
Microbial L-asparaginase for Application in Acrylamide Mitigation from Food: Current Research Status and Future Perspectives


Abstract
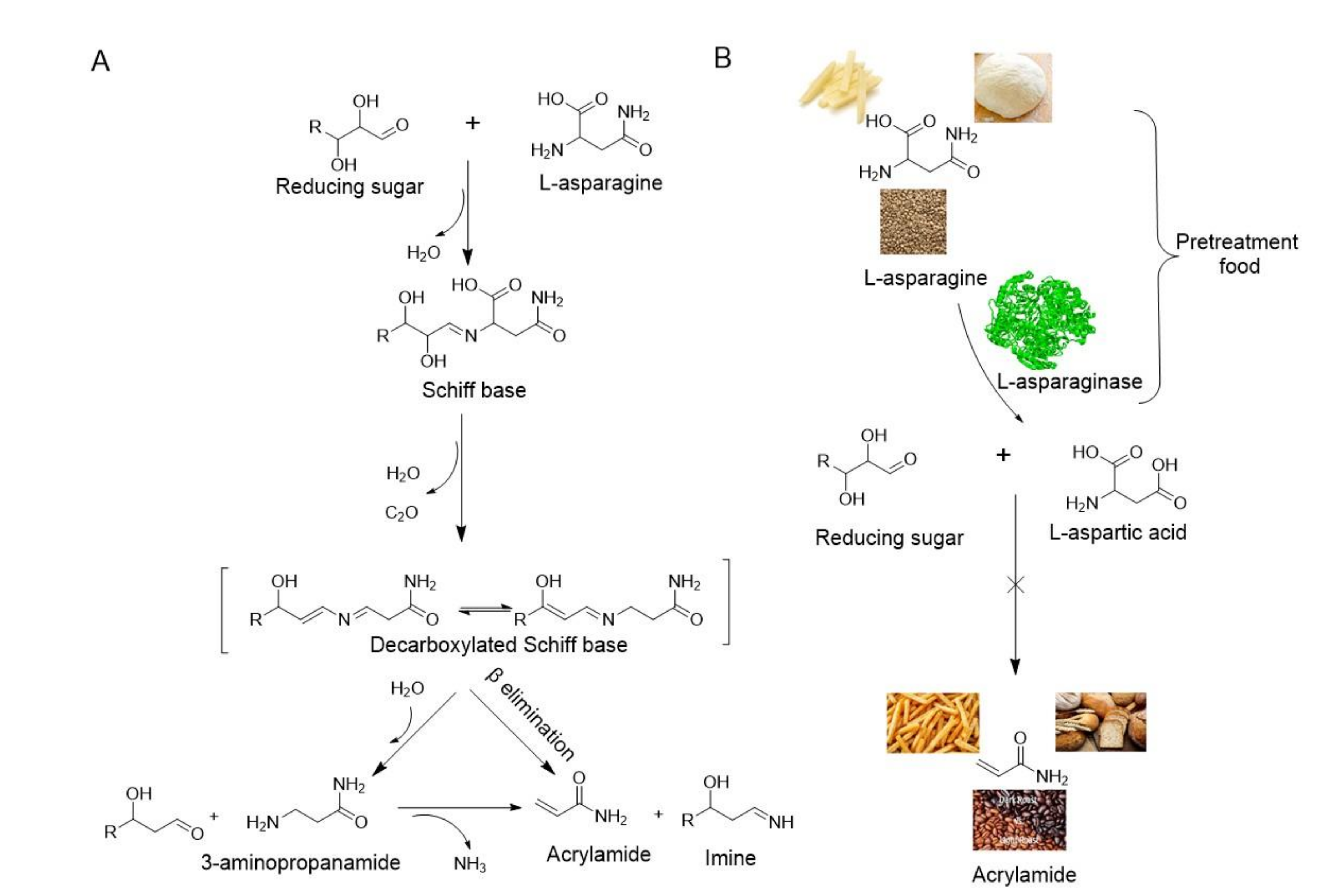
:1. Introduction
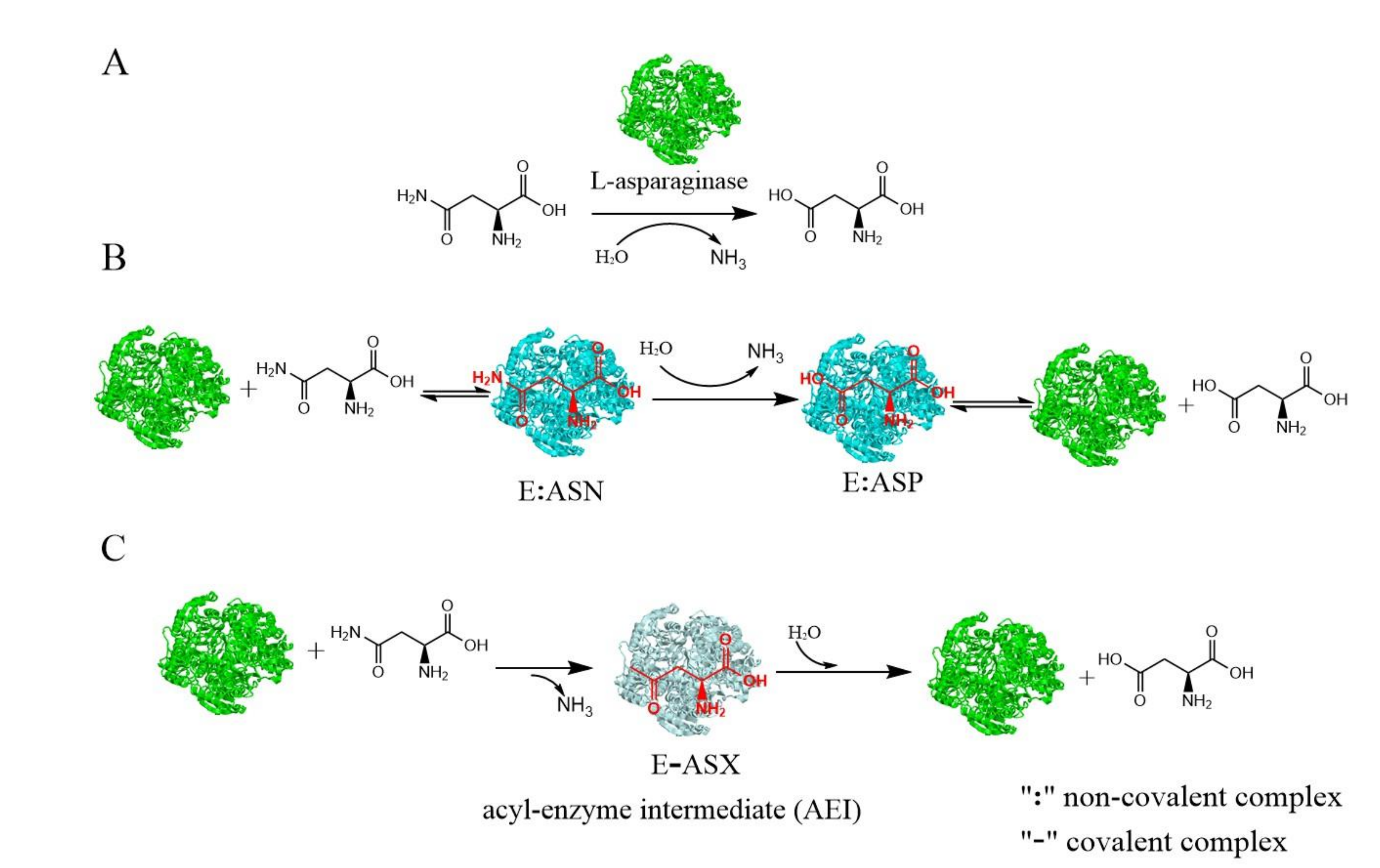
2. Classification and Catalytic Mechanism of L-asparaginase
3. Sources of L-asparaginase
4. Production of L-asparaginase
5. Purification and Biochemical Properties of Microbial L-asparaginase
6. Application of Microbial L-asparaginase in Food
6.1. Fried Potato Products
6.2. Bakery Products
6.3. Coffee
6.4. Industrial Processes
7. Application of Immobilized L-asparaginase in Food
8. Discussion
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Clementi, A. La désamidation enzymatique de l’asparagine chez les différentes espéces animales et la signification physio logique de sa presence dans l’organisme. Arch. Int. Physiol. 1922, 19, 369–398. [Google Scholar] [CrossRef]
- Strzelczyk, P.; Zhang, D.; Dyba, M.; Wlodawer, A.; Lubkowski, J. Generalized enzymatic mechanism of catalysis by tetrameric L-asparaginases from mesophilic bacteria. Sci. Rep. 2020, 10, 17516. [Google Scholar] [CrossRef]
- Zuo, S.; Zhang, T.; Jiang, B.; Mu, W. Recent research progress on microbial L-asparaginases. Appl. Microbiol. Biotechnol. 2015, 99, 1069–1079. [Google Scholar] [CrossRef]
- Chand, S.; Mahajan, R.V.; Prasad, J.P.; Sahoo, D.K.; Mihooliya, K.N.; Dhar, M.S.; Sharma, G. A comprehensive review on microbial L-asparaginase: Bioprocessing, characterization, and industrial applications. Biotechnol. Appl. Biochem. 2020, 67, 619–647. [Google Scholar] [CrossRef] [PubMed]
- Muneer, F.; Siddique, M.H.; Azeem, F.; Rasul, I.; Muzammil, S.; Zubair, M.; Afzal, M.; Nadeem, H. Microbial L-asparaginase: Purification, characterization and applications. Arch. Microbiol. 2020, 202, 967–981. [Google Scholar] [CrossRef]
- Safary, A.; Moniri, R.; Hamzeh-Mivehroud, M.; Dastmalchi, S. Highly efficient novel recombinant L-asparaginase with no glutaminase activity from a new halo-thermotolerant Bacillus strain. BioImpacts BI 2019, 9, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Kataria, M.; Kaur, N.; Narula, R.; Kumar, K.; Kataria, S.; Verma, N. L-Asparaginase from Novel Source-Solanum nigrum and Development of Asparagine Biosensor. Pharma Innov. 2015, 4, 81. [Google Scholar]
- Tareke, E.; Rydberg, P.; Karlsson, P.; Eriksson, S.; Törnqvist, M. Analysis of Acrylamide, a Carcinogen Formed in Heated Foodstuffs. J. Agric. Food Chem. 2002, 50, 4998–5006. [Google Scholar] [CrossRef]
- Svensson, K.; Abramsson, L.; Becker, W.; Glynn, A.; Hellenäs, K.E.; Lind, Y.; Rosén, J. Dietary intake of acrylamide in Sweden. Food Chem. Toxicol. 2003, 41, 1581–1586. [Google Scholar] [CrossRef]
- Stadler, R.H.; Blank, I.; Varga, N.; Robert, F.; Hau, J.; Guy, P.A.; Robert, M.-C.; Riediker, S. Acrylamide from Maillard reaction products. Nature 2002, 419, 449–450. [Google Scholar] [CrossRef] [PubMed]
- Mottram, D.S.; Wedzicha, B.L.; Dodson, A.T. Acrylamide is formed in the Maillard reaction. Nature 2002, 419, 448–449. [Google Scholar] [CrossRef]
- Zuo, S.; Zhang, T.; Jiang, B.; Mu, W. Reduction of acrylamide level through blanching with treatment by an extremely thermostable L-asparaginase during French fries processing. Extremophiles 2015, 19, 841–851. [Google Scholar] [CrossRef]
- Friedman, M. Chemistry, Biochemistry, and Safety of Acrylamide. A Review. J. Agric. Food Chem. 2003, 51, 4504–4526. [Google Scholar] [CrossRef]
- Zyzak, D.V.; Sanders, R.A.; Stojanovic, M.; Tallmadge, D.H.; Eberhart, B.L.; Ewald, D.K.; Gruber, D.C.; Morsch, T.R.; Strothers, M.A.; Rizzi, G.P.; et al. Acrylamide Formation Mechanism in Heated Foods. J. Agric. Food Chem. 2003, 51, 4782–4787. [Google Scholar] [CrossRef]
- Mohan Kumar, N.S.; Shimray, C.A.; Indrani, D.; Manonmani, H.K. Reduction of Acrylamide Formation in Sweet Bread with L-Asparaginase Treatment. Food Bioprocess Technol. 2014, 7, 741–748. [Google Scholar] [CrossRef]
- Corrêa, C.L.O.; das Merces Penha, E.; dos Anjos, M.R.; Pacheco, S.; Freitas-Silva, O.; Luna, A.S.; Gottschalk, L.M.F. Use of asparaginase for acrylamide mitigation in coffee and its influence on the content of caffeine, chlorogenic acid, and caffeic acid. Food Chem. 2021, 338, 128045. [Google Scholar] [CrossRef]
- Lyon, F. IARC monographs on the evaluation of carcinogenic risks to humans. Some Ind. Chem. 1994, 60, 389–433. [Google Scholar]
- Commission, E. Commission Regulation (EU) 2017/2158 of 20 November 2017 establishing mitigation measures and benchmark levels for the reduction of the presence of acrylamide in food. Off. J. Eur. Union 2017, 304, 24–44. [Google Scholar]
- Khalil, N.M.; Rodríguez-Couto, S.; El-Ghany, M.N.A. Characterization of Penicillium crustosum L-asparaginase and its acrylamide alleviation efficiency in roasted coffee beans at non-cytotoxic levels. Arch. Microbiol. 2021. [Google Scholar] [CrossRef]
- Sun, Z.; Qin, R.; Li, D.; Ji, K.; Wang, T.; Cui, Z.; Huang, Y. A novel bacterial type II L-asparaginase and evaluation of its enzymatic acrylamide reduction in French fries. Int. J. Biol. Macromol. 2016, 92, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Kornbrust, B.A.; Stringer, M.A.; Lange, N.E.K.; Hendriksen, H.V.; Whitehurst, R.; Oort, M. Asparaginase–An enzyme for acrylamide reduction in food products. Enzym. Food Technol. 2010, 2, 59–87. [Google Scholar]
- Adebo, O.A.; Kayitesi, E.; Adebiyi, J.A.; Gbashi, S.; Temba, M.C.; Lasekan, A.; Phoku, J.Z.; Njobeh, P.B. Mitigation of acrylamide in foods: An African perspective. Acrylic Polym. Healthc. 2017, 152–172. [Google Scholar]
- Xu, F.; Oruna-Concha, M.-J.; Elmore, J.S. The use of asparaginase to reduce acrylamide levels in cooked food. Food Chem. 2016, 210, 163–171. [Google Scholar] [CrossRef]
- Hendriksen, H.V.; Kornbrust, B.A.; Østergaard, P.R.; Stringer, M.A. Evaluating the Potential for Enzymatic Acrylamide Mitigation in a Range of Food Products Using an Asparaginase from Aspergillus oryzae. J. Agric. Food Chem. 2009, 57, 4168–4176. [Google Scholar] [CrossRef] [PubMed]
- Chohan, S.M.; Sajed, M.; Naeem, S.U.; Rashid, N. Heterologous gene expression and characterization of TK2246, a highly active and thermostable plant type l-asparaginase from Thermococcus kodakarensis. Int. J. Biol. Macromol. 2020, 147, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Li, D.; Liu, P.; Wang, W.; Ji, K.; Huang, Y.; Cui, Z. A novel L-asparaginase from Aquabacterium sp. A7-Y with self-cleavage activation. Antonie van Leeuwenhoek 2016, 109, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Karamitros, C.S.; Konrad, M. Human 60-kDa lysophospholipase contains an N-terminal L-asparaginase domain that is allosterically regulated by L-asparagine. J. Biol. Chem. 2014, 289, 12962–12975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bejger, M.; Imiolczyk, B.; Clavel, D.; Gilski, M.; Pajak, A.; Marsolais, F.; Jaskolski, M. Na+/K+ exchange switches the catalytic apparatus of potassium-dependent plant L-asparaginase. Acta Crystallogr. Sect. D Biol. Crystallogr. 2014, 70, 1854–1872. [Google Scholar] [CrossRef]
- Moreno-Enriquez, A.; Evangelista-Martinez, Z.; Gonzalez-Mondragon, E.G.; Calderon-Flores, A.; Arreguin, R.; Perez-Rueda, E.; Huerta-Saquero, A. Biochemical characterization of recombinant L-asparaginase (AnsA) from Rhizobium etli, a member of an increasing rhizobial-type family of L-asparaginases. J. Microbiol. Biotechnol. 2012, 22, 292–300. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, X.; Xu, S.; Xu, M.; Yang, T.; Wang, L.; Zhang, H.; Fang, H.; Osire, T.; Rao, Z. Insight into the thermostability of thermophilic L-asparaginase and non-thermophilic L-asparaginase II through bioinformatics and structural analysis. Appl. Microbiol. Biotechnol. 2019, 103, 7055–7070. [Google Scholar] [CrossRef]
- Maggi, M.; Chiarelli, L.R.; Valentini, G.; Scotti, C. Tackling Critical Catalytic Residues in Helicobacter pylori L-Asparaginase. Biomolecules 2015, 5, 306–317. [Google Scholar] [CrossRef] [Green Version]
- Radadiya, A.; Zhu, W.; Coricello, A.; Alcaro, S.; Richards, N.G.J. Improving the Treatment of Acute Lymphoblastic Leukemia. Biochemistry 2020, 59, 3193–3200. [Google Scholar] [CrossRef] [PubMed]
- Schalk, A.M.; Antansijevic, A.; Caffrey, M.; Lavie, A. Experimental Data in Support of a Direct Displacement Mechanism for Type I/II L-Asparaginases. J. Biol. Chem. 2016, 291, 5088–5100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lubkowski, J.; Vanegas, J.; Chan, W.K.; Lorenzi, P.L.; Weinstein, J.N.; Sukharev, S.; Fushman, D.; Rempe, S.; Anishkin, A.; Wlodawer, A. Mechanism of Catalysis by L-Asparaginase. Biochemistry 2020, 59, 1927–1945. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, N.E.-A.; El-Shweihy, N.M. Bioprocess development for L-asparaginase production by Streptomyces rochei, purification and in-vitro efficacy against various human carcinoma cell lines. Sci. Rep. 2020, 10, 7942. [Google Scholar] [CrossRef] [PubMed]
- Barati, M.; Faramarzi, M.A.; Nafissi-Varcheh, N.; Khoshayand, M.R.; Tehrani, M.H.H.; Vahidi, H.; Adrangi, S. L-asparaginase activity in cell lysates and culture media of halophilic bacterial isolates. Iran. J. Pharm. Res. IJPR 2016, 15, 435. [Google Scholar]
- Dias, F.F.G.; Santos Aguilar, J.G.D.; Sato, H.H. L-Asparaginase from Aspergillus spp.: Production based on kinetics, thermal stability and biochemical characterization. 3 Biotech 2019, 9, 289. [Google Scholar] [CrossRef] [PubMed]
- Abdelrazek, N.A.; Elkhatib, W.F.; Raafat, M.M.; Aboulwafa, M.M. Production, characterization and bioinformatics analysis of L-asparaginase from a new Stenotrophomonas maltophilia EMCC2297 soil isolate. AMB Express 2020, 10, 71. [Google Scholar] [CrossRef] [Green Version]
- Freire, R.K.B.; Mendonça, C.M.N.; Ferraro, R.B.; Moguel, I.S.; Tonso, A.; Lourenço, F.R.; Santos, J.H.P.M.; Sette, L.D.; Pessoa Junior, A. Glutaminase-free L-asparaginase production by Leucosporidium muscorum isolated from Antarctic marine-sediment. Prep. Biochem. Biotechnol. 2021, 51, 277–288. [Google Scholar] [CrossRef]
- Guo, J.; Coker, A.R.; Wood, S.P.; Cooper, J.B.; Chohan, S.M.; Rashid, N.; Akhtar, M. Structure and function of the thermostable L-asparaginase from Thermococcus kodakarensis. Acta Crystallogr. Sect. D Struct. Biol. 2017, 73, 889–895. [Google Scholar] [CrossRef] [Green Version]
- Chohan, S.M.; Rashid, N. TK1656, a thermostable L-asparaginase from Thermococcus kodakaraensis, exhibiting highest ever reported enzyme activity. J. Biosci. Bioeng. 2013, 116, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.-J.; Lee, Y.-H.; Khan, A.R.; Ullah, I.; Lee, C.; Park, C.K.; Shin, J.-H. Cloning, expression, and characterization of thermophilic L-asparaginase from Thermococcus kodakarensis KOD1. J. Basic Microbiol. 2014, 54, 500–508. [Google Scholar] [CrossRef]
- Chohan, S.M.; Rashid, N.; Sajed, M.; Imanaka, T. Pcal_0970: An extremely thermostable L-asparaginase from Pyrobaculum calidifontis with no detectable glutaminase activity. Folia Microbiol. 2019, 64, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, X.; Xu, S.; Zhang, H.; Xu, M.; Yang, T.; Wang, L.; Qian, H.; Zhang, H.; Fang, H.; et al. Simultaneous cell disruption and semi-quantitative activity assays for high-throughput screening of thermostable L-asparaginases. Sci. Rep. 2018, 8, 7915. [Google Scholar] [CrossRef]
- Jiao, L.; Chi, H.; Lu, Z.; Zhang, C.; Chia, S.R.; Show, P.L.; Tao, Y.; Lu, F. Characterization of a novel type I L-asparaginase from Acinetobacter soli and its ability to inhibit acrylamide formation in potato chips. J. Biosci. Bioeng. 2020, 129, 672–678. [Google Scholar] [CrossRef]
- Ameen, F.; Alshehri, W.A.; Al-Enazi, N.M.; Almansob, A. L-Asparaginase activity analysis, ansZ gene identification and anticancer activity of a new Bacillus subtilis isolated from sponges of the Red Sea. Biosci. Biotechnol. Biochem. 2020, 84, 2576–2584. [Google Scholar] [CrossRef] [PubMed]
- Aly, N.; El-Ahwany, A.; Ataya, F.S.; Saeed, H. Bacillus sonorensis L. Asparaginase: Cloning, Expression in E. coli and Characterization. Protein J. 2020, 39, 717–729. [Google Scholar] [CrossRef]
- Abdelrazek, N.A.; Elkhatib, W.F.; Raafat, M.M.; Aboulwafa, M.M. Experimental and bioinformatics study for production of L-asparaginase from Bacillus licheniformis: A promising enzyme for medical application. AMB Express 2019, 9, 39. [Google Scholar] [CrossRef] [PubMed]
- Azmi, W.; Narta, U.K.; Singh, N.; Gupta, V.; Singh Kanwar, S.; Devi, S. An Antileukemic Glutaminase Free L-Asparaginase from Bacillus brevis. Curr. Biotechnol. 2017, 6, 58–68. [Google Scholar] [CrossRef]
- Sanghvi, G.; Bhimani, K.; Vaishnav, D.; Oza, T.; Dave, G.; Kunjadia, P.; Sheth, N. Mitigation of acrylamide by L-asparaginase from Bacillus subtilis KDPS1 and analysis of degradation products by HPLC and HPTLC. SpringerPlus 2016, 5, 533. [Google Scholar] [CrossRef] [Green Version]
- Aishwarya, S.S.; Selvarajan, E.; Iyappan, S.; Rajnish, K.N. Recombinant l-Asparaginase II from Lactobacillus casei subsp. casei ATCC 393 and Its Anticancer Activity. Indian J. Microbiol. 2019, 59, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Husain, I. Evaluation of Antitumor Activity of Glutaminase-Free Periplasmic Asparaginase from Indigenous Bacterial Isolates as Candidates for Cancer Therapy. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2017, 87, 997–1004. [Google Scholar] [CrossRef]
- Bhagat, J.; Kaur, A.; Chadha, B.S. Single step purification of asparaginase from endophytic bacteria Pseudomonas oryzihabitans exhibiting high potential to reduce acrylamide in processed potato chips. Food Bioprod. Process. 2016, 99, 222–230. [Google Scholar] [CrossRef]
- Ghasemi, A.; Asad, S.; Kabiri, M.; Dabirmanesh, B. Cloning and characterization of Halomonas elongata L-asparaginase, a promising chemotherapeutic agent. Appl. Microbiol. Biotechnol. 2017, 101, 7227–7238. [Google Scholar] [CrossRef] [PubMed]
- Soliman, H.M.; El-Naggar, N.E.-A.; El-Ewasy, S.M. Bioprocess Optimization for Enhanced Production of L-asparaginase via Two Model-Based Experimental Designs by Alkaliphilic Streptomyces fradiae NEAE-82. Curr. Biotechnol. 2020, 9, 23–37. [Google Scholar] [CrossRef]
- da Silva Lacerda, G.R.; Cantalice, J.C.L.L.; de Souza Lima, G.M.; de Albuquerque, L.E.F.; da Silva, I.D.E.G.; de Melo, M.E.B.; Adam, M.L.; do Nascimento, S.C. Genotoxic activity of L-asparaginase produced by Streptomyces ansochromogenes UFPEDA 3420. World J. Microbiol. Biotechnol. 2019, 35, 41. [Google Scholar] [CrossRef]
- da Silva Lacerda, G.R.; de Melo, C.M.L.; de Araújo Soares, A.K.; Moreira, L.R.; Coriolano, M.C.; de Souza Lima, G.M.; Napoleão, T.H.; de Lorena, V.M.B.; de Oliveira da Silva, L.A.; do Nascimento, S.C. L-asparaginase isolated from Streptomyces ansochromogenes promotes Th1 profile and activates CD8+ T cells in human PBMC: An in vitro investigation. J. Appl. Microbiol. 2018, 124, 1122–1130. [Google Scholar] [CrossRef]
- El-Hadi, A.A.; Ahmed, H.M.; Hamzawy, R.A. Optimization and characterization of L-asparaginase production by a novel isolated streptomyces spp. strain. Egypt. Pharm. J. 2019, 18, 111. [Google Scholar]
- El-Naggar, N.E.-A.; Deraz, S.F.; El-Ewasy, S.M.; Suddek, G.M. Purification, characterization and immunogenicity assessment of glutaminase free L-asparaginase from Streptomyces brollosae NEAE-115. BMC Pharmacol. Toxicol. 2018, 19, 51. [Google Scholar] [CrossRef]
- Mangamuri, U.; Vijayalakshmi, M.; Ganduri, V.S.R.K.; Rajulapati, S.B.; Poda, S. Extracellular L-asparaginase from Streptomyces labedae VSM-6: Isolation, production and optimization of culture conditions using RSM. Pharmacogn. J. 2017, 9. [Google Scholar] [CrossRef] [Green Version]
- Meena, B.; Anburajan, L.; Dheenan, P.S.; Begum, M.; Vinithkumar, N.V.; Dharani, G.; Kirubagaran, R. Novel glutaminase free L-asparaginase from Nocardiopsis alba NIOT-VKMA08: Production, optimization, functional and molecular characterization. Bioprocess Biosyst. Eng. 2015, 38, 373–388. [Google Scholar] [CrossRef]
- Cardoso, S.L.; de Freitas, M.M.; de Souza, P.M.; Homem-de-Mello, M.; Silveira, D.; Fonseca-Bazzo, Y.M.; Filho, E.X.; Junior, A.P.; Magalhães, P.O. Optimization of aqueous two-phase micellar system for partial purification of L-asparaginase from Penicillium sp. grown in wheat bran as agro-industrial residue. Braz. J. Microbiol. 2020, 51, 979–988. [Google Scholar] [CrossRef]
- Gonçalves, A.B.; Maia, A.C.F.; Rueda, J.A.; Vanzela, A.P.d.F.C. Fungal production of the anti-leukemic enzyme L-asparaginase: From screening to medium development. Acta Sci. Biol. Sci. 2016, 38, 387–394. [Google Scholar] [CrossRef] [Green Version]
- Chow, Y.; Ting, A.S.Y. Endophytic l-asparaginase-producing fungi from plants associated with anticancer properties. J. Adv. Res. 2015, 6, 869–876. [Google Scholar] [CrossRef] [PubMed]
- El-Gendy, M.M.A.A.; Awad, M.F.; El-Shenawy, F.S.; El-Bondkly, A.M.A. Production, purification, characterization, antioxidant and antiproliferative activities of extracellular L-asparaginase produced by Fusarium equiseti AHMF4. Saudi J. Biol. Sci. 2021, 28, 2540–2548. [Google Scholar] [CrossRef]
- Benchamin, D.; Sreejai, R.; Sujitha, S.; Jensy Roshan, F.; Albert, C.; Rishad, K. Anti-proliferative activity of L-Asparaginase enzyme from fungi on breast cancer. J Pharmacogn. Phytochem. 2019, 8, 407–410. [Google Scholar]
- Dias, F.F.G.; Ruiz, A.L.T.G.; Torre, A.D.; Sato, H.H. Purification, characterization and antiproliferative activity of l-asparaginase from Aspergillus oryzae CCT 3940 with no glutaminase activity. Asian Pac. J. Trop. Biomed. 2016, 6, 785–794. [Google Scholar] [CrossRef] [Green Version]
- Golbabaie, A.; Nouri, H.; Moghimi, H.; Khaleghian, A. L-asparaginase production and enhancement by Sarocladium strictum: In vitro evaluation of anti-cancerous properties. J. Appl. Microbiol. 2020, 129, 356–366. [Google Scholar] [CrossRef]
- Ashok, A.; Doriya, K.; Rao, J.V.; Qureshi, A.; Tiwari, A.K.; Kumar, D.S. Microbes Producing L-Asparaginase free of Glutaminase and Urease isolated from Extreme Locations of Antarctic Soil and Moss. Sci. Rep. 2019, 9, 1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correa, H.T.; Vieira, W.F.; Pinheiro, T.M.A.; Cardoso, V.L.; Silveira, E.; Sette, L.D.; Pessoa, A.; Filho, U.C. L-asparaginase and Biosurfactants Produced by Extremophile Yeasts from Antarctic Environments. Ind. Biotechnol. 2020, 16, 107–116. [Google Scholar] [CrossRef]
- Nouri, H.; Moghimi, H.; khaleghian, A. Production and characterization of biochemical properties of L-Asparaginase by indigenous yeast isolated from soil of Iran. Koomesh J. 2020, 22, 178–184. [Google Scholar] [CrossRef]
- Darvishi, F.; Faraji, N.; Shamsi, F. Production and structural modeling of a novel asparaginase in Yarrowia lipolytica. Int. J. Biol. Macromol. 2019, 125, 955–961. [Google Scholar] [CrossRef] [PubMed]
- Costa, I.M.; Schultz, L.; de Araujo Bianchi Pedra, B.; Leite, M.S.M.; Farsky, S.H.P.; de Oliveira, M.A.; Pessoa, A.; Monteiro, G. Recombinant L-asparaginase 1 from Saccharomyces cerevisiae: An allosteric enzyme with antineoplastic activity. Sci. Rep. 2016, 6, 36239. [Google Scholar] [CrossRef] [PubMed]
- Abd El Baky, H.H.; El Baroty, G.S. Optimization of Growth Conditions for Purification and Production of L-Asparaginase by Spirulina maxima. Evid. Based Complementary Altern. Med. 2016, 2016, 1785938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abd El-Baky, H.H.; El-Baroty, G.S. Spirulina maxima L-asparaginase: Immobilization, Antiviral and Antiproliferation Activities. Recent Pat. Biotechnol. 2020, 14, 154–163. [Google Scholar] [CrossRef]
- Ebrahiminezhad, A.; Rasoul-Amini, S.; Ghoshoon, M.B.; Ghasemi, Y. Chlorella vulgaris, a novel microalgal source for L-asparaginase production. Biocatal. Agric. Biotechnol. 2014, 3, 214–217. [Google Scholar] [CrossRef]
- Meghavarnam, A.K.; Janakiraman, S. Solid state fermentation: An effective fermentation strategy for the production of L-asparaginase by Fusarium culmorum (ASP-87). Biocatal. Agric. Biotechnol. 2017, 11, 124–130. [Google Scholar] [CrossRef]
- Izadpanah Qeshmi, F.; Homaei, A.; Fernandes, P.; Javadpour, S. Marine microbial L-asparaginase: Biochemistry, molecular approaches and applications in tumor therapy and in food industry. Microbiol. Res. 2018, 208, 99–112. [Google Scholar] [CrossRef]
- Datar, R. Economics of primary separation steps in relation to fermentation and genetic engineering. Process. Biochem. 1986, 21, 19–26. [Google Scholar]
- Souza, P.M.; de Freitas, M.M.; Cardoso, S.L.; Pessoa, A.; Guerra, E.N.S.; Magalhaes, P.O. Optimization and purification of L-asparaginase from fungi: A systematic review. Crit. Rev. Oncol. Hematol. 2017, 120, 194–202. [Google Scholar] [CrossRef]
- da Cunha, M.C.; Dos Santos Aguilar, J.G.; de Melo, R.R.; Nagamatsu, S.T.; Ali, F.; de Castro, R.J.S.; Sato, H.H. Fungal L-asparaginase: Strategies for production and food applications. Food Res. Int. 2019, 126, 108658. [Google Scholar] [CrossRef]
- Mukherjee, J.; Majumdar, S.; Scheper, T. Studies on nutritional and oxygen requirements for production of L-asparaginase by Enterobacter aerogenes. Appl. Microbiol. Biotechnol. 2000, 53, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Baskar, G.; Renganathan, S. Optimization of L-asparaginase production by Aspergillus terreus MTCC 1782 using response surface methodology and artificial neural network-linked genetic algorithm. Asia Pac. J. Chem. Eng. 2012, 7, 212–220. [Google Scholar] [CrossRef]
- Batool, T.; Makky, E.A.; Jalal, M.; Yusoff, M.M. A Comprehensive Review on L-Asparaginase and Its Applications. Appl. Biochem. Biotechnol. 2016, 178, 900–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nageswara, S.; Guntuku, G.; Tadimalla, P. Production of L-asparaginase by solid state fermentation using marine fungus. BioMed Res. 2014, 1, 1–9. [Google Scholar]
- Pandey, A.; Larroche, C.; Soccol, C.R. General Considerations about Solid-state Fermentation Processes. In Current Developments in Solid-state Fermentation; Pandey, A., Soccol, C.R., Larroche, C., Eds.; Springer: New York, NY, USA, 2008; pp. 13–25. [Google Scholar]
- da Cunha, M.C.; Silva, L.C.; Sato, H.H.; de Castro, R.J.S. Using response surface methodology to improve the L-asparaginase production by Aspergillus niger under solid-state fermentation. Biocatal. Agric. Biotechnol. 2018, 16, 31–36. [Google Scholar] [CrossRef]
- Kumar, N.M.; Ramasamy, R.; Manonmani, H. Production and optimization of L-asparaginase from Cladosporium sp. using agricultural residues in solid state fermentation. Ind. Crop. Products 2013, 43, 150–158. [Google Scholar] [CrossRef]
- Abusham, R.A.; Rahman, R.N.Z.R.A.; Salleh, A.B.; Basri, M. Optimization of physical factors affecting the production of thermo-stable organic solvent-tolerant protease from a newly isolated halo tolerant Bacillus subtilis strain Rand. Microb. Cell Factories 2009, 8, 20. [Google Scholar] [CrossRef] [Green Version]
- Bezerra, M.A.; Santelli, R.E.; Oliveira, E.P.; Villar, L.S.; Escaleira, L.A. Response surface methodology (RSM) as a tool for optimization in analytical chemistry. Talanta 2008, 76, 965–977. [Google Scholar] [CrossRef]
- Choi, B.; Rempala, G.A.; Kim, J.K. Beyond the Michaelis-Menten equation: Accurate and efficient estimation of enzyme kinetic parameters. Sci. Rep. 2017, 7, 17018. [Google Scholar] [CrossRef] [Green Version]
- Krishnapura, P.R.; Belur, P.D.; Subramanya, S. A critical review on properties and applications of microbial L-asparaginases. Crit. Rev. Microbiol. 2016, 42, 720–737. [Google Scholar] [CrossRef] [PubMed]
- Javed, S.; Azeem, F.; Hussain, S.; Rasul, I.; Siddique, M.H.; Riaz, M.; Afzal, M.; Kouser, A.; Nadeem, H. Bacterial lipases: A review on purification and characterization. Prog. Biophys. Mol. Biol. 2018, 132, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Sindhu, R.; Manonmani, H.K. Expression and characterization of recombinant L-asparaginase from Pseudomonas fluorescens. Protein Expr. Purif. 2018, 143, 83–91. [Google Scholar] [CrossRef]
- Susan Aishwarya, S.; Iyappan, S.; Vijaya Lakshmi, K.; Rajnish, K.N. In silico analysis, molecular cloning, expression and characterization of L-asparaginase gene from Lactobacillus reuteri DSM 20016. 3 Biotech 2017, 7, 348. [Google Scholar] [CrossRef]
- Husain, I.; Sharma, A.; Kumar, S.; Malik, F. Purification and characterization of glutaminase free asparaginase from Pseudomonas otitidis: Induce apoptosis in human leukemia MOLT-4 cells. Biochimie 2016, 121, 38–51. [Google Scholar] [CrossRef]
- Momeni, V.; Alemzadeh, I.; Vosoughi, M. Enhancement of L-asparaginase production by Candida utilis in a 13l fermenter and its purification. Int. J. Eng. Trans. B: Appl. 2015, 28, 1134–1139. [Google Scholar]
- Keramat, J.; LeBail, A.; Prost, C.; Jafari, M. Acrylamide in baking products: a review article. Food Bioprocess Technol. 2011, 4, 530–543. [Google Scholar] [CrossRef]
- Mesias, M.; Delgado-Andrade, C.; Holgado, F.; Morales, F.J. Acrylamide content in French fries prepared in households: A pilot study in Spanish homes. Food Chem. 2018, 260, 44–52. [Google Scholar] [CrossRef] [Green Version]
- Muttucumaru, N.; Powers, S.J.; Elmore, J.S.; Dodson, A.; Briddon, A.; Mottram, D.S.; Halford, N.G. Acrylamide-forming potential of potatoes grown at different locations, and the ratio of free asparagine to reducing sugars at which free asparagine becomes a limiting factor for acrylamide formation. Food Chem. 2017, 220, 76–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinci, R.M.; Mestdagh, F.; De Meulenaer, B. Acrylamide formation in fried potato products–Present and future, a critical review on mitigation strategies. Food Chem. 2012, 133, 1138–1154. [Google Scholar] [CrossRef]
- Amrein, T.M.; Bachmann, S.; Noti, A.; Biedermann, M.; Barbosa, M.F.; Biedermann-Brem, S.; Grob, K.; Keiser, A.; Realini, P.; Escher, F. Potential of acrylamide formation, sugars, and free asparagine in potatoes: A comparison of cultivars and farming systems. J. Agric. Food Chem. 2003, 51, 5556–5560. [Google Scholar] [CrossRef] [PubMed]
- Mohan Kumar, N.S.; Manonmani, H.K. Purification, characterization and kinetic properties of extracellular L-asparaginase produced by Cladosporium sp. World J. Microbiol. Biotechnol. 2013, 29, 577–587. [Google Scholar] [CrossRef]
- Shi, R.; Liu, Y.; Mu, Q.; Jiang, Z.; Yang, S. Biochemical characterization of a novel L-asparaginase from Paenibacillus barengoltzii being suitable for acrylamide reduction in potato chips and mooncakes. Int. J. Biol. Macromol. 2017, 96, 93–99. [Google Scholar] [CrossRef]
- Ehrik, B.; Maganlal, D.P.; Allen, E.V.; Gregori, F.D.; Kin-Khang, L.G.; Vu, L.; Grant, T.M. Method for Reducing Acrylamide Formation. U.S. Patent RU2423876C2, 25 March 2004. [Google Scholar]
- Sarion, C.; Codină, G.G.; Dabija, A. Acrylamide in Bakery Products: A Review on Health Risks, Legal Regulations and Strategies to Reduce Its Formation. Int. J. Environ. Res. Public Health 2021, 18, 4332. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Peters, R.J.; Van Boekel, M.A. Acrylamide and 5-hydroxymethylfurfural formation during baking of biscuits: Part I: Effects of sugar type. Food Chem. 2016, 192, 575–585. [Google Scholar] [CrossRef]
- Meghavarnam, A.K.; Janakiraman, S. Evaluation of acrylamide reduction potential of L-asparaginase from Fusarium culmorum (ASP-87) in starchy products. LWT 2018, 89, 32–37. [Google Scholar] [CrossRef]
- Matouri, M.; Alemzadeh, I. Suppressed Acrylamide Formation during Baking in Yeast-Leavened Bread Based on added Asparaginase, Baking Time and Temperature Using Response Surface Methodology. Appl. Food Biotechnol. 2018, 5, 29–36. [Google Scholar] [CrossRef]
- Budolfsen, G.; Jensen, M.T.; Heldt-Hansen, H.P.; Stringer, M.A.; Lange, L. Asparaginases and method of preparing a heat-treated product. U.S. Patent US8124396B2, 24 April 2008. [Google Scholar]
- Naidu, M.M.; Sulochanamma, G.; Sampathu, S.; Srinivas, P. Studies on extraction and antioxidant potential of green coffee. Food Chem. 2008, 107, 377–384. [Google Scholar] [CrossRef]
- Butt, M.S.; Sultan, M.T. Coffee and its consumption: Benefits and risks. Crit. Rev. Food Sci. Nutr. 2011, 51, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Ponte, S. Thelatte revolution’? Regulation, markets and consumption in the global coffee chain. World Dev. 2002, 30, 1099–1122. [Google Scholar] [CrossRef]
- Cano-Marquina, A.; Tarín, J.; Cano, A. The impact of coffee on health. Maturitas 2013, 75, 7–21. [Google Scholar] [CrossRef]
- Hall, S.; Yuen, J.W.; Grant, G.D. Bioactive constituents in caffeinated and decaffeinated coffee and their effect on the risk of depression—A comparative constituent analysis study. Beverages 2018, 4, 79. [Google Scholar] [CrossRef] [Green Version]
- Porto, A.C.V.; Freitas-Silva, O.; Souza, E.F.d.; Gottschalk, L.M.F. Effect of asparaginase enzyme in the reduction of asparagine in green coffee. Beverages 2019, 5, 32. [Google Scholar] [CrossRef] [Green Version]
- Authority, E.F.S. Update on acrylamide levels in food from monitoring years 2007 to 2010. EFSA J. 2012, 10, 2938. [Google Scholar] [CrossRef]
- Akgün, B.; Arıcı, M.; Çavuş, F.; Karataş, A.B.; Ekşi Karaağaç, H.; Uçurum, H.Ö. Application of L-asparaginase to produce high-quality Turkish coffee and the role of precursors in acrylamide formation. J. Food Process. Preserv. 2021, e15486. [Google Scholar] [CrossRef]
- Porto, A.C.V.; Freitas-Silva, O.; da Rosa, J.S.; Gottschalk, L.M.F. Estimated acrylamide intake from coffee consumption in Latin America. Am. J. Agric. Biol. Sci. 2015, 10, 91–98. [Google Scholar]
- Mesías, M.; Morales, F.J. Acrylamide in coffee: Estimation of exposure from vending machines. J. Food Compos. Anal. 2016, 48, 8–12. [Google Scholar] [CrossRef] [Green Version]
- Method for Reducing Acrylamide in Roasted Coffee Beans, Roasted Coffee Beans Having Reduced Concentration of Acrylamide, and Goods. Patent JP4467517B2, 2 February 2006.
- Rottmann, E.; Hauke, K.F.; Krings, U.; Berger, R.G. Enzymatic acrylamide mitigation in French fries—An industrial-scale case study. Food Control 2021, 123, 107739. [Google Scholar] [CrossRef]
- Medeiros Vinci, R.; Mestdagh, F.; Van Poucke, C.; Kerkaert, B.; de Muer, N.; Denon, Q.; Van Peteghem, C.; De Meulenaer, B. Implementation of Acrylamide Mitigation Strategies on Industrial Production of French Fries: Challenges and Pitfalls. J. Agric. Food Chem. 2011, 59, 898–906. [Google Scholar] [CrossRef]
- Vidya, J.; Sajitha, S.; Ushasree, M.V.; Sindhu, R.; Binod, P.; Madhavan, A.; Pandey, A. Genetic and metabolic engineering approaches for the production and delivery of L-asparaginases: An overview. Bioresour. Technol. 2017, 245, 1775–1781. [Google Scholar] [CrossRef]
- Alam, S.; Ahmad, R.; Pranaw, K.; Mishra, P.; Khare, S.K. Asparaginase conjugated magnetic nanoparticles used for reducing acrylamide formation in food model system. Bioresour. Technol. 2018, 269, 121–126. [Google Scholar] [CrossRef]
- Ravi, A.; Gurunathan, B. Acrylamide Mitigation in Fried Kochchi Kesel Chips Using Free and Immobilized Fungal Asparaginase. Food Technol. Biotechno.l 2018, 56, 51–57. [Google Scholar] [CrossRef]
- Li, R.; Zhang, Z.; Pei, X.; Xia, X. Covalent Immobilization of L-Asparaginase and Optimization of Its Enzyme Reactor for Reducing Acrylamide Formation in a Heated Food Model System. Front. Bioeng. Biotechnol. 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Vo, T.D.; Sulaiman, C.; Tafazoli, S.; Lynch, B.; Roberts, A.; Chikamatsu, G. Safety assessment of glutaminase from Aspergillus niger. Food Sci. Nutr. 2020, 8, 1433–1450. [Google Scholar] [CrossRef] [PubMed]
- Claeys, W.L.; de Vleeschouwer, K.; Hendrickx, M.E. Effect of Amino Acids on Acrylamide Formation and Elimination Kinetics. Biotechnol. Prog. 2005, 21, 1525–1530. [Google Scholar] [CrossRef] [PubMed]
- Mestdagh, F.; Maertens, J.; Cucu, T.; Delporte, K.; Van Peteghem, C.; De Meulenaer, B. Impact of additives to lower the formation of acrylamide in a potato model system through pH reduction and other mechanisms. Food Chem. 2008, 107, 26–31. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Microbe | Strain | Kind of Enzyme | Source | Reference |
|---|---|---|---|---|
| Archaea | Thermococcus kodakarensis KOD1 | plant type | cloning gene | [25] |
| Thermococcus kodakarensis KOD1 | type I | cloning gene | [40,41,42] | |
| Thermococcus zilligii AN1 TziAN1_1 | – | cloning gene | [12] | |
| Pyrobaculum calidifontis | – | cloning gene | [43] | |
| Pyrococcus yayanosii CH1 | – | cloning gene | [44] | |
| Bacteria | Acinetobacter soli Y-3 | – | cloning gene | [45] |
| Bacillus subtilis WSA3 | extracellular | marine sponges | [46] | |
| Bacillus sonorensis | – | cloning gene | [47] | |
| Bacillus licheniformis | extracellular | soil | [48] | |
| Bacillus sp. SL-1 | – | cloning gene | [6] | |
| Bacillus brevis | intracellular | soil | [49] | |
| Bacillus subtilis KDPS1 | extracellular | soil | [50] | |
| Stenotrophomonas maltophilia EMCC2297 | extracellular | soil | [38] | |
| Lactobacillus casei subsp.casei ATCC 393 | – | cloning gene | [51] | |
| Pseudomonas otitidis Enterobacter cloacae Ochrobactrum anthropi Escherichia fergusonii | periplasmic | soils and water | [52] | |
| Pseudomonas oryzihabitans | extracellular | plant | [53] | |
| Halomonas elongate IBRC-M10216 | – | cloning gene | [54] | |
| Rhizobium etli | – | cloning gene | [29] | |
| Actinomycetes | Streptomyces fradiae NEAE-82 | extracellular | soil | [55] |
| Streptomyces rochei subsp. chromatogenes NEAE-K | extracellular | soil | [35] | |
| Streptomyces ansochromogenes UFPEDA 3420 | extracellular | collection | [56,57] | |
| Streptomyces spp. | extracellular | soil | [58] | |
| Streptomyces brollosae NEAE-115 | extracellular | soil | [59] | |
| Streptomyces labedae VSM-6 | extracellular | marine sediment | [60] | |
| Nocardiopsis alba NIOT-VKMA08 | extracellular | marine sediment | [61] | |
| Fungi | Penicillium crustosum NMKA 511 | extracellular | soil | [19] |
| Penicillium sp.–encoded 2DSST1 | extracellular | soil | [62] | |
| Penicillium sp. T6.2 Fusarium sp. T22.2 | extracellular | collection | [63] | |
| Penicillium simplicissimum Dothiodeomycetes sp. Ascomycota sp. Fusarium oxysporum | extracellular | plants | [64] | |
| Fusarium equiseti AHMF4 | extracellular | soil | [65] | |
| Aspergillus oryzae CCT 3940 | extracellular | collection | [37] | |
| Aspergillus fumigatus | extracellular | plant | [66] | |
| Aspergillus oryzae CCT 3940 | extracellular | collection | [67] | |
| Sarocladium strictum | extracellular | soil | [68] | |
| Trichosporon asahii IBBLA1 | extracellular | soil and mosses | [69] | |
| Yeast | Leucosporidium muscorum CRM 1648 | intracellular | marine sediment | [39] |
| Leucosporidium scotti L120 | - | marine sediment | [70] | |
| Sarocladium sp. AG90 | extracellular | soil | [71] | |
| Yarrowia lipolytica DSM3286 | extracellular | collection | [72] | |
| Saccharomyces cerevisiae BY4741 | - | cloning gene | [73] | |
| Algae | Spirulina maxima | extracellular | collection | [74,75] |
| Chlorella vulgaris | intracellular | water and soil | [76] |
| Strains | Purification | Characterization | References | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Method | SA (IU/mg) | PF | PY (%) | Opt. pH | Opt. temp (°C) | Km (mM) | Vmax/Kcat(s−1) 1 | Mol. Wt. (kDa) (Structural Form) | ||
| Thermococcus kodakarensis | Superdex 200 10/300 GL gel column | 767 μmol/min/mg | - | - | 7.0 | 85 | 3.1 | 833 μmol/min/mg | 70 (homodimer) | [25] |
| Pyrobaculum calidifontis | Superdex 200 10/300 GL gel filtration column | - | - | - | 6.5 | at or above 100 | 4.5 | 355 μmol/min/mg | 65 (dimer) | [43] |
| Pyrococcus yayanosii CH1 | nickel affinity column | 1483.81 | 50.62 | 60.92 | 8.0 | 95 | 6.5 | 2929 μmol/min | 72.2 (homodimer) | [44] |
| Thermococcus zilligii AN1 TziAN1_1 | nickel affinity column | 5278 | - | - | 8.5 | 90 | 6.08 | 3267 | 71 (homodimer) | [12] |
| Lactobacillus casei subsp.casei ATCC 393 | nickel affinity column | 0.419 | 9.78 | 99.8 | 6 | 40 | 0.012 | 1.576 mM/min | 35 (monomer) | [51] |
| Pseudomonas fluorescens MTCC 8127 | nickel affinity column | 26 | 4.12 | 85.4 | 7.5 | 37 | 50 | 4.032 M/min | 140 (homotetramer) | [94] |
| Lactobacillus reuteri DSM 20016 | nickel affinity column | 0.63 | 10.5 | 92 | 6 | 30 | 0.3332 | 14.06 mM/min | 35 (monomer) | [95] |
| Halomonas elongate IBRC-M10216 | nickel affinity column | 1510 | 14.5 | 27 | 6–9 | 37 | 5.6 | 2.2 μmol/min | 70–80 (homodimer) | [54] |
| Bacillus brevis | sulphopropyl Sephadex column | 9.89 | 89.9 | 15 | 6.5 | 37 | 35 | 0.77 IU | 32 (monomer) | [49] |
| Bacillus subtilis sp. KDPS-1 | ammonium sulphate precipitation and DEAE column | 97.04 | 12.11 | 84.89 | 5.0 | 37 | - | - | 97.4 - | [50] |
| Pseudomonas otitidis | ammonium sulphate precipitation, DEAE-cellulose Column and Sephadex G-100 Column | 107.84 | 151.88 | 38.9 | 7.5 | 40 | - | - | 205 (homohexamer) | [96] |
| Aquabacterium sp. A7-Y (abASNase2) | nickel affinity column | 458.9 | - | - | 9.0 | 60 | 1.8 | 241.9 | 33 (monomer) | [20] |
| Penicillium crustosum NMKA 511 | ammonium sulphate precipitation, DEAE-Sephadex column and Sephadex G-100 column | 9.84 | 6.47 | 36.3 | 6.67 | 36.9 | 3.79 | 499.8 µmol/min/mg | 41.3 and 44.4 (heterodimer) | [19] |
| Aspergillus oryzae CCT 3940 | ammonium sulphate precipitation, Q Sepharose™ column, SP Sepharose™ column and CM Sepharose™ column | 282 | 28.6 | 6 | 5–8 | 40–50 | 2.10 | 35.8 U/mL | 115 - | [37,67] |
| Saccharomyces cerevisiae | nickel affinity column and PD-10 Desalting column | - | - | - | 8.6 | 40 | 0.075 2 | 0.042 μmol/min | 41.4 - | [73] |
| Candida utilis ATCC9950 | acetone precipitation and Q-sepharose column | 7853 | 10.02 | 82.7 | - | - | - | - | 40 - | [97] |
| Spirulina maxima | ammonium sulphate precipitation and Sephadex G-200 column | 19.1 | 10.91 | 86.45 | 8.5 | 37 | - | - | - - | [74] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, R.; Wan, X.; Geng, X.; Xue, D.; Xie, Z.; Chen, C. Microbial L-asparaginase for Application in Acrylamide Mitigation from Food: Current Research Status and Future Perspectives. Microorganisms 2021, 9, 1659. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9081659
Jia R, Wan X, Geng X, Xue D, Xie Z, Chen C. Microbial L-asparaginase for Application in Acrylamide Mitigation from Food: Current Research Status and Future Perspectives. Microorganisms. 2021; 9(8):1659. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9081659
Chicago/Turabian StyleJia, Ruiying, Xiao Wan, Xu Geng, Deming Xue, Zhenxing Xie, and Chaoran Chen. 2021. "Microbial L-asparaginase for Application in Acrylamide Mitigation from Food: Current Research Status and Future Perspectives" Microorganisms 9, no. 8: 1659. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9081659