
Microbial Lipopeptide-Producing Strains and Their Metabolic Roles under Anaerobic Conditions

 ,
, 
Abstract
:1. Introduction
2. Lipopeptide-Producing Strains under Anaerobic Conditions

{kind=link}
{kind=link}
{kind=link}
| Year | Source | Strain | Biosurfactant | References |
|---|---|---|---|---|
| 1980 | Unknown | Clostridium pasteurianum | Neutral lipid | [24] |
| 1985 | Oklahoma oilfield, US | Bacillus mojavensis JF-2 | Lipopeptide | [33] |
| 1995 | Oilfield, North Germany | Bacillus licheniformis BAS50 | Lichenysin A | [34] |
| 1997 | Oilfield, North Germany | Bacillus licheniformis BNP29 | Lipopeptide | [35] |
| 1997 | Oilfield, North Germany | Bacillus licheniformis BNP36 | Lipopeptide | [35] |
| 1997 | Oilfield, North Germany | Bacillus licheniformis Mep132 | Lipopeptide | [35] |
| 1997 | Oilfield, Russian | Bacillus subtilis C9 | Surfactin | [36] |
| 1999 | Marine sediments | Bacillus subtilis ATCC 21332 | Lipopeptide | [37] |
| 2000 | Noyabrskyi oilfield, Russian | Bacillus subtilis BS2202 | Lipopeptide | [38] |
| 2001 | Water buffalo | Bacillus licheniformis 26 L-10 | Lipopeptide | [39] |
| 2004 | Oklahoma oilfield, US | Bacillus subtilis ATCC 12332 | Lipopeptide | [40] |
| 2007 | Sahara desert, Tunisia | Bacillus subtilis RS-1 | Lipopeptide | [41] |
| 2007 | Oilfield, Iran | Bacillus subtilis PTCC 1365 | Lipopeptide | [42] |
| 2008 | Unknown | Bacillus subtilis ATCC6633 | Mycosubtilin, surfactin | [43] |
| 2009 | Russia | Bacillus licheniformis VKM B-511 | Licheniformin A | [44] |
| 2009 | Hot springs, US | Bacillus licheniformis TT33 | Lipopeptide | [45] |
| 2011 | Oilfield, Iran | Bacillus mojavensis GMTB-C1-2 | Lipopeptide | [25] |
| 2011 | Oilfield, US | Bacillus cereus ATCC14579 | Lipopeptide | [46] |
| 2012 | Oilfield, Brazil | Bacillus subtilis 309 | Sufactin | [47] |
| 2012 | Oilfield, Brazil | Bacillus subtilis 191 | Lipopeptide | [47] |
| 2012 | Oilfield, Brazil | Bacillus subtilis 311 | Sufactin | [47] |
| 2012 | Oilfield, Brazil | Bacillus subtilis 552 | Lipopeptide | [47] |
| 2012 | Oilfield, Brazil | Bacillus subtilis 573 | Sufactin | [47] |
| 2012 | Daqing oilfield, China | Bacillus licheniformis WJ-2 | Glycosides, Lipopeptides | [26] |
| 2012 | Soil from Ituri, Congo | Bacillusamyloliquefaciens S499 | Surfatin, Iturin and Fengycin | [48] |
| 2015 | Oilfield, Germany | Bacillus subtilis DSM 10T | Lipopeptide | [49] |
| 2017 | Daqing oilfield, China | Bacillus licheniformis DQ4 | Lipopeptide | [50] |
| 2017 | Xinjiang oilfield, China | Bacillus amyloliquefaciens 702 | Glycosides, Lipopeptides | [51] |
| 2018 | Shengli oilfield, China | Bacillus licheniformis | Glycosides, Lipopeptides | [52] |
| 2019 | Shengli oilfield, China | Bacillus tequilensis | Glycosides, Lipopeptides | [53] |
| 2021 | Xinjiang oilfield, China | Bacillus subtilis AnPL-1 | Sufactin | [54] |
| Year | Source | Strain | Biosurfactant | References |
|---|---|---|---|---|
| 1955 | Ain-ez-Zania lake, Lybia | Desulfovibrio desulfuricans sp. DSM 1926 | Unidentified | [32] |
| 1991 | Hot springs, USA | Thermoanaerobacter pseudethanolicus ATCC 33233 | Unidentified | [55] |
| 2000 | Petroleum contaminated soil | Pseudomonas sp.BS2201 | Unidentified | [56] |
| 2000 | Petroleum contaminated soil | Pseudomonas sp. BS2203 | Unidentified | [56] |
| 2000 | Oilfield, India | Pseudomonas aeruginosa ATCC 10145 | Rhamnolipid | [57] |
| 2002 | Oilfield, Germany | Isolate Glc2 | Unidentified | [58] |
| 2007 | Antarctic soil | Pantoa A-13 | Rhamnolipid | [59] |
| 2007 | Hospital Wastewater | Pseudomonas aeruginosa PAO1 | Rhamnolipid | [60] |
| 2008 | Shengli oilfield, China | Pseudomonas aeruginosa SH6 | Rhamnolipid | [61] |
| 2010 | Municipal Sewage Sludge | Pseudomonas aeruginosa ANBIOSURF-1 | Rhamnolipid | [62] |
| 2012 | Oilfield; Veracruz, Mexico | Thermoanaerobacter sp. | Unidentified | [63] |
| 2012 | Menggulin oilfield, China | Pseudomonas aeruginosa WJ-1 | Rhamnolipid | [64] |
| 2013 | Soil samples, Iowa, USA | Pseudomonas aeruginosa E03-40 | Rhamnolipid | [65] |
| 2013 | Oil contaminated soil, Iran | Enterobacter cloacae | Unidentified | [66] |
| 2014 | Gachsaran oilfield, Iran | Bacillusstearothermophilus SUCPM#14 | Unidentified | [67] |
| 2015 | Xinjiang oilfield, China | Pseudomonas aeruginosa SG | Rhamnolipid | [68,69,70] |
| 2017 | Xinjiang oilfield, China | Pseudomonas aeruginosa 709 | Rhamnolipid | [60] |
| 2018 | Daqing oilfield, China | Pseudomonas aeruginosa DQ3 | Rhamnolipid | [71] |
| 2018 | Daqing oilfield, China | Pseudomonas aeruginosa DQ1 | Rhamnolipid | [71] |
| 2018 | Daqing oilfield, China | Pseudomonas aeruginosa DQ5 | Rhamnolipid | [71] |
| 2018 | Daqing oilfield, China | Pseudomonas aeruginosa DQ6 | Rhamnolipid | [71] |
| 2020 | Dagang oilfield, China | Bacillus licheniformis DM-1 | Exopolysaccharide | [72] |
3. The Mechanism of Anaerobic Growth of Bacillus subtilis
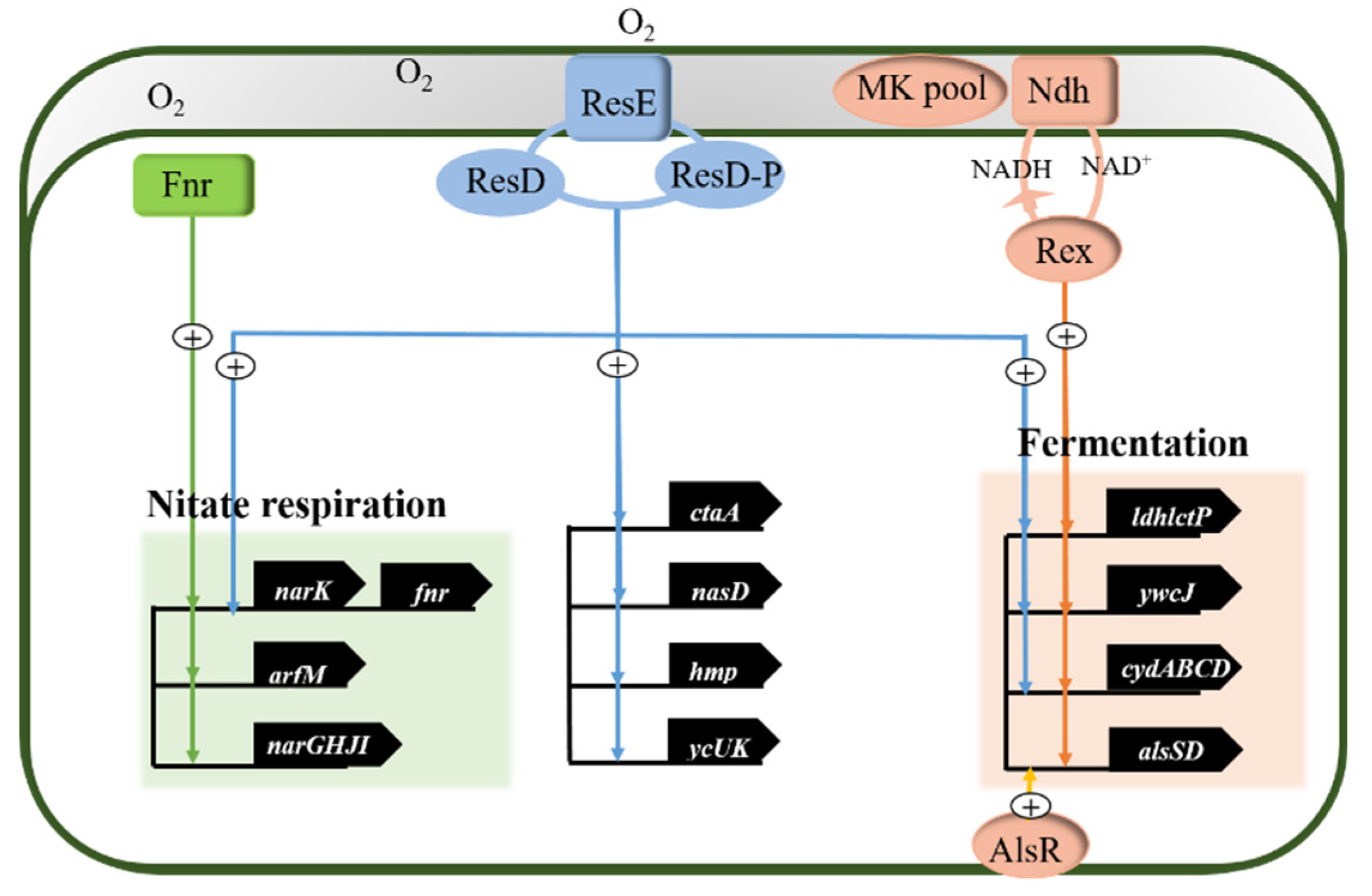
3.1. Anaerobic Regulatory Network of Bacillus subtilis
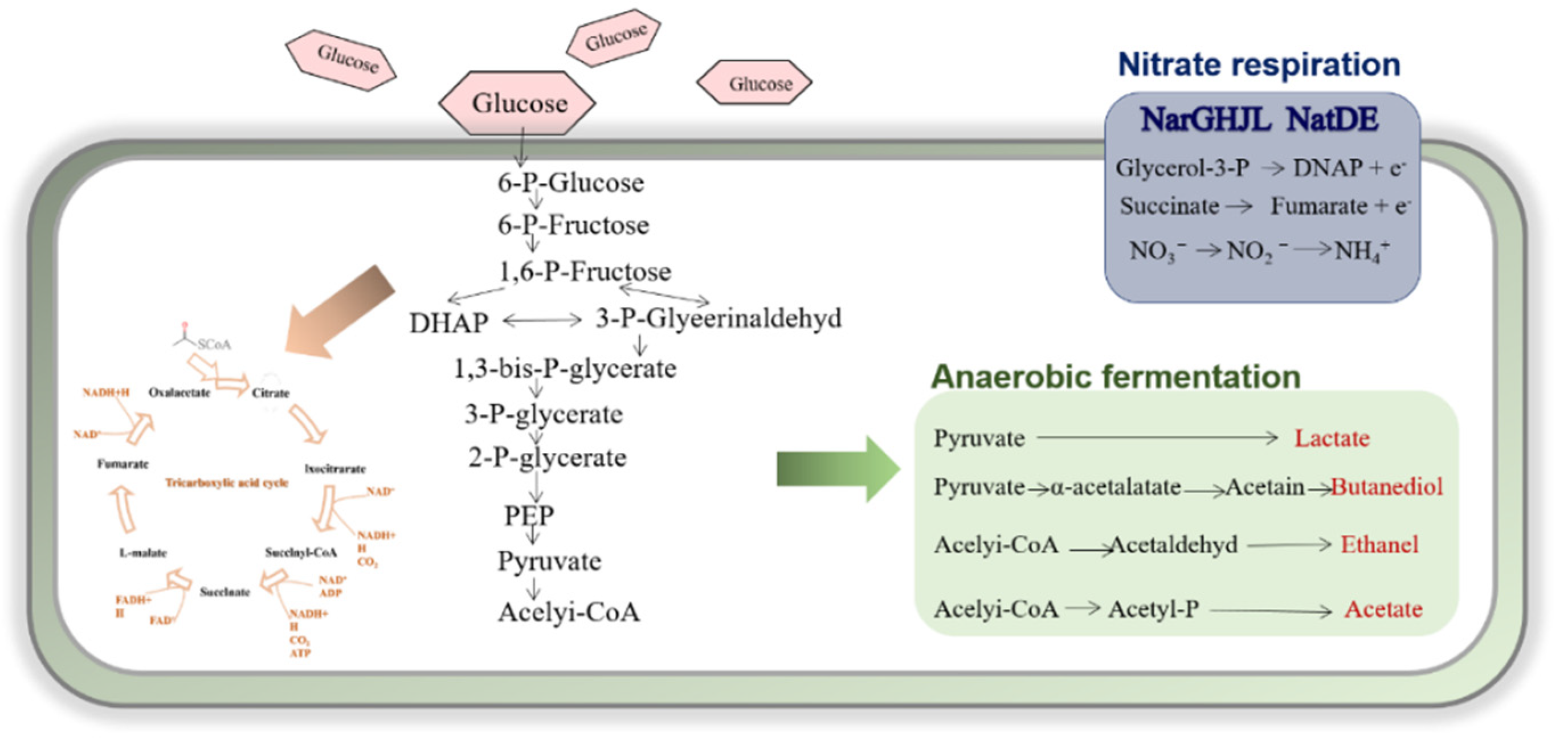
3.2. Anaerobic Energy Metabolism of Bacillus subtilis
4. Non-Ribosomal Peptide Synthetases (NRPSs) of Surfactin
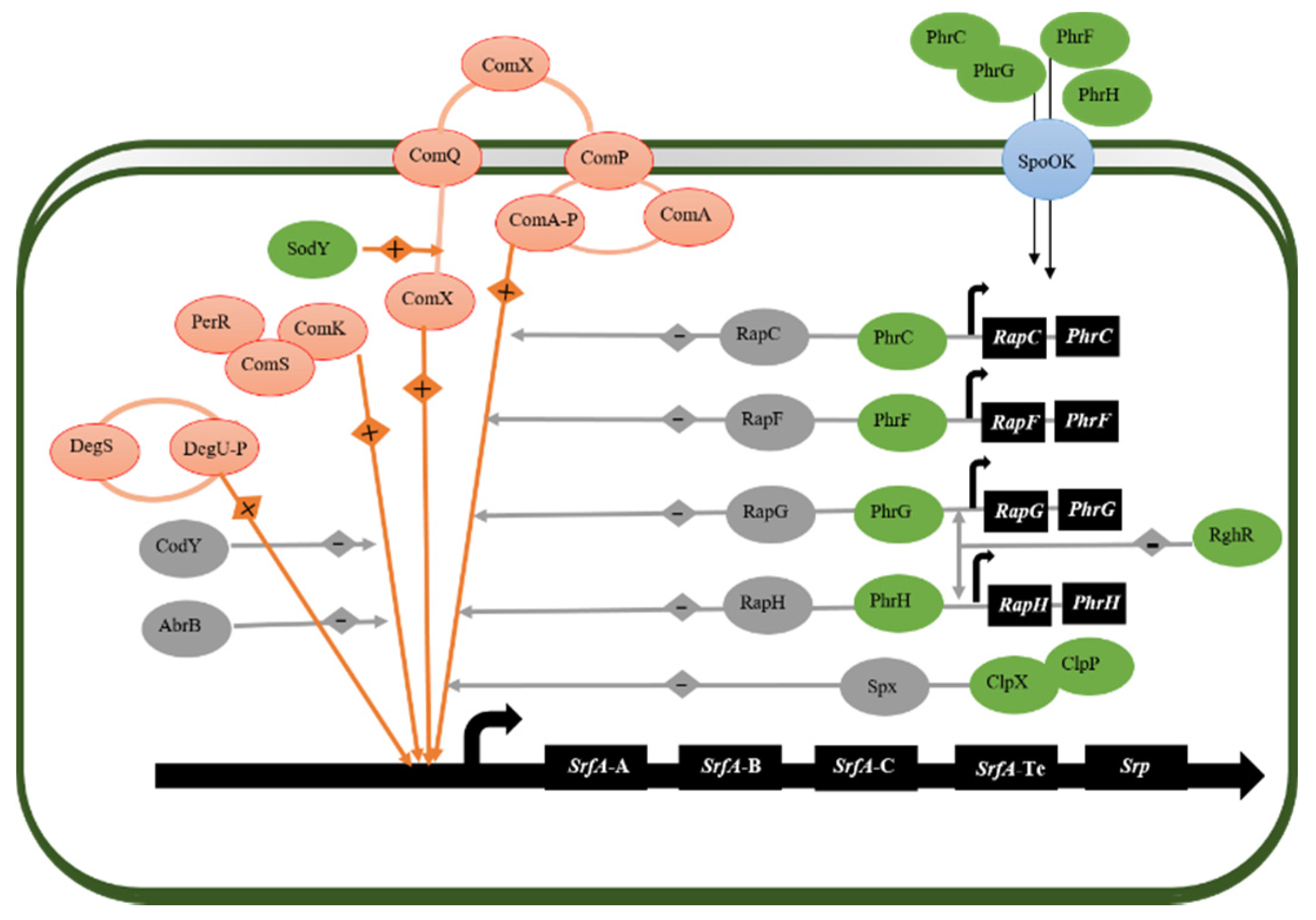
4.1. Regulator of Surfactin Synthesis in Bacillus subtilis
4.2. Non-Ribosomal Peptide Sythetases (NRPSs)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Varjani, S.J.; Upasani, V.N. Critical Review on Biosurfactant Analysis, Purification and Characterization Using Rhamnolipid as a Model Biosurfactant. Bioresour. Technol. 2017, 232, 389–397. [Google Scholar]
- Liu, J.F.; Mbadinga, S.M.; Yang, S.Z.; Gu, J.D.; Mu, B.Z. Chemical Structure, Property and Potential Applications of Biosurfactants Produced by Bacillus subtilis in Petroleum Recovery and Spill Mitigation. Int. J. Mol. Sci. 2015, 16, 4814–4837. [Google Scholar] [CrossRef] [PubMed]
- Santos, V.S.V.; Silveira, E.; Pereira, B.B. Toxicity and Applications of Surfactin for Health and Environmental Biotechnology. J. Toxicol. Environ. Health B Crit. Rev. 2018, 21, 382–399. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Fan, T.; Chen, J.; Su, J.; Zhi, X.; Pan, P.; Zou, L.; Zhang, Q. Magnetic Bioinspired Micro/Nanostructured Composite Scaffold for Bone Regeneration. Colloids Surf. B Biointerfaces 2019, 174, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Ke, C.Y.; Lu, G.M.; Wei, Y.L.; Sun, W.J.; Hui, J.F.; Zheng, X.Y.; Zhang, Q.Z.; Zhang, X.L. Biodegradation of Crude Oil by Chelatococcus Daeguensis Hb-4 and Its Potential for Microbial Enhanced Oil Recovery (Meor) in Heavy Oil Reservoirs. Bioresour. Technol. 2019, 287, 121442. [Google Scholar] [CrossRef] [PubMed]
- Kalogerakis, N.; Fava, F.; Corvini, P.F. Bioremediation Advances. N Biotechnol. 2017, 38, 41–42. [Google Scholar] [CrossRef] [PubMed]
- Karas, J.A.; Carter, G.P.; Howden, B.P.; Turner, A.M.; Paulin, O.K.A.; Swarbrick, J.D.; Baker, M.A.; Li, J.; Velkov, T. Structure-Activity Relationships of Daptomycin Lipopeptides. J. Med. Chem. 2020, 63, 13266–13290. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; De Bruijn, I.; Nybroe, O.; Ongena, M. Natural Functions of Lipopeptides from Bacillus and Pseudomonas: More Than Surfactants and Antibiotics. FEMS Microbiol. Rev. 2010, 34, 1037–1062. [Google Scholar] [CrossRef] [Green Version]
- Vašíček, O.; Hájek, J.; Bláhová, L.; Hrouzek, P.; Babica, P.; Kubala, L.; Šindlerová, L. Cyanobacterial Lipopeptides Puwainaphycins and Minutissamides Induce Disruptive and Pro-Inflammatory Processes in Caco-2 Human Intestinal Barrier Model. Harmful Algae 2020, 96, 101849. [Google Scholar] [CrossRef]
- Kozakai, R.; Ono, T.; Hoshino, S.; Takahashi, H.; Katsuyama, Y.; Sugai, Y.; Ozaki, T.; Teramoto, K.; Teramoto, K.; Tanaka, K.; et al. Acyltransferase That Catalyses the Condensation of Polyketide and Peptide Moieties of Goadvionin Hybrid Lipopeptides. Nat. Chem. 2020, 12, 869–877. [Google Scholar] [CrossRef]
- Zhao, P.; Xue, Y.; Li, X.; Li, J.; Zhao, Z.; Quan, C.; Gao, W.; Zu, X.; Bai, X.; Feng, S. Fungi-Derived Lipopeptide Antibiotics Developed since 2000. Peptides 2019, 113, 52–65. [Google Scholar] [CrossRef] [PubMed]
- Mnif, I.; Ghribi, D. Review Lipopeptides Biosurfactants: Mean Classes and New Insights for Industrial, Biomedical, and Environmental Applications. Biopolymers 2015, 104, 129–147. [Google Scholar] [CrossRef] [PubMed]
- Walton, R.B.; Woodruff, H.B. A Crystalline Antifungal Agent, Mycosubtilin, Isolated from Subtilin Broth. J. Clin. Investig. 1949, 28, 924–926. [Google Scholar] [CrossRef]
- Nishikiori, T.; Naganawa, H.; Muraoka, Y.; Aoyagi, T.; Umezawa, H. Plipastatins: New Inhibitors of Phospholipase A2, Produced by Bacillus Cereus Bmg302-Ff67. J. Antibiot. 1968, 39, 745–754. [Google Scholar] [CrossRef] [Green Version]
- Fei, D.; Liu, F.F.; Gang, H.Z.; Liu, J.F.; Yang, S.Z.; Ye, R.Q.; Mu, B.Z. A New Member of the Surfactin Family Produced by Bacillus subtilis with Low Toxicity on Erythrocyte. Process. Biochem. 2020, 94, 164–171. [Google Scholar] [CrossRef]
- Liu, X.Y.; Yang, S.Z.; Mu, B.Z. Isolation and Characterization of a C12-Lipopeptide Produced by Bacillus subtilis Hso 121. J. Pept. Sci. 2008, 14, 864–875. [Google Scholar] [CrossRef]
- Li, Y.; Zou, A.H.; Ye, R.Q.; Mu, B.Z. Counterion-Induced Changes to the Micellization of Surfactin-C16 Aqueous Solution. J. Phys. Chem. B 2009, 113, 15272–15277. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Yang, S.Z.; Mu, B.Z. Production and Characterization of a C15-Surfactin-O-Methyl Ester by a Lipopeptide Producing Strain Bacillus subtilis Hso121. Process. Biochem. 2009, 44, 1144–1151. [Google Scholar] [CrossRef]
- Arima, K.; Kakinuma, A.; Tamura, G. Surfactin, a Crystalline Peptidelipid Surfactant Produced by Bacillus subtilis: Isolation, Characterization and Its Inhibition of Fibrin Clot Formation. Biochem. Biophys. Res. Commun. 1968, 31, 488–494. [Google Scholar] [CrossRef]
- Zou, A.; Liu, J.; Garamus, V.M.; Zheng, K.; Willumeit, R.; Mu, B. Interaction between the Natural Lipopeptide [Glu1, Asp5)] Surfactin-C15 and Hemoglobin in Aqueous Solution. Biomacromolecules 2010, 11, 593–599. [Google Scholar] [CrossRef]
- Mandal, S.M.; Sharma, S.; Pinnaka, A.K.; Kumari, A.; Korpole, S. Isolation and Characterization of Diverse Antimicrobial Lipopeptides Produced by Citrobacter and Enterobacter. BMC Microbiol. 2013, 13, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veloo, A.C.M.; Tokman, H.B.; Jean-Pierre, H.; Dumont, Y.; Jeverica, S.; Lienhard, R.; Novak, A.; Rodloff, A.; Rotimi, V.; Wybo, I.; et al. Antimicrobial Susceptibility Profiles of Anaerobic Bacteria, Isolated from Human Clinical Specimens, within Different European and Surrounding Countries. A Joint Esgai Study. Anaerobe 2020, 61, 102111. [Google Scholar] [CrossRef] [PubMed]
- Kivistö, A.T.; Karp, M.T. Halophilic Anaerobic Fermentative Bacteria. J. Biotechnol. 2011, 152, 114–124. [Google Scholar] [CrossRef]
- Cooper, D.; Zajic, J.; Gerson, D.; Manninen, K. Isolation and Identification of Biosurfactants Produced During Anaerobic Growth of Clostridium pasteurianum. J. Ferment. Technol. 1980, 58, 83–86. [Google Scholar]
- Ghojavand, H.; Vahabzadeh, F.; Azizmohseni, F. A Halotolerant, Thermotolerant, and Facultative Biosurfactant Producer: Identification and Molecular Characterization of a Bacterium and Evolution of Emulsifier Stability of a Lipopeptide Biosurfactant. Biotechnol. Bioprocess Eng. 2011, 16, 72–80. [Google Scholar] [CrossRef]
- Xia, W.J.; Dong, H.P.; Yu, L. Oil-Degrading Characterization of Thermophilic and Halotolerant Strain Geobacillus Sp. Wj-2. J. Cent. South Univ. (Sci. Technol.) 2012, 43, 8–16. [Google Scholar]
- Xia, W.J.; Yu, L.; Wang, P.; Xiu, J.L.; Dong, H.P. Characterization of a Thermophilic and Halotolerant Geobacillus Pallidus H9 and Its Application in Microbial Enhanced Oil Recovery (Meor). Ann. Microbiol. 2012, 62, 1779–1789. [Google Scholar]
- Denger, K.; Schink, B. New Halo-and Thermotolerant Fermenting Bacteria Producing Surface-Active Compounds. Appl. Microbiol. Biotechnol. 1995, 44, 161–166. [Google Scholar] [CrossRef]
- Zheng, C.; Yu, L.; Huang, L.; Xiu, J.; Huang, Z. Investigation of a Hydrocarbon-Degrading Strain, Rhodococcus Ruber Z25, for the Potential of Microbial Enhanced Oil Recovery. J. Pet. Sci. Eng. 2012, 81, 49–56. [Google Scholar] [CrossRef]
- Tan, H.Q.; Wu, X.Y.; Zhang, X.Q.; Wu, M.; Zhu, X.F. Tepidibacter mesophilus Sp. Nov., a Mesophilic Fermentative Anaerobe Isolated from Soil Polluted by Crude Oil, and Emended Description of the Genus Tepidibacter. Int. J. Syst. Evol. Microbiol. 2012, 62, 66–70. [Google Scholar] [CrossRef]
- Ke, C.Y.; Sun, W.J.; Li, Y.B.; Lu, G.M.; Zhang, Q.Z.; Zhang, X.L. Microbial Enhanced Oil Recovery in Baolige Oilfield Using an Indigenous Facultative Anaerobic Strain Luteimonas Huabeiensis Sp. Nov. J. Pet. Sci. Eng. 2018, 167, 160–167. [Google Scholar] [CrossRef]
- La Riviere, J. The Production of Surface Active Compounds by Micro-Organisms and Its Possible Significance in Oil Recovery. Antonie Van Leeuwenhoek 1955, 21, 9–27. [Google Scholar] [CrossRef] [PubMed]
- Javaheri, M.; Jenneman, G.E.; McInerney, M.J.; Knapp, R.M. Anaerobic Production of a Biosurfactant by Bacillus licheniformis Jf-2. Appl. Environ. Microbiol. 1985, 50, 698–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yakimov, M.M.; Timmis, K.N.; Wray, V.; Fredrickson, H.L. Characterization of a New Lipopeptide Surfactant Produced by Thermotolerant and Halotolerant Subsurface Bacillus licheniformis Bas50. Appl. Environ. Microbiol. 1995, 61, 1706–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yakimov, M.M.; Amro, M.M.; Bock, M.; Boseker, K.; Fredrickson, H.L.; Kessel, D.G.; Timmis, K.N. The Potential of Bacillus licheniformis Strains for in Situ Enhanced Oil Recovery. J. Pet. Sci. Eng. 1997, 18, 147–160. [Google Scholar] [CrossRef]
- Kim, H.S.; Yoon, B.D.; Lee, C.H.; Suh, H.H.; Oh, H.M.; Katsuragi, T.; Tani, Y. Production and Properties of a Lipopeptide Biosurfactant from Bacillus subtilis C9. J. Ferment. Bioeng. 1997, 84, 41–46. [Google Scholar] [CrossRef]
- Davis, D.; Lynch, H.; Varley, J. The Production of Surfactin in Batch Culture by Bacillus subtilis Atcc 21332 Is Strongly Influenced by the Conditions of Nitrogen Metabolism. Enzym. Microb. Technol. 1999, 25, 322–329. [Google Scholar] [CrossRef]
- Grishchenkov, V.; Townsend, R.; McDonald, T.; Autenrieth, R.; Bonner, J.; Boronin, A. Degradation of Petroleum Hydrocarbons by Facultative Anaerobic Bacteria under Aerobic and Anaerobic Conditions. Process. Biochem. 2000, 35, 889–896. [Google Scholar] [CrossRef]
- Pattnaik, P.; Kaushik, J.; Grover, S.; Batish, V. Purification and Characterization of a Bacteriocin-Like Compound (Lichenin) Produced Anaerobically by Bacillus licheniformis Isolated from Water Buffalo. J. Appl. Microbiol. 2001, 91, 636–645. [Google Scholar] [CrossRef]
- Wei, Y.H.; Lai, C.C.; Chang, J.S. Using Taguchi Experimental Design Methods to Optimize Trace Element Composition for Enhanced Surfactin Production by Bacillus subtilis Atcc 21332. Process. Biochem. 2007, 42, 40–45. [Google Scholar] [CrossRef]
- Youssef, N.; Simpson, D.; Duncan, K.; McInerney, M.; Folmsbee, M.; Fincher, T.; Knapp, R. In Situ Biosurfactant Production by Bacillus Strains Injected into a Limestone Petroleum Reservoir. Appl. Environ. Microbiol. 2007, 73, 1239–1247. [Google Scholar] [CrossRef] [Green Version]
- Soudmand-asli, A.; Ayatollahi, S.S.; Mohabatkar, H.; Zareie, M.; Shariatpanahi, S.F. The in Situ Microbial Enhanced Oil Recovery in Fractured Porous Media. J. Pet. Sci. Eng. 2007, 58, 161–172. [Google Scholar] [CrossRef]
- Guez, J.-S.; Müller, C.; Danze, P.; Büchs, J.; Jacques, P. Respiration Activity Monitoring System (Ramos), an Efficient Tool to Study the Influence of the Oxygen Transfer Rate on the Synthesis of Lipopeptide by Bacillus subtilis Atcc6633. J. Biotechnol. 2008, 134, 121–126. [Google Scholar] [CrossRef]
- Gogotov, I.; Miroshnikov, A. The Influence of Growth Medium Composition and Physicochemical Factors on Biosurfactant Production by the Bacterium Bacillus licheniformis Vkm B-511. Appl. Biochem. Microbiol. 2009, 45, 588–592. [Google Scholar] [CrossRef]
- Suthar, H.; Hingurao, K.; Desai, A.; Nerurkar, A. Selective Plugging Strategy-Based Microbial-Enhanced Oil Recovery Using Bacillus licheniformis Tt33. J. Microbiol. Biotechnol. 2009, 19, 1230–1237. [Google Scholar]
- Senouci-Rezkallah, K.; Schmitt, P.; Jobin, M.P. Amino Acids Improve Acid Tolerance and Internal Ph Maintenance in Bacillus Cereus Atcc14579 Strain. Food Microbiol. 2011, 28, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Gudiña, E.J.; Pereira, J.F.; Rodrigues, L.R.; Coutinho, J.A.; Teixeira, J.A. Isolation and Study of Microorganisms from Oil Samples for Application in Microbial Enhanced Oil Recovery. Int. Biodeterior. Biodegrad. 2012, 68, 56–64. [Google Scholar]
- Nihorimbere, V.; Cawoy, H.; Seyer, A.; Brunelle, A.; Thonart, P.; Ongena, M. Impact of Rhizosphere Factors on Cyclic Lipopeptide Signature from the Plant Beneficial Strain Bacillus Amyloliquefaciens S499. FEMS Microbiol. Ecol. 2012, 79, 176–191. [Google Scholar] [CrossRef] [PubMed]
- Willenbacher, J.; Rau, J.T.; Rogalla, J.; Syldatk, C.; Hausmann, R. Foam-Free Production of Surfactin Via Anaerobic Fermentation of Bacillus subtilis Dsm 10(T). AMB Express 2015, 5, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, F.; Shi, R.; Cui, Q.; Han, S.; Dong, H.; Zhang, Y. Biosurfactant Production under Diverse Conditions by Two Kinds of Biosurfactant-Producing Bacteria for Microbial Enhanced Oil Recovery. J. Pet. Sci. Eng. 2017, 157, 124–130. [Google Scholar] [CrossRef]
- Li, Z.; Lin, Z.; Qian, Q.; Feng, Y.; Wang, L.; Wang, W.; Li, X. Anaerobic Growth and Metabolism of a Bacillus Licheniformis for Oil Production. J. China Univ. Pet. (Nat. Sci. Ed.) 2018, 42, 179–184. [Google Scholar]
- Lin, J.Z.; Feng, Y.; Tan, X.M.; Wang, J.; Wang, W.D.; Li, X.M. Study on Anaerobic Growth and Metabolism of a Strain of Bacillus Thermophilus for Oil Production. J. Xi’an Univ. Pet. 2019, 6, 115–119. [Google Scholar]
- Domingues, P.M.; Almeida, A.; Serafim Leal, L.; Gomes, N.C.M.; Cunha, Â. Bacterial Production of Biosurfactants under Microaerobic and Anaerobic Conditions. Rev. Environ. Sci. Bio/Technol. 2017, 16, 239–272. [Google Scholar] [CrossRef]
- Zhao, F.; Zhu, H.; Cui, Q.; Wang, B.; Su, H.; Zhang, Y. Anaerobic Production of Surfactin by a New Bacillus subtilis Isolate and the in Situ Emulsification and Viscosity Reduction Effect Towards Enhanced Oil Recovery Applications. J. Pet. Sci. Eng. 2021, 201, 108508. [Google Scholar] [CrossRef]
- Yen, T.; Park, J.; Lee, K.; Li, Y. Fate of Surfactant Vesicles Surviving from Thermophilic, Halotolerant, Spore Forming, Clostridium thermohydrosulfuricum. Microb. Enhanc. Oil Recovery 1991, 31, 297–309. [Google Scholar]
- Chayabutra, C.; Ju, L.K. Degradation of N-Hexadecane and Its Metabolites by Pseudomonas Aeruginosa under Microaerobic and Anaerobic Denitrifying Conditions. Appl. Environ. Microbiol. 2000, 66, 493–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chayabutra, C.; Wu, J.; Ju, L.K. Rhamnolipid Production by Pseudomonas Aeruginosa under Denitrification: Effects of Limiting Nutrients and Carbon Substrates. Biotechnol. Bioeng. 2001, 72, 25–33. [Google Scholar] [CrossRef]
- Denger, K.; Warthmann, R.; Ludwig, W.; Schink, B. Anaerophaga Thermohalophila Gen. Nov., Sp. Nov., a Moderately Thermohalophilic, Strictly Anaerobic Fermentative Bacterium. Int. J. Syst. Evol. Microbiol. 2002, 52, 173–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasileva-Tonkova, E.; Gesheva, V. Biosurfactant Production by Antarctic Facultative Anaerobe Pantoea Sp. During Growth on Hydrocarbons. Curr. Microbiol. 2007, 54, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Nozawa, T.; Tanikawa, T.; Hasegawa, H.; Takahashi, C.; Ando, Y.; Matsushita, M.; Nakagawa, Y.; Matsuyama, T. Rhamnolipid-Dependent Spreading Growth of Pseudomonas aeruginosa on a High-Agar Medium: Marked Enhancement under Co2-Rich Anaerobic Conditions. Microbiol. Immunol. 2007, 51, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Hao, D.H.; Lin, J.Q.; Song, X.; Lin, J.Q.; Su, Y.J.; Qu, Y.B. Isolation, Identification, and Performance Studies of a Novel Paraffin-Degrading Bacterium of Gordonia Amicalis Lh3. Biotechnol. Bioprocess Eng. 2008, 13, 61–68. [Google Scholar] [CrossRef]
- Albino, J.D.; Nambi, I.M. Partial Characterization of Biosurfactant Produced under Anaerobic Conditions by Pseudomonas Sp Anbiosurf-1. Adv. Mater. Res. 2010, 93, 623–626. [Google Scholar] [CrossRef]
- Cortés, G.C.; Zapata PeAsco, I.; Carrillo, T.R.; Reyes Avila, J.; Mayol Castillo, M.; Vargas, S.R.; Lora, P.O. Evaluation of Indigenous Anaerobic Microorganisms from Mexican Carbonate Reservoirs with Potential Meor Application. J. Pet. Sci. Eng. 2012, 81, 86–93. [Google Scholar] [CrossRef]
- Xia, W.J.; Luo, Z.B.; Dong, H.P.; Yu, L.; Cui, Q.F.; Bi, Y.Q. Synthesis, Characterization, and Oil Recovery Application of Biosurfactant Produced by Indigenous Pseudomonas Aeruginosa Wj-1 Using Waste Vegetable Oils. Appl. Biochem. Biotechnol. 2012, 166, 1148–1166. [Google Scholar] [CrossRef]
- Pinzon, N.M.; Cook, A.G.; Ju, L.K. Continuous Rhamnolipid Production Using Denitrifying Pseudomonas Aeruginosa Cells in Hollow-Fiber Bioreactor. Biotechnol. Prog. 2013, 29, 352–358. [Google Scholar] [CrossRef]
- Sarafzadeh, P.; Hezave, A.Z.; Ravanbakhsh, M.; Niazi, A.; Ayatollahi, S. Enterobacter Cloacae as Biosurfactant Producing Bacterium: Differentiating Its Effects on Interfacial Tension and Wettability Alteration Mechanisms for Oil Recovery During Meor Process. Colloids Surf. B Biointerfaces 2013, 105, 223–229. [Google Scholar] [CrossRef]
- Zargari, S.; Ramezani, A.; Ostvar, S.; Rezaei, R.; Niazi, A.; Ayatollahi, S. Isolation and Characterization of Gram-Positive Biosurfactant-Producing Halothermophilic Bacilli from Iranian Petroleum Reservoirs. Jundishapur J. Microbiol. 2014, 7, 10981–10991. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Mandlaa, M.; Hao, J.; Liang, X.; Shi, R.; Han, S.; Zhang, Y. Optimization of Culture Medium for Anaerobic Production of Rhamnolipid by Recombinant Pseudomonas Stutzeri Rhl for Microbial Enhanced Oil Recovery. Lett. Appl. Microbiol. 2014, 59, 231–237. [Google Scholar] [CrossRef]
- Zhao, F.; Cui, Q.; Han, S.; Dong, H.; Zhang, J.; Ma, F.; Zhang, Y. Enhanced Rhamnolipid Production of Pseudomonas Aeruginosa Sg by Increasing Copy Number of Rhlab Genes with Modified Promoter. RSC Adv. 2015, 5, 70546–70552. [Google Scholar] [CrossRef]
- Zhao, F.; Ma, F.; Shi, R.; Zhang, J.; Han, S.; Zhang, Y. Production of Rhamnolipids by Pseudomonas Aeruginosa Is Inhibited by H2s but Resumes in a Co-Culture with P. Stutzeri: Applications for Microbial Enhanced Oil Recovery. Biotechnol. Lett. 2015, 37, 1803–1808. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Li, P.; Guo, C.; Shi, R.J.; Zhang, Y. Bioaugmentation of Oil Reservoir Indigenous Pseudomonas Aeruginosa to Enhance Oil Recovery through in-Situ Biosurfactant Production without Air Injection. Bioresour. Technol. 2018, 251, 295–302. [Google Scholar] [CrossRef]
- Fan, Y.; Wang, J.; Gao, C.; Zhang, Y.; Du, W. A Novel Exopolysaccharide-Producing and Long-Chain N-Alkane Degrading Bacterium Bacillus licheniformis Strain Dm-1 with Potential Application for in-Situ Enhanced Oil Recovery. Sci. Rep. 2020, 10, 8519. [Google Scholar] [CrossRef] [PubMed]
- Riemenschneider, C.; Zerr, W.; Vater, N.; Brunn, H.; Mohring, S.A.I.; Hamscher, G. Analysis and Behavior of Colistin During Anaerobic Fermentation. Bioresour. Technol. 2014, 170, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Bulot, S.; Audebert, S.; Pieulle, L.; Seduk, F.; Baudelet, E.; Espinosa, L.; Pizay, M.C.; Camoin, L.; Magalon, A. Clustering as a Means to Control Nitrate Respiration Efficiency and Toxicity in Escherichia coli. mBio 2019, 10, e01832-01819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlson, H.K.; Lui, L.M.; Price, M.N.; Kazakov, A.E.; Carr, A.V.; Kuehl, J.V.; Owens, T.K.; Nielsen, T.; Arkin, A.P.; Deutschbauer, A.M. Selective Carbon Sources Influence the End Products of Microbial Nitrate Respiration. ISME 2020, 14, 2034–2045. [Google Scholar] [CrossRef] [PubMed]
- Nakano, M.M.; Dailly, Y.P.; Zuber, P.; Clark, D.P. Characterization of Anaerobic Fermentative Growth of Bacillus subtilis: Identification of Fermentation End Products and Genes Required for Growth. J. Bacteriol. 1997, 179, 6749–6755. [Google Scholar] [CrossRef] [Green Version]
- Clements, L.D.; Streips, U.N.; Miller, B.S. Differential Proteomic Analysis of Bacillus subtilis Nitrate Respiration and Fermentation in Defined Medium. Proteomics 2002, 2, 1724–1734. [Google Scholar] [CrossRef]
- Nakano, M.M.; Zuber, P. Anaerobic Growth of a "Strict Aerobe" (Bacillus subtilis). Annu. Rev. Microbiol. 1998, 52, 165–190. [Google Scholar] [CrossRef]
- Geng, H.; Zhu, Y.; Mullen, K.; Zuber, C.S.; Nakano, M.M. Characterization of Resde-Dependent Fnr Transcription in Bacillus subtilis. J. Bacteriol. 2007, 189, 1745–1755. [Google Scholar] [CrossRef] [Green Version]
- Whiteley, M.; Diggle, S.P.; Greenberg, E.P. Progress in and Promise of Bacterial Quorum Sensing Research. Nature 2017, 551, 313–320. [Google Scholar] [CrossRef]
- Härtig, E.; Jahn, D. Regulation of the Anaerobic Metabolism in Bacillus subtilis. Adv. Microb. Physiol. 2012, 61, 195–216. [Google Scholar] [PubMed]
- Ogawa, K.; Akagawa, E.; Yamane, K.; Sun, Z.W.; LaCelle, M.; Zuber, P.; Nakano, M.M. The Nasb Operon and Nasa Gene Are Required for Nitrate/Nitrite Assimilation in Bacillus subtilis. J. Bacteriol. 1995, 177, 1409–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marino, M.; Ramos, H.C.; Hoffmann, T.; Glaser, P.; Jahn, D. Modulation of Anaerobic Energy Metabolism of Bacillus subtilis by Arfm (Ywid). J. Bacteriol. 2001, 183, 6815–6821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puri-Taneja, A.; Schau, M.; Chen, Y.; Hulett, F.M. Regulators of the Bacillus subtilis Cydabcd Operon: Identification of a Negative Regulator, Ccpa, and a Positive Regulator, Resd. J. Bacteriol. 2007, 189, 3348–3358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gyan, S.; Shiohira, Y.; Sato, I.; Takeuchi, M.; Sato, T. Regulatory Loop between Redox Sensing of the Nadh/Nad(+) Ratio by Rex (Ydih) and Oxidation of Nadh by Nadh Dehydrogenase Ndh in Bacillus subtilis. J. Bacteriol. 2006, 188, 7062–7071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laouami, S.; Clair, G.; Armengaud, J.; Duport, C. Proteomic Evidences for Rex Regulation of Metabolism in Toxin-Producing Bacillus Cereus Atcc 14579. PLoS ONE 2014, 9, 107354–107363. [Google Scholar] [CrossRef] [Green Version]
- Sommer, B.; von Moeller, H.; Haack, M.; Qoura, F.; Langner, C.; Bourenkov, G.; Garbe, D.; Loll, B.; Brück, T. Detailed Structure-Function Correlations of Bacillus subtilis Acetolactate Synthase. Chembiochem 2015, 16, 110–118. [Google Scholar] [CrossRef]
- Atsumi, S.; Li, Z.; Liao, J.C. Acetolactate Synthase from Bacillus subtilis Serves as a 2-Ketoisovalerate Decarboxylase for Isobutanol Biosynthesis in Escherichia coli. Appl. Environ. Microbiol. 2009, 75, 6306–6311. [Google Scholar] [CrossRef] [Green Version]
- Huo, Y.; Zhan, Y.; Wang, Q.; Li, S.; Yang, S.; Nomura, C.T.; Wang, C.; Chen, S. Acetolactate Synthase (Alss) in Bacillus licheniformis Wx-02: Enzymatic Properties and Efficient Functions for Acetoin/Butanediol and L-Valine Biosynthesis. Bioprocess Biosyst. Eng. 2018, 41, 87–96. [Google Scholar] [CrossRef]
- Skjoedt, M.L.; Snoek, T.; Kildegaard, K.R.; Arsovska, D.; Eichenberger, M.; Goedecke, T.J.; Rajkumar, A.S.; Zhang, J.; Kristensen, M.; Lehka, B.J.; et al. Engineering Prokaryotic Transcriptional Activators as Metabolite Biosensors in Yeast. Nat. Chem. Biol. 2016, 12, 951–958. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Cai, Q.; Wang, Y.; Li, W.; Yu, J.; Yang, G.; Lin, W.; Lin, X. Four Lysr-Type Transcriptional Regulator Family Proteins (Lttrs) Involved in Antibiotic Resistance in Aeromonas hydrophila. World J. Microbiol. Biotechnol. 2019, 35, 127. [Google Scholar] [CrossRef]
- Wang, E.; Bauer, M.C.; Rogstam, A.; Linse, S.; Logan, D.T.; von Wachenfeldt, C. Structure and Functional Properties of the Bacillus subtilis Transcriptional Repressor Rex. Mol. Microbiol. 2008, 69, 466–478. [Google Scholar] [CrossRef]
- Abbasian, B.; Shair, A.; O’Gorman, D.B.; Pena-Diaz, A.M.; Brennan, L.; Engelbrecht, K.; Koenig, D.W.; Reid, G.; Burton, J.P. Potential Role of Extracellular Atp Released by Bacteria in Bladder Infection and Contractility. mSphere 2019, 4. [Google Scholar] [CrossRef] [Green Version]
- Kühlbrandt, W. Structure and Mechanisms of F-Type Atp Synthases. Annu. Rev. Biochem. 2019, 88, 515–549. [Google Scholar] [CrossRef]
- Joicy, A.; Song, Y.C.; Yu, H.; Chae, K.J. Nitrite and Nitrate as Electron Acceptors for Bioelectrochemical Ammonium Oxidation under Electrostatic Field. J. Environ. Manag. 2019, 250, 109517. [Google Scholar] [CrossRef] [PubMed]
- Clements, L.D.; Miller, B.S.; Streips, U.N. Comparative Growth Analysis of the Facultative Anaerobes Bacillus subtilis, Bacillus licheniformis, and Escherichia coli. Syst. Appl. Microbiol. 2002, 25, 284–286. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Zhao, Y.; Lou, W.; Su, J.; Wei, S.; Yang, X.; Wang, R.; Guan, R.; Pu, H.; Shen, W. Nitrate Reductase-Dependent Nitric Oxide Is Crucial for Multi-Walled Carbon Nanotube-Induced Plant Tolerance against Salinity. Nanoscale 2019, 11, 10511–10523. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.J.; Avila, S.; Bedmar, E.J.; Delgado, M.J. Overexpression of the Periplasmic Nitrate Reductase Supports Anaerobic Growth by Ensifer meliloti. FEMS Microbiol. Lett. 2018, 365, fny041. [Google Scholar] [CrossRef]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, A.; Galvan, A.; Fernandez, E. Nitrate Reductase Regulates Plant Nitric Oxide Homeostasis. Trends Plant Sci. 2017, 22, 163–174. [Google Scholar] [CrossRef]
- Stock, D.W.; Quattro, J.M.; Whitt, G.S.; Powers, D.A. Lactate Dehydrogenase (Ldh) Gene Duplication During Chordate Evolution: The Cdna Sequence of the Ldh of the Tunicate Styela Plicata. Mol. Biol. Evol. 1997, 14, 1273–1284. [Google Scholar] [CrossRef] [Green Version]
- Zheng, T.; Olson, D.G.; Murphy, S.J.; Shao, X.; Tian, L.; Lynd, L.R. Both Adhe and a Separate Nadph-Dependent Alcohol Dehydrogenase Gene, Adha, Are Necessary for High Ethanol Production in Thermoanaerobacterium saccharolyticum. J. Bacteriol. 2017, 199, e00542-00516. [Google Scholar] [CrossRef] [Green Version]
- Bareia, T.; Pollak, S.; Eldar, A. Self-Sensing in Bacillus subtilis Quorum-Sensing Systems. Nat. Microbiol. 2018, 3, 83–89. [Google Scholar] [CrossRef]
- Reimer, J.M.; Eivaskhani, M.; Harb, I.; Guarné, A.; Weigt, M.; Schmeing, T.M. Structures of a Dimodular Nonribosomal Peptide Synthetase Reveal Conformational Flexibility. Science 2019, 366, 706–711. [Google Scholar] [CrossRef]
- Boraker, D.K. Degu Colony Available. Science 1976, 192, 322. [Google Scholar] [CrossRef] [PubMed]
- Cairns, L.S.; Martyn, J.E.; Bromley, K.; Stanley-Wall, N.R. An Alternate Route to Phosphorylating Degu of Bacillus subtilis Using Acetyl Phosphate. BMC Microbiol. 2015, 15, 78–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zschiedrich, C.P.; Keidel, V.; Szurmant, H. Molecular Mechanisms of Two-Component Signal Transduction. J. Mol. Biol. 2016, 428, 3752–3775. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Sun, D.; Zhu, J.; Liu, W. Two-Component Signal Transduction Systems: A Major Strategy for Connecting Input Stimuli to Biofilm Formation. Front. Microbiol. 2018, 9, 3279. [Google Scholar] [CrossRef] [PubMed]
- Bacon Schneider, K.; Palmer, T.M.; Grossman, A.D. Characterization of Comq and Comx, Two Genes Required for Production of Comx Pheromone in Bacillus subtilis. J. Bacteriol. 2002, 184, 410–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, P.; Morrison, D.A. Transient Association of an Alternative Sigma Factor, Comx, with Rna Polymerase During the Period of Competence for Genetic Transformation in Streptococcus pneumoniae. J. Bacteriol. 2003, 185, 349–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, V.; Latour, J.M. Perr Vs Ohrr: Selective Peroxide Sensing in Bacillus subtilis. Mol Biosyst 2010, 6, 316–323. [Google Scholar] [CrossRef]
- Aspiras, M.B.; Ellen, R.P.; Cvitkovitch, D.G. Comx Activity of Streptococcus Mutans Growing in Biofilms. FEMS Microbiol. Lett. 2004, 238, 167–174. [Google Scholar]
- Gupta, M.; Rao, K.K. Epr Plays a Key Role in Degu-Mediated Swarming Motility of Bacillus subtilis. FEMS Microbiol. Lett. 2009, 295, 187–194. [Google Scholar] [CrossRef]
- O’Loughlin, C.T.; Miller, L.C.; Siryaporn, A.; Drescher, K.; Semmelhack, M.F.; Bassler, B.L. A Quorum-Sensing Inhibitor Blocks Pseudomonas Aeruginosa Virulence and Biofilm Formation. Proc. Natl. Acad. Sci. USA 2013, 110, 17981–17986. [Google Scholar] [CrossRef] [Green Version]
- Gallego del Sol, F.; Marina, A. Structural Basis of Rap Phosphatase Inhibition by Phr Peptides. PLoS Biol. 2013, 11, e1001511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Defoirdt, T. Quorum-Sensing Systems as Targets for Antivirulence Therapy. Trends Microbiol. 2018, 26, 313–328. [Google Scholar] [CrossRef] [PubMed]
- Duitman, E.H.; Wyczawski, D.; Boven, L.G.; Venema, G.; Kuipers, O.P.; Hamoen, L.W. Novel Methods for Genetic Transformation of Natural Bacillus subtilis Isolates Used to Study the Regulation of the Mycosubtilin and Surfactin Synthetases. Appl. Environ. Microbiol. 2007, 73, 3490–3496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haque, S.; Yadav, D.K.; Bisht, S.C.; Yadav, N.; Singh, V.; Dubey, K.K.; Jawed, A.; Wahid, M.; Dar, S.A. Quorum Sensing Pathways in Gram-Positive and -Negative Bacteria: Potential of Their Interruption in Abating Drug Resistance. J. Chemother. 2019, 31, 161–187. [Google Scholar] [CrossRef]
- Silpe, J.E.; Bassler, B.L. A Host-Produced Quorum-Sensing Autoinducer Controls a Phage Lysis-Lysogeny Decision. Cell 2019, 176, 268–280. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.V.; Johnson, R.D.; Arcus, V.L.; Lott, J.S. Prediction of the Substrate for Nonribosomal Peptide Synthetase (Nrps) Adenylation Domains by Virtual Screening. Proteins 2015, 83, 2052–2066. [Google Scholar] [CrossRef] [PubMed]
- Hacker, C.; Cai, X.; Kegler, C.; Zhao, L.; Weickhmann, A.K.; Wurm, J.P.; Bode, H.B.; Wöhnert, J. Structure-Based Redesign of Docking Domain Interactions Modulates the Product Spectrum of a Rhabdopeptide-Synthesizing Nrps. Nat. Commun. 2018, 9, 4366. [Google Scholar] [CrossRef] [Green Version]
- Sussmuth, R.D.; Mainz, A. Nonribosomal Peptide Synthesis-Principles and Prospects. Angew. Chem. Int. Ed. Engl. 2017, 56, 3770–3821. [Google Scholar] [CrossRef] [PubMed]
- Berry, D.; Mace, W.; Grage, K.; Wesche, F.; Gore, S.; Schardl, C.L.; Young, C.A.; Dijkwel, P.P.; Leuchtmann, A.; Bode, H.B.; et al. Efficient Nonenzymatic Cyclization and Domain Shuffling Drive Pyrrolopyrazine Diversity from Truncated Variants of a Fungal Nrps. Proc. Natl. Acad. Sci. USA 2019, 116, 25614–25623. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Liu, H.; Ma, Z.; Han, J.; Lu, Z.; Dai, C.; Lv, F.; Bie, X. Translocation of the Thioesterase Domain for the Redesign of Plipastatin Synthetase. Sci. Rep. 2016, 6, 38467. [Google Scholar] [CrossRef]
- Drake, E.J.; Miller, B.R.; Shi, C.; Tarrasch, J.T.; Sundlov, J.A.; Allen, C.L.; Skiniotis, G.; Aldrich, C.C.; Gulick, A.M. Structures of Two Distinct Conformations of Holo-Non-Ribosomal Peptide Synthetases. Nature 2016, 529, 235–238. [Google Scholar] [CrossRef] [Green Version]
- Marahiel, M.A. A Structural Model for Multimodular Nrps Assembly Lines. Nat. Prod. Rep. 2016, 33, 136–140. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.-Y.; Wang, L.; Liu, Y.-F.; Zhou, L.; Gang, H.-Z.; Liu, J.-F.; Yang, S.-Z.; Mu, B.-Z. Microbial Lipopeptide-Producing Strains and Their Metabolic Roles under Anaerobic Conditions. Microorganisms 2021, 9, 2030. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102030
Li J-Y, Wang L, Liu Y-F, Zhou L, Gang H-Z, Liu J-F, Yang S-Z, Mu B-Z. Microbial Lipopeptide-Producing Strains and Their Metabolic Roles under Anaerobic Conditions. Microorganisms. 2021; 9(10):2030. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102030
Chicago/Turabian StyleLi, Jia-Yi, Lu Wang, Yi-Fan Liu, Lei Zhou, Hong-Ze Gang, Jin-Feng Liu, Shi-Zhong Yang, and Bo-Zhong Mu. 2021. "Microbial Lipopeptide-Producing Strains and Their Metabolic Roles under Anaerobic Conditions" Microorganisms 9, no. 10: 2030. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9102030