
Persistent Cryptosporidium parvum Infection Leads to the Development of the Tumor Microenvironment in an Experimental Mouse Model: Results of a Microarray Approach

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cryptosporidium Oocysts
2.2. Mouse Model of Cryptosporidiosis
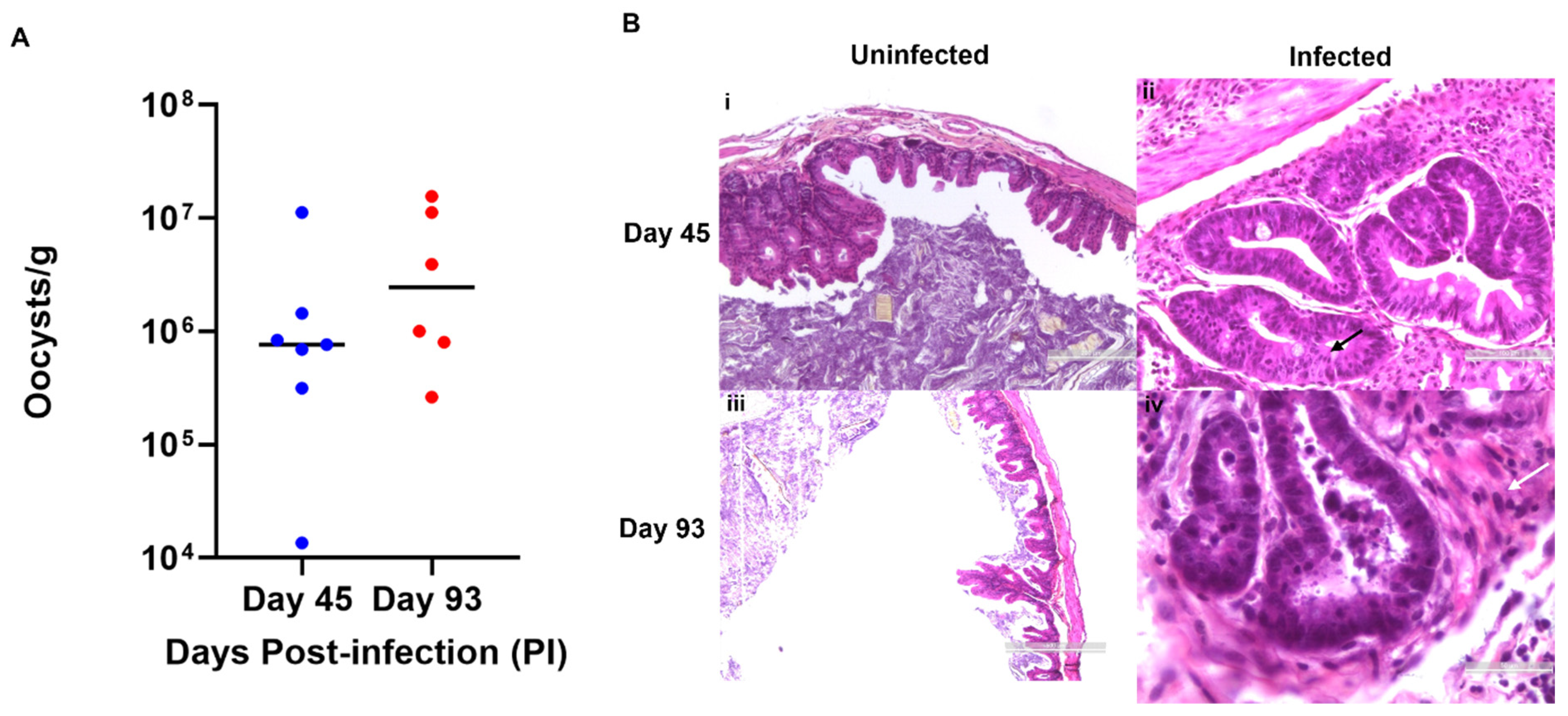
2.3. Oocyst Shedding
2.4. Histological Examination
2.5. Agilent Microarray Analysis
2.6. Data Analysis
2.7. Validation by Quantitative Reverse Transcription PCR
3. Results
3.1. C. parvum Induces Chronic Infection and Neoplasia in a Mouse Model
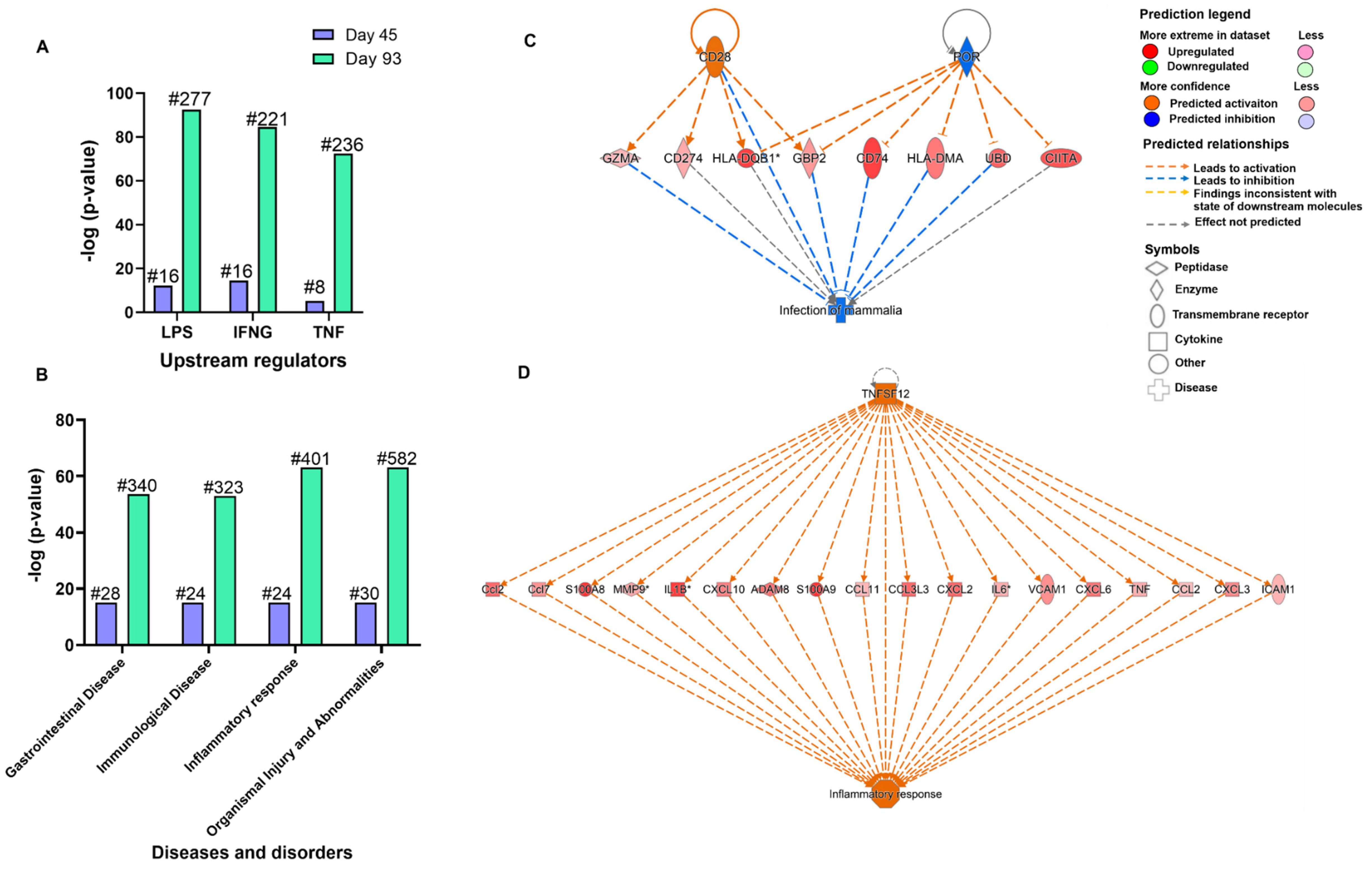
3.2. Chronic Cryptosporidiosis Induced Neoplasia Results in Global Gene Profile Alterations
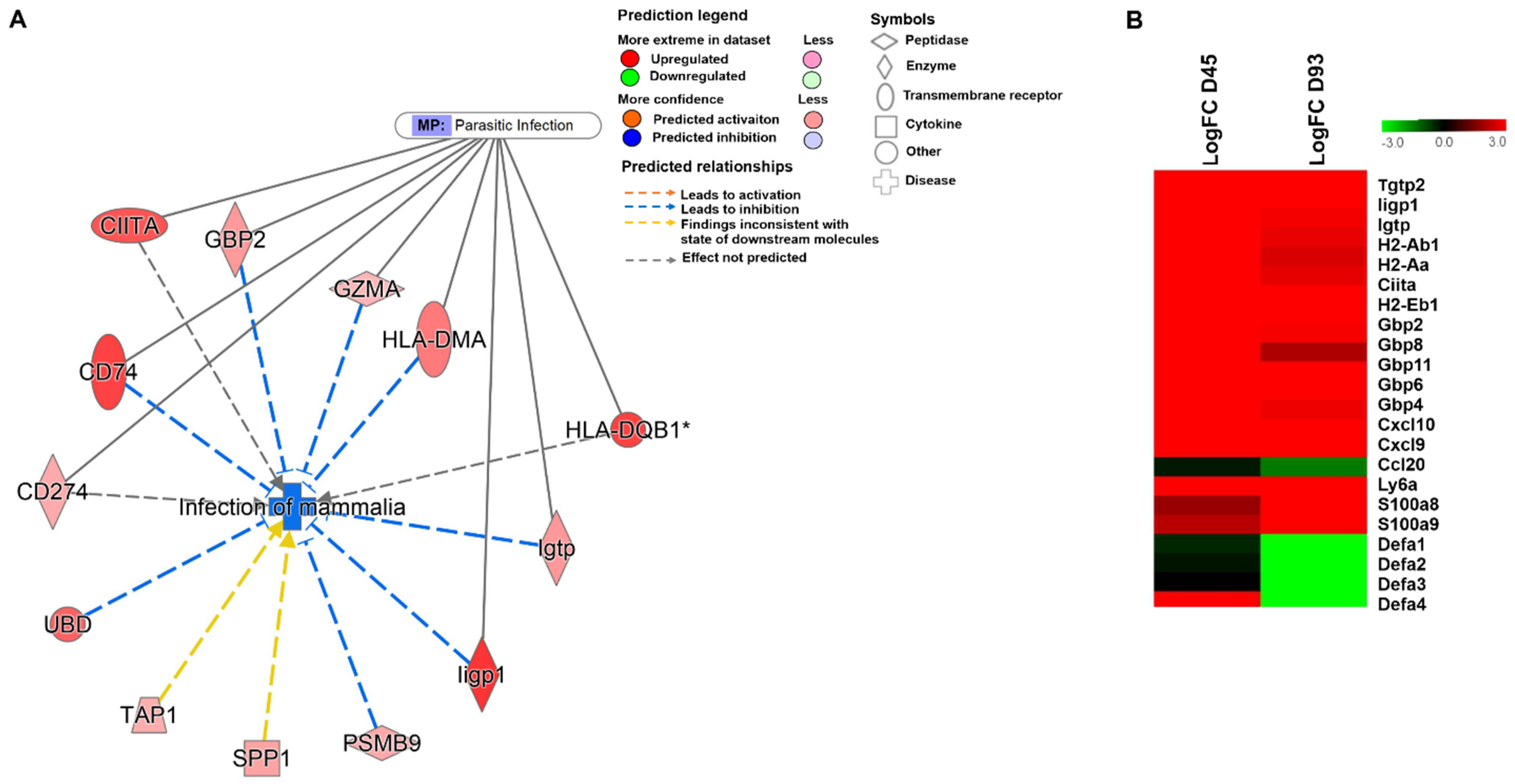
3.3. Regulation of Anti-Microbial Peptides Like α-Defensins during the Course of C. parvum Infection
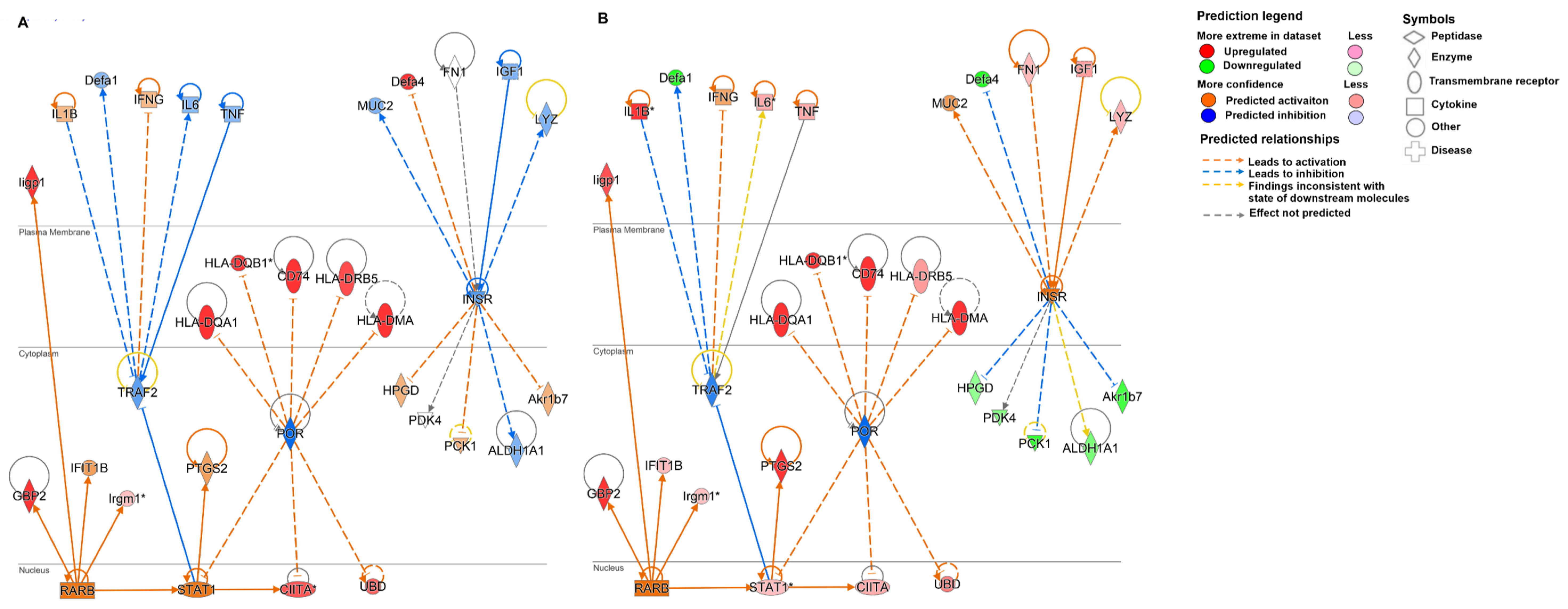
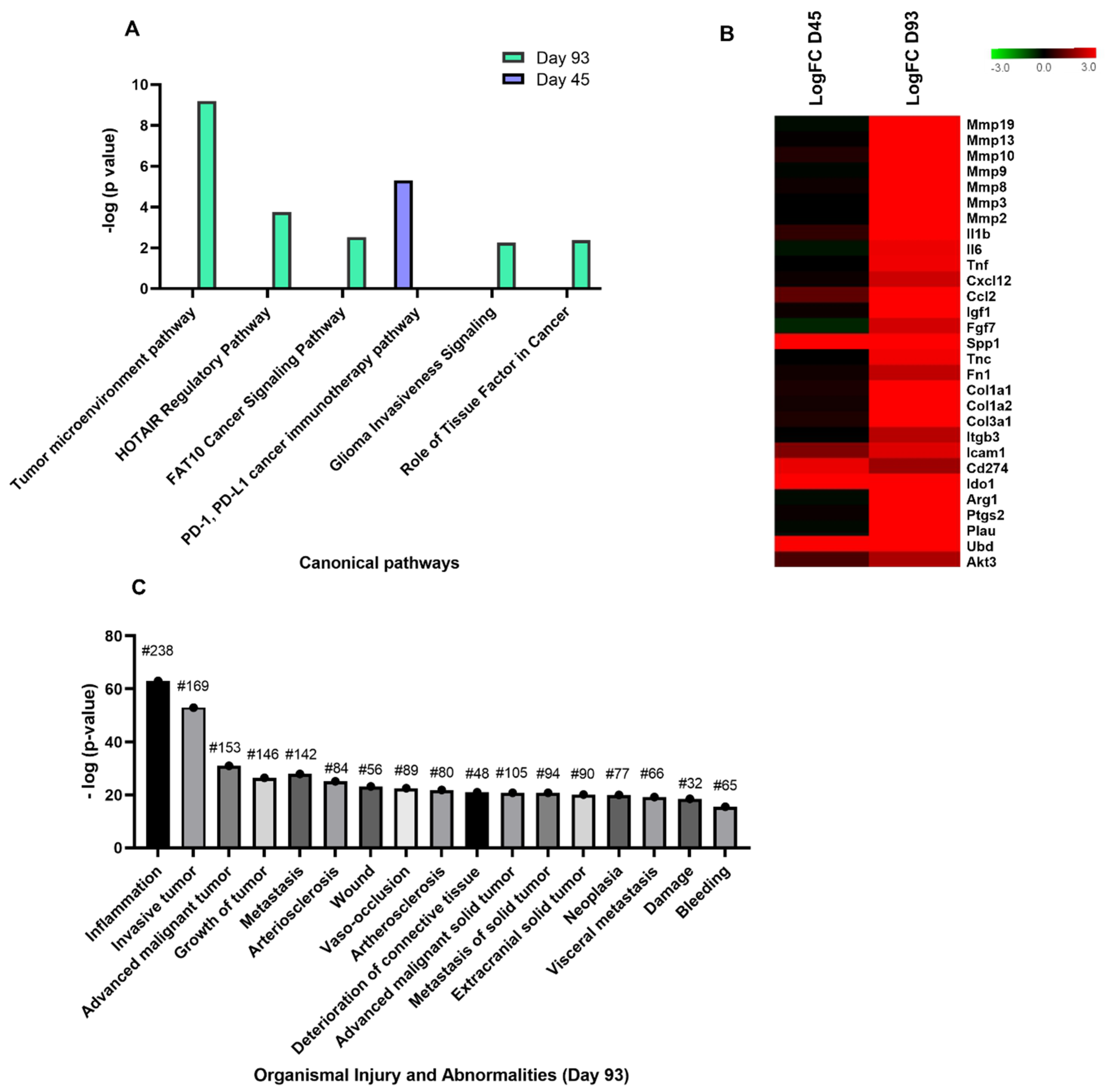
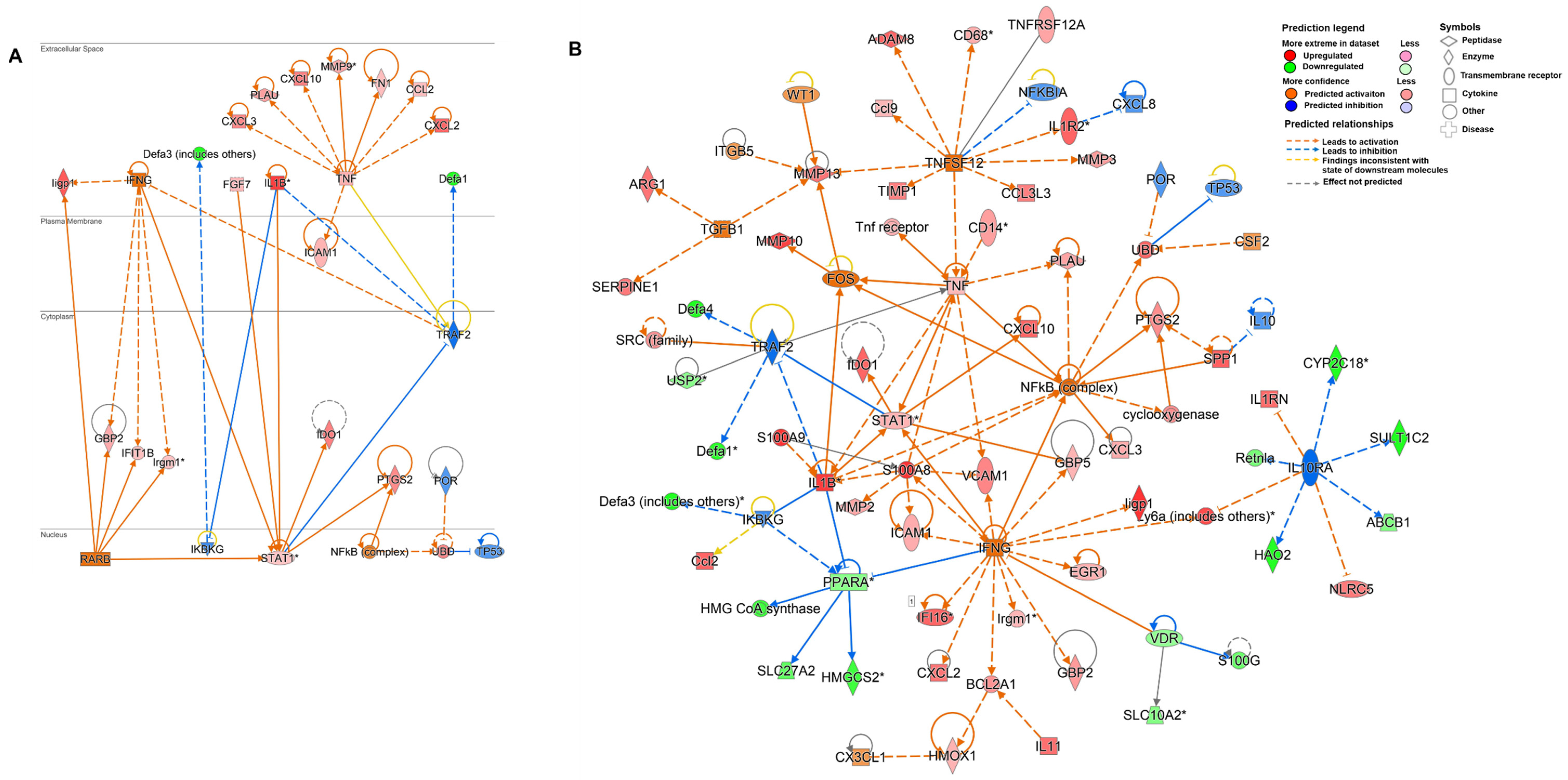
3.4. Chronic Cryptosporidiosis Induces Tumor Microenvironment
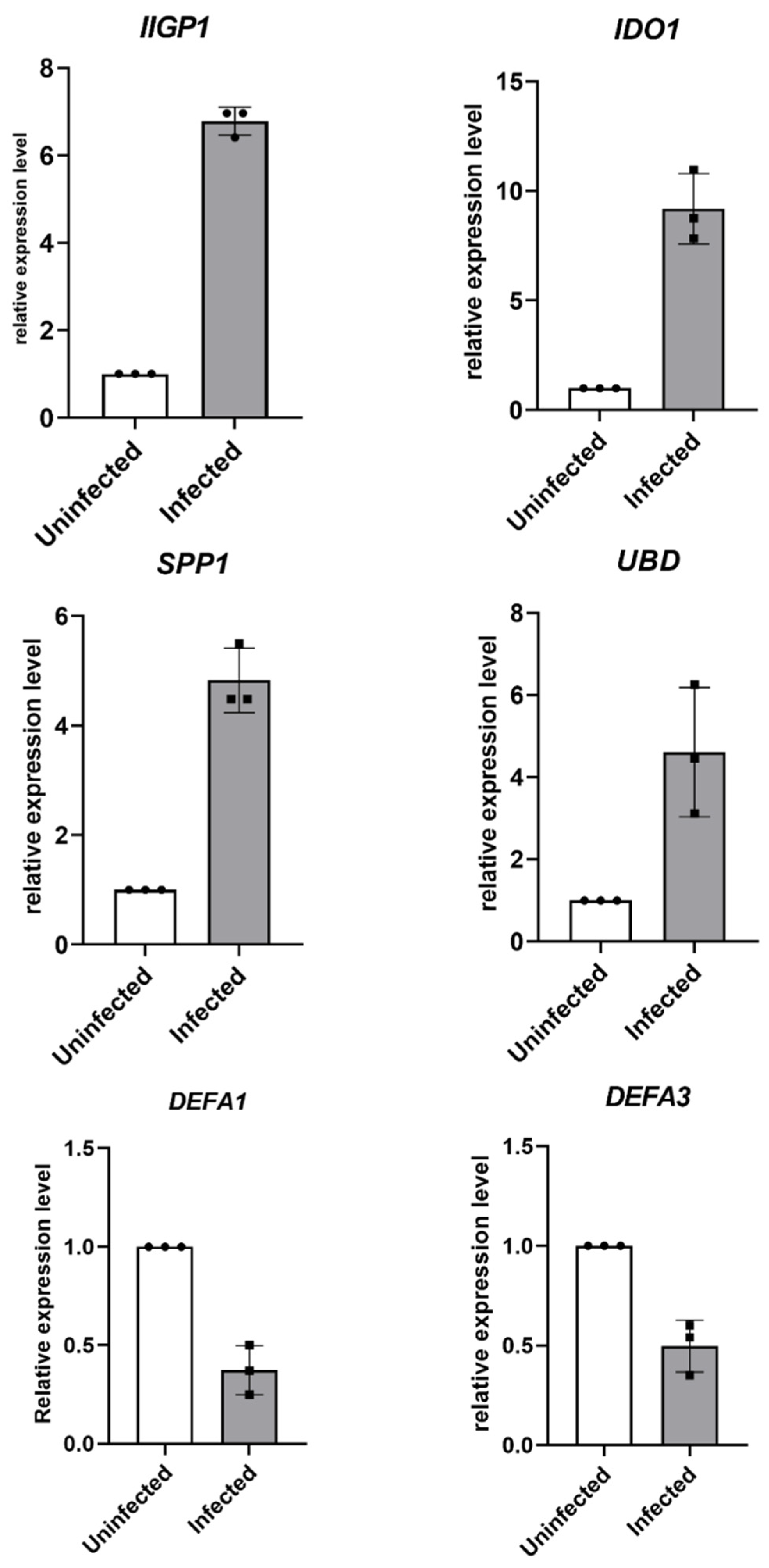
3.5. Validation of Target Genes by RT-qPCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ramirez, N.E.; Ward, L.A.; Sreevatsan, S. A Review of the Biology and Epidemiology of Cryptosporidiosis in Humans and an-imals. Microbes Infect. 2004, 6, 773–785. [Google Scholar] [CrossRef] [PubMed]
- Kotloff, K.L.; Nataro, J.P.; Blackwelder, W.C.; Nasrin, D.; Farag, T.H.; Panchalingam, S.; Wu, Y.; Sow, S.O.; Sur, D.; Breiman, R.F.; et al. Burden and Aetiology of Diarrhoeal Disease in Infants and Young Children in Developing Countries (The Global Enteric Multicenter Study, GEMS): A Prospective, Case-Control Study. Lancet 2013, 382, 209–222. [Google Scholar] [CrossRef]
- Khalil, I.A.; Troeger, C.; Rao, P.C.; Blacker, B.F.; Brown, A.; Brewer, T.G.; Colombara, D.V.; De Hostos, E.L.; Engmann, C.; Guerrant, R.L.; et al. Morbidity, Mortality, and Long-Term Consequences Associated with Diarrhoea from Cryptosporidium Infection in Children Younger Than 5 Years: A Meta-Analyses Study. Lancet Glob. Health 2018, 6, e758–e768. [Google Scholar] [CrossRef] [Green Version]
- Baldursson, S.; Karanis, P. Waterborne Transmission of Protozoan Parasites: Review of Worldwide Outbreaks—An Update 2004–2010. Water Res. 2011, 45, 6603–6614. [Google Scholar] [CrossRef] [PubMed]
- Efstratiou, A.; Ongerth, J.E.; Karanis, P. Waterborne Transmission of Protozoan Parasites: Review of Worldwide Outbreaks—An Update 2011–2016. Water Res. 2017, 114, 14–22. [Google Scholar] [CrossRef]
- CDC Crypto Outbreaks Linked to Swimming Have Doubled since 2014. Available online: https://www.cdc.gov/media/releases/2017/p0518-Cryptosporidium-outbreaks.html (accessed on 1 September 2021).
- Okhuysen, P.C.; Chappell, C.L.; Crabb, J.H.; Sterling, C.R.; Dupont, H.L. Virulence of Three Distinct Cryptosporidium parvum Isolates for Healthy Adults. J. Infect. Dis. 1999, 180, 1275–1281. [Google Scholar] [CrossRef] [Green Version]
- Rowan, N.J. Defining Established and Emerging Microbial Risks in the Aquatic Environment: Current Knowledge, Implications, and Outlooks. Int. J. Microbiol. 2010, 2011, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Yoder, J.S.; Beach, M.J. Cryptosporidium Surveillance and Risk Factors in the United States. Exp. Parasitol. 2010, 124, 31–39. [Google Scholar] [CrossRef]
- Gargala, G. Drug Treatment and Novel Drug Target against Cryptosporidium. Parasite 2008, 15, 275–281. [Google Scholar] [CrossRef] [Green Version]
- Osman, M.; Benamrouz, S.; Guyot, K.; Baydoun, M.; Fréalle, E.; Chabé, M.; Gantois, N.; Delaire, B.; Goffard, A.; Aoun, A.; et al. High Association of Cryptosporidium spp. Infection with Colon Adenocarcinoma in Lebanese patients. PLoS ONE 2017, 12, e0189422. [Google Scholar] [CrossRef]
- Zhang, N.; Yu, X.; Zhang, H.; Cui, L.; Li, X.; Zhang, X.; Gong, P.; Li, J.; Li, Z.; Wang, X.; et al. Prevalence and Genotyping of Cryptosporidium parvum in Gastrointestinal Cancer Patients. J. Cancer 2020, 11, 3334–3339. [Google Scholar] [CrossRef] [Green Version]
- Sulżyc-Bielicka, V.; Kołodziejczyk, L.; Jaczewska, S.; Bielicki, D.; Safranow, K.; Bielicki, P.; Kładny, J.; Rogowski, W. Colorectal Cancer and Cryptosporidium spp. Infection. PLoS ONE 2018, 13, e0195834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopacz, Ż.; Kváč, M.; Karpiński, P.; Hendrich, A.B.; Sąsiadek, M.M.; Leszczyński, P.; Sak, B.; McEvoy, J.; Kicia, M. The First Evidence of Cryptosporidium meleagridis Infection in a Colon Adenocarcinoma from an Immunocompetent Patient. Front. Cell Infect. Microbiol. 2019, 9, 35. [Google Scholar] [CrossRef]
- Shebl, F.M.; Engels, E.A.; Goedert, J.J. Opportunistic Intestinal Infections and Risk of Colorectal Cancer Among People with AIDS. AIDS Res. Hum. Retrovir. 2012, 28, 994–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Certad, G.; Ngouanesavanh, T.; Guyot, K.; Gantois, N.; Chassat, T.; Mouray, A.; Fleurisse, L.; Pinon, A.; Cailliez, J.-C.; Dei-Cas, E.; et al. Cryptosporidium parvum, a Potential Cause of Colic Adenocarcinoma. Infect. Agents Cancer 2007, 2, 22. [Google Scholar] [CrossRef]
- Benamrouz, S.; Guyot, K.; Gazzola, S.; Mouray, A.; Chassat, T.; Delaire, B.; Chabé, M.; Gosset, P.; Viscogliosi, E.; Dei-Cas, E.; et al. Cryptosporidium parvum Infection in SCID Mice Infected with Only One Oocyst: qPCR Assessment of Parasite Replication in Tissues and Development of Digestive Cancer. PLoS ONE 2012, 7, e51232. [Google Scholar] [CrossRef] [PubMed]
- Benamrouz, S.; Conseil, V.; Chabé, M.; Praet, M.; Audebert, C.; Blervaque, R.; Guyot, K.; Gazzola, S.; Mouray, A.; Chassat, T.; et al. Cryptosporidium parvum-Induced Ileo-Caecal Adenocarcinoma and Wnt Signaling in a Mouse Model. Dis. Model. Mech. 2014, 7, 693–700. [Google Scholar]
- Baydoun, M.; Vanneste, S.B.; Creusy, C.; Guyot, K.; Gantois, N.; Chabe, M.; Delaire, B.; Mouray, A.; Baydoun, A.; Forzy, G.; et al. Three-Dimensional (3D) Culture of Adult Murine Colon as an In Vitro Model of Cryptosporidiosis: Proof of Concept. Sci. Rep. 2017, 7, 17288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sablet, T.; Potiron, L.; Marquis, M.; Bussiere, F.I.; Lacroix-Lamande, S.; Laurent, F. Cryptosporidium parvum Increases the in-Testinal Permeability through Interaction with Epithelial Cells and IL-1β and TNF-α Released by Inflammatory Monocytes. Cell Microbiol. 2016, 18, 1871–1880. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.M.; Levine, S.A.; Splinter, P.L.; Tietz, P.S.; Ganong, A.L.; Jobin, C.; Gores, G.J.; Paya, C.V.; LaRusso, N.F. Cryp-Tosporidium Parvum Activates Nuclear Factor kappaB in Biliary Epithelia Preventing Epithelial Cell Apoptosis. Gastroenterology 2001, 120, 1774–1783. [Google Scholar] [CrossRef] [PubMed]
- Currey, N.; Jahan, Z.; Caldon, C.E.; Tran, P.N.; Benthani, F.; De Lacavalerie, P.; Roden, D.L.; Gloss, B.S.; Campos, C.; Bean, E.G.; et al. Mouse Model of Mutated in Colorectal Cancer Gene Deletion Reveals Novel Pathways in Inflammation and Cancer. Cell. Mol. Gastroenterol. Hepatol. 2019, 7, 819–839. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.L.; Fan, X.C.; Li, Y.H.; Yuan, Y.J.; Yin, Y.L.; Wang, X.T.; Zhang, L.X.; Zhao, G.H. Expression Profiles of mRNA and IncRNA in HCT-8 Cells Infected with Cryptosporidium parvum IId Subtype. Front. Microbiol. 2018, 9, 1409. [Google Scholar] [CrossRef]
- He, W.; Li, J.; Gong, A.-Y.; Deng, S.; Li, M.; Wang, Y.; Mathy, N.W.; Feng, Y.; Xiao, L.; Chen, X.M. Cryptosporidial Infection Suppresses Intestinal Epithelial Cell MAPK Signaling Imparing Host Anti-Parasitic Defense. Microorganisms 2021, 9, 151. [Google Scholar] [CrossRef] [PubMed]
- Mary, C.; Chapey, E.; Dutoit, E.; Guyot, K.; Hasseine, L.; Jeddi, F.; Menotti, J.; Paraud, C.; Pomares, C.; Rabodonirina, M.; et al. Multicentric Evaluation of a New Real-Time PCR Assay for Quantification of Cryptosporidium spp. and Identification of Cryptosporidium parvum and Cryptosporidium hominis. J. Clin. Microbiol. 2013, 51, 2556–2563. [Google Scholar] [CrossRef] [Green Version]
- Boivin, G.P.; Washington, K.; Yang, K.; Ward, J.M.; Pretlow, T.P.; Russell, R.; Besselsen, D.G.; Godfrey, V.L.; Doetschman, T.; Dove, W.F.; et al. Pathology of Mouse Models of Intestinal Cancer: Consensus Report and Recommendations. Gastroenterology 2003, 124, 762–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Certad, G.; Creusy, C.; Ngouanesavanh, T.; Guyot, K.; Gantois, N.; Mouray, A.; Chassat, T.; Flament, N.; Fleurisse, L.; Pinon, A.; et al. Development of Cryptosporidium parvum-Induced Gastrointestinal Neoplasia in Severe Combined Immunodeficiency (SCID) Mice: Severity of Lesions Is Correlated with Infection Intensity. Am. J. Trop. Med. Hyg. 2010, 82, 257–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stolte, M. The New Vienna Classification of Epithelial Neoplasia of the Gastrointestinal Tract: Advantages and Disadvantages. Virchows Arch. 2003, 442, 99–106. [Google Scholar] [CrossRef]
- Smyth, G.K.; Yang, Y.H.; Speed, T.; Brownstein, M.J.; Khodursky, A. Statistical Issues in cDNA Microarray Data Analysis. Methods Mol. Biol. 2003, 224, 111–136. [Google Scholar] [CrossRef]
- Ihaka, R.; Gentleman, R. A Language for Data Analysis and Graphics. J. Comput. Graph. Stat. 1996, 5, 299–314. [Google Scholar]
- Yang, Y.H.; Dudoit, S.; Luu, P.; Lin, D.M.; Peng, V.; Ngai, J.; Speed, T.P. Normalization for cDNA Microarray Data: A Robust Composite Method Addressing Single and Multiple Slide Systematic Variation. Nucleic Acids Res. 2002, 30, e15. [Google Scholar] [CrossRef] [Green Version]
- Lonnstedt, I.; Speed, T. Replicated Microarray Data. Stat. Sin. 2002, 12, 31–46. [Google Scholar]
- Thomas, S.; Bonchev, D. A Survey of Current Software for Network Analysis in Molecular Biology. Hum. Genom. 2010, 4, 353–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, A.; Green, J.; Pollard, J.; Tugendreich, S. Casual Analysis Approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(−ΔΔC(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, S.H.; Foster, D.; Sherry, B.; Magness, S.T.; Nielsen, D.M.; Gookin, J.L. Interferon-λ3 Promotes Epithelial Defense and Barrier Function against Cryptosporidium parvum Infection. Cell. Mol. Gastroenterol. Hepatol. 2019, 8, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Hara, S.P.; Chen, X.-M. The Cell Biology of Cryptosporidium Infection. Microbes Infect. 2011, 13, 721–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurent, F.; Lacroix-Lamandé, S. Innate Immune Responses Play a Key Role in Controlling Infection of the Intestinal Epithelium by Cryptosporidium. Int. J. Parasitol. 2017, 47, 711–721. [Google Scholar] [CrossRef]
- Heuberger, C.; Pott, J.; Maloy, K.J. Why Do Intestinal Epithelial Cells Express MHC Class II? Immunology 2021, 162, 357–367. [Google Scholar] [CrossRef]
- Foltz, C.; Napolitano, A.; Khan, R.; Clough, B.; Hirst, E.M.; Frickel, E.M. TRIM21 Is Critical for Survival of Toxoplasma Gondii Infection and Localizes to GBP-Positive Parasite Vacuoles. Sci. Rep. 2017, 7, 5209. [Google Scholar] [CrossRef]
- Guesdon, W.; Auray, G.; Pezier, T.; Bussière, F.I.; Drouet, F.; Le Vern, Y.; Marquis, M.; Potiron, L.; Rabot, S.; Bruneau, A.; et al. CCL20 Displays Antimicrobial Activity against Cryptosporidium parvum, but Its Expression Is Reduced During Infection in the Intestine of Neonatal Mice. J. Infect. Dis. 2015, 212, 1332–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aley, S.B.; Zimmerman, M.; Hetsko, M.; Selsted, M.E.; Gillin, F.D. Killing of Giardia Lamblia by Cryptdins and Cationic Neu-trophil Peptides. Infect. Immun. 1994, 62, 5397–5403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, V.; Pollok, R.C.G.; Dhaliwal, W.; Naik, S.; Farthing, M.J.G.; Bajaj-Elliott, M. A Potential Role for Interleukin-18 in Inhibition of the Development of Cryptosporidium parvum. Clin. Exp. Immunol. 2006, 145, 555–562. [Google Scholar] [CrossRef]
- Ming, Z.; Gong, A.-Y.; Wang, Y.; Zhang, X.-T.; Li, M.; Dolata, C.E.; Chen, X.-M. Trans-Suppression of Defense DEFB1 Gene in Intestinal Epithelial Cells Following Cryptosporidium parvum Infection Is Associated with Host Delivery of Parasite Cdg7_FLc_1000 RNA. Parasitol. Res. 2018, 117, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Dubois, R.N. Role of Prostanoids in Gastrointestinal Cancer. J. Clin. Investig. 2018, 128, 2732–2742. [Google Scholar] [CrossRef]
- Fu, Y.; Liu, S.; Zeng, S.; Shen, H. From Bench to Bed: The Tumor Immune Microenvironment and Current Immunotherapeutic Strategies for Hepatocellular Carcinoma. J. Exp. Clin. Cancer Res. 2019, 38, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofman, P.; Vouret-Craviari, V. Microbes-Induced EMT at the Crossroad of Inflammation and Cancer. Gut Microbes 2012, 3, 176–185. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, H.; Rokudai, S.; Kawabata-Iwakawa, R.; Sakakura, K.; Oyama, T.; Nishiyama, M.; Chikamatsu, K. AKT3 Is a Novel Regulator of Cancer-Associated Fibroblasts in Head and Neck Squamous Cell Carcinoma. Cancers 2021, 13, 1233. [Google Scholar] [CrossRef] [PubMed]
- Ricciardi, M.; Zanotto, M.; Malpeli, G.; Bassi, G.; Perbellini, O.; Chilosi, M.; Bifari, F.; Krampera, M. Epithelial-to-Mesenchymal Transition (EMT) Induced by Inflammatory Priming Elicits Mesenchymal Stromal Cell-like Immune-Modulatory Properties in Cancer Cells. Br. J. Cancer 2015, 112, 1067–1075. [Google Scholar] [CrossRef]
- Nagaraj, S.; Gabrilovich, D.I. Tumor Escape Mechanism Governed by Myeloid-Derived Suppressor Cells. Cancer Res. 2008, 68, 2561–2563. [Google Scholar] [CrossRef] [Green Version]
- Pflügler, S.; Svinka, J.; Scharf, I.; Crncec, I.; Filipits, M.; Charoentong, P.; Tschurtschenthaler, M.; Kenner, L.; Awad, M.; Stift, J.; et al. IDO1+ Paneth Cells Promote Immune Escape of Colorectal Cancer. Commun. Biol. 2020, 3, 252. [Google Scholar] [CrossRef]
- Choudhry, N.; Korbel, D.S.; Edwards, L.A.; Bajaj-Elliott, M.; McDonald, V. Dysregulation of Interferon-Gamma-Mediated Signalling Pathway in Intestinal Epithelial Cells by Cryptosporidium parvum Infection. Cell. Microbiol. 2009, 11, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Peddareddigari, V.G.; Wang, D.; DuBois, R.N. The Tumor Microenvironment in Colorectal Carcinogenesis. Cancer Microenviron. 2010, 3, 149–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solinas, G.; Germano, G.; Mantovani, A.; Allavena, P. Tumor-Associated Macrophages (TAM) as Major Players of the Cancer-Related Inflammation. J. Leukoc. Biol. 2009, 86, 1065–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shurin, M.R. Osteopontin Controls Immunosuppression in the Tumor Microenvironment. J. Clin. Investig. 2018, 128, 5209–5212. [Google Scholar] [CrossRef]
- Eberhart, C.E.; Coffey, R.J.; Radhika, A.; Giardiello, F.M.; Ferrenbach, S.; DuBois, R.N. Up-Regulation of Cyclooxygenase 2 Gene Expression in Human Colorectal Adenomas and Adenocarcinomas. Gastroenterology 1994, 107, 1183–1188. [Google Scholar] [CrossRef]
- Wang, D.; Dubois, R.N. Immunosuppression Associated with Chronic Inflammation in the Tumor Microenvironment. Carcinogenesis 2015, 36, 1085–1093. [Google Scholar] [CrossRef] [Green Version]
- Yan, D.-W.; Li, D.-W.; Yang, Y.-X.; Xia, J.; Wang, X.-L.; Zhou, C.-Z.; Fan, J.-W.; Wen, Y.-G.; Sun, H.-C.; Wang, Q.; et al. Ubiquitin D Is Correlated with Colon Cancer Progression and Predicts Recurrence for Stage II-III Disease after Curative Surgery. Br. J. Cancer 2010, 103, 961–969. [Google Scholar] [CrossRef] [Green Version]
- Barriga, V.; Kuol, N.; Nurgali, K.; Apostolopoulos, V. The Complex Interaction between the Tumor Micro-Environment and Immune Checkpoints in Breast Cancer. Cancers 2019, 11, 1205. [Google Scholar] [CrossRef] [Green Version]
- Fang, K.; Bruce, M.; Pattillo, C.B.; Zhang, S.; Stone, R.; Clifford, J.; Kevil, C.G. Temporal Genome Wide Expression Profiling of DSS Colitis Reveals Novel Inflammatory and Angiogenesis Genes Similar to Ulcerative Colitis. Physiol. Genom. 2011, 43, 43–56. [Google Scholar] [CrossRef]
- Spandidos, A.; Wang, X.; Wang, H.; Brian, S. PrimerBank: A resource of human and mouse PCR primer pairs for gene expression detection and quantification. Nucl. Acids Res. 2010, 38, D792–D799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spandidos, A.; Wang, X.; Wang, H.; Dragnev, S.; Thurber, T.; Brian, S. A comprehensive collection of experimentally validated primers for Polymerase Chain Reaction quantitation of murine transcript abundance. BMC Genomics. 2008, 9, 633. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Brain, S. A PCR primer bank for quantitative gene expression analysis. Nucl. Acids Res. 2003, 31, e154. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Yokoi, Y.; Fukaya, R.; Ohira, S.; Shinozaki, R.; Nishida, T.; Kikuchi, M.; Ayabe, T. Expression and localization of Paneth cells and their α-defensins in the small intestine of adult mouse. Front. Immunol. 2020, 11, 570296. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbols and Names | Direction of Regulation | LogFC Day 45 PI | LogFC Day 93 PI |
|---|---|---|---|
| Iigp1; interferon inducible GTPase 1 | Up | 6.78 | 5.74 |
| Defa4; defensin, alpha, 4 | Up (day 45 PI) Down (day 93 PI) | 6.37 | −6.07 |
| H2-DMb1;histocompatibility 2, class II, locus Mb1 | Up | 6.35 | 3.20 |
| Tgtp2; T cell specific GTPase 2 | Up | 6.31 | 5.13 |
| Cd74; CD74 antigen | Up | 6.23 | 2.99 |
| H2-Ab1; histocompatibility 2, class II antigen A, β | Up | 6.21 | 2.91 |
| H2-Eb1; histocompatibility 2, class II antigen E, β | Up | 5.89 | 3.27 |
| Ciita; class II transactivator | Up | 5.63 | 2.73 |
| H2-Aa; histocompatibility 2, class II antigen A, α | Up | 5.31 | 2.56 |
| UBD; ubiquitin D | Up | 5.15 | 4.25 |
| Ido1; indoleamine 2,3-dioxygenase 1 | Up | 5.07 | 4.41 |
| Cxcl10; chemokine (C-X-C motif) ligand 10 | Up | 4.56 | 4.44 |
| Gbp11; guanylate binding protein 11 | Up | 4.39 | 3.11 |
| Cxcl9; chemokine (C-X-C motif) ligand 9 | Up | 4.38 | 3.68 |
| Gbp4; guanylate binding protein 4 | Up | 4.14 | 2.81 |
| Gbp2b; guanylate binding protein 2b | Up | 3.90 | 2.93 |
| Gbp6; guanylate binding protein 6 | Up | 3.90 | 3.20 |
| Il18bp; interleukin 18 binding protein | Up | 3.68 | 3.21 |
| Gbp8; guanylate-binding protein 8 | Up | 3.60 | 2.06 |
| Ly6a; lymphocyte antigen 6 complex, locus A | Up | 3.35 | 5.52 |
| Igtp; interferon gamma induced GTPase | Up | 3.37 | 2.90 |
| Gbp2; guanylate binding protein 2 | Up | 3.32 | 2.92 |
| Spp1; secreted phosphoprotein 1 | Up | 3.04 | 5.07 |
| Ly6e; lymphocyte antigen 6 complex, locus E | Up | 2.13 | 5.25 |
| S100a9; calgranulin B | Up | N/A a | 9.14 |
| S100a8; calgranulin A | Up | N/A a | 8.42 |
| Mmp10; matrix metallopeptidase 10 | Up | N/A a | 6.76 |
| Il1b; interleukin 1 beta | Up | N/A a | 6.74 |
| Defa2; defensin, alpha, 2 | Down | N/A a | −6.02 |
| Il1rl1; interleukin 1 receptor-like 1 | Up | N/A a | 5.84 |
| Ifi44l; interferon-induced protein 44 like | Up | N/A a | 5.77 |
| Arg1; arginase | Up | N/A a | 5.34 |
| Defa3; defensin, alpha, 3 | Down | N/A a | −5.76 |
| Mmp8; matrix metallopeptidase 8 | Up | N/A a | 5.74 |
| Il11; interleukin 11 | Up | N/A a | 5.74 |
| Mmp13; matrix metallopeptidase 13 | Up | N/A a | 5.49 |
| Defa1; defensin, alpha 1 | Down | N/A a | −5.76 |
| Ifi202b; interferon activated gene 202B | Up | N/A a | 5.27 |
| Cxcl1; chemokine (C-X-C motif) ligand 1 | Up | N/A a | 5.27 |
| Ccl3; chemokine (C-C motif) ligand 3 | Up | N/A a | 5.23 |
| Ccl2; chemokine (C-C motif) ligand 2 | Up | N/A a | 5.19 |
| Slc37a2; solute carrier family 37 (glycerol-3-phosphate transporter), member 2 | Down | N/A a | −4.50 |
| Col1a1; collagen, type I, alpha 1 | Up | N/A a | 4.87 |
| Il1r2; interleukin 1 receptor, type II | Up | N/A a | 4.95 |
| Adam8; a dis integrin and metallopeptidase domain 8 | Up | N/A a | 4.89 |
| Cxcl5; chemokine (C-X-C motif) ligand 5 | Up | N/A a | 4.87 |
| Sult1c2; sulfotransferase family, cytosolic, 1C, member 2 | Down | N/A a | −4.68 |
| Ifit2; interferon-induced protein with tetratricopeptide repeats 2 | Up | N/A a | 4.63 |
| Mmp3; matrix metallopeptidase 3 | Up | N/A a | 4.58 |
| Cyp2c40; cytochrome P450, family 2, subfamily c, polypeptide 40 | Down | N/A a | −4.33 |
| Cxcl2; chemokine (C-X-C motif) ligand 2 | Up | N/A a | 4.14 |
| Col1a2; collagen, type I, alpha 2 | Up | N/A a | 4.14 |
| Ptgs2; prostaglandin-endoperoxide synthase 2 | Up | N/A a | 4.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sawant, M.; Benamrouz-Vanneste, S.; Mouray, A.; Bouquet, P.; Gantois, N.; Creusy, C.; Duval, E.; Mihalache, A.; Gosset, P.; Chabé, M.; et al. Persistent Cryptosporidium parvum Infection Leads to the Development of the Tumor Microenvironment in an Experimental Mouse Model: Results of a Microarray Approach. Microorganisms 2021, 9, 2569. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9122569
Sawant M, Benamrouz-Vanneste S, Mouray A, Bouquet P, Gantois N, Creusy C, Duval E, Mihalache A, Gosset P, Chabé M, et al. Persistent Cryptosporidium parvum Infection Leads to the Development of the Tumor Microenvironment in an Experimental Mouse Model: Results of a Microarray Approach. Microorganisms. 2021; 9(12):2569. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9122569
Chicago/Turabian StyleSawant, Manasi, Sadia Benamrouz-Vanneste, Anthony Mouray, Peggy Bouquet, Nausicaa Gantois, Colette Creusy, Erika Duval, Adriana Mihalache, Pierre Gosset, Magali Chabé, and et al. 2021. "Persistent Cryptosporidium parvum Infection Leads to the Development of the Tumor Microenvironment in an Experimental Mouse Model: Results of a Microarray Approach" Microorganisms 9, no. 12: 2569. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9122569