
Development of Various Leishmania (Sauroleishmania) tarentolae Strains in Three Phlebotomus Species

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Parasites and Sand Flies
2.2. Sand Fly Infections
2.3. Morphometry of Parasites
3. Results
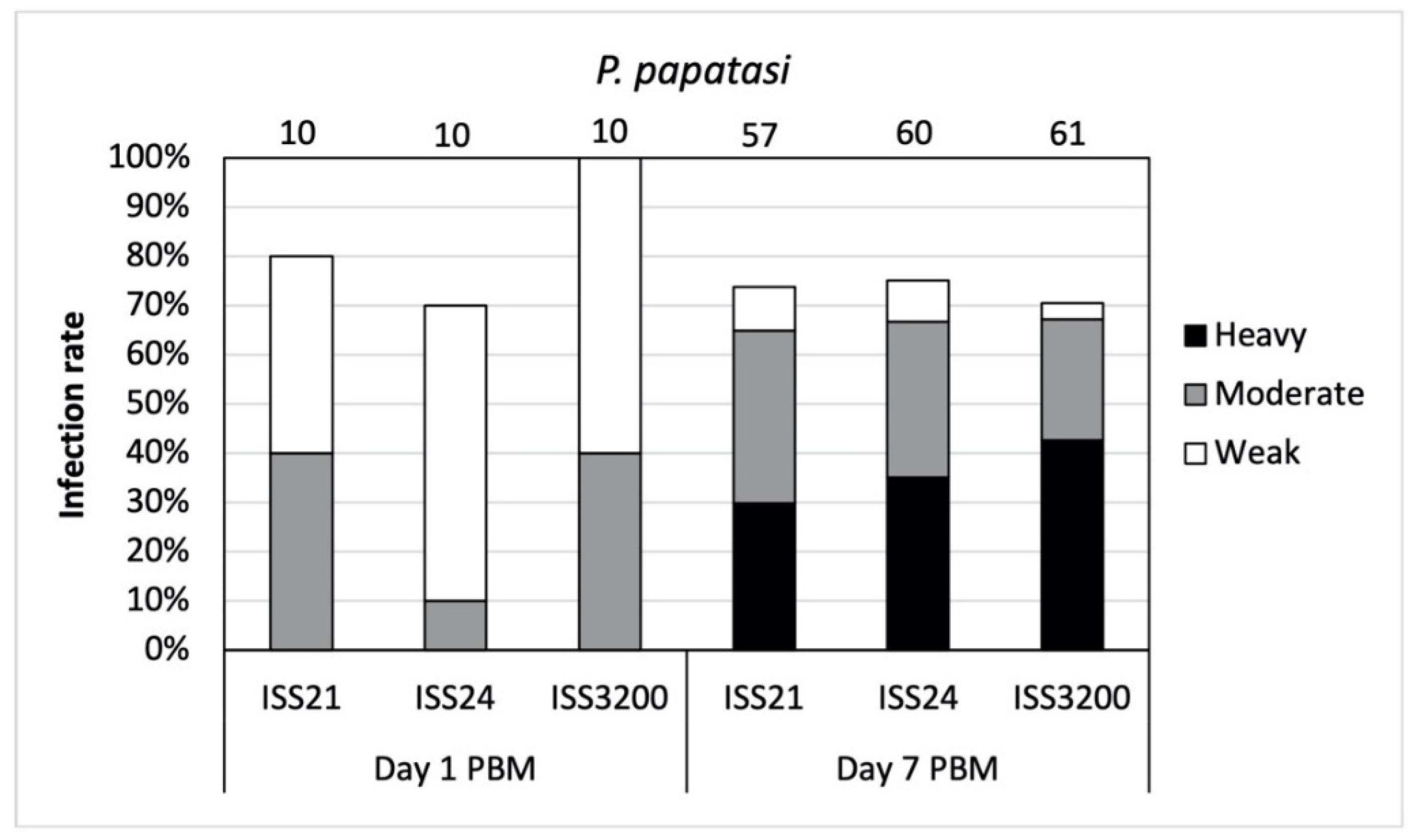
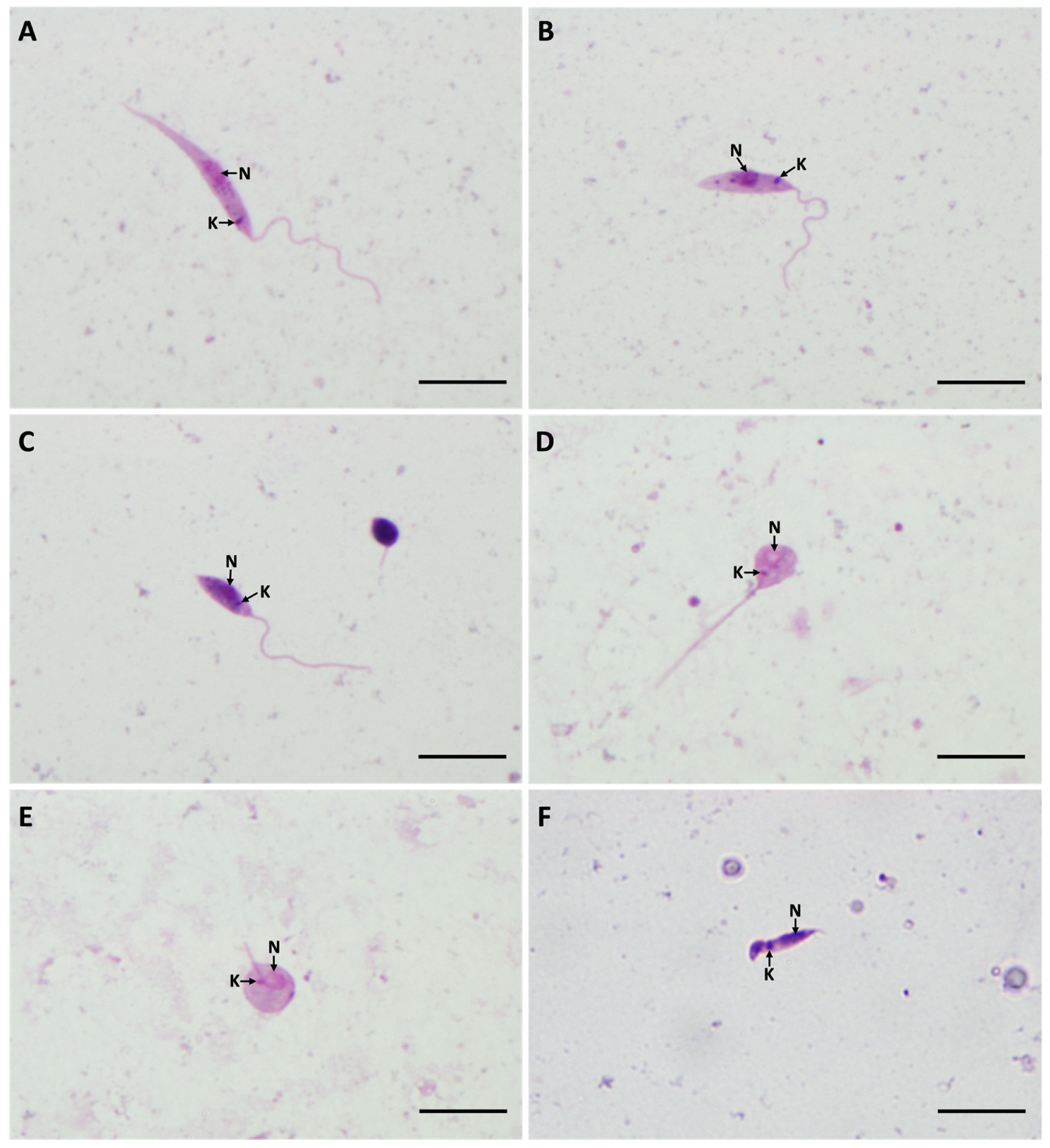
3.1. Development of L. (S.) tarentolae in P. papatasi
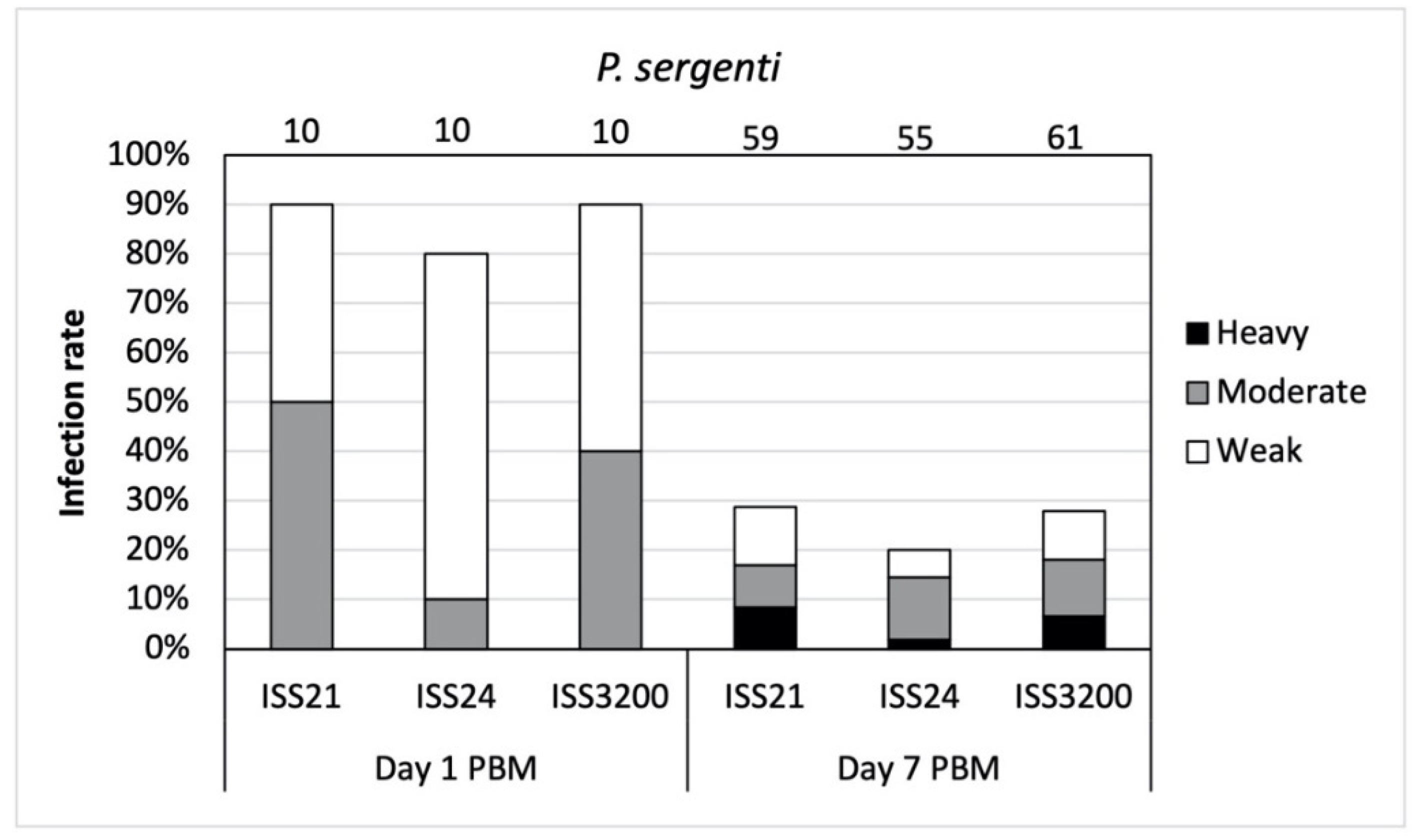
3.2. Development of L. (S.) tarentolae in P. sergenti
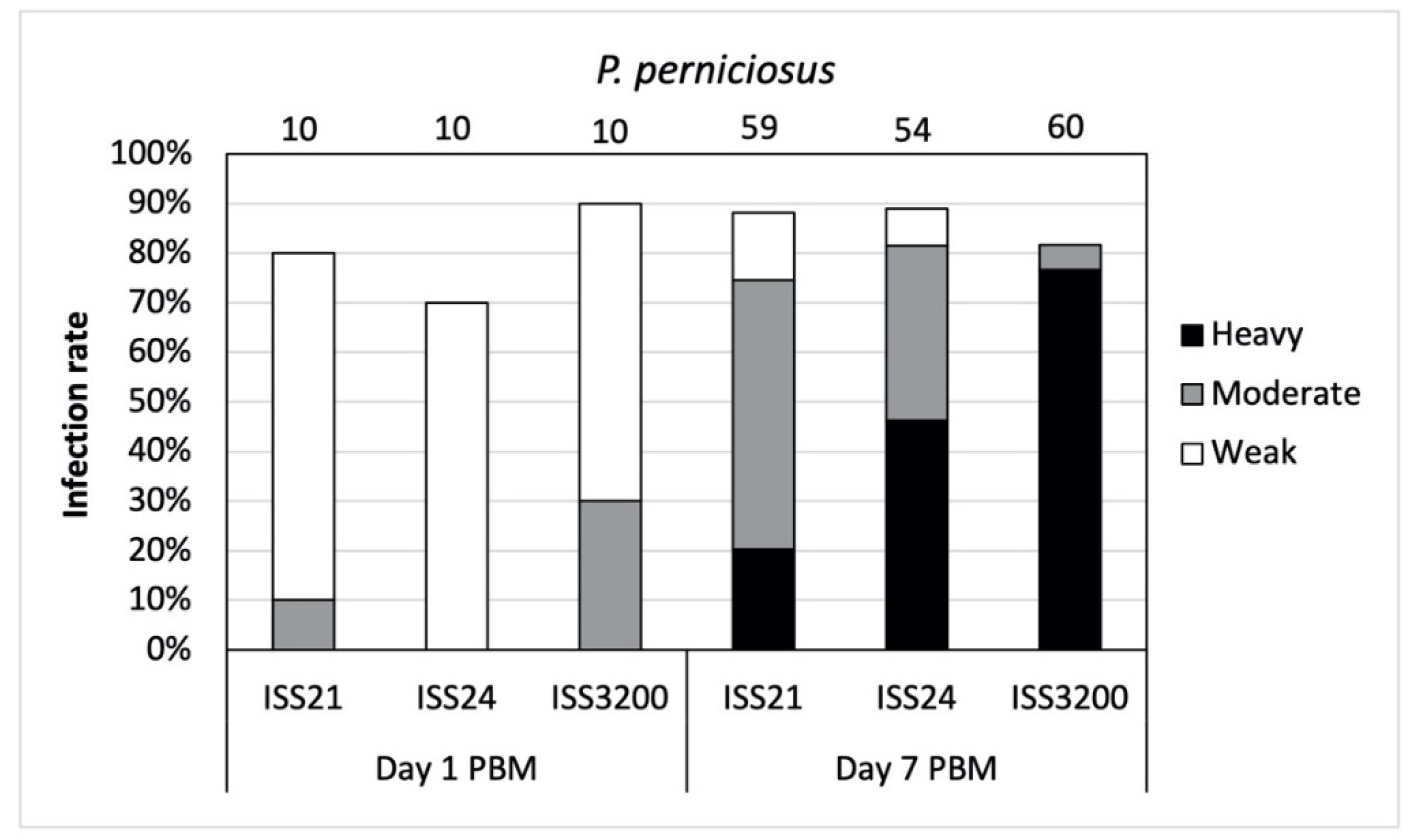
3.3. Development of L. (S.) tarentolae in P. perniciosus
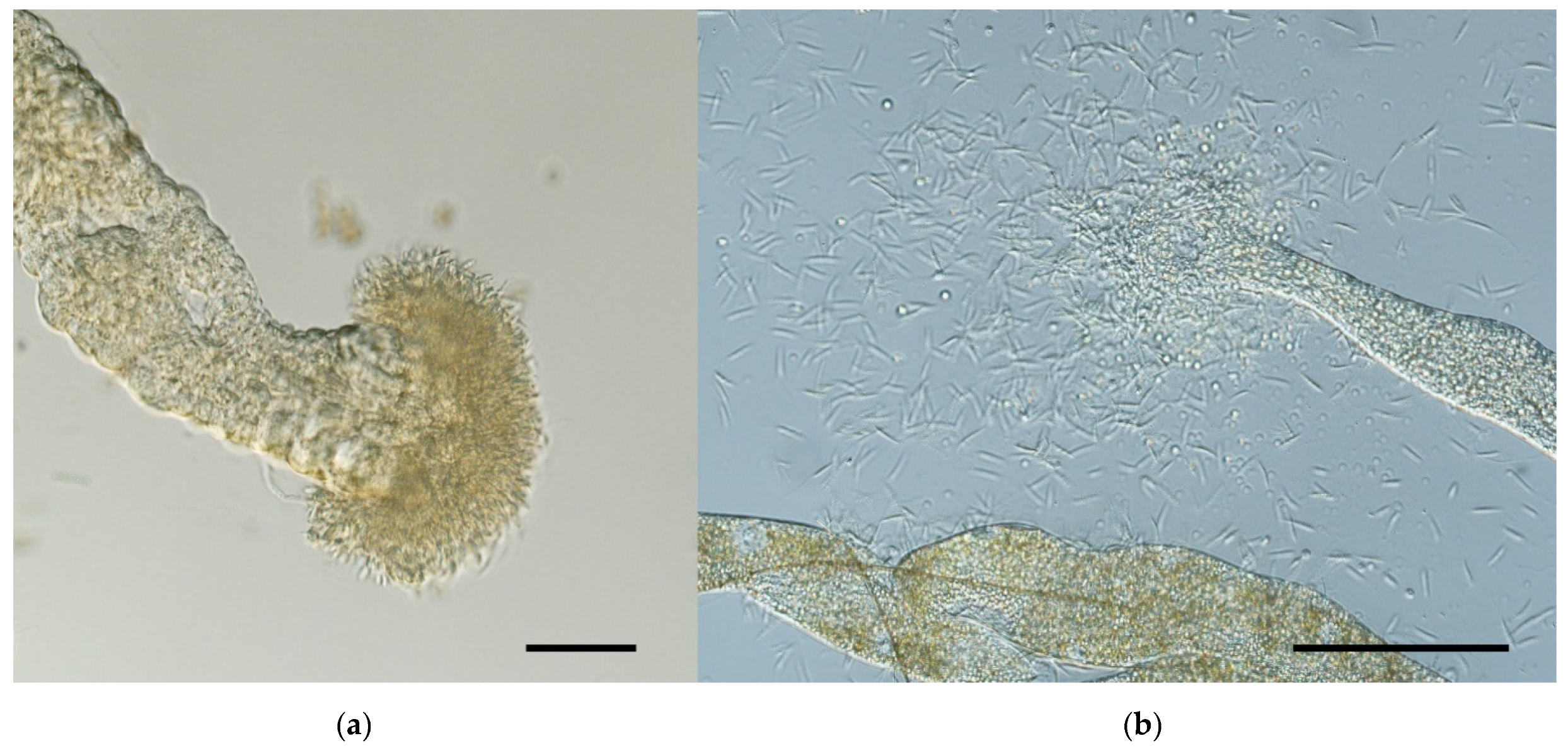
3.4. Morphometry of Promastigotes from Gut Smears
4. Discussion
5. Conclusions
- We demonstrated the ability of L. (S.) tarentolae to develop in the midgut of three Phlebotomus sand flies. The most permissive to parasite development was P. perniciosus and we suggest that this species, together with P. papatasi, could be involved in L. (S.) tarentolae circulation and should be considered as potential secondary vectors of this parasite.
- L. (S.) tarentolae was frequently found in anterior midgut and stomodeal valve of infected sand fly females, which challenges previous definition of its hypopylarian development (limited to hindgut only). Interestingly, heavy parasite loads were frequently found in Malpighian tubules, which suggests that this localization, found previously in monoxenous Crithidia parasites, is unique among Leishmania but typical for L. (S.) tarentolae development in Phlebotomus sand flies.
- For better understanding of Sauroleishmania life cycle, its morphological forms, and localization in the sand fly gut, it would be important to study the development of L. (S.) tarentolae in its proven natural vector Sergentomyia minuta.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Espinosa, O.A.; Serrano, M.G.; Camargo, E.P.; Teixeira, M.M.G.; Shaw, J.J. An appraisal of the taxonomy and nomenclature of trypanosomatids presently classified as Leishmania and Endotrypanum. Parasitology 2016, 145, 430–442. [Google Scholar] [CrossRef]
- Ranque, P. Étude morphologique et biologique de quelques Trypanosomidés récoltés au Sénégal. Ph.D. Thesis, Université d’Aix-Marseille II, Marseille, France, 28 April 1973. [Google Scholar]
- Saf’janova, V.M. The problem of taxonomy with Leishmania. In The Leishmanias; Protozoology, Academy of Sciences; USSR All Union Society of Protozoologists: Leningrad, Russia, 1982; Volume 7, pp. 95–101. [Google Scholar]
- Killick-Kendrick, R.; Lainson, R.; Rioux, J.A.; Saf’janova, V.M. The taxonomy of Leishmania-like parasites of reptiles. In Leishmania: Taxonomie et Phylogenèse; Rioux, J.A., Ed.; Applications Eco-epidéemiologiques (Colloque international du CNRS/INSERM, 1984), IMEE: Montpelier, France, 1986; pp. 143–148. [Google Scholar]
- Akhoundi, M.; Kuhls, K.; Cannet, A.; Votypka, J.; Marty, P.; Delaunay, P.; Sereno, D. A historical overview of the classification, evolution, and dispersion of Leishmania parasites and sandflies. PLoS Negl. Trop. Dis. 2016, 10, e0004349. [Google Scholar] [CrossRef]
- Noyes, H. Implications of a Neotropical origin of the genus Leishmania. Memórias do Instituto Oswaldo Cruz 1998, 93, 657–662. [Google Scholar] [CrossRef]
- Klatt, S.; Simpson, L.; Maslov, D.A.; Konthur, Z. Leishmania tarentolae: Taxonomic classification and its application as a promising biotechnological expression host. PLoS Negl. Trop. Dis. 2019, 13, e0007424. [Google Scholar] [CrossRef] [Green Version]
- Belova, E.M. Reptiles and their importance in the epidemiology of leishmaniasis. Bull. World Health Organ. 1971, 44, 553–560. [Google Scholar]
- Wilson, V.C.L.C.; Southgate, B.A. Lizard Leishmania. In Biology of the Kinetoplastida; Lumsden, W.H.R., Evans, D.A., Eds.; Academic Press: London, UK, 1979; Volume 2, pp. 241–268. ISBN 978-0124602014. [Google Scholar]
- Adler, S. The behaviour of a lizard Leishmania in hamsters and baby mice. Rev. Do Inst. De Med. Trop. De Sao Paulo 1962, 4, 61–64. [Google Scholar]
- Breton, M.; Tremblay, M.J.; Ouellette, M.; Papadopoulou, B. Live nonpathogenic parasitic vector as a candidate vaccine against visceral leishmaniasis. Infect. Immun. 2005, 73, 6372–6382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, V.M.; Munoz, D.L.; Cedeno, D.L.; Velez, I.D.; Jones, M.A.; Robledo, S.M. Leishmania tarentolae: Utility as an in vitro model for screening of antileishmanial agents. Exp. Parasitol. 2010, 126, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Novo, S.P.; Leles, D.; Bianucci, R.; Araujo, A. Leishmania tarentolae molecular signatures in a 300 hundred-years-old human Brazilian mummy. Parasites Vectors 2015, 8, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coughlan, S.; Mulhair, P.; Sanders, M.; Schonian, G.; Cotton, J.A.; Downing, T. The genome of Leishmania adleri from a mammalian host highlights chromosome fission in Sauroleishmania. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Mendoza-Roldan, J.A.; Latrofa, M.S.; Iatta, R.; Manoj, R.R.S.; Panarese, R.; Annoscia, G.; Pombi, M.; Zatelli, A.; Beugnet, F.; Otranto, D. Detection of Leishmania tarentolae in lizards, sand flies and dogs in southern Italy, where Leishmania infantum is endemic: Hindrances and opportunities. Parasites Vectors 2021, 14, 461. [Google Scholar] [CrossRef] [PubMed]
- Pombi, M.; Giacomi, A.; Barlozzari, G.; Mendoza-Roldan, J.A.; Macrì, G.; Otranto, D.; Gabrielli, S. Molecular detection of Leishmania (Sauroleishmania) tarentolae in human blood and Leishmania (Leishmania) infantum in Sergentomyia minuta: Unexpected host-parasite contacts. Med. Vet. Entomol. 2020, 34, 470–475. [Google Scholar] [CrossRef]
- Telford, S.R. Hemoparasites of the Reptilia; Color Atlas and Text; CRC Press: Boca Raton, FL, USA, 2009; Volume 1, pp. 311–376. ISBN 978-1-4200-8040-7. [Google Scholar]
- Maroli, M.; Gramiccia, M.; Gradoni, L.; Ready, P.D.; Smith, D.F.; Aquino, C. Natural infections of phlebotomine sandflies with Trypanosomatidae in central and south Italy. Trans. R. Soc. Trop. Med. Hyg. 1988, 82, 227–228. [Google Scholar] [CrossRef]
- Quate, L.W. Phlebotomus sandflies of the Paloich area in the Sudan (Diptera, Psychodidae). J. Med Entomol. 1964, 1, 213–268. [Google Scholar] [CrossRef] [PubMed]
- Mutinga, M.J.; Ngoka, J.M. Suspected vectors of lizard leishmaniasis in Kenya and their possible role in partial immunization of the human populationl against Leishmania donovani in kala-azar endemic areas. Int. J. Trop. Insect Sci. 1981, 1, 207–210. [Google Scholar] [CrossRef]
- Adler, S.; Theodor, O. Observations on Leishmania ceramodactyli n. sp. Trans. R. Soc. Trop. Med. Hyg. 1929, 22, 343–355. [Google Scholar] [CrossRef]
- Lainson, R.; Shaw, J.J. Evolution, classification and geographical distribution. In The Leishmaniases in Biology and Medicine; Peters, W., Killick-Kendrick, R., Eds.; Academic Press: Cambridge, MA, USA, 1987; Volume 1, pp. 1–120. ISBN 0-12-552101-4. [Google Scholar]
- Adler, S.; Theodor, O. Investigation on Mediterranean kala azar X—A note on Trypanosoma platydactyli and Leishmania tarentolae. Proc. R. Soc. Lond. Ser. B-Biol. Sci. 1935, 116, 543–544. [Google Scholar] [CrossRef]
- Heisch, R.B. On Leishmania adleri sp. nov. from lacertid lizards (Latastia sp.) in Kenya. Ann. Trop. Med. Parasitol. 1985, 52, 68–71. [Google Scholar] [CrossRef]
- Killick-Kendrick, R. Biology of Leishmania in phlebotomine sandflies. In Biology of the Kinetoplastida; Lumsden, W.H.R., Evans, D.A., Eds.; Academic Press: London, UK, 1979; Volume 2, pp. 395–460. ISBN 978-0124602014. [Google Scholar]
- Bates, P.A. Transmission of Leishmania metacyclic promastigotes by phlebotomine sand flies. Int. J. Parasitol. 2007, 37, 1097–1106. [Google Scholar] [CrossRef]
- Wenyon, C.M. Observations on the intestinal protozoa of three Egyptian lizards, with a note on a cell-invading fungus. Parasitology 1921, 12, 350–365. [Google Scholar] [CrossRef] [Green Version]
- Wallbanks, K.R.; Maazoun, R.; Canning, E.U.; Rioux, J.A. The identity of Leishmania tarentolae Wenyon 1921. Parasitology 1985, 90, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Pozio, E.; Gramiccia, M.; Gradoni, L.; Maroli, M. Hémoflagellés de Tarentola mauritanica L., 1758 (Reptilia, Gekkonidae). In Leishmania. Taxonomie et phylogenèse; Rioux, J.A., Ed.; IMEEE: Montpellier, France, 1986; pp. 149–155. [Google Scholar]
- Rioux, J.A.; Knoepfler, L.P.; Martini, A.; Callot, J.; Kremer, M. Présence en France de Leishmania tarentolae Wenyon, 1921. Parasite du gecko Tarentola mauritanica (L. 1758). Ann. De Parasitol. Hum. Et Comparée 1969, 44, 115–118. [Google Scholar] [CrossRef]
- Pozio, E.; Gramiccia, M.; Gradoni, L.; Maroli, M. Hemoflagellates in Cyrtodactylus kotschyi (Steindachner, 1870) (Reptilia, Gekkonidae) in Italy. Acta Trop. 1983, 40, 399–400. [Google Scholar]
- Elwasila, M. Leishmania tarentolae Wenyon, 1921 from the gecko Tarentola annularis in the Sudan. Parasitol. Res. 1988, 74, 591–592. [Google Scholar] [CrossRef]
- Bongiorno, G.; Di Muccio, T.; Gradoni, L.; Giacomi, A.; Pombi, M.; Gabrielli, S.; Gramiccia, M. Natural infections of Sergentomyia minuta with kinetoplastid flagellates detected by gold standard methods in Rome province. In Proceedings of the XXXI Congresso SoIPa & 2021 ESDA EVENT, Teramo, Italy, 16–19 June 2021; p. 230. [Google Scholar]
- Di Muccio, T.; Scalone, A.; Bruno, A.; Marangi, M.; Grande, R.; Armignacco, O.; Gradoni, L.; Gramiccia, M. Epidemiology of Imported Leishmaniasis in Italy: Implications for a European Endemic Country. PLoS ONE 2015, 10, e0129418. [Google Scholar] [CrossRef]
- Diamond, L.S.; Herman, C.M. Incidence of trypanosomes in the Canada goose as revealed by bone marrow culture. J. Parasitol. 1954, 40, 195–202. [Google Scholar] [CrossRef]
- Volf, P.; Volfova, V. Establishment and maintenance of sand fly colonies. J. Vector Ecol. 2011, 36, 1–9. [Google Scholar] [CrossRef]
- Myskova, J.; Votypka, J.; Volf, P. Leishmania in sand flies: Comparison of quantitative polymerase chain reaction with other techniques to determine the intensity of infection. J. Med. Entomol. 2008, 45, 133–138. [Google Scholar] [CrossRef]
- Killick-Kendrick, R. The life-cycle of Leishmania in the sandfly with special reference to the form infective to the vertebrate host. Ann. De Parasitol. Hum. Et Comparée 1990, 65, 37–42. [Google Scholar] [CrossRef] [Green Version]
- Sadlova, J.; Price, H.P.; Smith, B.A.; Votypka, J.; Volf, P.; Smith, D.F. The stage-regulated HASPB and SHERP proteins are essential for differentiation of the protozoan parasite Leishmania major in its sand fly vector, Phlebotomus papatasi. Cell. Microbiol. 2010, 12, 1765–1779. [Google Scholar] [CrossRef] [Green Version]
- Blum, B.; Bakalara, N.; Simpson, L. A model for RNA editing in kinetoplastid mitochondria: RNA molecules transcribed from maxicircle DNA provide the edited information. Cell 1990, 60, 189–198. [Google Scholar] [CrossRef]
- Aphasizhev, R.; Aphasizheva, I.; Nelson, R.E.; Gao, G.; Simpson, A.M.; Kang, X.; Falick, A.M.; Sbicego, S.; Simpson, L. Isolation of a U-insertion/deletion editing complex from Leishmania tarentolae mitochondria. EMBO J. 2003, 22, 913–924. [Google Scholar] [CrossRef] [Green Version]
- Aphasizhev, R.; Aphasizheva, I. Mitochondrial RNA editing in trypanosomes: Small RNAs in control. Biochimie 2013, 100, 125–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdossamadi, Z.; Taheri, T.; Seyed, N.; Montakhab-Yeganeh, H.; Zahedifard, F.; Taslimi, Y.; Habibzadeh, S.; Gholami, E.; Gharibzadeh, A.; Rafati, S. Live Leishmania tarentolae secreting HNP1 as an immunotherapeutic tool against Leishmania infection in BALB/c mice. Immunotherapy 2017, 9, 1089–1102. [Google Scholar] [CrossRef]
- Montakhab-Yeganeh, H.; Abdossamadi, Z.; Zahedifard, F.; Taslimi, Y.; Badirzadeh, A.; Saljoughian, N.; Taheri, T.; Taghikhani, M.; Rafati, S. Leishmania tarentolae expressing CXCL-10 as an efficient immunotherapy approach against Leishmania major-infected BALB/c mice. Parasite Imunol. 2017, 39, e12461. [Google Scholar] [CrossRef]
- Pimenta, P.F.P.; Saraiva, E.M.; Rowton, E.; Modi, G.B.; Garraway, L.A.; Beverley, S.M.; Turco, S.J.; Sacks, D.L. Evidence that the vectorial competence of phlebotomine sand flies for different species of Leishmania is controlled by structural polymorphisms in the surface lipophosphoglycan. Proc. Natl. Acad. Sci. USA 1994, 91, 9155–9159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chajbullinova, A.; Votypka, J.; Sadlova, J.; Kvapilova, K.; Seblova, V.; Kreisinger, J.; Jirku, M.; Sanjoba, C.; Gantuya, S.; Matsumoto, Y.; et al. The development of Leishmania turanica in sand flies and competition with L. major. Parasites Vectors 2012, 5, 219. [Google Scholar] [CrossRef] [Green Version]
- Svobodova, M.; Sadlova, J.; Chang, K.P.; Volf, P. Distribution and feeding preference of the sand flies Phlebotomus sergenti and P. papatasi in a cutaneous leishmaniasis focus in Sanliurfa, Turkey. Am. J. Trop. Med. Hyg. 2003, 68, 6–9. [Google Scholar] [CrossRef] [PubMed]
- Palit, A.; Bhattacharya, S.K.; Kundu, S.N. Host preference of Phlebotomus argentipes and Phlebotomus papatasi in different biotopes of West Bengal, India. Int. J. Environ. Health Res. 2005, 15, 449–454. [Google Scholar] [CrossRef]
- McMillan, B. Leishmaniasis in the Sudan Republic. 22. Leishmania hoogstraali sp. n. in the gecko. J. Parasitol. 1965, 51, 336–339. [Google Scholar] [CrossRef]
- Kamhawi, S.; Modi, G.B.; Pimenta, P.F.P.; Rowton, E.; Sacks, D.L. The vectorial competence of Phlebotomus sergenti is specific for Leishmania tropica and is controlled by species-specific, lipophosphoglycan-mediated midgut attachment. Parasitology 2000, 121, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Maroli, M.; Jalouk, L.; Al Ahmed, M.; Bianchi, R.; Bongiorno, G.; Khoury, C.; Gradoni, L. Aspects of the bionomics of Phlebotomus sergenti sandflies from an endemic area of anthroponotic cutaneous leishmaniasis in Aleppo Governorate, Syria. Med. Vet. Entomol. 2009, 23, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Maroli, M.; Gramiccia, M.; Gradoni, L.; Troiani, M.; Ascione, R. Natural infection of Phlebotomus perniciosus with MON 72 zymodeme of Leishmania infantum in the Campania region of Italy. Acta Trop. 1994, 57, 333–335. [Google Scholar] [CrossRef]
- Volf, P.; Myskova, J. Sand flies and Leishmania: Specific versus permissive vectors. Trends Parasitol. 2007, 23, 91–92. [Google Scholar] [CrossRef] [Green Version]
- Bongiorno, G.; Habluetzel, A.; Khoury, C.; Maroli, M. Host preferences of phlebotomine sand flies at a hypoendemic focus of canine leishmaniasis in central Italy. Acta Trop. 2003, 88, 109–116. [Google Scholar] [CrossRef]
- Rossi, E.; Bongiorno, G.; Ciolli, E.; Di Muccio, T.; Scalone, A.; Gramiccia, M.; Gradoni, L.; Maroli, M. Seasonal phenology, host-blood feeding preferences and natural Leishmania infection of Phlebotomus perniciosus (Diptera, Psychodidae) in a high-endemic focus of canine leishmaniasis in Rome province, Italy. Acta Trop. 2008, 105, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Remadi, L.; Chargui, N.; Jimenez, M.; Molina, R.; Haouas, N.; González, E.; Chaabane-Banaouas, R.; Salah, E.B.; Haddaji, M.; Chaabouni, Y.; et al. Molecular detection and identification of Leishmania DNA and blood meal analysis in Phlebotomus (Larroussius) species. PLoS Negl. Trop. Dis. 2020, 14, e0008077. [Google Scholar] [CrossRef]
- Bettini, S.; Gramiccia, M.; Gradoni, L.; Atzeni, M.C. Leishmaniasis in Sardinia: II. Natural infection of Phlebotomus perniciosus Newstead 1911, by Leishmania infantum Nicolle 1908, in the province of Cagliari. Trans. R. Soc. Trop. Med. Hyg. 1986, 80, 458–459. [Google Scholar] [CrossRef]
- Bongiorno, G.; Lisi, O.; Severini, F.; Vaccalluzzo, V.; Khoury, C.; Di Muccio, T.; Gradoni, L.; Maroli, M.; D’Urso, V.; Gramiccia, M. Investigations on sand fly bionomics and Leishmania natural infections in Eastern Sicily, Italy, with particular reference to Phlebotomus sergenti. In Proceedings of the VIII International Symposium on Phlebotomine Sandflies, Puerto Iguazu, Argentina, 22–25 September 2014. id 45-O. [Google Scholar]
- Latrofa, M.S.; Mendoza-Roldan, J.A.; Manoj, R.; Dantas-Torres, F.; Otranto, D. A duplex real-time PCR assay for the detection and differentiation of Leishmania infantum and Leishmania tarentolae in vectors and potential reservoir hosts. Entomol. Gen. 2021. [Google Scholar] [CrossRef]
- Franco, A.M.R.; Tesh, R.B.; Guzman, H.; Deane, M.P.; Grimaldi, G., Jr. Development of Endotrypanum (Kinetoplastida: Trypanosomatidae) in experimentally infected phlebotomine sand flies (Diptera: Psychodidae). J. Med Entomol. 1997, 34, 189–192. [Google Scholar] [CrossRef]
- McConville, M.J.; Turco, S.J.; Ferguson, M.A.; Sacks, D.L. Developmental modification of lipophosphoglycan during the differentiation of Leishmania major promastigotes to an infectious stage. EMBO J. 1992, 11, 3593–3600. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sauroleishmania Strain | Culture | P. papatasi | P. sergenti | P. perniciosus |
|---|---|---|---|---|
| ISS21 | 18% | 15% | 15% | 16% |
| ISS24 | 21% | 12% | 12% | 12% |
| ISS3200 | 22% | 11% | 15% | 16% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ticha, L.; Kykalova, B.; Sadlova, J.; Gramiccia, M.; Gradoni, L.; Volf, P. Development of Various Leishmania (Sauroleishmania) tarentolae Strains in Three Phlebotomus Species. Microorganisms 2021, 9, 2256. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112256
Ticha L, Kykalova B, Sadlova J, Gramiccia M, Gradoni L, Volf P. Development of Various Leishmania (Sauroleishmania) tarentolae Strains in Three Phlebotomus Species. Microorganisms. 2021; 9(11):2256. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112256
Chicago/Turabian StyleTicha, Lucie, Barbora Kykalova, Jovana Sadlova, Marina Gramiccia, Luigi Gradoni, and Petr Volf. 2021. "Development of Various Leishmania (Sauroleishmania) tarentolae Strains in Three Phlebotomus Species" Microorganisms 9, no. 11: 2256. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9112256