
Cross-Reactive Immune Responses toward the Common Cold Human Coronaviruses and Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): Mini-Review and a Murine Study

Abstract
:1. Severe Acute Respiratory Syndrome Coronavirus 2
2. Common Cold Human Coronaviruses (HCoV)
3. Human Responses toward SARS-CoV-2 in SARS-CoV-2 Unexposed Individuals
4. Upregulation of Cross-Reactive Antibodies after SARS-CoV-2 Infections
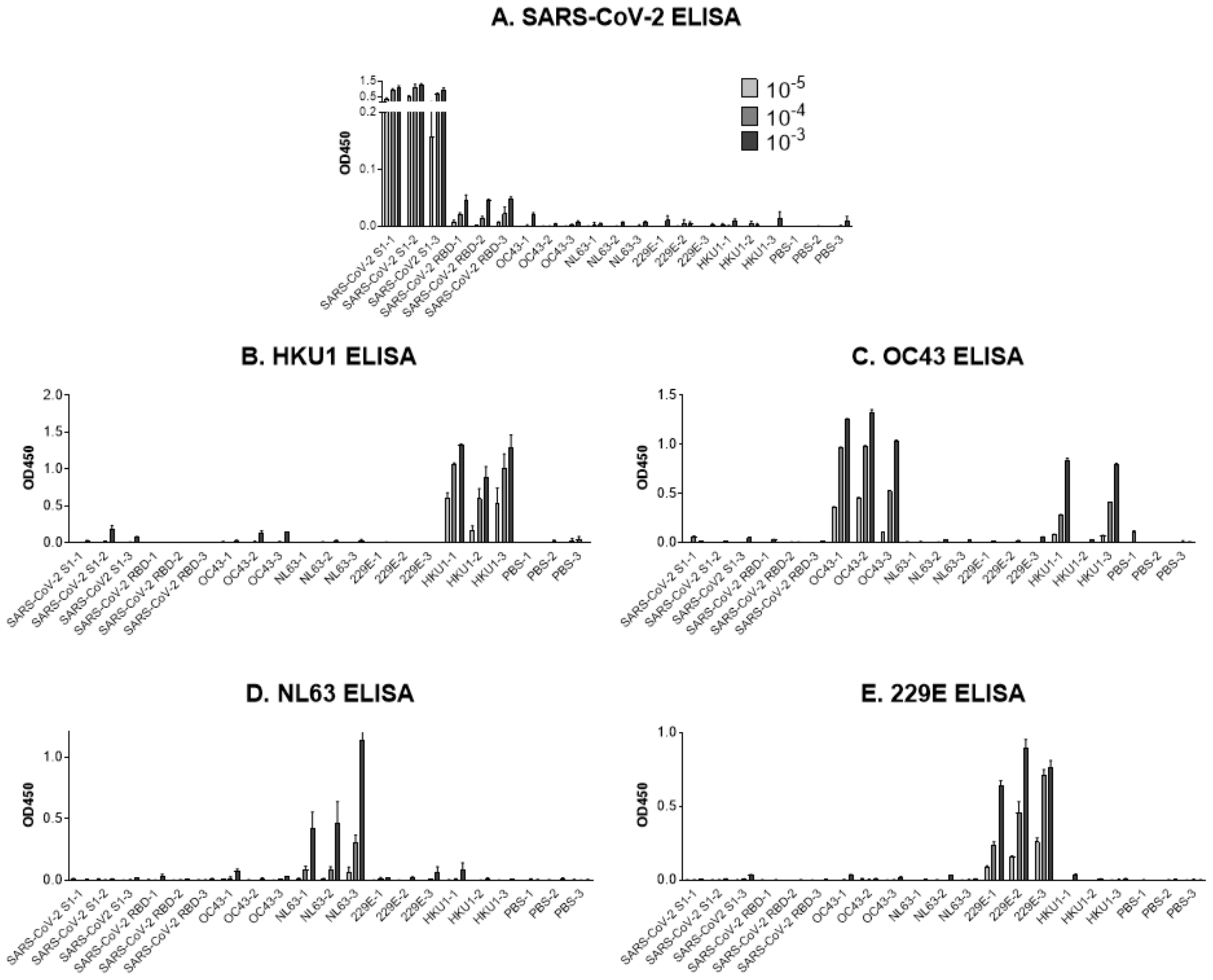
5. Testing Cross-Reactive S1 Antibody Induction in a Controlled Research Setting
6. T Cells Cross-React with Common Cold HCoVs and SARS-CoV-2
7. Influence of Cross-Reactive Immune Responses on SARS-CoV-2 Disease in Humans
8. Why Has the Study of Cross-Reactive Immune Responses Yielded Conflicting Messages?
9. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| SARS-CoV-2 | severe acute respiratory syndrome coronavirus 2; |
| HCoV | human coronavirus; |
| MIS-C | multisystem inflammatory syndrome in children; |
| RBD | receptor binding domain; |
| ACE2 | angiotensin-converting enzyme 2 |
References
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Johns Hopkins University and Medicine. Coronavirus Resource Center. Available online: https://coronavirus.jhu.edu (accessed on 6 June 2021).
- Xiao, F.; Tang, M.; Zheng, X.; Liu, Y.; Li, X.; Shan, H. Evidence for Gastrointestinal Infection of SARS-CoV-2. Gastroenterology 2020, 158, 1831–1833.e3. [Google Scholar] [CrossRef]
- Coronavirus Disease 2019. COVID-19 in Children and Teens, Updated 18 December 2020. Available online: www.cdc.gov (accessed on 25 February 2021).
- Guruprasad, L. Human coronavirus spike protein-host receptor recognition. Prog. Biophys. Mol. Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, A.A.T.; Fatima, K.; Mohammad, T.; Fatima, U.; Singh, I.K.; Singh, A.; Atif, S.M.; Hariprasad, G.; Hasan, G.M.; Hassan, M.I. Insights into SARS-CoV-2 genome, structure, evolution, pathogenesis and therapies: Structural genomics approach. Biochim Biophys. Acta Mol. Basis Dis. 2020, 1866, 165878. [Google Scholar] [CrossRef] [PubMed]
- Mandala, V.S.; McKay, M.J.; Shcherbakov, A.A.; Dregni, A.J.; Kolocouris, A.; Hong, M. Structure and drug binding of the SARS-CoV-2 envelope protein transmembrane domain in lipid bilayers. Nat. Struct. Mol. Biol. 2020, 27, 1202–1208. [Google Scholar] [CrossRef]
- Weiss, S.R.; Navas-Martin, S. Coronavirus pathogenesis and the emerging pathogen severe acute respiratory syndrome coronavirus. Microbiol. Mol. Biol. Rev. 2005, 69, 635–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widge, A.T.; Rouphael, N.G.; Jackson, L.A.; Anderson, E.J.; Roberts, P.C.; Makhene, M.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; Pruijssers, A.J.; et al. Durability of Responses after SARS-CoV-2 mRNA-1273 Vaccination. N. Engl. J. Med. 2021, 384, 80–82. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the mRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2020. [Google Scholar] [CrossRef]
- Walsh, E.E.; Frenck, R.W., Jr.; Falsey, A.R.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Neuzil, K.; Mulligan, M.J.; Bailey, R.; et al. Safety and Immunogenicity of Two RNA-Based Covid-19 Vaccine Candidates. N. Engl. J. Med. 2020. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Perez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Rawat, K.; Kumari, P.; Saha, L. COVID-19 vaccine: A recent update in pipeline vaccines, their design and development strategies. Eur. J. Pharm. 2021, 892, 173751. [Google Scholar] [CrossRef]
- Walensky, R.P.; Walke, H.T.; Fauci, A.S. SARS-CoV-2 Variants of Concern in the United States-Challenges and Opportunities. JAMA 2021, 325, 1037–1038. [Google Scholar] [CrossRef]
- Monto, A.S.; DeJonge, P.M.; Callear, A.P.; Bazzi, L.A.; Capriola, S.B.; Malosh, R.E.; Martin, E.T.; Petrie, J.G. Coronavirus Occurrence and Transmission Over 8 Years in the HIVE Cohort of Households in Michigan. J. Infect. Dis. 2020, 222, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Liu, J.; Ma, Z.; Bramer, W.M.; Peppelenbosch, M.P.; Pan, Q. Estimating Global Epidemiology of Low-Pathogenic Human Coronaviruses in Relation to the COVID-19 Context. J. Infect. Dis. 2020, 222, 695–696. [Google Scholar] [CrossRef] [PubMed]
- Gorse, G.J.; Patel, G.B.; Vitale, J.N.; O’Connor, T.Z. Prevalence of antibodies to four human coronaviruses is lower in nasal secretions than in serum. Clin. Vaccine Immunol. 2010, 17, 1875–1880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Hoek, L. Human coronaviruses: What do they cause? Antivir. Ther. 2007, 12, 651–658. [Google Scholar]
- Falsey, A.R.; Walsh, E.E.; Hayden, F.G. Rhinovirus and coronavirus infection-associated hospitalizations among older adults. J. Infect. Dis. 2002, 185, 1338–1341. [Google Scholar] [CrossRef] [Green Version]
- Kronvall, G.; Nordenfelt, E. On the history of human coronaviruses. APMIS 2021. [Google Scholar] [CrossRef]
- Hamre, D.; Procknow, J.J. A new virus isolated from the human respiratory tract. Proc. Soc. Exp. Biol. Med. 1966, 121, 190–193. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, H.; Pyrc, K.; van der Hoek, L.; Geier, M.; Berkhout, B.; Pohlmann, S. Human coronavirus NL63 employs the severe acute respiratory syndrome coronavirus receptor for cellular entry. Proc. Natl. Acad. Sci. USA 2005, 102, 7988–7993. [Google Scholar] [CrossRef] [Green Version]
- van der Hoek, L.; Pyrc, K.; Berkhout, B. Human coronavirus NL63, a new respiratory virus. FEMS Microbiol. Rev. 2006, 30, 760–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Hoek, L.; Pyrc, K.; Jebbink, M.F.; Vermeulen-Oost, W.; Berkhout, R.J.; Wolthers, K.C.; Wertheim-van Dillen, P.M.; Kaandorp, J.; Spaargaren, J.; Berkhout, B. Identification of a new human coronavirus. Nat. Med. 2004, 10, 368–373. [Google Scholar] [CrossRef]
- Hoorn, B.; Tyrrell, D.A. On the Growth of Certain “Newer” Respiratory Viruses in Organ Cultures. Br. J. Exp. Pathol. 1965, 46, 109–118. [Google Scholar]
- Tyrrell, D.A.; Bynoe, M.L. Cultivation of a Novel Type of Common-Cold Virus in Organ Cultures. Br. Med. J. 1965, 1, 1467–1470. [Google Scholar] [CrossRef] [Green Version]
- Tyrrell, D.A.; Almeida, J.D. Direct electron-microscopy of organ culture for the detection and characterization of viruses. Arch. Gesamte Virusforsch 1967, 22, 417–425. [Google Scholar] [CrossRef]
- Almeida, J.D.; Tyrrell, D.A. The morphology of three previously uncharacterized human respiratory viruses that grow in organ culture. J. Gen. Virol. 1967, 1, 175–178. [Google Scholar] [CrossRef]
- Hoorn, B.; Tyrrell, D.A. A new virus cultivated only in organ cultures of human ciliated epithelium. Arch. Gesamte Virusforsch 1966, 18, 210–225. [Google Scholar] [CrossRef] [PubMed]
- Hoorn, B.; Tyrrell, D.A. Organ cultures in virology. Prog. Med. Virol 1969, 11, 408–450. [Google Scholar] [PubMed]
- Bruckova, M.; McIntosh, K.; Kapikian, A.Z.; Chanock, R.M. The adaptation of two human coronavirus strains (OC38 and OC43) to growth in cell monolayers. Proc. Soc. Exp. Biol. Med. 1970, 135, 431–435. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, K.; Bruckova, M.; Kapikian, A.Z.; Chanock, R.J.; Turner, H. Studies on new virus isolates recovered in tracheal organ culture. Ann. N. Y. Acad. Sci. 1970, 174, 983–989. [Google Scholar] [CrossRef]
- McIntosh, K.; Dees, J.H.; Becker, W.B.; Kapikian, A.Z.; Chanock, R.M. Recovery in tracheal organ cultures of novel viruses from patients with respiratory disease. Proc. Natl. Acad. Sci. USA 1967, 57, 933–940. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, K.; Becker, W.B.; Chanock, R.M. Growth in suckling-mouse brain of “IBV-like” viruses from patients with upper respiratory tract disease. Proc. Natl. Acad. Sci. USA 1967, 58, 2268–2273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyrc, K.; Berkhout, B.; van der Hoek, L. Identification of new human coronaviruses. Expert Rev. Anti Infect. Ther. 2007, 5, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Lau, S.K.; Woo, P.C.; Yip, C.C.; Tse, H.; Tsoi, H.W.; Cheng, V.C.; Lee, P.; Tang, B.S.; Cheung, C.H.; Lee, R.A.; et al. Coronavirus HKU1 and other coronavirus infections in Hong Kong. J. Clin. Microbiol. 2006, 44, 2063–2071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, P.C.; Lau, S.K.; Chu, C.M.; Chan, K.H.; Tsoi, H.W.; Huang, Y.; Wong, B.H.; Poon, R.W.; Cai, J.J.; Luk, W.K.; et al. Characterization and complete genome sequence of a novel coronavirus, coronavirus HKU1, from patients with pneumonia. J. Virol. 2005, 79, 884–895. [Google Scholar] [CrossRef] [Green Version]
- Pyrc, K.; Berkhout, B.; van der Hoek, L. The novel human coronaviruses NL63 and HKU1. J. Virol. 2007, 81, 3051–3057. [Google Scholar] [CrossRef] [Green Version]
- Gussow, A.B.; Auslander, N.; Faure, G.; Wolf, Y.I.; Zhang, F.; Koonin, E.V. Genomic determinants of pathogenicity in SARS-CoV-2 and other human coronaviruses. Proc. Natl. Acad. Sci. USA 2020, 117, 15193–15199. [Google Scholar] [CrossRef] [PubMed]
- Hamre, D.; Kindig, D.A.; Mann, J. Growth and intracellular development of a new respiratory virus. J. Virol. 1967, 1, 810–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fouchier, R.A.; Hartwig, N.G.; Bestebroer, T.M.; Niemeyer, B.; de Jong, J.C.; Simon, J.H.; Osterhaus, A.D. A previously undescribed coronavirus associated with respiratory disease in humans. Proc. Natl. Acad. Sci. USA 2004, 101, 6212–6216. [Google Scholar] [CrossRef] [Green Version]
- Corman, V.M.; Baldwin, H.J.; Tateno, A.F.; Zerbinati, R.M.; Annan, A.; Owusu, M.; Nkrumah, E.E.; Maganga, G.D.; Oppong, S.; Adu-Sarkodie, Y.; et al. Evidence for an Ancestral Association of Human Coronavirus 229E with Bats. J. Virol. 2015, 89, 11858–11870. [Google Scholar] [CrossRef] [Green Version]
- Vijgen, L.; Keyaerts, E.; Moes, E.; Thoelen, I.; Wollants, E.; Lemey, P.; Vandamme, A.M.; Van Ranst, M. Complete genomic sequence of human coronavirus OC43: Molecular clock analysis suggests a relatively recent zoonotic coronavirus transmission event. J. Virol. 2005, 79, 1595–1604. [Google Scholar] [CrossRef] [Green Version]
- Cueno, M.E.; Imai, K. Structural Comparison of the SARS CoV 2 Spike Protein Relative to Other Human-Infecting Coronaviruses. Front. Med. 2020, 7, 594439. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.M.; Goodwin, E.C.; Verma, A.; Arevalo, C.P.; Bolton, M.J.; Weirick, M.E.; Gouma, S.; McAllister, C.M.; Christensen, S.R.; Weaver, J.; et al. Seasonal human coronavirus antibodies are boosted upon SARS-CoV-2 infection but not associated with protection. medRxiv 2020. [Google Scholar] [CrossRef]
- Khan, S.; Nakajima, R.; Jain, A.; de Assis, R.R.; Jasinskas, A.; Obiero, J.M.; Adenaiye, O.; Tai, S.; Hong, F.; Milton, D.K.; et al. Analysis of Serologic Cross-Reactivity Between Common Human Coronaviruses and SARS-CoV-2 Using Coronavirus Antigen Microarray. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Doshi, P. Covid-19: Do many people have pre-existing immunity? BMJ 2020, 370, m3563. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.W.; Faulkner, N.; Cornish, G.H.; Rosa, A.; Harvey, R.; Hussain, S.; Ulferts, R.; Earl, C.; Wrobel, A.G.; Benton, D.J.; et al. Preexisting and de novo humoral immunity to SARS-CoV-2 in humans. Science 2020, 370, 1339–1343. [Google Scholar] [CrossRef]
- Long, Q.X.; Liu, B.Z.; Deng, H.J.; Wu, G.C.; Deng, K.; Chen, Y.K.; Liao, P.; Qiu, J.F.; Lin, Y.; Cai, X.F.; et al. Antibody responses to SARS-CoV-2 in patients with COVID-19. Nat. Med. 2020, 26, 845–848. [Google Scholar] [CrossRef]
- Shiakolas, A.R.; Kramer, K.J.; Wrapp, D.; Richardson, S.I.; Schafer, A.; Wall, S.; Wang, N.; Janowska, K.; Pilewski, K.A.; Venkat, R.; et al. Cross-reactive coronavirus antibodies with diverse epitope specificities and extra-neutralization functions. bioRxiv 2020. [Google Scholar] [CrossRef]
- Wec, A.Z.; Wrapp, D.; Herbert, A.S.; Maurer, D.P.; Haslwanter, D.; Sakharkar, M.; Jangra, R.K.; Dieterle, M.E.; Lilov, A.; Huang, D.; et al. Broad neutralization of SARS-related viruses by human monoclonal antibodies. Science 2020, 369, 731–736. [Google Scholar] [CrossRef]
- Laing, E.D.; Sterling, S.L.; Richard, S.A.; Phogat, S.; Samuels, E.C.; Epsi, N.J.; Yan, L.; Moreno, N.; Coles, C.; Mehalko, J.; et al. A betacoronavirus multiplex microsphere immunoassay detects early SARS-CoV-2 seroconversion and controls for pre-existing seasonal human coronavirus antibody cross-reactivity. medRxiv 2020. [Google Scholar] [CrossRef]
- Huang, A.T.; Garcia-Carreras, B.; Hitchings, M.D.T.; Yang, B.; Katzelnick, L.C.; Rattigan, S.M.; Borgert, B.A.; Moreno, C.A.; Solomon, B.D.; Trimmer-Smith, L.; et al. A systematic review of antibody mediated immunity to coronaviruses: Kinetics, correlates of protection, and association with severity. Nat. Commun. 2020, 11, 4704. [Google Scholar] [CrossRef]
- Song, G.; He, W.T.; Callaghan, S.; Anzanello, F.; Huang, D.; Ricketts, J.; Torres, J.L.; Beutler, N.; Peng, L.; Vargas, S.; et al. Cross-reactive serum and memory B-cell responses to spike protein in SARS-CoV-2 and endemic coronavirus infection. Nat. Commun. 2021, 12, 2938. [Google Scholar] [CrossRef]
- Guo, L.; Ren, L.; Yang, S.; Xiao, M.; Chang, D.; Yang, F.; Dela Cruz, C.S.; Wang, Y.; Wu, C.; Xiao, Y.; et al. Profiling Early Humoral Response to Diagnose Novel Coronavirus Disease (COVID-19). Clin. Infect. Dis. 2020, 71, 778–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mveang Nzoghe, A.; Essone, P.N.; Leboueny, M.; Maloupazoa Siawaya, A.C.; Bongho, E.C.; Mvoundza Ndjindji, O.; Avome Houechenou, R.M.; Agnandji, S.T.; Djoba Siawaya, J.F. Evidence and implications of pre-existing humoral cross-reactive immunity to SARS-CoV-2. Immun. Inflamm. Dis. 2021, 9, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Li, P.; Ji, Y.; Ikram, A.; Pan, Q. Cross-reactivity towards SARS-CoV-2: The potential role of low-pathogenic human coronaviruses. Lancet Microbe 2020, 1, e151. [Google Scholar] [CrossRef]
- To, K.K.; Cheng, V.C.; Cai, J.P.; Chan, K.H.; Chen, L.L.; Wong, L.H.; Choi, C.Y.; Fong, C.H.; Ng, A.C.; Lu, L.; et al. Seroprevalence of SARS-CoV-2 in Hong Kong and in residents evacuated from Hubei province, China: A multicohort study. Lancet Microbe 2020, 1, e111–e118. [Google Scholar] [CrossRef]
- Shrock, E.; Fujimura, E.; Kula, T.; Timms, R.T.; Lee, I.H.; Leng, Y.; Robinson, M.L.; Sie, B.M.; Li, M.Z.; Chen, Y.; et al. Viral epitope profiling of COVID-19 patients reveals cross-reactivity and correlates of severity. Science 2020, 370. [Google Scholar] [CrossRef] [PubMed]
- Tso, F.Y.; Lidenge, S.J.; Pena, P.B.; Clegg, A.A.; Ngowi, J.R.; Mwaiselage, J.; Ngalamika, O.; Julius, P.; West, J.T.; Wood, C. High prevalence of pre-existing serological cross-reactivity against severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) in sub-Saharan Africa. Int. J. Infect. Dis. 2021, 102, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Nguyen-Contant, P.; Embong, A.K.; Kanagaiah, P.; Chaves, F.A.; Yang, H.; Branche, A.R.; Topham, D.J.; Sangster, M.Y. S Protein-Reactive IgG and Memory B Cell Production after Human SARS-CoV-2 Infection Includes Broad Reactivity to the S2 Subunit. mBio 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Dalakas, M.C.; Bitzogli, K.; Alexopoulos, H. Anti-SARS-CoV-2 Antibodies Within IVIg Preparations: Cross-Reactivities with Seasonal Coronaviruses, Natural Autoimmunity, and Therapeutic Implications. Front. Immunol 2021, 12, 627285. [Google Scholar] [CrossRef] [PubMed]
- Reche, P.A. Potential Cross-Reactive Immunity to SARS-CoV-2 From Common Human Pathogens and Vaccines. Front. Immunol. 2020, 11, 586984. [Google Scholar] [CrossRef]
- Yonker, L.M.; Neilan, A.M.; Bartsch, Y.; Patel, A.B.; Regan, J.; Arya, P.; Gootkind, E.; Park, G.; Hardcastle, M.; St John, A.; et al. Pediatric Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): Clinical Presentation, Infectivity, and Immune Responses. J. Pediatr. 2020, 227, P45–52.e5. [Google Scholar] [CrossRef]
- Amanat, F.; Stadlbauer, D.; Strohmeier, S.; Nguyen, T.H.O.; Chromikova, V.; McMahon, M.; Jiang, K.; Arunkumar, G.A.; Jurczyszak, D.; Polanco, J.; et al. A serological assay to detect SARS-CoV-2 seroconversion in humans. Nat. Med. 2020, 26, 1033–1036. [Google Scholar] [CrossRef]
- Loos, C.; Atyeo, C.; Fischinger, S.; Burke, J.; Slein, M.D.; Streeck, H.; Lauffenburger, D.; Ryan, E.T.; Charles, R.C.; Alter, G. Evolution of Early SARS-CoV-2 and Cross-Coronavirus Immunity. mSphere 2020, 5. [Google Scholar] [CrossRef]
- Caton, A.J.; Brownlee, G.G.; Yewdell, J.W.; Gerhard, W. The antigenic structure of the influenza virus A/PR/8/34 hemagglutinin (H1 subtype). Cell 1982, 31, 417–427. [Google Scholar] [CrossRef]
- Gerhard, W.; Yewdell, J.; Frankel, M.E.; Webster, R. Antigenic structure of influenza virus haemagglutinin defined by hybridoma antibodies. Nature 1981, 290, 713–717. [Google Scholar] [CrossRef]
- Gerhard, W.; Webster, R.G. Antigenic drift in influenza A viruses I. Selection and characterization of antigenic variants of A/PR/8/34 [H0N1] influenza virus with monoclonal antibodies. J. Exp. Med. 1978, 148, 383–392. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Seiler, P.; Jones, J.C.; Ridout, G.; Camp, K.P.; Fabrizio, T.P.; Jeevan, T.; Miller, L.A.; Throm, R.E.; Ferrara, F.; et al. Antibody Responses to SARS-CoV-2 Antigens in Humans and Animals. Vaccines 2020, 8, 684. [Google Scholar] [CrossRef] [PubMed]
- Premkumar, L.; Segovia-Chumbez, B.; Jadi, R.; Martinez, D.R.; Raut, R.; Markmann, A.; Cornaby, C.; Bartelt, L.; Weiss, S.; Park, Y.; et al. The RBD of the Spike Protein of SARS-Group Coronaviruses Is A Highly Specific Target of SARS-CoV-2 Antibodies But Not Other Pathogenic Human and Animal Coronavirus Antibodies. medRxiv 2020. [Google Scholar] [CrossRef]
- Mateus, J.; Grifoni, A.; Tarke, A.; Sidney, J.; Ramirez, S.I.; Dan, J.M.; Burger, Z.C.; Rawlings, S.A.; Smith, D.M.; Phillips, E.; et al. Selective and cross-reactive SARS-CoV-2 T cell epitopes in unexposed humans. Science 2020, 370, 89–94. [Google Scholar] [CrossRef]
- Sette, A.; Crotty, S. Pre-existing immunity to SARS-CoV-2: The knowns and unknowns. Nat. Rev. Immunol. 2020, 20, 457–458. [Google Scholar] [CrossRef]
- Le Bert, N.; Tan, A.T.; Kunasegaran, K.; Tham, C.Y.L.; Hafezi, M.; Chia, A.; Chng, M.H.Y.; Lin, M.; Tan, N.; Linster, M.; et al. SARS-CoV-2-specific T cell immunity in cases of COVID-19 and SARS, and uninfected controls. Nature 2020, 584, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Meckiff, B.J.; Ramirez-Suastegui, C.; Fajardo, V.; Chee, S.J.; Kusnadi, A.; Simon, H.; Grifoni, A.; Pelosi, E.; Weiskopf, D.; Sette, A.; et al. Single-cell transcriptomic analysis of SARS-CoV-2 reactive CD4 (+) T cells. bioRxiv 2020. [Google Scholar] [CrossRef]
- Braun, J.; Loyal, L.; Frentsch, M.; Wendisch, D.; Georg, P.; Kurth, F.; Hippenstiel, S.; Dingeldey, M.; Kruse, B.; Fauchere, F.; et al. SARS-CoV-2-reactive T cells in healthy donors and patients with COVID-19. Nature 2020, 587, 270–274. [Google Scholar] [CrossRef]
- Saletti, G.; Gerlach, T.; Jansen, J.M.; Molle, A.; Elbahesh, H.; Ludlow, M.; Li, W.; Bosch, B.J.; Osterhaus, A.; Rimmelzwaan, G.F. Older adults lack SARS CoV-2 cross-reactive T lymphocytes directed to human coronaviruses OC43 and NL63. Sci. Rep. 2020, 10, 21447. [Google Scholar] [CrossRef] [PubMed]
- Nelde, A.; Bilich, T.; Heitmann, J.S.; Maringer, Y.; Salih, H.R.; Roerden, M.; Lubke, M.; Bauer, J.; Rieth, J.; Wacker, M.; et al. SARS-CoV-2-derived peptides define heterologous and COVID-19-induced T cell recognition. Nat. Immunol. 2021, 22, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Schulien, I.; Kemming, J.; Oberhardt, V.; Wild, K.; Seidel, L.M.; Killmer, S.; Sagar; Daul, F.; Salvat Lago, M.; Decker, A.; et al. Characterization of pre-existing and induced SARS-CoV-2-specific CD8(+) T cells. Nat. Med. 2021, 27, 78–85. [Google Scholar] [CrossRef]
- Woldemeskel, B.A.; Kwaa, A.K.; Garliss, C.C.; Laeyendecker, O.; Ray, S.C.; Blankson, J.N. Healthy donor T cell responses to common cold coronaviruses and SARS-CoV-2. J. Clin. Investig. 2020, 130, 6631–6638. [Google Scholar] [CrossRef]
- Scherle, P.A.; Gerhard, W. Functional analysis of influenza-specific helper T cell clones in vivo. T cells specific for internal viral proteins provide cognate help for B cell responses to hemagglutinin. J. Exp. Med. 1986, 164, 1114–1128. [Google Scholar] [CrossRef] [Green Version]
- Sagar, M.; Reifler, K.; Rossi, M.; Miller, N.S.; Sinha, P.; White, L.F.; Mizgerd, J.P. Recent endemic coronavirus infection is associated with less-severe COVID-19. J. Clin. Investig. 2021, 131. [Google Scholar] [CrossRef] [PubMed]
- Malani, A.; Shah, D.; Kang, G.; Lobo, G.N.; Shastri, J.; Mohanan, M.; Jain, R.; Agrawal, S.; Juneja, S.; Imad, S.; et al. Seroprevalence of SARS-CoV-2 in slums versus non-slums in Mumbai, India. Lancet Glob. Health 2021, 9, e110–e111. [Google Scholar] [CrossRef]
- Capoor, M.N.; Ahmed, F.S.; McDowell, A.; Slaby, O. Is the “Common Cold” Our Greatest Ally in the Battle Against SARS-CoV-2? Front. Cell Infect. Microbiol. 2020, 10, 605334. [Google Scholar] [CrossRef]
- Shukla, R.; Ramasamy, V.; Shanmugam, R.K.; Ahuja, R.; Khanna, N. Antibody-Dependent Enhancement: A Challenge for Developing a Safe Dengue Vaccine. Front. Cell Infect. Microbiol. 2020, 10, 572681. [Google Scholar] [CrossRef]
- Beretta, A.; Cranage, M.; Zipeto, D. Is Cross-Reactive Immunity Triggering COVID-19 Immunopathogenesis? Front. Immunol 2020, 11, 567710. [Google Scholar] [CrossRef]
- Mudd, P.A.; Crawford, J.C.; Turner, J.S.; Souquette, A.; Reynolds, D.; Bender, D.; Bosanquet, J.P.; Anand, N.J.; Striker, D.A.; Martin, R.S.; et al. Distinct inflammatory profiles distinguish COVID-19 from influenza with limited contributions from cytokine storm. Sci. Adv. 2020, 6. [Google Scholar] [CrossRef]
- Callow, K.A.; Parry, H.F.; Sergeant, M.; Tyrrell, D.A. The time course of the immune response to experimental coronavirus infection of man. Epidemiol. Infect. 1990, 105, 435–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, S.E. The behaviour of recent isolates of human respiratory coronavirus in vitro and in volunteers: Evidence of heterogeneity among 229E-related strains. J. Med. Virol. 1984, 13, 179–192. [Google Scholar] [CrossRef]
- COVID-19: World’s First Human Challenge Trials to Start in UK, 17 February 2021. Available online: https://www.bbc.com (accessed on 6 June 2021).
- Kirby, T. COVID-19 human challenge studies in the UK. Lancet Respir. Med. 2020, 8, e96. [Google Scholar] [CrossRef]
- Kuiper, V.P.; Rosendaal, F.R.; Kamerling, I.M.C.; Visser, L.G.; Roestenberg, M. Assessment of risks associated with SARS-CoV-2 experimental human infection studies. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Sia, S.F.; Yan, L.M.; Chin, A.W.H.; Fung, K.; Choy, K.T.; Wong, A.Y.L.; Kaewpreedee, P.; Perera, R.; Poon, L.L.M.; Nicholls, J.M.; et al. Pathogenesis and transmission of SARS-CoV-2 in golden hamsters. Nature 2020, 583, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Sealy, R.; Chaka, W.; Surman, S.; Brown, S.A.; Cresswell, P.; Hurwitz, J.L. Target peptide sequence within infectious human immunodeficiency virus type 1 does not ensure envelope-specific T-helper cell reactivation: Influences of cysteine protease and gamma interferon-induced thiol reductase activities. Clin. Vaccine Immunol. 2008, 15, 713–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Authors | Manuscript Title | Results | Paper |
|---|---|---|---|
| Khan et al. | Cross-reactivity between common human coronaviruses and SARS-Cov-2 using coronavirus antigen microarray | Khan et al. used pre-pandemic sera and antigen microarrays to study antibody responses. They observed IgG reactivity toward each of the common cold HCoVs in four of five tested serum samples, but only weak IgG reactivity toward SARS-CoV-2 in all five samples. Weak signals were directed toward SARS-CoV-2 N and S2 antigens. | [47] |
| Guo et al. | Profiling early humoral response to diagnose novel coronavirus disease (COVID-19) | Guo et al. did not observe SARS-CoV-2 N-specific antibodies within pre-pandemic samples that exhibited positive antibodies toward N proteins of NL63, 229E, OC43, and HKU1. | [56] |
| Mveang Nzoghet al. | Evidence and implications of pre-existing humoral cross-reactive immunity to SARS-CoV-2. | Mveang Nzoghe et al. tested samples from healthy volunteers taken in 2014. Of 135 samples, 32 (23.7%) tested positive for antibodies against SARS-CoV-2 N. | [57] |
| To et al. | Seroprevalence of SARS-CoV-2 in Hong Kong and in residents evacuated from Hubei province, China: a multicohort study. | To et al. found that in a Hong Kong population presumed to be SARS-CoV-2-unexposed, 53 of 1938 samples (2.73%) were positive for antibody binding in a SARS-CoV-2 enzyme-linked immunosorbent assay (ELISA). | [17,58,59] |
| Shrock et al. | Viral epitope profiling of COVID-19 patients reveals cross-reactivity and correlates of severity. | Using VirScan technology, Shrock et al. observed antibodies to SARS-CoV-2 ORF-1, but relatively weak or no activity toward SARS-CoV-2 S or N among 190 pre-pandemic samples. | [60] |
| Anderson et al. | Seasonal human coronavirus antibodies are boosted upon SARS-CoV-2 infection but not associated with protection. | Anderson et al. reported that in pre-pandemic samples taken in 2017, there was antibody binding to SARS-CoV-2 full-length S in 5.4% samples and antibody binding to SARS-CoV-2 RBD of the S protein in 2% of the samples. SARS-CoV-2 N-specific antibodies were detected in 18.6% of the samples. In this case, antibodies were non-neutralizing. | [46] |
| Tso et al. | High prevalence of pre-existing serological cross-reactivity against severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) in sub-Saharan Africa. | Tso et al. identified higher frequencies of SARS-CoV-2-specific antibodies in their study of pre-pandemic samples from individuals in African countries compared to individuals in the USA. Specifically, the prevalence of serological activity against SARS-COV-2 in assays was 19% among samples from Tanzania and 14.1% among samples from Zambia, but only 2.4% among samples from the USA. Among the samples from Africa, responses toward N were higher than those toward S. | [61] |
| Nguyen-Contant et al. | S protein-reactive IgG and memory B cell production after human SARS-Cov-2 infection includes broad reactivity to the S2 subunit. | Nguyen-Contant et al. examined 21 pre-pandemic samples collected from 2011 to 2014 at the University of Rochester (Rochester, New York, USA). Antibody responses toward SARS-CoV-2 were generally weak. Nonetheless, authors detected serum IgG toward a stabilized ectodomain of SARS-CoV-2 S. The responses were also identified toward SARS-CoV-2 N. Responses toward the SARS-CoV-2 RBD were not identified, but 86% of samples bound SARS-CoV-2 S2. The authors contemplated that responses toward S2 might provide neutralizing activity and a degree of protection against SARS-CoV-2 in humans. | [62] |
| Ng et al. | Preexisting and de novo humoral immunity to SARS-CoV-2 in humans. | Ng et al. frequently observed SARS-CoV-2 S-specific antibodies in SARS-CoV-2 unexposed individuals. The highest frequencies were observed in children. In a population of SARS-CoV-2-uninfected healthy children between the ages of 1 and 16 years, at least 21 of 48 samples scored positively for SARS-CoV-2 S-specific IgG. Anti-S2 antibodies from SARS-CoV-2-uninfected patients exhibited neutralizing activity against SARS-CoV-2. | [49] |
| Dalakas et al. | Anti-SARS-Cov-2 antibodies within IVIG preparations: cross-reactivities with seasonal coronaviruses, natural autoimmunity and therapeutic implications. | Dalakas et al. found that the majority of tested, pre-pandemic IVIG preparations (immunoglobulins from pooled serum samples) contained antibodies that cross-reacted with SARS-CoV-2. | [63] |
| Song et al. | Cross-reactive serum and memory B cell responses to spike protein in SARS-CoV-2 and endemic coronavirus infection. | Song et al. observed minimal or no reactivity to SARS-CoV-2 S among pre-pandemic samples. | [55] |
| Authors | Manuscript Title | Results | Paper |
|---|---|---|---|
| Nguyen-Contant et al. | S protein-reactive IgG and memory B cell production after human SARS-CoV-2 infection includes broad reactivity to the S2 subunit. | Nguyen-Contant et al. observed upregulation of antibodies toward common cold HCoVs in COVID-19 patients compared to SARS-CoV-2-unexposed controls. As an example, IgG titers toward OC43 S were higher in SARS-CoV-2 convalescent patients compared to unexposed donors. Both S2- and N-specific responses were significantly upregulated in the SARS-CoV-2 -exposed blood donors.. | [62] |
| Shrock et al. | Viral epitope profiling of COVID-19 patients reveals cross-reactivity and correlates of severity. | Shrock et al. used VirScan technology to identify peptide-specific responses that increased in SARS-CoV-2-exposed individuals. They observed elevated antibodies toward peptides in S and N. For example, antibodies toward SARS-CoV-2 S peptides that spanned the region of amino acids 811–830 were increased. An 11 amino acid stretch in this region was found to be highly conserved between SARS-CoV-2 and all four of the common cold HCoVs. This peptide overlapped with the S2 fusion peptide. Additionally, antibodies that targeted another region of the SARS-CoV-2 S2 peptide, amino acids 1144–1163, increased in SARS-CoV-2-exposed individuals compared to unexposed donors. A ten amino acid stretch within this sequence was well matched between SARS-CoV-2 and OC43. | [60] |
| Yonker et al. | Pediatric Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): Clinical Presentation, Infectivity, and Immune Responses. | Yonker et al. identified improved responses to the RBD of common cold HCoVs (229E, NL63, HKU1, and OC43) in children who were suffering MIS-C due to SARS-CoV2. However, these children also experienced increases in antibodies toward respiratory syncytial virus and influenza virus, suggesting that B cells were activated non-specifically. | [65] |
| Song et al. | Cross-reactive serum and memory B cell responses to spike protein in SARS-CoV-2 and endemic coronavirus infection. | Authors suggested that pre-existing cross-reactive memory B cells were activated during SARS-CoV-2 infection. Authors identified elevated antibodies toward the HKU1 and NL63 S proteins in SARS-CoV-2-exposed individuals compared to pre-pandemic samples. They also characterized a cross-reactive, S2-specific monoclonal antibody with neutralizing activity. Specifically, single-particle negative stain electron microscopy was used to show that monoclonal CC40.8 bound to the HKU1 S trimer near the bottom of the S2 domain. | [55] |
| Anderson et al. | Seasonal human coronavirus antibodies are boosted upon SARS-CoV-2 infection but not associated with protection. | Anderson et al. tested samples from 27 hospitalized COVID-19 patients longitudinally and observed increases in antibodies that recognized the S protein of OC43 and SARS-CoV-2 over the course of hospitalization. | [46] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sealy, R.E.; Hurwitz, J.L. Cross-Reactive Immune Responses toward the Common Cold Human Coronaviruses and Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): Mini-Review and a Murine Study. Microorganisms 2021, 9, 1643. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9081643
Sealy RE, Hurwitz JL. Cross-Reactive Immune Responses toward the Common Cold Human Coronaviruses and Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): Mini-Review and a Murine Study. Microorganisms. 2021; 9(8):1643. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9081643
Chicago/Turabian StyleSealy, Robert E., and Julia L. Hurwitz. 2021. "Cross-Reactive Immune Responses toward the Common Cold Human Coronaviruses and Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): Mini-Review and a Murine Study" Microorganisms 9, no. 8: 1643. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9081643