
Error Distribution Model to Standardize LPUE, CPUE and Survey-Derived Catch Rates of Target and Non-Target Species

 , ,
, ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Datasets
2.2. Statistical Models
- Lognormal. This distribution assumed that the logarithm of the catch rate was normally distributed, using the identity as the link function. This error distribution also required the addition of a positive constant (1 or c) to deal with zero catches.
- Tweedie. This distribution is part of the exponential family of distributions and is defined by a mean (μ) and variance (φμp), in which φ is the dispersion parameter and p is an index parameter [22]. Given that this distribution can handle a certain proportion of zeros, the nominal catches were used directly. The power parameter (p) of the variance function was calculated by maximum likelihood estimation.
- Hurdle models. The catch estimates involved fitting two sub-models to the data [19,35]. The first sub-model modeled the probability that a positive observation (non-zero catch) occurred, assuming a binomial error distribution and logit link function. Positive observations were analyzed using a second sub-model assuming (1) a lognormal (hurdle–lognormal) error distribution with an identity link function and log-transformed catch rates, and (2) a gamma (hurdle–gamma) error distribution with a log link function.
- LPUE ~ Year + Quarter + Vessel + Métier + Target + Year × Quarter + Year × Vessel + Year × Métier + Year × Target
- CPUE ~ Year + Quarter + Vessel + Gear + Depth + Target + Year × Quarter + Year × Vessel + Year × Gear + Year × Target
- RPN ~ Year + Month + Moon + Area + Depth + Soak time + Year × Month + Year × Moon + Year × Area + Year × Soak time
2.3. Error–Model Selection (Methodology)
- Pearson residuals were plotted against the fitted values as a check of the assumed variance function;
- Standardized deviance residuals were plotted against the estimated linear predictor () to check for systematic deviations from the assumptions underlying the error distribution; and
- The dependent variable was plotted against the estimated linear predictor () as a check of the assumed link function.
2.4. Standardization Procedure
2.5. Catch Trend Comparison between Datasets
3. Results and Discussion
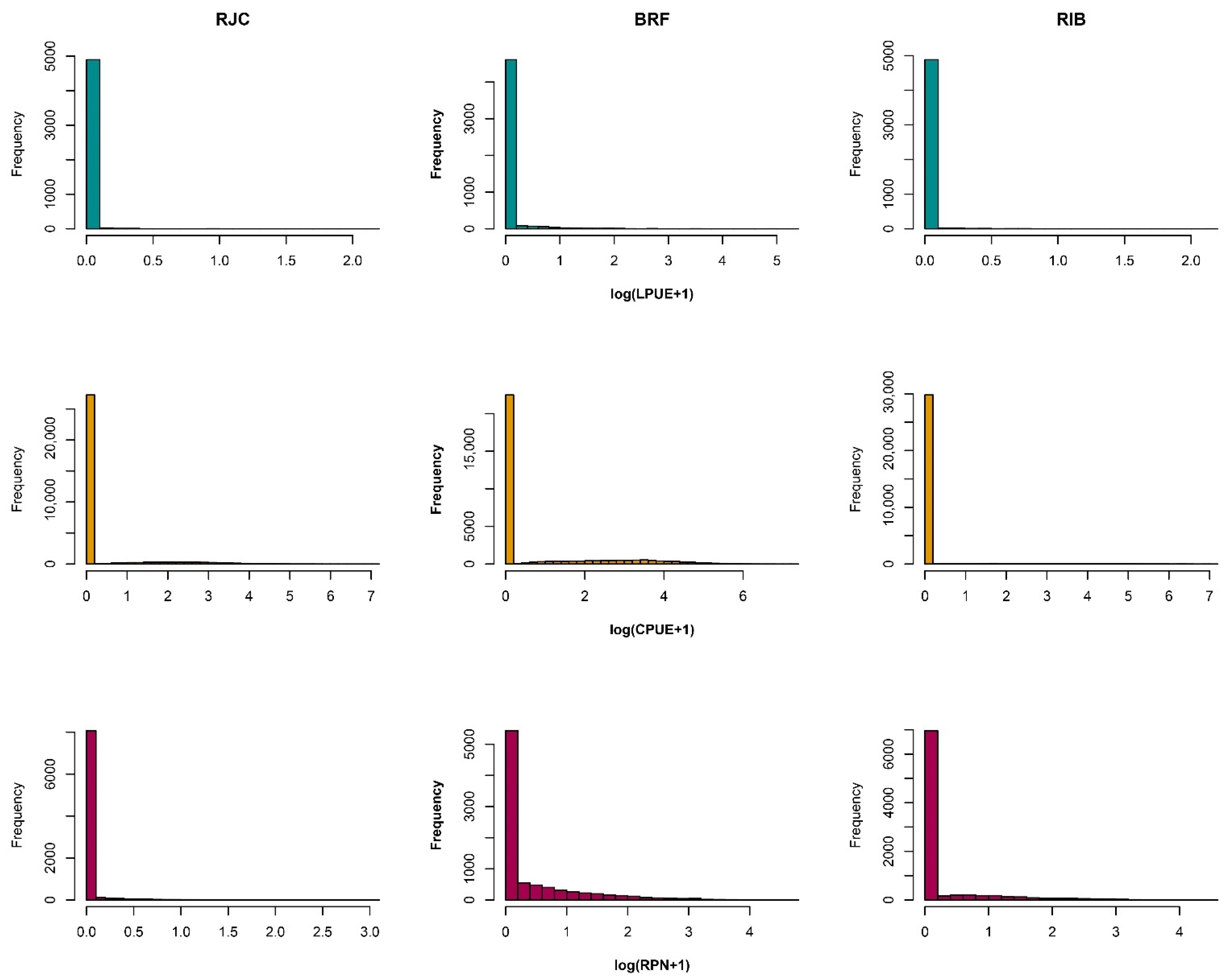
3.1. Nominal Catch Data
3.2. Error–Model Selection (Application)
3.3. Standardization Procedure
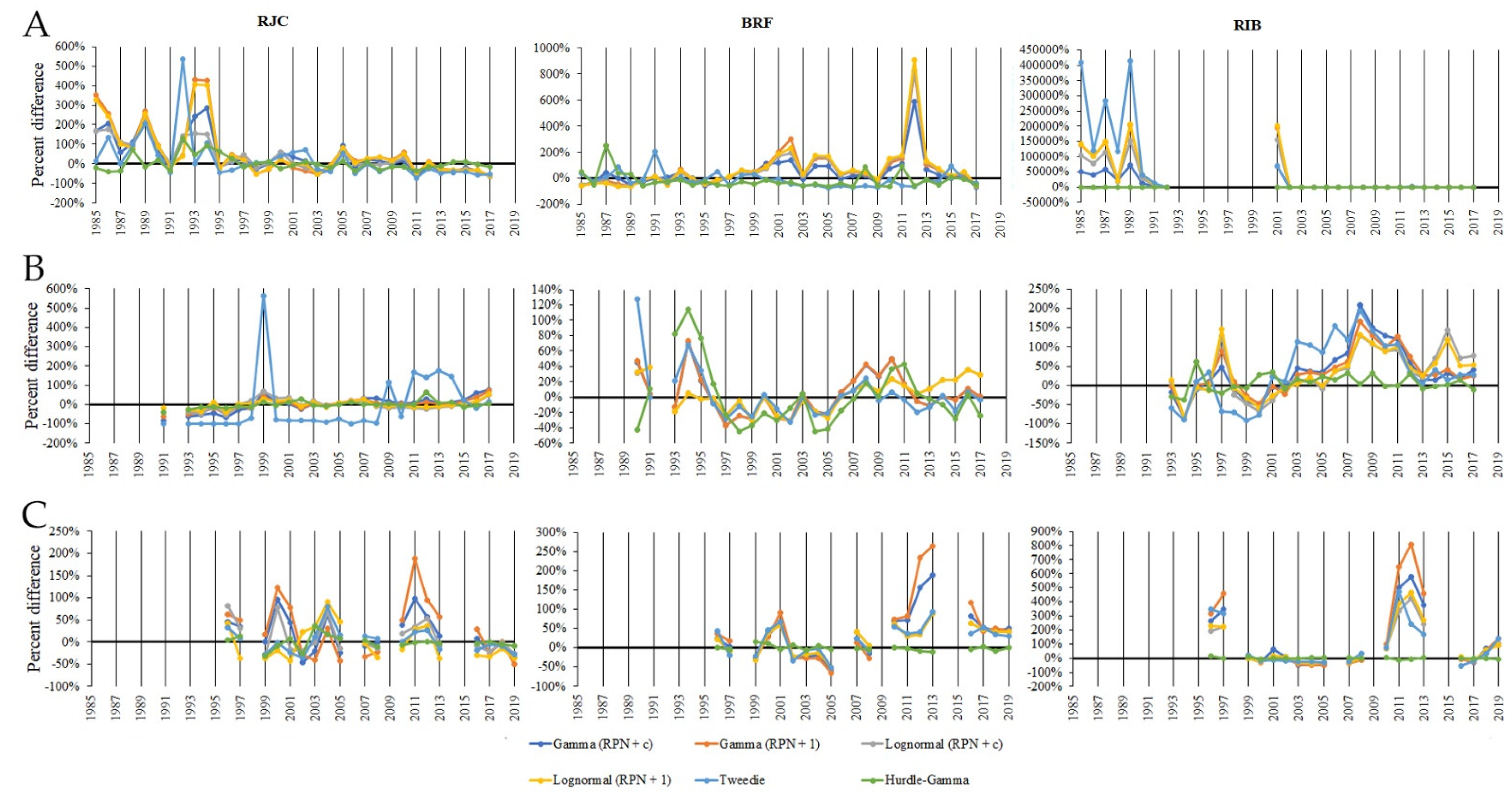
3.4. Consequences of Choosing a Wrong Error–Model
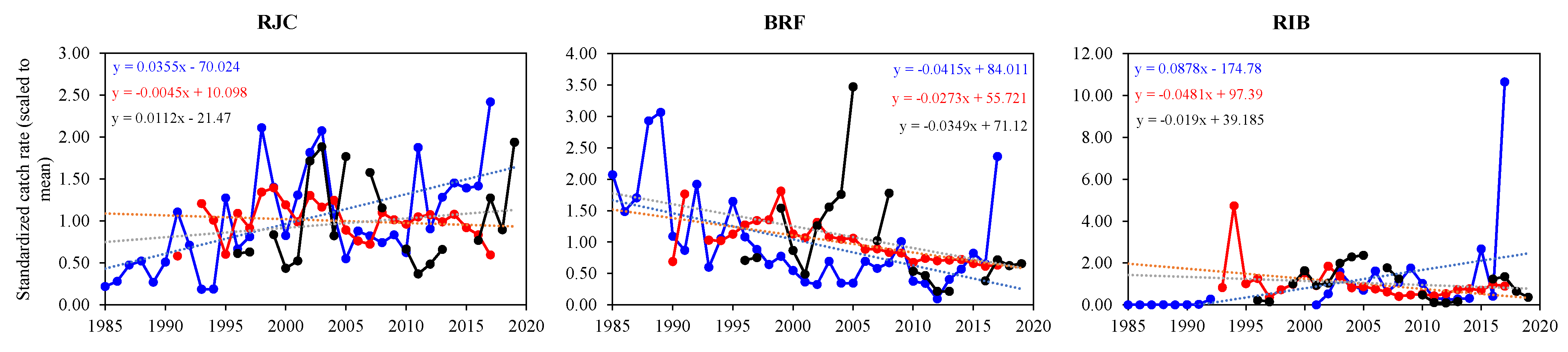
3.5. Catch Trend Comparison between Datasets
3.6. Final Considerations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hilborn, R.; Walters, C.J. Quantitative Fisheries Stock Assessment: Choice, Dynamics and Uncertainty; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013; ISBN 9781461535980. [Google Scholar]
- ICES Guide to ICES advisory framework and principles. Rep. ICES Advis. Comm. 2020, 1–8. [CrossRef]
- Cadrin, S.X.; Dickey-Collas, M. Stock assessment methods for sustainable fisheries. ICES J. Mar. Sci. 2015, 72, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Cadima, E.L. Fish Stock Assessment Manual; FAO Fisher; FAO: Rome, Italy, 2003; ISBN 9251045054. [Google Scholar]
- Sparre, P.; Venema, S.C. Introduction to Tropical Fish Stock Assessment. Part 1 Manual. Part 2 Exercises; FAO: Rome, Italy, 1998; pp. 1–423. [Google Scholar]
- Gulland, J.A. Fish population analysis. In Manual of Methods for Fish Stock Assessment; FAO: Rome, Italy, 1969; Volume 4. [Google Scholar]
- Quinn, T.J., II; Hoag, S.H.; Southward, G.M. Comparison of two methods of combining catch-per-unit-effort data from geographic regions. Can. J. Fish. Aquat. Sci. 1982, 39, 837–846. [Google Scholar] [CrossRef]
- Maunder, M.N.; Punt, A.E. Standardizing catch and effort data: A review of recent approaches. Fish. Res. 2004, 70, 141–159. [Google Scholar] [CrossRef]
- Campbell, R.A. CPUE standardisation and the construction of indices of stock abundance in a spatially varying fishery using general linear models. Fish. Res. 2004, 70, 209–227. [Google Scholar] [CrossRef]
- Pinho, M.; Medeiros-Leal, W.; Sigler, M.; Santos, R.; Novoa-Pabon, A.; Menezes, G.; Silva, H. Azorean demersal longline survey abundance estimates: Procedures and variability. Reg. Stud. Mar. Sci. 2020, 39, 101443. [Google Scholar] [CrossRef]
- Garrod, D.J. Effective fishing effort and the catchability coefficient q. Rapport et process verbaux des réunions du Conseil International pour l’Exploration de la Mer 1964, 155, 66–70. [Google Scholar]
- Bishop, J. Standardizing fishery-dependent catch and effort data in complex fisheries with technology change. Rev. Fish Biol. Fish. 2006, 16, 21–38. [Google Scholar] [CrossRef]
- Ye, Y.; Dennis, D. How reliable are the abundance indices derived from commercial catch—Effort standardization? Can. J. Fish. Aquat. Sci. 2009, 66, 1169–1178. [Google Scholar] [CrossRef]
- Maunder, M.N. A general framework for integrating the standardization of catch per unit of effort into stock assessment models. Can. J. Fish. Aquat. Sci. 2001, 58, 795–803. [Google Scholar] [CrossRef]
- Nelder, J.A.; Wedderburn, R.W.M. Generalized linear models. J. R. Stat. Soc. Ser. A 1972, 135, 370–384. [Google Scholar] [CrossRef]
- McCullagh, P.; Nelder, J.A. Generalized Linear Models, 2nd ed.; Chapman and Hall: London, UK, 1989. [Google Scholar]
- Jorgensen, B. The Theory of Dispersion Models; CRC Press: Boca Rato, FL, USA, 1997. [Google Scholar]
- Jorgensen, B. Exponential Dispersion Models. J. R. Stat. Soc. Ser. B 1987, 49, 127–162. [Google Scholar] [CrossRef]
- Lo, N.C.; Jacobson, L.D.; Squire, J.L. Indices of Relative Abundance from Fish Spotter Data based on Delta-Lognornial Models. Can. J. Fish. Aquat. Sci. 1992, 49, 2515–2526. [Google Scholar] [CrossRef]
- Stefánsson, G. Analysis of groundfish survey abundance data: Combining the GLM and delta approaches. ICES J. Mar. Sci. 1996, 53, 577–588. [Google Scholar] [CrossRef]
- Ortiz, M.; Arocha, F. Alternative error distribution models for standardization of catch rates of non-target species from a pelagic longline fishery: Billfish species in the Venezuelan tuna longline fishery. Fish. Res. 2004, 70, 275–297. [Google Scholar] [CrossRef]
- Shono, H. Application of the Tweedie distribution to zero-catch data in CPUE analysis. Fish. Res. 2008, 93, 154–162. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, F.C.; Murie, D.J.; Hazin, H.G.; Leite-Mourato, B.; Travassos, P.; Burgess, G.H. Catch rates and size composition of blue sharks (Prionace glauca) caught by the Brazilian pelagic longline fleet in the southwestern Atlantic Ocean. Aquat. Living Resour. 2010, 23, 373–385. [Google Scholar] [CrossRef] [Green Version]
- Pons, M.; Domingo, A.; Sales, G.; Fiedler, F.N.; Miller, P.; Giffoni, B.; Ortiz, M. Standardization of CPUE of loggerhead sea turtle (Caretta caretta) caught by pelagic longliners in the Southwestern Atlantic Ocean. Aquat. Living Resour. 2010, 23, 65–75. [Google Scholar] [CrossRef]
- Thorson, J.T.; Cunningham, C.J.; Jorgensen, E.; Havron, A.; Hulson, P.J.F.; Monnahan, C.C.; von Szalay, P. The surprising sensitivity of index scale to delta-model assumptions: Recommendations for model-based index standardization. Fish. Res. 2021, 233, 105745. [Google Scholar] [CrossRef]
- Simpfendorfer, C.A.; Hueter, R.E.; Bergman, U.; Connett, S.M.H. Results of a fishery-independent survey for pelagic sharks in the western North Atlantic, 1977–1994. Fish. Res. 2002, 55, 175–192. [Google Scholar] [CrossRef]
- EU Council Regulation (EC) No 199/2008 of 25 February 2008 concerning the establishment of a Community framework for the collection, management and use of data in the fisheries sector and support for scientific advice regarding the Common Fisheries Policy. Off. J. Eur. Union L 2008, 60, 1–12.
- Menezes, G.M.; Sigler, M.F.; Silva, H.M.; Pinho, M.R. Structure and zonation of demersal fish assemblages off the Azores Archipelago (mid-Atlantic). Mar. Ecol. Prog. Ser. 2006, 324, 241–260. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.V.S.; Silva, W.M.M.L.; Novoa-Pabon, A.M.; Silva, H.M.; Pinho, M.R. Long-term changes in the diversity, abundance and size composition of deep sea demersal teleosts from the Azores assessed through surveys and commercial landings. Aquat. Living Resour. 2019, 32, 25. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.; Novoa-Pabon, A.; Silva, H.; Pinho, M. Elasmobranch species richness, fisheries, abundance and size composition in the Azores archipelago (NE Atlantic). Mar. Biol. Res. 2020, 16, 103–116. [Google Scholar] [CrossRef]
- Santos, R.; Pabon, A.; Silva, W.; Silva, H.; Pinho, M. Population structure and movement patterns of blackbelly rosefish in the NE Atlantic Ocean (Azores archipelago). Fish. Oceanogr. 2020, 29, 227–237. [Google Scholar] [CrossRef]
- Santos, R.; Medeiros-Leal, W.; Pinho, M. Stock assessment prioritization in the Azores: Procedures, current challenges and recommendations. Arquipelago Life Mar. Sci. 2020, 37, 45–64. [Google Scholar]
- Santos, R.; Medeiros-Leal, W.; Pinho, M. Synopsis of biological, ecological and fisheries-related information on priority marine species in the Azores region. Arquipelag. Life Mar. Sci. 2020, 1, 1–138. [Google Scholar]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer Science & Business Media: New York, NY, USA, 2009; ISBN 978-0-387-98957-0. [Google Scholar]
- Zuur, A.F.; Ieno, E.N. Beginner′s Guide to Zero-Inflated Models with R; Highland Statistics Ltd.: Newburgh, UK, 2016; ISBN 978-0-9571741-8-4. [Google Scholar]
- Ortiz, M.; Legault, C.M.; Ehrhardt, N.M. An alternative method for estimating bycatch from the U.S. shrimp trawl fi shery in the Gulf of Mexico, 1972–1995. Fish. Bull. 2000, 98, 583–599. [Google Scholar]
- Cooke, J.G. A procedure for using catch-effort indices in bluefin tuna assessments. Collect. Vol. Sci. Pap. ICCAT 1997, 46, 228–232. [Google Scholar]
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-PLUS; Springer: New York, NY, USA, 2000; ISBN 978-0-387-98957-0. [Google Scholar]
- Walter, J.; Ortiz, M. Derivation of the delta-lognormal variance estimator and recommendation for approximating variances for two-stage cpue standardization models. Collect. Vol. Sci. Pap. ICCAT 2012, 68, 365–369. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Prentice-Hall/Pearson: Teaneck, NJ, USA, 2010. [Google Scholar]
- McDonald, J.H. Handbook of Biological Statistics—Paired t-test. Sparky House Publ. 2014, 180–185. [Google Scholar]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020.
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Sarkar, D. Lattice: Multivariate Data Visualization with R; Springer: New York, NY, USA, 2008. [Google Scholar]
- Lenth, R. V Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Dunn, P.K. Tweedie: Evaluation of Tweedie Exponential Family Models, R package version 2.3; 2017. [Google Scholar]
- Pennington, M. Estimating the mean and variance from highly skewed marine data. Fish. Bull. 1996, 94, 498–505. [Google Scholar]
- Santos, R.; Medeiros-Leal, W.; Novoa-Pabon, A.; Crespo, O.; Pinho, M. Biological Knowledge of Thornback Ray (Raja clavata) from the Azores: Improving Scientific Information for the Effectiveness of Species-Specific Management Measures. Biology 2021, 10, 676. [Google Scholar] [CrossRef]
- Santos, R.; Medeiros-Leal, W.; Crespo, O.; Novoa-Pabon, A.; Pinho, M. Contributions to Management Strategies in the NE Atlantic Regarding the Life History and Population Structure of a Key Deep-Sea Fish (Mora moro). Biology 2021, 10, 522. [Google Scholar] [CrossRef]
- Santos, R.; Medeiros-Leal, W.; Novoa-Pabon, A.; Silva, H.; Pinho, M. Demersal fish assemblages on seamounts exploited by fishing in the Azores (NE Atlantic). J. Appl. Ichthyol. 2021, 37, 198–215. [Google Scholar] [CrossRef]
- Ono, K.; Punt, A.E.; Hilborn, R. Think outside the grids: An objective approach to define spatial strata for catch and effort analysis. Fish. Res. 2015, 170, 89–101. [Google Scholar] [CrossRef]
- Potts, S.E.; Rose, K.A. Evaluation of GLM and GAM for estimating population indices from fishery independent surveys. Fish. Res. 2018, 208, 167–178. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Landing Reports | Fishing Inquiries | Scientific Survey | ||||||
|---|---|---|---|---|---|---|---|---|
| Variable | Type | Observations | Variable | Type | Observations | Variable | Type | Observations |
| Year | Categorical (33) | Period: 1985–2017 | Year | Categorical (27) | Period: 1990–2017 | Year | Categorical (19) | Period: 1996–2019 (except 1998, 2006, 2009, 2014 and 2015) |
| Quarter | Categorical (4) | 1: January–March | Quarter | Categorical (4) | 1: January–March | |||
| 2: April–June | 2: April–June | Month | Categorical (5) | March | ||||
| 3: July–September | 3: July–September | April | ||||||
| 4: October–December | 4: October–December | May | ||||||
| Vessel length | Categorical (5) | 1: ≤10 m | Vessel length | Categorical (5) | 1: ≤10 m | June | ||
| 2: >10 and ≤12 m | 2: >10 and ≤12 m | July | ||||||
| 3: >12 and ≤18 m | 3: >12 and ≤18 m | Moon | Categorical (4) | 1: New moon | ||||
| 4: >18 and ≤24 m | 4: >18 and ≤24 m | 2: First quarter | ||||||
| 5: >24 and ≤40 m | 5: >24 and ≤40 m | 3: Full moon | ||||||
| Métier | Categorical (12) | HDP: hand picking | Gear | Categorical (5) | LL: Longlines | 4: Last quarter | ||
| HUN: species removal by hunting | HL: Handlines | Area | Categorical (10) | AÇO: Açores bank | ||||
| FPO_CRU: pots and traps for crustaceans | NT: Nets | PAL: Princess Alice bank | ||||||
| FPO_FIF: pots and traps for fish | TP: Traps and pots | FPI: Faial and Pico | ||||||
| GNS_FIF: gillnets for coastal demersal and pelagic fish | MG: Multigear | GRA: Graciosa | ||||||
| LHP_CEP: handlines for cephalopods—squids | Depth (mean depth of fishing operation) | Categorical (3) | 1: Shallow (<200 m) | SJO: São Jorge | ||||
| LHP_FIF: handlines for demersal fish | 2: Intermediate (200–600 m) | TER: Terceira | ||||||
| LHP_MDP: handlines locally called “corrico” for pelagic fish | 3: Deep (>600 m) | SMA: Santa Maria | ||||||
| LHP_LPF (pole and lines for pelagic fish) | Target effect (percentage of species-specific catch related to the total catch) | Categorical (4) | 1: 1st quartile (≤25%) | SMI: São Miguel | ||||
| LLD: drifting longlines for pelagic and demersal fish | 2: 2nd quartile (>25% and ≤50%) | MPR: Mar da Prata bank | ||||||
| LLS_DEF: set longlines for pelagic and demersal fish | 3: 3rd quartile (>50% and ≤75%) | FCO: Flores and Corvo | ||||||
| PS_SPF: purse seines for small pelagic fish | 4: 4th quartile (>75%) | Depth | Categorical (24) | from 0 to 1200 m by 50 m intervals (1: 0–50 m, 2: 50–100 m, …, 24: 1150–1200 m) | ||||
| Target effect (percentage of species-specific catch related to the total catch) | Categorical (4) | 1: 1st quartile (≤ 25%) | ||||||
| 2: 2nd quartile (> 25% and ≤ 50%) | Soak time (time during which the hooks were in the water) | Categorical (8) | Time expressed in hours from 2 to 8 by 1 h intervals (2: ≥1.5 and <2.5, 3: ≥2.5 and <3.5, …, 8: ≥7.5 and <8.5) | |||||
| 3: 3rd quartile (> 50% and ≤ 75%) | ||||||||
| 4: 4th quartile (> 75%) | ||||||||
| RJC | BRF | RIB | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Estimate | Std. Error | t | p | Estimate | Std. Error | t | p | Estimate | Std. Error | t | p | |
| Intercept | ||||||||||||
| LPUE | −70.024 | 15.933 | −4.395 | <0.001 | 84.011 | 21.019 | 3.997 | <0.001 | −174.777 | 50.969 | −3.429 | 0.001 |
| CPUE | 80.122 | 27.675 | 2.895 | 0.005 | −28.290 | 35.019 | −0.808 | 0.422 | 272.167 | 89.886 | 3.028 | 0.004 |
| RPN | 48.554 | 32.460 | 1.496 | 0.139 | −12.891 | 42.821 | −0.301 | 0.764 | 213.962 | 100.356 | 2.132 | 0.037 |
| Slope | ||||||||||||
| LPUE | 0.035 | 0.008 | 4.458 | <0.001 | −0.041 | 0.011 | −3.949 | <0.001 | 0.088 | 0.025 | 3.449 | 0.001 |
| CPUE | −0.040 | 0.014 | −2.898 | 0.005 | 0.014 | 0.017 | 0.811 | 0.420 | −0.136 | 0.045 | −3.029 | 0.004 |
| RPN | −0.024 | 0.016 | −1.502 | 0.138 | 0.007 | 0.021 | 0.307 | 0.760 | −0.107 | 0.050 | −2.135 | 0.037 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, R.; Crespo, O.; Medeiros-Leal, W.; Novoa-Pabon, A.; Pinho, M. Error Distribution Model to Standardize LPUE, CPUE and Survey-Derived Catch Rates of Target and Non-Target Species. Modelling 2022, 3, 1-13. https://0-doi-org.brum.beds.ac.uk/10.3390/modelling3010001
Santos R, Crespo O, Medeiros-Leal W, Novoa-Pabon A, Pinho M. Error Distribution Model to Standardize LPUE, CPUE and Survey-Derived Catch Rates of Target and Non-Target Species. Modelling. 2022; 3(1):1-13. https://0-doi-org.brum.beds.ac.uk/10.3390/modelling3010001
Chicago/Turabian StyleSantos, Régis, Osman Crespo, Wendell Medeiros-Leal, Ana Novoa-Pabon, and Mário Pinho. 2022. "Error Distribution Model to Standardize LPUE, CPUE and Survey-Derived Catch Rates of Target and Non-Target Species" Modelling 3, no. 1: 1-13. https://0-doi-org.brum.beds.ac.uk/10.3390/modelling3010001