains. Int. J. Food Microbiol. 2018, 280, 66–77. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Y. Engineering the antioxidative properties of lactic acid bacteria for improving its robustness. Curr. Opin. Biotechnol. 2013, 24, 142–147. [Google Scholar] [CrossRef] [PubMed]
- An, H.; Zhou, H.; Huang, Y.; Wang, G.; Luan, C.; Mou, J.; Luo, Y.; Hao, Y. High-level expression of heme-dependent catalase gene katA from Lactobacillus sakei protects Lactobacillus rhamnosus from oxidative stress. Mol. Biotechnol. 2010, 45, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Zotta, T.; Ricciardi, A.; Guidone, A.; Sacco, M.; Muscariello, L.; Mazzeo, M.F.; Cacace, G.; Parente, E. Inactivation of ccpA and aeration affect growth, metabolite production and stress tolerance in Lactobacillus plantarum WCFS1. Int. J. Food Microbiol. 2012, 155, 51–59. [Google Scholar] [CrossRef]
- Igarashi, T.; Kono, Y.; Tanaka, K. Molecular cloning of manganese catalase from Lactobacillus plantarum. J. Biol. Chem. 1996, 271, 29521–29524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, M.; van der Veen, S.; Nakajima, H.; Abee, T. Effect of respiration and manganese on oxidative stress resistance of Lactobacillus plantarum WCFS1. Microbiology 2012, 158, 293–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berlec, A.; Škrlec, K.; Kocjan, J.; Olenic, M.; Štrukelj, B. Single plasmid systems for inducible dual protein expression and for CRISPR-Cas9/CRISPRi gene regulation in lactic acid bacterium Lactococcus lactis. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Tan, S. A modular polycistronic expression system for overexpressing protein complexes in Escherichia coli. Protein Expr. Purif. 2001, 21, 224–234. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.-J.; Kim, H.-E.; Lee, K.-H.; Han, W.; Yi, M.-J.; Jeong, J.; Oh, B.-H. Two-promoter vector is highly efficient for overproduction of protein complexes. Protein Sci. 2004, 13, 1698–1703. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Relevant Features | References |
|---|---|---|
| Escherichia coli | ||
| NEB5α | Cloning host | NEB |
| JM101 | Cloning host recA+ | [15] |
| Lactococcus lactis (ll) | ||
| NZ9000 | MG1363 pepN::nisRK, cloning host | [16] |
| ∆menA | NZ9000 menA deficient by lox72 replacement | This study |
| ∆menB | NZ9000 menB deficient by lox72 replacement | This study |
| ∆menE | NZ9000 menE deficient by lox72 replacement | This study |
| ∆menG | NZ9000 menG deficient by lox72 replacement | This study |
| Plasmids | Relevant Features | References |
|---|---|---|
| pSIP409gusA | spp-based expression vector, PSIP401 derivative, gusA controlled by PsppQ(PorfX), sppKR expression driven by eryB read through | [25] |
| pSIP409∆BsaI | ery, PSIP401 derivative with removal of BsaI restriction site | This study |
| pSIP_lpMK_2 | ery, PSIP401∆BsaI derivative, menaquinone biosynthesis genes for Lactipl. plantarum controlled by two PsppQ | This study |
| pSIP_lpMK_1 | ery, PSIP401∆BsaI derivative, menaquinone biosynthesis genes for Lactipl. plantarum controlled by one PsppQ | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watthanasakphuban, N.; Virginia, L.J.; Haltrich, D.; Peterbauer, C. Analysis and Reconstitution of the Menaquinone Biosynthesis Pathway in Lactiplantibacillus plantarum and Lentilactibacillus buchneri. Microorganisms 2021, 9, 1476. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071476
Watthanasakphuban N, Virginia LJ, Haltrich D, Peterbauer C. Analysis and Reconstitution of the Menaquinone Biosynthesis Pathway in Lactiplantibacillus plantarum and Lentilactibacillus buchneri. Microorganisms. 2021; 9(7):1476. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071476
Chicago/Turabian StyleWatthanasakphuban, Nisit, Ludovika Jessica Virginia, Dietmar Haltrich, and Clemens Peterbauer. 2021. "Analysis and Reconstitution of the Menaquinone Biosynthesis Pathway in Lactiplantibacillus plantarum and Lentilactibacillus buchneri" Microorganisms 9, no. 7: 1476. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071476