
Bacteriophages of Thermophilic ‘Bacillus Group’ Bacteria—A Review
by
 ,
,
Beata Łubkowska
1,2,*, 
Joanna Jeżewska-Frąckowiak
1,
Ireneusz Sobolewski
1 and
Piotr M. Skowron
1 1
Department of Molecular Biotechnology, Faculty of Chemistry, University of Gdansk, Wita Stwosza 63, 80-308 Gdansk, Poland
2
The High School of Health in Gdansk, Pelplinska 7, 80-335 Gdansk, Poland
*
Author to whom correspondence should be addressed.
Microorganisms 2021, 9(7), 1522; https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071522
Submission received: 3 June 2021
/
Revised: 3 July 2021
/
Accepted: 12 July 2021
/
Published: 16 July 2021
(This article belongs to the Special Issue Bacillus subtilis as a Model Organism to Study Basic Cell Processes)
Abstract
:Bacteriophages of thermophiles are of increasing interest owing to their important roles in many biogeochemical, ecological processes and in biotechnology applications, including emerging bionanotechnology. However, due to lack of in-depth investigation, they are underrepresented in the known prokaryotic virosphere. Therefore, there is a considerable potential for the discovery of novel bacteriophage-host systems in various environments: marine and terrestrial hot springs, compost piles, soil, industrial hot waters, among others. This review aims at providing a reference compendium of thermophages characterized thus far, which infect the species of thermophilic ‘Bacillus group’ bacteria, mostly from Geobacillus sp. We have listed 56 thermophages, out of which the majority belong to the Siphoviridae family, others belong to the Myoviridae and Podoviridae families and, apparently, a few belong to the Sphaerolipoviridae, Tectiviridae or Corticoviridae families. All of their genomes are composed of dsDNA, either linear, circular or circularly permuted. Fourteen genomes have been sequenced; their sizes vary greatly from 35,055 bp to an exceptionally large genome of 160,590 bp. We have also included our unpublished data on TP-84, which infects Geobacillus stearothermophilus (G. stearothermophilus). Since the TP-84 genome sequence shows essentially no similarity to any previously characterized bacteriophage, we have defined TP-84 as a new species in the newly proposed genus Tp84virus within the Siphoviridae family. The information summary presented here may be helpful in comparative deciphering of the molecular basis of the thermophages’ biology, biotechnology and in analyzing the environmental aspects of the thermophages’ effect on the thermophile community.
1. Introduction
Bacterial viruses (bacteriophages or phages) are the most abundant, genetically and evolutionary diversified biological entities in the biosphere [1]. They play key roles in the regulation and evolution of bacterial communities in essentially all ecosystems. They are found in all environments, including extreme ones, such as those associated with: very low or high temperatures, hot/cold deserts, hypersaline systems, low or high pH, such as soda lakes, high hydrodynamic pressures or containing poisonous elements or compounds, among others.
Here we focus on a narrow segment of such bacteriophages, meeting two criteria: (i) those living in a moderately high temperature segment of approximately 45–70 °C. Ecosystems providing such a condition are diverse and include: terrestrial hot springs, deep-sea hydrothermal vents areas, shallow ocean waters above geothermally active beds, human generated hot waters, hot deserts, compost piles, greenhouse soils, silage, rotting straw, river sludge, stable manure, digested sewage sludge, among others. These contain thermophilic bacterial communities and specific viruses that infect them (thermophilic bacteriophages or thermophages) [2,3] and (ii) thermophages infecting a diverse ‘Bacillus group’ bacteria. A number of moderately thermophilic, spore-forming, aerobic bacteria with a growth range of 45–70 °C were classified into the genera Aneurinibacillus, Alicyclobacillus, Sulfobacillus, Brevibacillus, Thermoactinomyces and Thermobacillus [4,5,6,7]. However, genomes and proteomes have revealed that the majority of these thermophiles belong to the Bacillus sp. (B.) genetic groups 1 and 5 [8,9]. These findings further led to a more in-depth evaluation of thermophiles from group 5. In its course, it was determined, on the basis of high 16S rRNA sequence similarity (98.5–99.2%), that they form a phenotypically and phylogenetically coherent group of thermophilic bacilli. Thus, in 2001, the thermophiles belonging to Bacillus genetic group 5 were reclassified to be included into Geobacillus (G.) gen. nov. with the well-known B. stearothermophilus being assigned as the type strain G. stearothermophilus [9,10]. The Geobacillus name means earth or soil Bacillus and includes moderately thermophilic, aerobic or facultatively anaerobic, neutrophilic, motile, spore-forming rods [10]. The genus may also include some non-spore-forming bacteria, in which sporulation-related genes were inactivated by genome-integrated probacteriophage-like DNA segments [11]. The increasing number of discoveries of many novel thermophages, including those infecting the ‘Bacillus group’, are providing a more complete understanding of thermophiles’ biology, the mechanisms of biochemical adaptations needed for the life in high temperatures and the thermophilic host-bacteriophage evolution. From this perspective, especially interesting is the characterization of microbial and viral thermophilic communities in deep-sea hydrothermal vents fields, as this leads to deciphering the origin of ecosystems on the sea floor, the subsurface biosphere, the driving forces of evolution and the origin of life on the Earth [12,13]. Furthermore, it has been found that a horizontal gene transfer between thermophilic and mesophilic Bacillus species and bacteriophages takes place. This results in genetic mosaicism, as exemplified by the D6E thermophage, possibly leading to the contribution of substantial amounts of foreign DNAs by mobile DNA elements such as bacteriophages to entire microbial communities, including remote ones, such as the deep-sea vent community [14]. From a practical perspective, thermophilic Bacillus species are dominant bacterial workhorses in industrial fermentation processes, thus, these bacteria and their bacteriophages provide a novel source of genetic material and enzymes with great potential for application in biotechnology industry and science. Thus we provide an overview of the bacteriophages that infect thermophilic Bacillus sp. (many reclassified later as Geobacillus sp.), Geobacillus sp. and other related hosts that have been isolated from hot ecosystems around the world.
2. Environment Relation
Thermophilic bacilli, including Geobacillus, grow in essentially all environments, except those with ultra-harsh conditions, such as temperatures near or above boiling and extreme pH, among others. As mentioned above, geobacilli can be found in temperatures ranging from those typical for mesophiles (20–45 °C) up to those characteristic for moderate thermophiles (approximately 45–70 °C) in substantial numbers in ecological niches: geothermal areas, terrestrial hot springs, deep-sea hydrothermal vents areas, shallow ocean waters above geothermally active beds, human generated hot waters, hot deserts, compost piles, greenhouse soils, silage, rotting straw, river sludge, stable manure, digested sewage sludge, regardless of geographic location [15,16]. Especially rich sources of thermophilic Geobacillus bacteria, infected with bacteriophages, are present in humid organic matter, such as active compost piles and greenhouse soils due to active microbial metabolism and increased temperature, not reaching extreme values [2,17]. The G. kaustophilus-infecting thermophage GBK2 was found in a backyard compost pile after enrichment in liquid culture with G. kaustophilus (ATCC 8005) at 55 °C [18]. The minimum detectable growth temperature for Geobacillus isolates was described to be 45 °C [4], although in our hands, laboratory conditions allowed for a slow growth of. G. stearothermophilus strain 10 also in the mesophilic range. As typical soils rarely reach temperatures above 45 °C, this seems to be an evolutionary adaptation, allowing Geobacillus to grow extremely slowly, until more environmentally favorable growth temperatures occur. On the other hand, the temperatures in active organic matter easily exceed 45 °C. Very diverse conditions are present in hot springs: their temperatures range from 40 °C to 98 °C, and the bacteria capable of growing there, including geobacilli, are classified either as moderate thermophiles (40–71 °C) or hyperthermophiles (72–98 °C) [19]. Furthermore, the hot springs’ pH values vary widely, depending on hot water dissolving components of various minerals—from pH 1 to 9—and are defined in three functional categories [20]: acidic (pH 1–5), approximately neutral (pH 6–7.5) and alkaline (pH ˃ 7.5). This translates to a high diversity of ‘Bacillus group’ hosts and their specific bacteriophages, which are able to infect most of the major taxonomic groups of Bacillus thermophiles [21], regardless of the type of ecosystem. For example, two thermophages—Bacillus virus BVW1 and Geobacillus virus GVE1—were isolated from deep-sea hydrothermal fields in the east Pacific [22]. Thermostability assays showed that GVE1, whose host is Geobacillus sp. E26323, isolated from the same location, was most stable at 60 °C [23]. During the cultivation of Geobacillus sp. strain E263, many uncharacterized bacteriophage plaques were observed, indicating the presence of a rich host-bacteriophage community. Another deep-sea hydrothermal fields thermophage GVE2 was found in geobacilli cultured at 65 °C [14,24]. Additionally, a Geobacillus thermophage D6E was characterized in deep-sea hydrothermal vent communities, where viruses play very important roles [14]. Yellowstone National Park is one of the best studied sites regarding the thermophages that infect the species of Geobacillus Thermus as well as Archea, among others. This is due to the fact that Yellowstone has one of the highest concentrations of hot springs in the world, which vary greatly in terms of temperature, pH and minerals content [10]. Bacteriophage GBSV1, infecting Geobacillus sp. 6k51 was isolated from a shallow (100 m depth) off-shore hot spring [25]. Similarly, the bacteriophage ϕOH2, which infects G. kaustophilus NBRC 102445(T), was isolated from the sediment of a hot spring [26] and bacteriophage AP45 and its host strain Aeribacillus sp. CEMTC656 were isolated from the Valley of Geysers, Kamchatka, Russia [27].
Regardless of the environmental diversity, the morphologies and compositions of the ‘Bacillus group’ thermophilic bacteriophages are less diverse than that of the overall community of thermophages. In general, the thermophilic bacteriophages that have been found in all taxonomic groups of Bacteria and Archaea, exhibit diverse morphologies (e.g., tailed capsids, tail-less capsids, filamentous, lipid-containing), unique structures (e.g., very long-tailed siphoviruses), and extend through five taxonomic families: Myoviridae, Siphoviridae, Inoviridae, Tectiviridae and Sphaerolipoviridae [3]. Interestingly, in the diverse ‘Bacillus group’ bacteria, the bacteriophages found thus far belong to a limited number of families: vastly dominating Siphoviridae (head–tail bacteriophages with non-contractile tails), Myoviridae (head–tail bacteriophages with contractile tails), Podoviridae (very short tail) and, possibly scarce, Sphaerolipoviridae, Tectiviridae or Corticoviridae members (icosahedral, with a lipid membrane) (Figure 1). All of them contain dsDNA genome, either linear or circular, or circularly permitted. On the whole, the ubiquity of thermophages and their impact on various trophic levels seems to have a powerful direct influence on diverse microbial communities that play a crucial role in biogeochemical cycles. All this, in turn, plays a role in altering those cycles [27,28,29]. As a result, thermophages can affect/stimulate the modifications of those cycles.
3. ‘Bacillus Group’ Bacteriophages—Chronological Review
The thermophilic ‘Bacillus group’ bacteriophages, reviewed here, comprise 56 species, described in varying detail in original publications (Table 1). Although there have certainly been more such bacteriophages detected, they have not been sufficiently analyzed or have been just briefly mentioned in publications. They infect various Geobacillus (previously Bacillus) sp., ‘B. stearothermophilus’, B. acidocaldarius, B. caldotenax, B. caldolyticus, G. kaustophilus, G. thermoglucosidasius, G. icigianus, B. pseudoalcaliphilus, B. bogoriensis, B. pseudofirmus, B. thermocatenulatus, B. cohnii, undefined Bacillus sp. and Aeribacillus sp. As older species determinations are not always precise, we tend to retain the original author’s nomenclature to avoid confusion and for the reader’s convenience, when reading original publications. Some of these thermophages were discovered and analyzed as long ago as the 1950s, 1960s and 1970s, using the techniques available at the time. The data thus obtained is often incomplete or contains errors (such as the nearly 2-fold underestimation of TP-84’s genome size) especially at the molecular level. Apparently, the earliest thorough/reliable report is that of the thermophage TP-84, discovered in 1952 and preliminarily characterized in the 1960s and 1970s [30]. Much later, its genome was sequenced, analyzed and its proteomics data was established by our group in 2018 [2] (Table 1). Below, we chronologically review the major features of each thermophage and its interactions with its corresponding host. Interestingly, all 56 bacteriophages contain dsDNA genomes. The hosts and bacteriophages were cultivated typically on rich, ‘nutrient broth’-type media at approximately pH 7, unless otherwise noted.
3.1. ‘Thermophilic Lytic Principle’ (Family/Genus Not Determined (ND), Host Putative G. stearothermophilus)
This is apparently the earliest discovered and reported thermophilic bacteriophage, with no assigned name, just mentioned as thermophilic lytic principle [31]. Its host, T60 bacteria was described as medium length, Gram-positive rods, isolated from milk, forming spores and growing between 20 °C and 60 °C, with the optimum at 45–52 °C, which suggests that the host belongs to G. stearothermophilus. The thermophilic lytic principle was capable of inoculating between series of cultures and was optimally lysing the host’s cells at 52–55 °C; the lysis capability declining sharply at temperatures of 60 °C and with no lysis occurring at 62 °C [31].
3.2. ‘Thermophilic Bacteriophage’ (1) (Family/Genus ND, Host Thermophilic Bacillus sp.)
3.3. ’Thermophilic Bacteriophage’ (2) (Family/Genus ND, Host ‘Thermophilic Bacterium No. 10′)
Scarce data was published concerning thermophilic bacteriophage [33]. It was found to infect a thermophilic bacterium no. 10, aerobic, Gram-positive, isolated from thermal springs in Yellowstone National Park, as being or closely resembling B. stearothermophilus [34]. The bacteria exhibited temperature growth range of 35–75 °C and the bacteriophage was propagated with stimulating adsorption Ca2+ ions at 50–70 °C, with optimum at 65 °C, and a latent period of 80–90 min. The bacteriophage produced a halo around clear plaques, suggesting the existence of a diffusing envelope depolymerase. It exhibited substantial thermostability. At 2 h incubation at 95 °C in its growth media, significant phage titer remained, whereas at 100 °C after 30 min, some infective particles were still detectable. Transmission electron microscopy (TEM) images showing the bacteriophage morphology suggest that it belongs to the Siphoviridae family [33,35].
3.4. ‘Thermophilic Bacteriophage’ (3) (Family/Genus ND, Host Thermophilic Bacillus sp.)
One of the first isolated thermophilic bacteriophages included an undefined species of very high thermostability. As a host served gram-positive bacillus, spore-forming, strictly aerobic, capable of growth at over 70 °C, isolated from a compost pile in Tokyo, Japan [36]. The bacteriophage was capable of lysing a host’s cells at between 55 °C and 70 °C, optimally at 65 °C, with a burst size of 205–220. Remarkably, that bacteriophage exhibited a very high thermostability. Suspended in its cultivation media, it was capable of surviving over 120 min at 100 °C with approximately a 1000-fold decrease in a titer. The bacteriophage TEM analysis has shown spherical particles of approximately 20 nm diameter with a tendency to aggregate. Aside from that, the results of the coding nucleic acid determination are questionable. The author claims, as based on chemical analysis, that the bacteriophage is composed of 74% nucleic acids, both DNA—42%, and RNA—32% [37,38].
3.5. TP-84 (Siphoviridae Family, Tp84virus Genus, Host B. stearothermophilus)
Early studies determined a number of microbiological and physical properties of bacteriophage TP-84. This was discovered in 1952 in greenhouse soil, infecting B. stearothermophilus strain 2184 (according to current classification—G. stearothermophilus) [30]. TP-84 exhibits high host specificity. Out of +24 related strains of B. stearothermophilus, two unclassified thermophilic Bacillus strains, T-27 and 194, also supported TP-84 infection as well as G. stearothermophilus strain 10 and 4 supporting TP-84 growth. The most effective host was G. stearothermophilus strain 10, thus it was further used for more detailed studies [39]. Here, we evaluated more G. stearothermophilus hosts that have been used by others for recombinant DNA studies in these bacteria, including: NUB3625 9A5 and NUB R [40]. These hosts supported TP-84 growth but much less effectively than G. stearothermophilus 10. The cultivation conditions, optimal for TP-84, included a rich media of pH 6.5, supplemented with fructose or glucose as a sporulation suppressor and essential Ca2+ and Mg2+ ions. TP-84 can reach a high titer of app. 1012. However, omitting the sugar addition decreased the titer approximately 1000-fold [39], while omitting divalent cations had a dramatic effect on final TP-84 yields, causing an approximate 200-fold decrease in titer [41]. The temperature growth range of 43–76 °C (optimal 55–60 °C) matched that of the hosts growth range. The latent period was 22–24 min. Clear plaques were surrounded by a turbid ‘halo’ ([41] Lubkowska et al., unpublished data), indicating diffusion of an enzyme, possibly glycosylase-depolymerase, degrading the bacterial envelopes. The bacteriophage has a hexagonal head, 53 nm in diameter with a 131 nm long 3–5 nm wide tail. It has a Svedberg coefficient of 436 and bands in CsCl at a density of 1.508 g/cm3 (pH 8.5). The sedimentation and diffusion data allowed for calculation of the TP-84 particle molecular weight of approximately 50 MDa [39]. In our previous work, we characterized and sequenced the genome, and established the proteomics data on TP-84 [2]. TP-84 contains 47.7 kb dsDNA, 42% G+C genome. The bioinformatics analysis revealed the presence of 81 coding sequences (CDSs), coding for polypeptides of 4 kDa or larger. Interestingly, all CDSs are oriented in the same direction, pointing to a dominant transcription direction of one DNA strand only. Based on homology analysis, a hypothetical function could be assigned to 31 CDSs and a total of 33 biosyntheses of proteins was confirmed experimentally [2]. To further confirm the genome bioinformatics data, we analyzed: (i) purified TP-84 bacteriophage particles, subjected to SDS-PAGE, resolving individual proteins and (ii) purified TP-84 proteins from TP-84/G. stearothermophilus lysates, followed by LC-MS analysis (to be published elsewhere). Nineteen new TP-84-encoded proteins were found (Table 1) with at least two peptides sequence coverage. Those include: (i) proteins involved in DNA replication/packaging—TP84_01 (putative terminase, small subunit), TP84_02 (putative terminase, large subunit), TP84_59, replicative DNA helicase and TP84_58, replicative helicase inhibitor; (ii) TP84_27—holin, involved in cell membrane disintegration during host lysis; (iii) TP84_61—HNH homing endonuclease, which may be involved in recombination processes; (iv) TP84_26—glycosylase, possibly involved in disintegration of a very thick host’s envelope. This is the largest protein of TP-84 (112 kDa) and, interestingly, it shows homology to a number of proteins from Bacillus and Geobacillus species, and one Geobacillus bacteriophage GBK2 protein (GenBank YP_009010491). No RNA or DNA polymerase-coding genes were found on the TP-84 genome, thus the bacteriophage relies on the host’s transcription and replication enzymes. The TP-84 genome has a ‘condensed’ genome with CDSs, typically spaced by several-to-tens of bp only and often overlapping. The genome contains five putative promoter-like sequences resembling the host promoter consensus sequence with allowed 2-bp mismatches. Furthermore, ten putative rho-independent terminators were found. The TP-84 genome sequence shows essentially no similarity to any previously characterized bacteriophage, thus, TP-84 was established as a holotype of a new genus Tp84virus within the Siphoviridae family [2].
3.6. φμ-4 (Family/Genus Not Determined, Host B. stearothermophilus)
Bacteriophage φμ-4 was isolated from lysogenic B. stearothermophilus NU strain 4, and was propagated in B. stearothermophilus 10. It was cultivated at 50–65 °C with required Ca2+ ions. The latent period at 65 °C was 35 min with a burst size of 175. TEM images have shown the bacteriophage particles size of 88–100 nm. The particles shape was ‘roughly spherical’ [42].
3.7. TP-1 (Putative Siphoviridae Family, Host B. stearothermophilus)
TP-1 bacteriophage was isolated as a mitomycin- or UV-induced lysogenic B. stearothermophilus 1503-4R, growing in the narrow temperature range of 50–65 °C, with optimum at 55 °C. Ca2+ ions enhanced the bacteriophage propagation. TP-1 formed turbid plaques when titrated on B. stearothermophilus 4S. Furthermore, a clear-plaque mutant of TP-1C was found. Lysogen induction was rapid, causing a culture lysis within 45–60 min. Spontaneous induction was detected at a rate of 1 particle/app. 3 × 106 cells. Morphologically, TP-1 has a head of 65 nm and a long, flexible tail of 240 nm and 12 nm wide, which suggests that TP-1 belongs to the Siphoviridae family. The bacteriophage genome is composed of 42% G+C, dsDNA, with estimated molecular weight of 1.21 × 107 [43,44], which corresponds to approximately 18,516 bp. One of the authors [45] isolated a number of thermophilic bacteriophages, exemplified by TP-42, TP-56 and TP-68, but no characterization was provided.
3.8. ST1 (Siphoviridae Family, Genus ND, Host B. stearothermophilus)
Bacteriophage ST1 was propagated at 60 °C in B. stearothermophilus S13, forming clear plaques. TEM micrographs showed the head of ST1 as a regular icosahedral form of 50 nm in diameter, a tail of 100 nm length and 2 nm wide, ended with a terminal plate. The genetic material was dsDNA, containing 43% G+C [46].
3.9. Tφ3 (Tphi3) (Siphoviridae Family, Genus ND, Host G. stearothermophilus)
Tphi3 bacteriophage was isolated from a soil sample and infects the thermophilic B. stearothermophilus ATCC 8005. Along with Tphi3, there were two more bacteriophages isolated, apparently infecting B. stearothermophilus, not further characterized. Tphi3 is highly host-specific, as it does not infect B. stearothermophilus ATCC 7953, 7954, 10,149, 12,016. Optimal growth occurs at 60 °C with a latent period of 18 min and burst size of approximately 200. The Tphi3 exhibits high thermostability, up to 75 °C with a half-life of 12 min. Prolonged plates incubation resulted in a formation of a turbid ‘halo’ around clear plaques, suggesting a secretion of an enzyme, depolymerase, degrading the bacterial envelope. TEM imaging of Tphi3 shows that the bacteriophage has a 57 nm long regular icosahedron head. Edges of the head are 29 nm long, the tail is 125 nm long and 10 nm wide. The tail contains a stacked-ring structure. There are about 30 cross-strations that are spaced at 3.9-nm intervals along the tail and TEM images also suggest that the tail end contains small fibers. The buoyant density of the bacteriophage in a cesium chloride density gradient was 1.526 g/mL. Ca2+ ions were required for efficient adsorption and/or growth [47]. Further reported Tphi3 DNA studies, indicated that its average genome length was 12.1 mm, which corresponds to app. 23.2 MDa or app. 35,700 bp with G+C content of 40.2% [48].
3.10. GH5, GH8 (Family/Genus ND, Host B. stearothermophilus)
A brief report by Humbert and Fields (1972) showed two bacteriophages, GH5 and GH8, with distinct features, both infecting B. stearothermophilus NCA1518 [49]. Both share the same plaque-forming temperature range of 42.5–67 °C. Bacteriophage ND8 was examined under TEM and exhibited an icosahedral head with a diameter of approximately 100 nm, a long tail of 330 nm and approximately 10 nm wide, thus, apparently belonging to the Siphoviridae family. The latent periods were 47 min (GH5) or 35 min (GH8), burst size 51 (GH5) or 72 (GH8), buoyant density 1.473 (GH5) or 1.506 (GH8), sensitivity to osmotic shock from 2 M sucrose (both), sensitivity to osmotic shock from 4 M NaCl (GH5 no, GH8 yes), extrapolated 1 min half-life 88 °C (GH5) or 76 °C (GH8), maximum half-life at pH 9.0 (GH5) or 7.0 (GH8) [49].
3.11. PhB1 (Siphoviridae Family, Genus ND, Host Thermophilic Bacillus sp.)
PhB1 was isolated from a farm soil sample, using a soil-isolated bacterium as a host, described by the authors as facultatively thermophilic Bacillus sp. strain B1, Gram-positive rods, aerobic, spore-forming, motile with temperature growth range approximately 30–60 °C and optimum between 45–50 °C. The bacteriophage was cultivated on the hosts, growing at 55 °C. Small plaques were obtained with PhB1 and had a clear center with a diameter less than one millimeter, surrounded by a turbid halo [50], which suggests the presence of secreted envelope depolymerases. It was determined that Ca2+ ions and Mg2+ ions were stabilizing the bacteriophage particles against heat inactivation. TEM showed a long-tailed particle with a probable octahedral head and small spikes at the tail tip.
3.12. D5–D8 (Family/Genus ND, Host B. stearothermophilus)
A report by Reanney and Marsch (1973) presented 4 bacteriophages: D5, D6, D7, D8 infecting thermophilic B. stearothermophilus NRS T91 and ATCC7953. The hosts were grown on rich media supplemented with glucose and Ca2+ ions. Remarkably, the plaque-forming range for the bacteriophages extended down to the mesophilic region of 30–37 °C, with an optimum at 45 °C and reaching the upper limit at 55 °C. Bacteriophage proliferation in soil was greatest at 45 °C, while at 55 °C this was much less intense. The authors raised a hypothesis concerning the ecology of bacteriophages infecting the thermophilic soil organism B. stearothermophilus. The authors suggested that, in soil, these bacteria and their bacteriophages proliferate optimally at mesophilic rather than thermophilic temperatures. They proposed that, in the field, the vast majority of the metabolizing population of B. stearothermophilus are mesophilic in their biochemistry and that the ‘thermophilic’ strains, maintained in laboratories, represent a selection obtained by the isolation procedure of a minority of atypical, thermophilic cells [51].
3.13. φNS11 (PhiNS11) (Putative Sphaerolipoviridae, Tectiviridae or Corticoviridae Family, Host Acidophilic-Thermophilic B. acidocaldarius)
φNS11 is a ‘double’ extremophilic bacteriophage, as it infects both acidophilic and thermophilic bacterium, A. acidocaldarius strain TA6, growing in the Beppu hot springs in Japan. The bacteriophage was propagated in a pH range of 2 to 5 and exhibited optimum growth at 60 °C and pH 3.5. Above pH 6, it became unstable. φNS11 has a polyhedral shape, is tail-less and has a lipid-containing capsid with a diameter of 60–70 nm and spike-like structures. SDS-PAGE showed that φNS11 contains only six proteins. The genome was determined to be composed of dsDNA. The buoyant density of the bacteriophage was relatively low, 1.27 g/cm3, similar to that of the lipid-containing phages PM2, PR4, φ6. Furthermore, it was highly sensitive to organic solvents. The fatty acid composition of the bacteriophage was slightly different from that of the host bacterium. However, o-cyclohexyl fatty acids typical of B. acidocaldarius, were also found in φNS11 as its main components [52]. These features point to classification of φNS11 as belonging to the Sphaerolipoviridae, Tectiviridae or Corticoviridae families.
3.14. JS001-JS027 Series (Section 3.14, Section 3.15, Section 3.16, Section 3.17, Section 3.18, Section 3.19, Section 3.20, Section 3.21, Section 3.22, Section 3.23, Section 3.24, Section 3.25, Section 3.26, Section 3.27, Section 3.28, Section 3.29, Section 3.30, Section 3.31, Section 3.32, Section 3.33, Section 3.34, Section 3.35, Section 3.36 and Section 3.37 Below)
Sharp et al. (1986) have discovered and partially characterized 24 thermophilic bacteriophages, designated JS001-JS027, isolated from diversified sources: compost piles, soil, mud, sewage and river sludges, stable manure, silage and rotting straw, capable of infection of most of the major taxonomic groups of Bacillus thermophiles [21]. Various species/strains were used for the bacteriophages detection and propagation, including: ‘B. caldotenax’ DSM 406, DSM 411, ‘B. caldolyticus’ DSM 405, the Bacillus thermophile RS 93, B. stearothermophilus NCA 1503, NW 10, EP 240, EP 262, EP 136, DSM 2334, ATCC 12016 [53,54], streptomycin-resistant strains of: B. stearothermophilus NCA 1503, Bacillus thermophiles RS 239, RS 240, RS 241 and RS 242 [55]. The bacteriophages were uniformly detected/propagated in rich media of pH app. 7, and supplemented with Ca2+ ions, at 55 °C. They exhibited significant differences in capsid morphology, comprising 4 groups: (i) comparatively short stubby tail and no evidence of a tail plate; (ii) icosahedral heads of approximately 40–65 nm in diameter with tails between 120 nm and 135 nm long and no evidence of a tail plate; (iii) icosahedral head of 85–90 nm in diameter and a long (400 nm) flexible tail with helical symmetry; (iv) cylindrical heads with rounded ends, with tails varying between 115 nm and 120 nm long. All the genomes were determined as dsDNA. A number of isolates were lost when chloroform was used to sterilize them, indicating vital lipids content or denaturation-sensitive capsid protein(s). Table 2 contains selected data, concerning JS001–JS027.
3.15. JS001 (Family/Genus ND, Hosts Thermophilic Bacillus sp.)
JS001 was obtained by induction of B. stearothermophilus NCA 1503 and propagated on B. caldotenax. Other susceptible hosts included: B. stearothermophilus: ATCC 12016, RS 93, EP 262; Bacillus thermophile: RS 15, RS 108, RS 125 [21].
3.16. JS004 (Putative Podoviridae Family, Hosts Thermophilic Bacillus sp.)
JS004 was isolated from a silage, propagated on Bacillus thermophile RS 239, yielding small 0.5–1 mm circular, hazy plaques. High thermal stability was observed up to 60 °C, then gradually declining, but still substantial at 70 °C. Other susceptible hosts included: B. stearothermophilus RS 93, Bacillus thermophile RS 241. The bacteriophage belongs to the group 1: it possesses an icosahedral head 55–60 nm long, a short cylindrical 15 nm stubby tail with collar and no evidence of a base plate, which suggests that JS004 belongs to the Podoviridae family [21].
3.17. JS005 (Family/Genu ND, Hosts Thermophilic Bacillus sp.)
JS005 was isolated from rotting straw, propagated on Bacillus thermophile RS 239 and yielding small 0.5–1 mm circular, slightly hazy plaques. The only other susceptible host was Bacillus thermophile RS 241 [21].
3.18. JS006 (Family/Genus ND, Hosts Thermophilic Bacillus sp.)
JS006 was isolated from compost, propagated on Bacillus thermophile RS 239 and yielding small 0.5–0.75 mm circular, clear plaques. High thermal stability was observed up to 60 °C, then gradually declining, but still substantial at 70 °C. Other susceptible hosts included: B. stearothermophilus: RS 93, NW 4S, EP 262, Bacillus thermophile RS 242 [21].
3.19. JS007 (Family/Genus ND, Hosts Thermophilic Bacillus sp.)
JS007 was isolated from silage, propagated on Bacillus thermophile RS 240, and yielding small 0.5–0.75 mm circular, clear plaques. High thermal stability was observed up to 60 °C, then gradually declining, but still substantial at 70 °C. Other susceptible hosts included: B. stearothermophilus RS 93, and Bacillus thermophile RS 239, RS 240, RS 241. The bacteriophage belongs to the group 3: it possesses an icosahedral head 80–85 nm, an exceptionally long flexible tail of 400–420 nm with cross-strations and a fine fiber at tip, which suggests that JS007 belongs to the Siphoviridae family [21].
3.20. JS008 (Family/Genus ND, Hosts Thermophilic Bacillus sp.)
JS008 was isolated from rotting straw, propagated on Bacillus thermophile RS 241, yielding 1–3.5 mm circular, hazy plaques. Another susceptible host was Bacillus thermophile RS 239 [21].
3.21. JS009 (Family/Genus ND, Hosts Thermophilic Bacillus sp.)
JS009 was isolated from stable manure, propagated on Bacillus thermophile RS 242 and yielding small 0.5 mm circular plaques with a hazy center. Other susceptible hosts included: B. stearothermophilus RS 93, Bacillus thermophile RS 241 [21].
3.22. JS010 (Family/Genus ND, Hosts Thermophilic Bacillus sp.)
JS010 was isolated from compost, propagated on Bacillus thermophile RS 242 and yielding 3 mm, irregular, hazy plaques with clear areas at the edge. Other susceptible hosts included: B. stearothermophilus RS93, Bacillus thermophile RS 241 [21].
3.23. JS011 (Family/Genus ND, Hosts Thermophilic Bacillus sp.)
J7S011 was isolated from silage, propagated on Bacillus thermophile RS 239 and yielding 3 mm, irregular, hazy plaques with clear areas at the edge [21].
3.24. JS012 (Family/Genus ND, Hosts Thermophilic Bacillus sp.)
JS012 was isolated from compost, propagated on Bacillus thermophile RS 239 and yielding 0.5–2 mm, irregular, clear plaques [21].
3.25. JS013 (Family/Genus ND, Hosts Thermophilic Bacillus sp.)
JS013 was isolated from soil, propagated on B. stearothermophilus NCA 1503 and yielding 0.75–1 mm, circular, hazy plaques [21].
3.26. JS014 (Putative Siphoviridae, Hosts Thermophilic Bacillus sp.)
JS014 was isolated from rotting straw, propagated on B. stearothermophilus NCA 1503 and yielding small 0.5–1 mm circular, slightly hazy plaques. High thermal stability was observed up to 60 °C, then gradually declining, but still substantial at 70 °C. Another susceptible host was B. stearothermophilus NW 10. The bacteriophage belongs to the group 3: it possesses icosahedral head 85–90 nm, an exceptionally long flexible tail of 400 nm with cross-strations, which suggests that JS007 belongs to the Siphoviridae family [21].
3.27. JS015 (Family/Genus ND, Hosts Thermophilic Bacillus sp.)
JS015 was isolated from compost, propagated on B. stearothermophilus NCA 1503 and yielding 0.5–2 mm, circular, slightly hazy plaques. Another susceptible host was B. stearothermophilus NW 10 [21].
3.28. JS017 (Putative Myoviridae Family, Hosts Thermophilic Bacillus sp.)
JS017 was isolated from compost, propagated on B. caldotenax and yielding small 1–1.5 mm circular, turbid/hazy center plaques. High thermal stability was observed up to 60 °C, then gradually declining, but still substantial at 70 °C. Other susceptible hosts included: B. caldotenax DSM 406, B. caldovelox DSM 411, B. caldolyticus DSM 405, B. stearothermophilus: ATCC 12016, NW 10, RS 93, EP 262, EP 240, ATCC 8005, B. thermocatenulatus DSM 730. The JS017 belongs to the group 4: it possesses a cylindrical head with round ends 80 × 40 nm, a rigid tail of 115 nm with cross-strations and triangular base plate, which suggests that JS017 belongs to the Myoviridae family. Interestingly, a 1–5% portion of the bacteriophage population had heads of twice the normal length and/or less frequently longer tails [21].
3.29. JS018 (Putative Myoviridae Family, Hosts Thermophilic B. caldotenax)
JS018 was isolated from rotting vegetation, propagated on B. stearothermophilus NCA 1503 and yielding 1 mm, circular, clear center plaques. Other susceptible hosts included: B. caldotenax DSM 406, B. caldovelox DSM 411; B. caldolyticus DSM 405, B. stearothermophilus: RS 93, EP 240 [21].
3.30. JS019 (Putative Myoviridae Family, Hosts Thermophilic B. caldotenax)
JS019 was isolated from rotting vegetation, propagated on B. caldotenax and yielding small 1–1.5 mm circular, clear center, hazy at edges plaques. High thermal stability was observed up to 60 °C, then gradually declining, but still substantial at 70 °C. The bacteriophage exhibited broad specificity. Other susceptible hosts included: B. caldotenax DSM 406, B. caldovelox DSM 411; B. caldolyticus DSM 405, B. stearothermophilus: RS 93, EP 240. The JS019 belongs to the group 2: it possesses an icosahedral head of 50 nm and rigid tail of 130, which suggests that JS019 belongs to the Myoviridae family [21].
3.31. JS020 (Family/Genus ND, Hosts Thermophilic B. caldotenax)
JS020 was isolated from rotting vegetation, propagated on B. caldotenax and yielding 1–2 mm, circular, clear center, hazy outer halo 4–8 mm plaques. This suggest that the bacteriophage produces very active depolymerase, degrading the host’s envelope. Other susceptible hosts included: B. caldotenax DSM 406, B. caldovelox DSM 411, B. caldolyticus DSM 405, B. stearothermophilus: RS 93, EP 240 [21].
3.32. JS021 (Family/Genus ND, Hosts Thermophilic B. caldotenax)
JS021 was isolated from rotting vegetation, propagated on B. caldotenax and yielding 2–3 mm, circular, clear centre, hazy outer halo plaques. This suggests that the bacteriophage produces an envelope depolymerase. Other susceptible hosts included: B. caldotenax DSM 406, B. caldovelox DSM 411; B. caldolyticus DSM 405, B. stearothermophilus: RS 93, EP 240, ATCC 12016 [21].
3.33. JS022 (Putative Myoviridae Family, Hosts Thermophilic B. caldotenax)
JS022 was isolated from compost, propagated on B. caldotenax and yielding 1–2 mm, circular, clear centre plaques. Other susceptible hosts included: B. caldotenax DSM 406, B. caldovelox DSM 411; B. caldolyticus DSM 405, B. stearothermophilus: RS 93, EP 240. The JS022 belongs to the group 2: it possesses an icosahedral head 60–65 nm long and a rigid tail of 130 nm, which suggests that JS022 belongs to the Myoviridae family [21].
3.34. JS023 (Family/Genus ND, Hosts Thermophilic B. caldotenax)
JS023 was isolated from compost, propagated on B. caldotenax and yielding 0.5–1.5 mm, circular, clear centre, hazy edge plaques. Other susceptible hosts included: B. caldotenax DSM 406, B. caldovelox DSM 411; B. caldolyticus DSM 405, B. stearothermophilus: RS 93, EP 136 [21].
3.35. JS024 (Putative Siphoviridae or Myoviridae Family, Hosts Thermophilic B. caldotenax)
JS024 was isolated from compost, propagated on B. caldotenax and yielding 0.5–1.5 mm, circular, clear center, hazy edge plaques with a less hazy outer edge. Other susceptible hosts included: B. caldotenax DSM 406, B. caldovelox DSM 411; B. caldolyticus DSM 405, B. stearothermophilus: RS 93, EP 240, EP 136. The JS024 belongs to the group 2: it possesses an icosahedral head 60 nm long and a tail of limited flexibility 120 nm, which suggests that JS024 belongs to either the Siphoviridae or Myoviridae family [21].
3.36. JS025 (Putative Myoviridae Family, Hosts Thermophilic B. caldotenax)
JS025 was isolated from compost, propagated on B. caldotenax and yielding 1–1.5 mm, circular, hazy plaques. Other susceptible hosts included: B. caldotenax DSM 406, B. caldovelox DSM 411; B. caldolyticus DSM 405, B. stearothermophilus: RS 93, EP 240. The JS025 belongs to the group 2: it possesses an icosahedral head 45–50 nm long and a rigid tail of 120 nm, which suggests that JS025 belongs to the Myoviridae family [21].
3.37. JS026 (Putative Siphoviridae or Myoviridae Family, Hosts Thermophilic B. caldotenax)
JS026 was isolated from compost, propagated on B. caldotenax and yielding 1–1.5 mm, circular, hazy plaques. Other susceptible hosts included: B. caldotenax DSM 406, B. caldovelox DSM 411; B. caldolyticus DSM 405, B. stearothermophilus: RS 93, ATCC 12016. The JS026 belongs to the group 4: it possesses a cylindrical head with round ends 80 × 45–50 nm and a limited flexibility tail of 120 nm, which suggests that JS026 belongs to either the Siphoviridae or Myoviridae family [21].
3.38. JS027 (Putative Podoviridae Family, Hosts Thermophilic Bacillus sp.)
JS027 was isolated from compost, propagated on Bacillus thermophile RS241 and yielding 0.75 mm, circular, clear plaques. Another susceptible host is Bacillus thermophile RS239. The JS027 belongs to the group 1: it possesses an icosahedral head 55–60 nm long and a very short, stubby tail with no evidence of a tail plate, which suggests that JS027 belongs to the Podoviridae family [21].
3.39. W1 (BVW1) (Siphoviridae Family, Genus ND, Host Thermophilic Bacillus sp.)
W1 (BVW1) was isolated from a deep-sea hydrothermal field, located in the west Pacific (19_ 24′08″ N, 148_44′79″ E, depth of 5060 m), along with its host Bacillus sp. W13 (GenBank AY583457), which is aerobic, rod-shaped and spore-forming, growing within a temperature range of 45–85 °C with an optimum at 65–70 °C. The host range was narrow, as the W1 infects only one thermophilic strain out of 10 tested. The bacteriophage propagates most effectively at 60 °C. Morphologically, W1 has a hexagonal head of 70 nm in diameter and a very long tail of 300 nm and 15 nm in width. The bacteriophage genome is composed of dsDNA with an estimated size of 18 kb. As determined, using SDS-PAGE of purified virions, six major proteins were detected with estimated molecular weights: 32, 45, 50, 57, 60 and 70 kDa [22].
3.40. GVE1 (E1) (Siphoviridae Family, Genu NDs, Host Geobacillus sp.)
GVE1 was isolated from a deep-sea hydrothermal field, located in the east Pacific (12_42′29″ N, 104_02′01″ W, depth of 3083 m), along with its host Geobacillus sp. E26323 (GenBank DQ225186), which is aerobic, rod-shaped and spore-forming, growing in rich media of pH 7, within a temperature range of 45–85 °C and an optimum at 65–70 °C. The host range is narrow, as the GVE1 infects only one thermophilic strain out of 10 tested. The bacteriophage propagates most effectively at 60 °C. GVE1 possesses a large hexagonal head of 130 nm in diameter, a tail of 180 nm and 30 nm in width. The bacteriophage genome is composed of dsDNA with an estimated size of 41 kb. As determined on SDS-PAGE of purified virions, six major proteins were detected with estimated molecular weights: 34, 37, 43, 60, 66 and 100 kDa [22].
3.41. GVE2 (E2) (Siphoviridae Family, Genus ND, Hosts Geobacillus sp.)
Siphovirus GVE2 (also known as E2) is one of the very few deeply studied Geobacillus thermophages and was isolated from the same location as GVE1 [22]. The authors in the publications concerning GVE2, refer to their previous publication concerning the discovery of GVE1, thus, it is possible that the bacteriophages are identical or closely related.
GVE2 is a virulent Siphoviridae bacteriophage infecting deep-sea thermophilic Geobacillus sp. E263 (CGMCC1.7046), capable of growth at 45–80 °C, with optimum at 60–65 °C, which was selected for GVE2 propagation. The bacteriophage genome is a 40,863 bp, 44.8% G+C, linear dsDNA with 62 ORFs [22]. Its genome shows sequence similarity to the genomes of several Geobacillus sp. bacteriophages. Six GVE2 proteins were characterized in the laboratory. Transcriptomic analysis has shown that 74.2%, 46 of the ORFs, were transcribed and potentially expressed as proteins. The six structural proteins of purified GVE2 virions were also determined by mass spectrometry and, for one of those, novel portal protein VP411 protein, its coding gene was cloned/expressed in Escherichia coli (E. coli). The recombinant VP411 was characterised in details and confirmed by immune electron microscopy using gold-labelled secondary antibody, as located in the portal-neck region of the GVE2 [24]. Subsequently, the endolysin coding gene was cloned in E. coli, expressed and the protein purified and characterised [56]. Further, the holin-endolysin system was studied and it was shown that GFP–endolysin fusion aggregated in GVE2-infected Geobacillus sp. and revealed that the endolysin and holin interacted directly as well, as with the host Geobacillus protein ABC transporter system, which apparently participates in the regulation of the lysis process [57].
3.42. GBSV1 (Myoviridae Family, Svunavirus Genus, Hosts Geobacillus sp.)
GBSV1 was isolated from an off-shore location in shallow waters at the depth of 100 m. It was propagated in thermophilic Geobacillus sp. 6k51 (GenBank DQ141699), which is an aerobic rod-shaped bacterium growing at 45–85 °C [25]. It was cultivated and infected with GBSV1 at 65 °C, supplemented with Ca2+ and Mg2+ ions at pH app. 7. The 16S rDNA analysis revealed that this host shared 99.93% sequence identity with that of G. toebii mc-14 (GenBank EU214628). TEM analysis of GBSV1 showed a typical Myoviridae morphology with a 60 nm hexagonal head and 80–150 nm contractile tail. The GBSV1 genome analysis is composed of linear, 44.4% G+C dsDNA of 34,683 bp, coding for 54 putative ORFs. Thirty-three of them exhibit sequence similarities to genes from seven species of Geobacillus or Bacillus and to the bacteriophages that infect them. This may indicate their biological role of upmost importance in promoting genetic exchanges among Bacillus and/or Geobacillus bacteria: short-term adaptation, long term evolution of bacteria and population genetics. Twenty-two ORFs were bioinformatically annotated and five structural proteins of the purified GBSV1 were identified by proteomic analyses. The ORFs are grouped into functional clusters on the genome. Interestingly, seven of the GBSV1 ORFs exhibit sequence similarities with the genes from the bacteria relevant to human diseases. The authors hypothesize that thermophages may be the potential evolutionary connection between thermophiles and human pathogens. The GBSV1 shares over 90% proteome similarity with BV1, as based on genomes analysis. Two groups of genes exhibit high similarity between GBSV1, BV1 and phiOH2: (i) lysis-related proteins—a probacteriophage anti-repressor and lysin with >95% similarity, and (ii) bacteriophage genome maintenance proteins and integration—integrase, >95%, recombinase—>65% similarity [25]. These similarities indicate common, possibly interchangeable, infection modules across thermophilic host species of distinct bacterial phyla [3].
3.43. BV1 (Myoviridae, Family, Svunavirus Genus, Hosts Geobacillus sp.)
BV1 is highly similar to GBSV1 (see previous section) and the characteristics are essentially the same, with minor differences: a hexagonal head is 60 nm in diameter and a tail 90nm in length and 10 nm in width, the genome is 44.8% G+C, 35,055 bp. It contains 54 ORFs, classified into eight functional groups. SDS-PAGE analysis of purified BV1 revealed that it contains seven major polypeptides with molecular weights of app.: 100, 72, 42, 33, 31, 28 and 26 kDa. BLAST analysis revealed that 38 ORFs exhibited similarities to proteins present in databases. Interestingly, a diverse pattern of similarities suggests that BV1 could be involved in gene transfer between the Bacillus and Geobacillus species [58].
3.44. D6E (Myoviridae Family, Genus ND, Host Geobacillus sp.)
D6E was isolated from a deep-sea hydrothermal field (12°42′29″ N, 104°02′01″ W, depth of 3083 m) at the same location as GVE1, along with the same host Geobacillus sp. E26323 and was propagated at 65 °C. TEM results have shown that D6E is a myovirus with an icosahedral head (60 nm), a contractile tail (16 nm in width and 60 nm in length), decorated with a tail fiber (4 nm in width and 60 nm in length). The bacteriophage genome of 49,335 bp is composed of 46% G+C dsDNA, possibly circular, as based on restriction and PCR analysis. It contains 49 ORFs, with 30 of them having bioinformatically assigned homologies/function, grouped in functional clusters: (i) DNA packaging and capsid assembly, (ii) tail assembly, (iii) lysis and lysogeny, (iv) DNA replication and transcription. D6E genome does not code for any tRNA genes. Comparative genomics and proteomics analyses have indicated an extensive mosaicism of D6E genome with other mesophilic and thermophilic bacteriophages, including GVE1. The authors conclude, that there were at least three recombination events during GVE1 evolution: (i) the overall gene organization, (ii) recombination between gene boundaries and (iii) mutations in genes. As determined on SDS-PAGE and mass spectrometry 10 virion-associated proteins biosynthesis were confirmed [14].
3.45. ϕOH2 (phiOH2) (Family/Genus ND, Host G. kaustophilus)
Bacteriophage ϕOH2 is closely grouped with Bacillus virus 1 BV1 thermophage [3]. It was found in the sediment of a hot spring as lytic, infecting factor for G. kaustophilus NBRC102445. The genome of bacteriophage ϕOH2 is integrated into the lysogenic host genome G. kaustophilus GBlys, thus being a prophage. The host was cultivated at 55 °C. Its genome of 3,541,481 bp consists of 216 sequence contigs with OH2 prophage genome (38,099 bp, 45% G = C, 60 ORFs), situated astride contigs GBL0024 and GBL0110 and flanked by the host genes encoding GroES, GroEL. The host transposase gene is located next to the OH2 genome. A putative integrase gene is located at the terminus of the OH2 sequence. There are no excisionase gene homologues, and the attachment sites located next to the integrase gene are dissimilar to those coded by bacteriophages [59]. BLAST analysis show the presence of completely or partially homologous ϕOH2 sequences in many ‘Bacillus group’ bacteria and bacteriophages, which indicates intense horizontal gene transfer within the group.
3.46. GBK2 (Siphoviridae Family, Genus ND, Host G. kaustophilus)
GBK2 bacteriophage was extracted out of the compost pile using a liquid culture of G. kaustophilus ATCC8005, cultivated at 55 °C. GBK2 has a circularly permuted dsDNA genome of 39,078 bp and 43% G+C content. The genome contains seven copies of a 17-bp inverted DNA repeat, which forms a potential origin of replication for GBK2. Bioinformatics analysis of the genome reveals 62 putative ORFs. Twenty-five of them show homology to the known proteins and 37 are proteins with undetermined functions. The ORFs are grouped into functional clusters: (i) DNA packaging, (ii) structural proteins, (iii) lysis, (iv) unknown, (v) DNA replication/metabolism. Overall, the genome exhibits a limited DNA sequence similarity to any other sequenced bacteriophages [1]. It is intriguing that GBK2 is more closely related to the bacteriophages that infect mesophilic hosts than to those that infect thermophilic Geobacillus sp. or other related thermophiles species. The overall genomic arrangement of GBK2 is quite similar to that of mesophilic bacteriophage SPP1, infecting B. subtilis [22]. Fourteen ORFs are coding proteins that are homologous to SPP1 bacteriophage and most of them are involved in DNA replication, while two ORFs are predicted to encode capsid structural proteins. DNA-related genes, include those coding for: exonuclease/recombinase, recombinase of RecT-type, helicase, helicase loader and three DNA metabolism enzymes [18]. Furthermore, in SPP1 genome there are three genes, non-essential for growth, of unknown function. Nevertheless, homologs of these genes are found in GBK2 bacteriophage. It was concluded that they may play a significant role in the thermophage life cycles under certain conditions [60]. These functional and sequence similarities point to a common origin of SPP1 and GBK2 bacteriophages or a massive horizontal gene transfer.
3.47. GVE3 (Siphoviridae Family, Genus ND, Host G. thermoglucosidasius)
GVE3 bacteriophage is a B1 morphotype group siphovirus that infects, very specifically, the thermophilic bacteria G. thermoglucosidasius as the only susceptible host, propagated at 60 °C. The bacteriophage has an isometric head of 90–100 nm diameter and a non-contractile tail of 210 nm. The plaques had characteristic “bulls-eye” morphology, suggesting host lysogeny [61] with several bacterial colonies growing inside plaques. Once isolated, the bacteria from those colonies were resistant to GVE3 infection. The lysogenic host genome sequencing revealed that the GVE3 genome had inserted into the specific location, attB site, with a 23-bp sequence that was duplicated upon insertion. The bacteriophage GVE3 encodes a putative pyrimidine nucleoside phosphorylase, situated downstream of GVE3 resolvase gene. The attP site is located within the gene. Apparently, the host’s pyrimidine nucleoside phosphorylase is inactivated upon the GVE3 genome insertion.
The presence of a GVE3-encoded pyrimidine nucleoside phosphorylase suggests an obligatory requirement for its activity: upon GVE3 genome integration the host relies on the bacteriophage enzyme. There is an alternative explanation: if the enzyme is not essential for either the GVE3 or the host, since the mutation in the pyrimidine nucleoside phosphorylase-coding gene is non-lethal, this may suggest that GVE3 is a specialized transducing thermophage [62]. Interestingly, the genome size is 141,298 bp comprising the largest known among the bacteriophages that infect Geobacillus sp., and has a unique DNA sequence, with no close relatives. A large number of 202 putative ORFs were bioinformatically detected and, out of those, putative functions to 62 of them were assigned. As observed in a number of other members of the Siphoviridae, the GVE3 genome displays the functional clusters [62]. Remarkably, GVE3 resembles GBK2 in that it is apparently most closely related to the mesophilic B. anthracis bacteriophage vB_BanS_Tsamsa, rather than the Geobacillus-infecting bacteriophages. Tetranucleotide usage deviation analysis confirms this finding, indicating that the GVE3 genome sequence correlates mostly with B. anthracis and B. cereus sequences, rather than Geobacillus sp. sequences [62,63], which again points to widespread genetic recombination among meso- and thermophilic ‘Bacillus group’ bacteriophages. The analysis also suggests that there exist a different ‘natural’ host, unidentified Geobacillus sp., with tetranucleotide usage deviation patterns more similar to the two Bacillus species that might be the “natural” hosts for GVE3. Alternatively, the possibility exists that GVE3 has evolved from a mesophilic counterpart and that the correlation to mesophilic Bacillus species is an evolutionary link to its heritage. Similar findings have been shown for GBK2 [64]. The GVE3 genome sequence has a much lower G+C content (29.6%) than its host G. thermoglucosidasius (44%) [62], which is also observed for most bacteriophage–host pairs. It is suggested that its base composition bias might have resulted from a competition for metabolic resources and/or reflects an adaptation to thermophily [65]. The genome GC skew analysis has suggested a location of a replication terminus. A search for direct and inverted repeats of 7–30 bp resulted in finding as many as 582 repeats, including two invert repeats (TATTTTTT/TAATTAT) predicted to be part of the origin of the replication. The large genome segment, DNA replication, modification and repair, contains tens of genes, not all of them with assigned functions. These make the GVE3 enzymatic machinery remarkably independent, including three subunits diverging hosts DNA polymerase to the bacteriophage replication and three DNA modification methyltransferases, protecting the genome from restriction, among others [62].
3.48. AP45 (Siphoviridae Family, Genus ND, Host Aeribacillus sp.)
AP45 bacteriophage was isolated from a soil sample collected in the Valley of Geysers, Kamchatka, Russia. It was propagated in thermophilic Aeribacillus sp. strain CEMTC 656, a Gram-positive, aerobic, rod-shaped, non-motile, forming sub-terminally located endospores. The optimum growth temperature was at 50–65 °C in nutrient broth at pH 6.5, containing 0.3 M NaCl. AP45 was propagated at 55 °C. The host range is very narrow, out of 13 tested Geobacillus sp., only G. icigianus CMBI G1w1, also isolated from the Valley of Geysers, was supporting infection. AP45 exhibited high thermostability and a latent period of approximately 200 min, with a burst size of approximately 40 bacteriophage particles/infected cell. The bacteriophage produces both a ‘halo’ around clear plaque centers and ‘bulls-eye’ morphology, indicating secretion of a cell envelope depolymerase and suggesting formations of host lysogeny. The colonies isolated from the plaques proved to be resistant to AP45 infection, however, PCR using primers to AP45 has failed, thus, it is not clear whether other resistance mechanisms take place. TEM analysis of AP45 indicated Siphoviridae morphology with an icosahedral head of 60 nm with a diameter and a tail approximately 160 nm in length. The AP45 genome analysis has shown that it is composed of 38.3% G+C dsDNA of 51,606 bp, containing 71 ORFs, 40 of them have had putative functions predicted bioinformatically. Furthermore, ORFs were divided into three functional clusters: (i) nucleic acids metabolism and regulation of the viral life cycle; (ii) structural proteins; (iii) lysis of bacterial cells. No genes encoding DNA and RNA polymerases were identified, thus, AP45 apparently uses host polymerases. The AP45 genome exhibited limited similarity to other bacteriophage sequences. The highest value of 36% was detected for thermophilic Geobacillus myovirus D6E. Interestingly, the majority of putative AP45 proteins exhibited a higher similarity to proteins from bacteria belonging to the Bacillaceae family, than to bacteriophages. Moreover, over half of the putative AP45 ORFs were highly similar to probacteriophage sequences of the Aeribacillus pallidus strain 8m3. It was suggested that the AP45 bacteriophage and revealed probacteriophages might be members of a new genus within the Siphoviridae family [27].
3.49. Series of 5 Bacteriophages from Haloalkaline Lake Elmenteita of Kenyan Rift Valley (Section 3.49, Section 3.50, Section 3.51, Section 3.52, Section 3.53 and Section 3.54 Below)
All bacteriophages were isolated from Lake Elmenteita [66], situated at 0°27′ S 36°15′ E on the floor of the Kenyan Rift Valley, at 1776 m above sea level without a direct water outlet. This area has a hot, dry and semi-arid climate, which results in rapid evaporation rates. The alkalinity of the water is high alkaline with pH above 9 and with a high concentration of carbonates, chlorides and sulphates [67]. The water temperature ranges between 30 °C and 40 °C. Comparison of genome sizes, coding density, number of ORFs, GC% and gene organizations revealed that the bacteriophages had no relationship with each other as well as no significant similarities in the GenBank® sequence data. Furthermore, there is evidence of genes mosaicism to described genes with conserved domains [66]. The bacterial hosts were not described in details. Nevertheless these were extremophilic bacteria with regard to salinity and alkalinity from the ‘Bacillus group’, possibly also entering moderate thermophilic range. No bacteriophages characteristics are available at the time of this review submission, except bacteriophages genomes bioinformatics analysis. The authors cite ongoing publication, which will contain descriptions of the bacteriophages [66].
3.50. vB_BpsS-36 (Family/Genus ND, Host B. pseudalcaliphilus)
The vB_BpsS-36 genome is composed of 50,485 bp dsDNA with 41.1% GC content. It codes for 68 ORFs distributed on both forward and reverse strands and six transcription terminators with coding density of 91.6%. On the DNA level, the bacteriophage shows essentially no homologies to other bacteriophages, while on the amino acids level there exists weak similarities to short regions of Bacillus bacteriophages phages Tsamsa, Riggi and CampHawk. A conserved replication factor was detected in the genome, suggesting that vB_BpsS-36 employs DNA polymerase III subunit alpha for DNA replication. Furthermore, vB_BpsS-36 codes for replication proteins—domains DnaB and DnaG [66].
3.51. vB_BpsM-61 (Family/Genus ND, Host B. pseudofirmus)
The vB_BpsM-61 genome is composed of 48,160 bp dsDNA with 43.5% GC content. It codes for 75 ORFs distributed on both forward and reverse strands and eight transcription terminators with coding density of 93.0%. The bacteriophage exhibits weak similarities to Bacillus bacteriophage PM1 and Geobacillus phage GBK2 in cluster coding for structural components. The vB_BpsM-61 genome is also coding for putative proteins: replicative DnaC, helix destabilizing Ssb, dUTPase, Holliday junction resolvase [66].
3.52. vB_BboS-125 (Family/Genus ND, Host B. bogoriensis)
The vB_BboS-125 genome is composed of 58,528 bp dsDNA with 48.6% GC content. It codes for 81 ORFs distributed on forward strands only and six transcription terminators with coding density of 92.2%. The bacteriophage exhibits weak similarities to other bacteriophage proteins, more pronounced to structural components and DNA packaging located in genomic regions in Exiguobacterium sp. AB2 and Brevibacillus sp. CF112, apparently being not-annotated prophage segments. In addition, the vB_BboS-125 shows similarities to genes with conserved domains of the replication cluster, helicase and primase, present in the genome of Brevibacillus sp. CF112. Furthermore, the vB_BboS employs a host’s DNA polymerase I for replication. Interestingly, the bacteriophage terminase did not cluster with previously described bacteriophages, but formed a distinct phyletic line, which suggests that vB_BboS-125 belongs to a new genus of bacteriophages [66].
3.53. vB_BcoS-136 (Family/Genus ND, Host B. cohnii)
The vB_BcoS-136 has the largest genome of an entire group of eight bacteriophages found in haloalkaline in Lake Elmenteita, 160,590 bp dsDNA with low GC content of 32.2%. It codes for 240 ORFs distributed on both forward and reverse strands, 15 transcription terminators and 17 tRNA with a coding density of 93.0%. A very high number of tRNA coding genes suggest substantial differences in codon usage between the bacteriophage and its host, thus, needed for efficient translation and vB_BcoS-136 propagation. Furthermore, it may suggest that the bacteriophage relatively recently has adopted its host. The bacteriophage exhibits very low similarities to Bacillus bacteriophage Tsamsa. BcoS-136 terminase is dissimilar to other bacteriophage’s terminases with an already known DNA packaging strategy, but groups in a distinct phyletic line, thus pointing to the possibility of the existence of another new genus. vB_BcoS-136 utilizes a host’s DNA polymerase III subunit alpha and codes for other DNA replication/DNA metabolism putative proteins: DNA ligase, DNA gyrase, ribonuclease HI and integrase. The latter suggests that vB_BcoS-136 can also enter lysogenic mode [66].
3.54. vB_BpsS-140 (Family/Genus ND, Host B. pseudalcaliphilus)
The vB_BpsS-140 genome is composed of 55,091 bp dsDNA with 39.8% GC content. It codes for 68 ORFs distributed on forward strands only and four transcription terminators with coding density of 91%. The vB_BpsS-140 encoded genes for proteins similar to homologs of a terminase and some structural proteins of Bacillus bacteriophages IEBH and 250, respectively. Homology comparison of the large terminase subunit shows that vB_BpsS-140 clusters with P22 bacteriophage, which is established as a model bacteriophage that follows headful packaging strategy [68]. As this protein is considered the most universally conserved gene sequence among bacteriophages [69], it is used to get insight into evolutionary relationships among distant bacteriophages [70]. Interestingly, the bacteriophage codes for atypical endopeptidase endolysin, are absent in other bacteriophages published by the authors [66].
4. Scientific, Biotechnology, Environmental and Medical Potential—A Short Note
In general, bacteriophages, and their corresponding enzymatic activities, offer great potential for the development of agents for the control of human pathogens [71]. Bacteriophage bionanostructures are used in science, industry and medicine, as exemplified by bioimaging diagnostics, ultrasensitive biomarker detection, targeted drug and gene delivery, improved tissue formation, directed stem cell differentiation, new generation vaccines and nanotherapeutics for targeted disease treatment, among others [71]. These applications may be substantially improved if based on robust, thermostable bacteriophage bionanoparticles. The study of Geobacillus host bacteria, their bacteriophages and their mutual interaction, presents great potential for gaining knowledge about the functioning of high temperature ecosystems, including the thermophages crucial role as regulators of bacterial populations. No less importantly, this knowledge is likely to help improve the known processes or to develop new processes in biotechnology. The list of viral genome sequences reported thus far in the National Center for Biotechnology Information Genome database, as of June 2021, includes 10,607 genomes; 3746 of them being bacterial hosts, with only four hosts listed under ‘Geobacillus’. This proportion shows that there are significant gaps in the understanding of the genetics of the ‘Bacillus group’ thermophages as well as of the type and dynamics of their biological processes at a local and global scale.
Even though thermophiles and extreme thermophiles are presently attracting substantial attention, the area remains in large part a ‘terra incognita’ in terms of approaching understanding of mesophilic host-bacteriophage systems (such as E. coli/bacteriophage lambda). Nevertheless, they can provide model systems for deciphering the molecular biology and biochemistry of adaptations towards life at high temperatures and their effects on atypical ecosystems, and can affect many biogeochemical and ecological processes.
The scientific interest in thermophilic bacilli, especially Geobacillus sp., is fully justified, due to their potential applications in biotechnology and environment remediation processes. Nevertheless, thermophilic bacteriophages can cause huge economical losses, upon undesired infections of industrial fermentations. Bacteriophage-sensitive processes are shown in the following examples.
As Geobacillus sp. possesses the ubiquitous capability to metabolize various organic compounds, including hydrocarbons, the fermentation of C-5 and C-6 sugars is being used to biosynthesize ethanol for biofuel and organic acids production [72]. Even very ‘difficult’ substrates, such as cellulose, the most abundant organic carbon reservoir on Earth, are effectively degraded by Geobacillus sp. cellulases.
More thermostable enzymes, coded by Geobacillus sp., are also of interest to the biotechnology industry, such as: proteases, amylases, polysaccharides, lipases, glycanases and herbicides metabolizing enzymes for agricultural biotechnology [10,73,74,75,76].
Certain Geobacillus sp. are also capable of the destruction of quorum sensing in some Gram-negative bacteria [10]. Furthermore, some strains of G. thermoleovorans are secreting large bacteriocins that cause lysis of various other bacteria, including pathogenic Salmonella typhimurium [77]. These capabilities could be a basis for future therapeutic methods. Thus, cataloguing bacteriophages and studying their interactions with their hosts is also important for practical purposes. Furthermore, bacteriophages can affect the human microbiome, including facultatively thermophilic ‘Bacillus group’ species. Several clades from genus Bacillus of the Bacillaceae family involve bio-safe species, regarded as bimodal probiotic microbiobiota of humans and animals [78]. These are strains generally derived from environmental locations such as soil, marsh and plants, but are also found in animal’s and human’s skin, gastrointestinal tract or respiratory system. Several Bacillus species are typically encountered in traditional fermented foods, especially those of Asian or African origin that are prepared prevalently from protein rich soybeans or locust beans. The outstanding potential for the survival of environmental probiotics results from their ability to form durable spores [79] capable of enduring harsh conditions of draught, UV exposure, extreme temperatures and pH values, the presence of organic and inorganic chemicals, salts, detergents and heavy metal ions. During the past decade, probiotic Bacillus species, including B. subtilis, B. coagulans, B. clausii and B. licheniformis, are of growing interest and seem to be promising models for biotechnological applications, involving industrial, agricultural, medical or, typically, scientific applications [80,81]. Pathogenic activity cases of ‘Bacillus group’ are extremely rare [82], thus, no phage therapy against these bacteria has been developed for human use. However, for animal protection purposes, successful phage therapy has been used, employing FBL1 bacteriophage preparation for combating the mortality of the white Pacific shrimp, Litopenaeus vannamei, infected by B. licheniformis. Some strains of these bacteria are capable of extending their upper temperature growth limit up to approximately 50 °C, thus stepping into the thermophilic range. The FBL1 not only impaired its host growth, but also prevented the formation of biofilm [83]. Besides biofilm degradation/prevention, there are other molecular mechanisms that can be exploited to control bacterial population. The bacteriophage SPO1 of B. subtilis is coding for Gp44 protein, which is part of an unusual molecular mimicry strategy inhibiting host RNA polymerase. Gp44 is composed of a DNA binding motif, a flexible DNA mimic domain and a random-coiled domain. Such composition allows selective binding to bacterial RNA polymerase through β and β’ subunits, blocking bacterial growth. This a non-specific mechanism that SPO1 employs to target different bacterial transcription apparatus, regardless of the structural variations of RNA polymerases. This opens up the possibility for the generation of genetically modified SPO1 for the purpose of targeted gene therapy [84,85].
It is known that these industrial and medical biotechnology applications can be dramatically affected by undesired thermophage infections. On the other hand, thermophages are important in biotechnology, not only because of their potential to cause big economic losses but also because they also code for a number of valuable enzymes, which are often more robust than those of their hosts due to the rapid nature of a high temperature thermophage infection requirements. Some of these enzymes, with a big potential for novel applications, are the less recognized endolysins-lysozymes and depolymerases-glycosidases of thermophage origin [1,86,87]. These enzymes could be used for treating industrial installations overgrown with various bacteria as well as in biofilm removal for medical usage [1,86,87]. Since the critical initial step in a bacteriophage infection is the adsorption to its compatible receptor on the surface of the bacterial cell, the presence of exopolysaccharide envelopes can effectively block this stage. Some bacteriophages are producing a ‘halo’ around clear plaque centers, indicating a diffusing factor, hydrolyzing and allowing for the penetration of the extracellular exopolysaccharide matrix—a major bacterial biofilm component [88,89,90]. A number of thermophages produce thermostable depolymerases-glycosylases, endolysins (lysozymes) and holins, involved in actions against external bacterial layers, capsule, cell wall, cell membrane, protecting the internal cell cytoplasmatic environment [64]. The thermophage TP-84 codes for all three types of enzymes [2]. The combination of three such types of thermostable, robust enzymes in a cocktail for biofilms removal may become a powerful tool in industrial, environmental and medical applications. The ability of bacteria to form biofilms is an important element of their pathogenicity, and biofilm destruction is of high interest in medicine as a potential alternative or supplement to existing therapies [91]. TP-84 is a bacteriophage that deprives G. stearothermophilus bacteria of their capsule, thus facilitating the permeation of bacteriophages into deeper biofilm layers and lysis of the susceptible bacterial cells. Considering the rather broad specificity of TP-84 lytic enzymes, if applied to pathogen biofilms, it may prevent their formation and contribute to eradication of the biofilm bacteria [91]. Furthermore, thermophages often produce other useful enzymes, for example, GVE3 is encoding DNA ligase, DNA polymerase, among others [92]. Additionally, there are works being conducted to develop a GVE3 thermophage-based system for the introduction of novel or engineered metabolic and biosynthetic pathways [61]. Some of the lysogenic thermophages contain integrases, which are of growing importance for the genetic engineering of living eukaryotic cells, as they can mediate precise site-specific recombination between two different sequences [93,94]. An application unexplored thus far would be the construction of a robust, thermostable phage display system for directed evolution in vitro. Such systems have made a major breakthrough in molecular biology and biotechnology, so that the inventors were honored with the 2018 Nobel Prize.
Author Contributions
Conceptualization, B.Ł. and P.M.S.; methodology, B.Ł., P.M.S., I.S., J.J.-F.; resources, B.Ł. and P.M.S.; writing—original draft preparation B.Ł. and P.M.S.; writing—review and editing, B.Ł., I.S., J.J.-F. and P.M.S.; supervision P.M.S.; project administration, B.Ł. and P.M.S.; funding acquisition P.M.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by the National Center for Research and Development (Poland) grant TECHMATSTRATEG2/410747/11/NCBR/2019 to Piotr Skowron.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Brief data presented in the Table 1 will be published in details elsewhere.
Acknowledgments
Agnieszka-Zylicz-Stachula is appreciated for valuable suggestions. Jakub Zaleski is appreciated for English corrections.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shapiro, J.W.; Putonti, C. Gene co-occurrence networks reflect bacteriophage ecology and evolution. mBio 2018, 9, e01870-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skowron, P.M.; Kropinski, A.M.; Zebrowska, J.; Janus, L.; Szemiako, K.; Czajkowska, E.; Maciejewska, N.; Skowron, M.; Łoś, J.; Łoś, M.; et al. Sequence, genome organization, annotation and proteomics of the thermophilic, 47.7-kb Geobacillus stearothermophilus bacteriophage TP-84 and its classification in the new Tp84virus genus. PLoS ONE 2018, 13, e0195449. [Google Scholar] [CrossRef]
- Zablocki, O.; van Zyl, L.; Trindade, M. Biogeography and taxonomic overview of terrestrial hot spring thermophilic bacteriophages. Extremophiles 2018, 22, 827–837. [Google Scholar] [CrossRef]
- Wisotzkey, J.D.; Jurtshuk, P., Jr.; Fox, G.E.; Deinhard, G.; Poralla, K. Comparative sequence analyses on the 16S rRNA (rDNA) of Bacillus acidocaldarius, Bacillus acidoterrestris, and Bacillus cycloheptanicus and proposal for creation of a new genus, Alicyclobacillus gen. nov. Int. J. Syst. Bacteriol. 1992, 42, 263–269. [Google Scholar] [CrossRef]
- Dufresne, S.; Bousquet, J.; Boissinot, M.; Guay, R. Sulfobacillus disulfidooxidans sp. nov., a new acidophilic, disulfide-oxidizing, gram-positive, spore-forming bacterium. Int. J. Syst. Bacteriol. 1996, 46, 1056–1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyndrickx, M.; Lebbe, L.; Vancanneyt, M.; Kersters, K.; De Vos, P.; Logan, N.A.; Forsyth, G.; Nazli, S.; Ali, N.; Berkeley, R.C.W. A polyphasic reassessment of the genus Aneurinibacillus, reclassification of Bacillus thermoaerophilus (Meier-Stauffer et al. 1996) as Aneurinibacillus thermoaeropilus comb. nov., and emended descriptions of A. aneurinilyticus corrig., A migulanus, and A. thermoaerophilus. Int. J. Syst. Bacteriol. 1997, 47, 808–817. [Google Scholar]
- Touzel, J.P.; O’Donohue, M.; Debeire, P.; Samain, E.; Breton, C. Thermobacillus xylanilyticus gen. nov., sp. nov., a new aerobic thermophilic xylan-degrading bacterium isolated from farm soil. Int. J. Syst. Evol. Microbiol. 2000, 50, 315–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rainey, F.A.; Fritze, D.; Stackebrandt, E. The phylogenetic diversity of thermophilic members of the genus Bacillus as revealed by 16S rDNA analysis. FEMS Microbiol. Lett. 1994, 115, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Nazina, T.N.; Tourova, T.P.; Poltaraus, A.B.; Novikova, E.V.; Grigoryan, A.A.; Ivanova, A.E.; Lysenko, A.M.; Petrunyaka, V.V.; Osipov, G.A.; Belyaev, S.S.; et al. Taxonomic study of aerobic thermophilic bacilli: Descriptions of Geobacillus subterraneus gen. nov., sp. nov. and Geobacillus uzenensis sp. nov. from petroleum reservoirs and transfer of Bacillus stearothermophilus, Bacillus thermocatenulatus, Bacillus thermoleovorans, Bacillus kaustophilus, Bacillus thermoglucosidasius and Bacillus thermodenitrificans to Geobacillus as the new combinations G. stearothermophilus, G. thermocatenulatus, G. thermoleovorans, G. kaustophilus, G. thermoglucosidasius and G. thermodenitrificans. Int. J. Syst. Evol. Microbiol. 2001, 51, 433–446. [Google Scholar] [CrossRef] [Green Version]
- McMullan, G.; Christie, J.M.; Rahman, T.J.; Banat, I.M.; Ternan, N.G.; Marchant, R. Habitat, applications and genomics of the aerobic, thermophilic genus Geobacillus. Biochem. Soc. Trans. 2004, 32, 214–217. [Google Scholar] [CrossRef] [Green Version]
- Abe, K.; Yoshinari, A.; Aoyagi, T.; Hirota, Y.; Iwamoto, K.; Sato, T. Regulated DNA rearrangement during sporulation in Bacillus weihenstephanensis KBAB4. Mol. Microbiol. 2013, 90, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Roger, W.; Hendrix, R.W. Bacteriophages: Evolution of the mMajority. Theor. Popul. Biol. 2002, 61, 471–480. [Google Scholar]
- Reysenbach, A.-L.; Shock, E. Merging genomes with geochemistry in hydrothermal ecosystems. Science 2002, 296, 1077–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhang, X. Genome analysis of deep-sea thermophilic bacteriophage D6E. Appl. Environ. Microbiol. 2010, 76, 7861–7866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharp, R.J.; Riley, P.W.; White, D. Thermophilic Bacilli; Kristjansson, J.K., Ed.; CRC Press: Boca Raton, FL, USA, 1992; pp. 19–50. [Google Scholar]
- Maugeri, T.L.; Gugliandolo, C.; Caccamo, D.; Stackebrandt, E. Three novel halotolerant and thermophilic Geobacillus strains from shallow marine vents. Syst. Appl. Microbiol. 2002, 25, 450–455. [Google Scholar] [CrossRef]
- Blanc, M.; Marilley, L.; Beffa, T.; Aragno, M. Rapid identification of heterotrophic, thermophilic, spore-form-ing bacteria isolated from hot composts. Int. J. Syst. Bacteriol. 1997, 47, 1246–1248. [Google Scholar] [CrossRef] [Green Version]
- Marks, T.J.; Hamilton, P.T. Characterization of a thermophilic bacteriophage of Geobacillus kaustophilus. Arch. Virol. 2014, 159, 2771–2775. [Google Scholar] [CrossRef]
- Bell, E.M. Life at Extremes: Environments, Organisms, and Strategies for Survival; CABI: Wallingford/Oxfordshire, UK, 2012. [Google Scholar]
- Pentecost, A. High temperature ecosystems and their chemical interactions with their environment. In Ciba Foundation Symposium 202-Evolution of Hydrothermal Ecosystems on Earth (And Mars?): Ciba Foundation Symposium 202; Bock, G.R., Goode, J.A., Eds.; John Wiley & Sons Ltd.: New York, NY, USA, 1996; pp. 99–111. [Google Scholar] [CrossRef]
- Sharp, R.J.; Ahmad, S.I.; Munster, A.; Dowsett, B.; Atkinson, T. The isolation and characterization of bacterio-bacteriophages infecting obligately thermophilic strains of Bacillus. J. Gen. Microbiol. 1986, 132, 1709–1722. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Wu, S.; Song, Q.; Zhang, X.; Xie, L. Two novel bacteriophages of thermophilic bacteria isolated from deep-sea hydrothermal fields. Curr. Microbiol. 2006, 53, 163–166. [Google Scholar] [CrossRef]
- Nagayoshi, Y.; Kumagae, K.; Mori, K.; Tashiro, K.; Nakamura, A.; Fujino, Y.; Hiromasa, Y.; Iwamoto, T.; Kuhara, S.; Ohshima, T.; et al. Physiological properties and genome structure of the hyperthermophilic filamentous bacteriophage φOH3 which infects Thermus thermophilus HB8. Front. Microbiol. 2016, 7, 50. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Zhang, X. Deep-sea thermophilic Geobacillus bacteriophage GVE2 transcriptional profile and proteomic characterization of virions. Appl. Microbiol. Biotechnol. 2008, 80, 697–707. [Google Scholar] [CrossRef]
- Liu, B.; Zhou, F.; Wu, S.; Xu, Y.; Zhang, X. Genomic and proteomic characterization of a thermophilic Geobacillus bacteriophage GBSV1. Res. Microbiol. 2009, 16, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Doi, K.; Mori, K.; Martono, H.; Nagayoshi, Y.; Fujino, Y.; Tashiro, K.; Kuhara, S.; Ohshima, T. Draft genome sequence of Geobacillus kaustophilus GBlys, a lysogenic strain with bacteriophage ϕOH2. Genome Announc. 2013, 1, e00634-13. [Google Scholar] [CrossRef] [Green Version]
- Morozova, V.; Bokovaya, O.; Kozlova, Y.; Kurilshikov, A.; Babkin, I.; Tupikin, A.; Bondar, A.; Ryabchikova, E.; Brayanskaya, A.; Peltek, S.; et al. A novel thermophilic Aeribacillus bacteriophage AP45 isolated from the Valley of Geysers, Kamchatka: Genome analysis suggests the existence of a new genus within the Siphoviridae family. Extremophiles 2019, 23, 599–612. [Google Scholar] [CrossRef]
- Wei, D.; Zhang, X. Identification and characterization of a single-stranded DNA-binding protein from thermophilic bacteriophage GVE2. Virus Genes 2008, 36, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Brister, J.R.; Ako-Adjei, D.; Bao, Y.; Blinkova, O. NCBI viral genomes resource. Nucleic Acids Res. 2015, 43, D571–D577. [Google Scholar] [CrossRef] [Green Version]
- Saunders, G.F.; Campbell, L.L.; Postgate, J.R. Abstract. In Proceedings of the 148th National Meeting of the American Chemical Society, Chicago, IL, USA, 30 August–4 September 1964. [Google Scholar]
- Koser, S.A. Action of the bacteriophage on a thermophilic Bacillus. Exp. Biol. Med. 1926, 24, 109–111. [Google Scholar] [CrossRef]
- Adant, M. Les bacteriophages des microbes thermophiles. Compt. Rend. Soc. Biol. 1928, 99, 1244–1245. [Google Scholar]
- White, R.; Georgi, C.E.; Militzer, W. Heat studies on a thermophilic bacteriophage. Proc. Soc. Exp. Biol. Med. 1954, 85, 137–139. [Google Scholar] [CrossRef]
- Marsh, C.L.; Larsen, D.H. Characterization of some thermophilic bacteria from the Hot Springs of Yellowstone National Park. J. Bacteriol. 1953, 65, 193–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, R.; Georgi, C.E.; Militzer, W.E. Characteristics of a thermophilic bacteriophage. Proc. Soc. Exp. Biol. Med. 1955, 88, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Hirano, J.; Mutoh, Y.; Kitamura, M.; Asai, S.; Nakajima, I.; Takamiya, A. Effect of change of temperature upon the viability of a thermophilic bacterium. J. Gen. Appl. Microbiol. 1958, 4, 188–199. [Google Scholar] [CrossRef] [Green Version]
- Onodera, N. Electron-Microscopy 1959, 7, 2–3, 19. (In Japanese)
- Onodera, N. On some characteristics of a newly isolated thermophilic bacteriophage and consideration of its thermostability. J. Electron. Microsc. (Tokyo) 1961, 10, 91–102. [Google Scholar] [CrossRef]
- Saunders, G.F.; Campbell, L.L. Characterization of a thermophilic bacteriophage for Bacillus stearothermophilus. J. Bacteriol. 1966, 91, 340–348. [Google Scholar] [CrossRef] [Green Version]
- Zeigler, D.R. The genus Geobacillus. In Bacillus Genetic Stock Center, Catalog of Strains, 7th ed.; Department of Biochemistry, The Ohio State University: Columbus, OH, USA, 2001; Volume 3, pp. 1–25. [Google Scholar]
- Epstein, I.; Campbell, L. Production and purification of the thermophilic bacteriophage TP-84. Appl. Microbiol. 1975, 29, 219–223. [Google Scholar] [CrossRef]
- Shafiai, F.; Thompson, T.L. Isolation and preliminary characterization of bacteriophage φμ-4. J. Bacteriol. 1964, 87, 999–1002. [Google Scholar] [CrossRef] [Green Version]
- Walker, E.; Campbell, L. Induction and properties of a temperate bacteriophage from Bacillus stearothermophilus. J. Bacteriol. 1965, 89, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Walker, E.; Campbell, L. Biochemical changes in lysogenic Bacillus stearothermophilus after bacteriophage induction. J. Bacteriol. 1965, 90, 1129–1137. [Google Scholar] [CrossRef] [Green Version]
- Welker, E. Transduction in Bacillus stearothermophilus. J. Bacteriol. 1988, 170, 3761–3764. [Google Scholar] [CrossRef] [Green Version]
- Carnevali, F.; Donelli, G. Some properties of a thermophilic phage DNA. Arch. Biochem. Biophys. 1968, 125, 376–377. [Google Scholar] [CrossRef]
- Egbert, L.N.; Mitchell, H.K. Characteristics of Tφ3, a bacteriophage for Bacillus stearothermophilus. J. Virol. 1967, 1, 610–616. [Google Scholar] [CrossRef] [Green Version]
- Egbert, L.N. Characteristics of the deoxyribonucleic acid of T phi 3, a bacteriophage for Bacillus stearothermophilus. J. Virol. 1969, 3, 528–532. [Google Scholar] [CrossRef] [Green Version]
- Humbert, R.D.; Fields, M.L. Study of two bacteriophages of Bacillus stearothermophilus strain NCA1518. J. Virol. 1972, 9, 397–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurgen, C.L.; Edero, M. The influence of inorganic ions on the heat stability of a thermophilic bacteriophage. Physiol. Plant. 1972, 27, 182–186. [Google Scholar]
- Reanney, D.C.; Marsh, S.C.N. The ecology of viruses attacking Bacillus stearothermophilus in soil. Soil Biol. Biochem. 1973, 5, 399–406. [Google Scholar] [CrossRef]
- Sakaki, Y.; Oshima, T. A new lipid-containing phage infecting acidophilic thermophilic bacteria. Virology 1976, 75, 256–259. [Google Scholar] [CrossRef]
- Sharp, R.J.; Bown, K.J.; Atkinson, A. Phenotypic and genotypic characterization of some thermophilic species of Bacillus. J. Gen. Microbiol. 1980, 117, 201–210. [Google Scholar] [CrossRef] [Green Version]
- Wolf, J.; Sharp, R.J. Taxonomic and related aspects of thermophiles within the Genus Bacillus. In The Aerobic Endospore-Forming Bacteria: Classification and Identification; Berkeley, R.C.W., Goodfellow, M., Eds.; Academic Press: London, UK, 1981; pp. 251–296. [Google Scholar]
- Sharp, R.J.; Woodrow, R. Numerical taxonomy of Bacillus thermophiles. In Abstracts of the XHIth International Congress of Microbiology Boston USA; Nottingham Trent University: Nottingham, UK, 1982; pp. 61–62. [Google Scholar]
- Wang, Y.; Zhang, X. Characterization of a novel portal protein from deep-sea thermophilic bacteriophage GVE2. Gene 2008, 421, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Ye, T.; Zhang, X. Roles of bacteriophage GVE2 endolysin in host lysis at high temperatures. Microbiology 2013, 159, 1597–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Wu, S.; Xie, L. Complete genome sequence and proteomic analysis of a thermophilic bacteriophage BV1. Acta Oceanol. Sin. 2010, 29, 84–89. [Google Scholar] [CrossRef]
- Fujimoto, D.F.; Higginbotham, R.H.; Sterba, K.M.; Maleki, S.J.; Segall, A.M.; Smeltzer, M.S.; Hurlburt, B.K. Staphylococcus aureus SarA is a regulatory protein responsive to redox and pH that can support bacteriophage lambda integrase-mediated excision/recombination. Mol. Microbiol. 2009, 74, 1445–1458. [Google Scholar] [CrossRef] [Green Version]
- Alonso, J.C.; Lüder, G.; Stiege, A.C.; Chai, S.; Weise, F.; Trautner, T.A. The complete nucleotide sequence and functional organization of Bacillus subtilis bacteriophage SPP1. Gene 1997, 204, 201–212. [Google Scholar] [CrossRef]
- Levine, M.; Truesdall, S.; Ramakrishan, T.; Bronson, M.J. Dual control of lysogeny by bacteriophage P22: An antirepressor locus and its controlling elements. J. Mol. Biol. 1975, 91, 421–438. [Google Scholar] [CrossRef] [Green Version]
- van Zyl, L.J.; Sunda, F.; Taylor, M.P.; Cowan, D.; Trindade, M.I. Identification and characterization of a novel Geobacillus thermoglucosidasius bacteriophage, GVE3. Arch. Virol. 2015, 160, 2269–2282. [Google Scholar] [CrossRef] [Green Version]
- van Zyl, L.J.; Taylor, M.P.; Trindade, M. Engineering resistance to bacteriophage GVE3 in Geobacillus thermoglucosidasius. Appl. Microbiol. Biotechnol. 2016, 100, 1833–1841. [Google Scholar] [CrossRef]
- Maszewska, A. Bacteriophage associated polysaccharide depolymerases—Characteristics and application. Postepy Hig. Med. Dosw. 2015, 69, 690–702. [Google Scholar] [CrossRef] [PubMed]
- Rocha, E.P.C.; Danchin, A. Base composition bias might result from competition for metabolic resources. TRENDS Genet. 2002, 18, 291–294. [Google Scholar] [CrossRef]
- Akhwale, J.K.; Rohde, M.; Rohde, C.; Bunk, B.; Sproer, C.; Klenk, H.-P.; Boga, H.I.; Wittmann, J. Comparative genomic analysis of eight novel haloalkaliphilic bacteriophages from Lake Elmenteita, Kenya. PLoS ONE 2019, 14, e0212102. [Google Scholar] [CrossRef]
- Mwirichia, R.; Cousin, S.; Muigai, A.W.; Boga, H.I.; Stackebrandt, E. Archaeal diversity in the haloalkaline Lake Elmenteita in Kenya. Curr. Microbiol. 2010, 60, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Streisinger, G.; Emrich, J.; Stahl, M.M. Chromosome structure in phage T4, iii. Terminal redundancy and length determination. Proc. Natl. Acad. Sci. USA 1967, 57, 292–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casjens, S. Prophages and bacterial genomics: What have we learned so far? Mol. Microbiol. 2003, 49, 277–300. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, J.; Dreiseikelmann, B.; Rohde, C.; Rohde, M.; Sikorski, J. Isolation and characterization of numerous novel phages targeting diverse strains of the ubiquitous and opportunistic pathogen Achromobacter xylosoxidans. PLoS ONE 2014, 9, e86935. [Google Scholar] [CrossRef] [PubMed]
- Sunderland, K.S.; Yang, M.; Mao, C. Phage-Enabled Nanomedicine: From Probes to Therapeutics in Precision Medicine. Angew Chem. Int. Ed. Engl. 2017, 56, 1964–1992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustard, M.T.; Whiting, S.; Cowan, D.A.; Wright, P.C. Biodegradation of high-concentration isopropanol by a solvent-tolerant thermophile, Bacillus pallidus. Extremophiles 2002, 6, 319–323. [Google Scholar] [CrossRef]
- Taylor, M.P.; Eley, K.L.; Martin, S.; Tuffin, M.I.; Burton, S.G.; Cowan, D.A. Thermophilic ethanologenesis: Future prospects for second-generation bioethanol production. Trends Biotechnol. 2009, 27, 398–405. [Google Scholar] [CrossRef]
- Cuebasg, M.; Sannino, D.; Bini, B. Isolation and characterization of an arsenic resistant Geobacillus kaustophilus strain from geothermal soils. J. Basic Microbiol. 2011, 51, 364–371. [Google Scholar] [CrossRef]
- Feng, L.; Wang, W.; Cheng, J.; Ren, Y.; Zhao, G.; Gao, C.; Tang, Y.; Lui, X.; Han, W.; Peng, X.; et al. Genome and proteome of long-chain alkane degrading Geobacillus thermodenitrificans NG80-2 isolated from a deep-subsurface oil reservoir. Proc. Natl. Acad. Sci. USA 2007, 104, 5602–5607. [Google Scholar] [CrossRef] [Green Version]
- Moriello, V.S.; Lama, L.; Poli, A.; Gugliandolo, C.; Maugeri, T.L.; Gamacorta, A.; Nicolaus, B. Production of exopolysaccharides from a thermophilic microorganism isolated from a marine hot spring in flegrean areas. J. Ind. Microbiol. Biotechnol. 2003, 30, 95–101. [Google Scholar] [CrossRef]
- Novotny, J.F.; Perry, J.J. Characterization of bacteriocins from two strains of Bacillus thermoleovorans, a thermophilic hydrocarbon-utilizing species. Appl. Environ. Microbiol. 1992, 58, 2393–2396. [Google Scholar] [CrossRef] [Green Version]
- Jeżewska-Frąckowiak, J.; Seroczyńska, K.; Banaszczyk, J.; Jedrzejczak, G.; Żylicz-Stachula, A.; Skowron, P.M. The promises and risks of probiotic Bacillus species. Acta Biochim. Pol. 2018, 65, 509–519. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Korza, G.; De Marco, A.M.; Kuipers, O.P.; Li, Y.Q.; Setlow, P. Properties of spores of Bacillus subtilis with or without a transposon that decreases spore germination and increases spore wet heat resistance. J. Appl. Microbiol. 2021. [Google Scholar] [CrossRef]
- Kuebutornye, F.K.A.; Abarike, E.D.; Lu, Y. A review on the application of Bacillus as probiotics in aquaculture. Fish Shellfish Immunol. 2019, 87, 820–828. [Google Scholar] [CrossRef]
- Mu, Y.; Cong, Y. Bacillus coagulans and its applications in medicine. Benef. Microbes 2019, 10, 679–688. [Google Scholar] [CrossRef]
- Logan, N.A. Bacillus and relatives in foodborne illness. J. Appl. Microbiol. 2012, 112, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.; Ali, Z.; Khan, M.; Bostan, N.; Naseem, S. The dawn of phage therapy. Rev. Med. Virol. 2019, 29, e2041. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, H.; Mulvenna, N.; Sanz-Hernandez, M.; Zhang, P.; Li, Y.; Ma, J.; Wang, Y.; Matthews, S.; Wigneshweraraj, S.; et al. A Bacteriophage DNA Mimic Protein Employs a Non-specific Strategy to Inhibit the Bacterial RNA Polymerase. Front. Microbiol. 2021, 12, 692512. [Google Scholar] [CrossRef] [PubMed]
- Mulvenna, N.; Hantke, I.; Burchell, L.; Nicod, S.; Bell, D.; Turgay, K.; Wigneshweraraj, S. Xenogeneic modulation of the ClpCP protease of Bacillus subtilis by a phage-encoded adaptor-like protein. J. Biol. Chem. 2019, 294, 17501–17511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil, J.F.; Mesa, V.; Estrada-Ortiz, N.; Lopez-Obando, M.; Gómez, A.; Plácido, J. Viruses in Extreme Environments, Current Overview, and Biotechnological Potential. Viruses 2021, 13, 81. [Google Scholar] [CrossRef] [PubMed]
- Maat, D.S.; Biggs, T.; Evans, C.; Van Bleijswijk, J.D.L.; Van Der Wel, N.N.; Dutilh, B.E.; Brussaard, C.P.D. Characterization and temperature dependence of arctic micromonas polaris viruses. Viruses 2017, 9, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hughes, K.A.; Sutherland, I.W.; Clark, J.; Jones, M.V. Bacteriophage and associated polysaccharide depolymerases—Novel tools for study of bacterial biofilms. J. Appl. Microbiol. 1998, 85, 583–590. [Google Scholar] [CrossRef]
- Hughes, K.A.; Sutherland, I.W.; Clark, J.; Jones, M.V. Biofilm susceptibility to bacteriophage attack: The role of bacteriophageborne polysaccharide depolymerase. Microbiology 1998, 144, 3039–3047. [Google Scholar] [CrossRef] [Green Version]
- Donlan, R.M. Preventing biofilms of clinically relevant organisms using bacteriophage. Trends Microbiol. 2009, 17, 66–72. [Google Scholar] [CrossRef]
- Parasion, S.; Kwiatek, M.; Gryko, R.; Mizak, L.; Malm, A. Bacteriophages as an alternative strategy for fighting biofilm development. Pol. J. Microbiol. 2014, 63, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Szekera, K.; Zhou, X.; Schwab, T.; Casanueva, A.; Cowan, D.; Mikhailopulo, I.A.; Neubauer, P. Comparative investigations on thermostable pyrimidine nucleoside phosphorylases from Geobacillus thermoglucosidasius and Thermus thermophiles. J. Mol. Cat. B Enzym. 2012, 84, 27–34. [Google Scholar] [CrossRef]
- Julien, B. Characterization of the integrase gene and attachment site for the Myxococcus xanthus bacteriophage Mx9. J. Bacteriol. 2003, 185, 6325–6330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piazzolla, D.; Calì, S.; Spoldi, E.; Forti, F.; Sala, C.; Magnoni, F.; Dehò, G.; Ghisotti, D. Expression of phage P4 integrase is regulated negatively by both Int and Vis. J. Gen. Virol. 2006, 87, 2423–2431. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of morphological/structural forms of the ‘Bacillus group’ thermophages. (A). Siphoviridae; (B). Myoviridae; (C). Podoviridae; (D). Sphaerolipoviridae, Tectiviridae, Corticoviridae.
Figure 1.
Schematic representation of morphological/structural forms of the ‘Bacillus group’ thermophages. (A). Siphoviridae; (B). Myoviridae; (C). Podoviridae; (D). Sphaerolipoviridae, Tectiviridae, Corticoviridae.


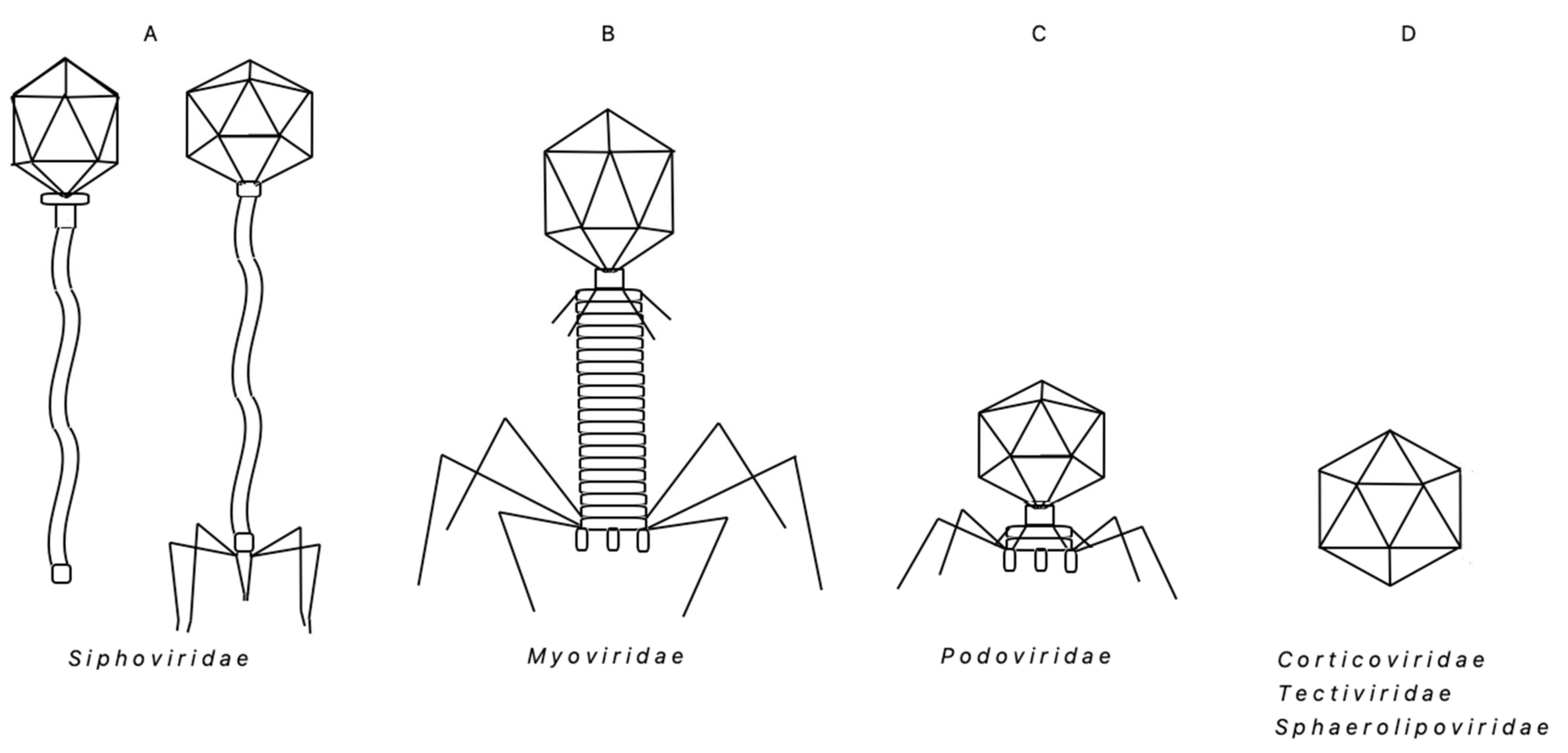{kind=link}
Table 1.
New (this work) experimentally confirmed functional CDSs of bacteriophage TP-84 genome—an update 1.
Table 1.
New (this work) experimentally confirmed functional CDSs of bacteriophage TP-84 genome—an update 1.
| CDS Name | CDS Length (bp) | Location in the Genome (bp) | CDS Arbitrary Orientation | Polypeptide Length (aa) | Predicted Polypeptide Molecular Weight (kDa) | Experimentally Determined Polypeptide Molecular Weight (kDa) | Predicted Isoelectric Point | Hypothetical Function (Analysis) | Confirmed by Proteomic Analysis |
|---|---|---|---|---|---|---|---|---|---|
| TP84_01 | 567 | 13–579 | + | 188 | 22.4 | 22.4 | 7.76 | terminase, small subunit | terminase, small subunit |
| TP84_02 | 1299 | 576–1874 | + | 432 | 50.0 | 50.1 | 7.74 | terminase, large subunit | terminase, large subunit |
| TP84_04 | 354 | 2586–2939 | + | 117 | 13.6 | ND | 4.36 | unknown | unknown |
| TP84_15 | 324 | 10358–10681 | + | 107 | 11.9 | ND | 6.26 | unknown | unknown |
| TP84_20 | 375 | 12687–13061 | + | 124 | 14.7 | 14.7 | 9.80 | unknown | unknown |
| TP84_26 | 2976 | 20179–23154 | + | 991 | 112.2 | 112.9 | 5.49 | glycosylase | glycosylase |
| TP84_27 | 432 | 23240–23671 | + | 143 | 15.5 | 15.5 | 6.20 | holin | holin |
| TP84_30 | 573 | 25329–25901 | + | 190 | 21.7 | 21.7 | 4.98 | unknown | unknown |
| TP84_33 | 291 | 26597–26887 | + | 96 | 11.0 | ND | 5.10 | unknown | unknown |
| TP84_41 | 273 | 28824–29096 | + | 90 | 10.6 | ND | 9.69 | unknown | unknown |
| TP84_43 | 309 | 29330–29639 | + | 102 | 11.0 | ND | 9.81 | unknown | unknown |
| TP84_48 | 2265 | 30527–32791 | + | 754 | 86 | ND | 7.09 | unknown | unknown |
| TP84_49 | 474 | 32871–33344 | + | 157 | 18.0 | ND | 5.85 | unknown | unknown |
| TP84_58 | 318 | 37942–38259 | + | 105 | 12.3 | ND | 4.92 | replicative helicase inhibitor | replicative helicase inhibitor |
| TP84_59 | 1305 | 38259–39563 | + | 434 | 49.1 | 49.2 | 5.53 | replicative DNA helicase | replicative DNA helicase |
| TP84_60 | 672 | 39632–40303 | + | 223 | 26.4 | 26.4 | 8.41 | unknown | unknown |
| TP84_61 | 435 | 40281–40715 | + | 144 | 16.7 | ND | 9.08 | HNH homing endonuclease | HNH homing endonuclease |
| TP84_70 | 330 | 43704–44033 | + | 109 | 12.7 | ND | 9.47 | unknown | unknown |
| TP84_73 | 264 | 44746–45009 | + | 87 | 10.4 | ND | 5.60 | unknown | unknown |
Table 2.
Classification and major data describing ‘Bacillus group’ thermophilic bacteriophages (chronologically as discovered).
Table 2.
Classification and major data describing ‘Bacillus group’ thermophilic bacteriophages (chronologically as discovered).
| N° | Bacteriophage Species | Virus Family, Genus | GenBank Accession Number | Host (Used for Propagation) | Genome | Original Discovery Reference | Isolate Location | Life Cycle | Growth Temperature (°C, Optimal/Range) and pH (Optimal/Range) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Type and Size [bp] | ORFs | G+C [%] | |||||||||
| 1 | ‘thermophilic lytic principle’ | ND | ND | B. stearothermophilus T60 | ND | ND | ND | Koser, 1926 | sewage polluted river water (USA) | lytic | 52–60, app. 7 |
| 2 | ‘thermophilic bacteriophage’ | ND | ND | ND | ND | ND | ND | Adant, 1928 | ND | lytic | 52–55 |
| 3 | ‘thermophilic bacteriophage’ | ND | ND | thermophilic bacterium no. 10 | ND | ND | ND | White et al., 1954, 1955 | greenhouse soil (USA) | lytic | 65 (50–70), 7 |
| 4 | ‘thermophilic bacteriophage’ | ND | ND | Bacillus sp. | DNA, RNA(?) | ND | ND | Onodera; 1959; Onodera 1961 | compost (Japan) | lytic | 65 (55–70), 7.2 |
| 5 | TP-84 | Siphoviridae, Tp84virus | KY565347.1 | B. stearothermophilus strain 10 | circular, dsDNA, 47,718 | 81 | 54.5 | Saunders & Campbell, 1964 | greenhouse soil (USA) | lytic | 58 (43–76), 7.2 |
| 6 | φμ-4 | ND | ND | B. stearothermophilus | ND | ND | ND | Shafia & Thompson, 1964 | ND | lytic/lysogenic | 50–65, 7 |
| 7 | TP-1 | Siphoviridae (putative) | ND | B. stearothermophilus | dsDNA, app. 18,516 bp, (MW 12.1 Mda) | ND | 42 | Welker and Campbell, 1965 | ND | lysogenic/lytic | 55 (50–65), 7 |
| 8 | ST1 | Siphoviridae Myoviridae (putative) | ND | B. stearothermophilus strain S13 | dsDNA | ND | 43 | Carnevali & Donelli, 1968 | ND | lytic | 60, app. 7 |
| 9 | Tφ3 | Siphoviridae | ND | B. stearothermophilus ATCC 8005 SR | ds DNA, app. 35,700 (MW app. 23.2 MDa) | ND | 40.2 | Egbert&Mitchel, 1967; Egbert, 1969 | soil (USA) | lytic | 60, 7.3 |
| 10 | GH5 | ND | ND | B. stearothermophilus NCA1518 | ND | ND | ND | Humbert & Fields, 1972 | greenhouse soil (USA) | lytic | 42.5–67, app. 7 |
| 11 | GH8 | Siphoviridae | ND | B.stearothermophilus NCA1518 | ND | ND | ND | Humbert & Fields, 1972 | greenhouse soli (USA) | lytic | 42.5–67, app. 7 |
| 12 | PhB1 | Siphoviridae | ND | Bacillus sp. strain B | ND | ND | ND | Junger & Edebo, 1972 | farm soil (Sweden) | lytic | 55, 7.3 |
| 13 | D5 | ND | ND | B. stearothermophilus NRS T91, ATCC7953 | ND | ND | ND | Reanney & Marsch, 1973 | ND | lytic | 45 (30–55), app. 7 |
| 14 | D6 | ND | ND | B. stearothermophilus NRS T91, ATCC7953 | ND | ND | ND | Reanney & Marsch, 1973 | ND | lytic | 45 (30–55), app. 7 |
| 15 | D7 | ND | ND | B. stearothermophilus NRS T91, ATCC7953 | ND | ND | ND | Reanney & Marsch, 1973 | ND | lytic | 45 (30–55), app. 7 |
| 16 | D8 | ND | ND | B. stearothermophilus NRS T91, ATCC7953 | ND | ND | ND | Reanney & Marsch, 1973 | ND | lytic | 45 (30–55), app. 7 |
| 17 | φNS11 | Sphaerolipoviridae, (putative) | ND | B.acidocaldarius TA6 | dsDNA | ND | ND | Sakaki & Oshima, 1976 | hot spring (Beppu, Japan) | lytic | 60, 3.5 (2–5) |
| 18 | JS001 | ND | ND | B. caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | ND | lytic/lysogenic | 55 (50–70), 7.3 ± 0.2 |
| 19 | JS004 | ND | ND | Bacillus thermophile RS 239 | dsDNA | ND | ND | Sharp et al., 1986 | silage | lytic | 55 (50–70), 7.3 ± 0.2 |
| 20 | JS005 | ND | ND | B. thermophile RS 239 | dsDNA | ND | ND | Sharp et al., 1986 | rotting straw | lytic | 55 (50–70), 7.3 ± 0.2 |
| 21 | JS006 | ND | ND | Bacillus thermophile RS 239 | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 22 | JS007 | ND | ND | Bacillus thermophile RS 240 | dsDNA | ND | ND | Sharp et al., 1986 | silage | lytic | 55 (50–70), 7.3 ± 0.2 |
| 23 | JS008 | ND | ND | Bacillus thermophile RS 241 | dsDNA | ND | ND | Sharp et al., 1986 | rotting straw | lytic | 55 (50–70), 7.3 ± 0.2 |
| 24 | JS009 | ND | ND | Bacillus thermophile RS 242 | dsDNA | ND | ND | Sharp et al., 1986 | stable manure | lytic | 55 (50–70), 7.3 ± 0.2 |
| 25 | JS010 | ND | ND | Bacillus thermophile RS 242 | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 26 | JS011 | unclassified family | ND | Bacillus thermophile RS 239 | dsDNA | ND | ND | Sharp et al., 1986 | silage | lytic | 55 (50–70), 7.3 ± 0.2 |
| 27 | JS012 | ND | ND | Bacillus thermophile RS 239 | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 28 | JS013 | ND | ND | B.stearothermophilus NCA 1503 | dsDNA | ND | ND | Sharp et al., 1986 | soil | lytic | 55 (50–70), 7.3 ± 0.2 |
| 29 | JS014 | ND | ND | B.stearothermophilus NCA 1503 | dsDNA | ND | ND | Sharp et al., 1986 | rotting straw | lytic | 55 (50–70), 7.3 ± 0.2 |
| 30 | JS015 | ND | ND | B.stearothermophilus NCA 1503 | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 31 | JS017 | ND | ND | B. caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 32 | JS018 | ND | ND | B. caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | rotting vegetation | lytic | 55 (50–70), 7.3 ± 0.2 |
| 33 | JS019 | ND | ND | B. caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | rotting vegetation | lytic | 55 (50–70), 7.3 ± 0.2 |
| 34 | JS020 | ND | ND | B.s caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | rotting vegetation | lytic | 55 (50–70), 7.3 ± 0.2 |
| 35 | JS021 | ND | ND | B. caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | rotting vegetation | lytic | 55 (50–70), 7.3 ± 0.2 |
| 36 | JS022 | ND | ND | B. caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 37 | JS023 | ND | ND | B. caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 38 | JS024 | ND | ND | B. caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 39 | JS025 | ND | ND | B. caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 40 | JS026 | ND | ND | B. caldotenax | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 41 | JS027 | ND | ND | Bacillus thermophile RS 241 | dsDNA | ND | ND | Sharp et al., 1986 | compost | lytic | 55 (50–70), 7.3 ± 0.2 |
| 42 | BVW1 (W1) | Siphoviridae | ND | Bacillus sp. w13 | dsDNA, app. 18 kb | ND | ND | Liu et al., 2006 | deep-sea hydrothermal fields (West Pacific) | lytic | 60, 7.0 |
| 43 | GVE1 (E1) | Siphoviridae | ND | Geobacillus sp. E 26323 | dsDNA, app. 41 kb | ND | ND | Liu et al., 2006 | deep-sea hydrothermal fields (East Pacific) | lytic | 60, 7.0 |
| 44 | GVE2 (E2) | Siphoviridae, unclassified Siphoviridae | NC_009552 DQ453159 | Geobacillus sp. E 263 | linear, dsDNA, 40,863 | 62 | 44.8 | Liu & Zhang, 2008a,b | deep sea (China) | lysogenic | 65, 7.0 |
| 45 | GBSV1 | Myoviridae, Svunavirus | Geobacillus sp. 6k512 | linear, dsDNA, 34,683 | 54 | 44.4 | Liu et al., 2009, 2010 | off shore hot spring, (Xiamen, China) | lytic | 65, 7.2 | |
| 46 | BV1 | Myoviridae ,Svunavirus | NC_009737.2, DQ840344 | Geobacillus sp. 6k512 | linear, dsDNA, 35,055 | 54 | 44.4 | Liu et al., 2009, 2010 | off shore hot spring, (Xiamen, China) | lytic | 65, 7.2 |
| 47 | D6E | Myoviridae | NC_019544 | Geobacillus sp. E 26323 | circular, dsDNA, 49,335 | 49 | 46 | Wang & Zhang, 2010 | deep-sea hydrothermal fields (East Pacific) | lytic | 65, 7.0 |
| 48 | ϕOH2 (phiOH2) | Siphoviridae | AB823818, NC_021784 | G. kaustophilus GBlys, G. kaustophilus NBRC 102445(T), lysogenic G. kaustophilus GBlys) | dsDNA, 38,099 | 60 | 45 | Doi et al., 2013 | hot spring sediment (Japan) | lytic/lysogenic | 55 |
| 49 | GBK2 | Siphoviridae | KJ159566 | G. kaustophilus | Circularly permuted, dsDNA, 39,078 | 62 | 43 | Marks & Hamilton, 2014 | compost (Cary, NC, USA) | lytic | 55, 7.3 |
| 50 | GVE3 (E3) | Siphoviridae | NC_029073, KP144388 | G. thermoglucosidasius | dsDNA 141,298 | 202 | 29.6 | Van Zyl et al., 2015 | ND | lytic/lysogenic | 60, 7.3 |
| 51 | AP45 | Siphoviridae | KX965989 | Aeribacillus sp. CEMTC656 | dsDNA 51,606 | 71 | 38.3 | Morozowa et al., 2019 | soil (Valley of Geysers, Kamchatka, Russia) | lytic/lysogenic | 55, 7.5 |
| 52 | vB_Bps-36 | ND | MH884513 | B. pseudalcaliphilus | dsDNA 50,485 | 68 | 41.1 | Akhwale et al., 2019 | Lake Elmenteita, (Kenya) | lytic/? | 30–40< 9< |
| 53 | vB_BpsM-61 | ND | MH884514 | B. pseudofirmus | dsDNA 48,160 | 75 | 43.5 | Akhwale et al., 2019 | Lake Elmenteita, (Kenya) | lytic/? | 30–40< 9< |
| 54 | vB_BboS-125 | ND | MH884509 | B. bogoriensis | dsDNA 58,528 | 81 | 48.6 | Akhwale et al., 2019 | Lake Elmenteita, (Kenya) | lytic/? | 30–40< 9< |
| 55 | vB_BcoS-136 | ND | MH884508 | B. cohnii | dsDNA 160,590 | 240 | 32.2 | Akhwale et al., 2019 | Lake Elmenteita, (Kenya) | lytic/? | 30–40< 9< |
| 56 | vB_BpsS-140 | ND | MH884512 | B. pseudalcaliphilus | dsDNA 55,091 | 68 | 39.8 | Akhwale et al., 2019 | Lake Elmenteita, (Kenya) | lytic/? | 30–40< 9< |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Łubkowska, B.; Jeżewska-Frąckowiak, J.; Sobolewski, I.; Skowron, P.M. Bacteriophages of Thermophilic ‘Bacillus Group’ Bacteria—A Review. Microorganisms 2021, 9, 1522. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071522
AMA Style
Łubkowska B, Jeżewska-Frąckowiak J, Sobolewski I, Skowron PM. Bacteriophages of Thermophilic ‘Bacillus Group’ Bacteria—A Review. Microorganisms. 2021; 9(7):1522. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071522
Chicago/Turabian StyleŁubkowska, Beata, Joanna Jeżewska-Frąckowiak, Ireneusz Sobolewski, and Piotr M. Skowron. 2021. "Bacteriophages of Thermophilic ‘Bacillus Group’ Bacteria—A Review" Microorganisms 9, no. 7: 1522. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071522
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.