
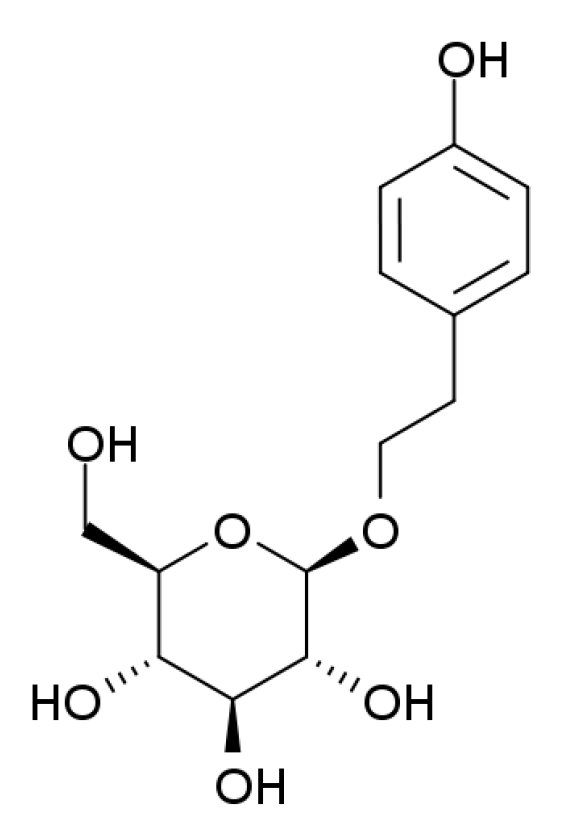
Salidroside Suppresses HUVECs Cell Injury Induced by Oxidative Stress through Activating the Nrf2 Signaling Pathway

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
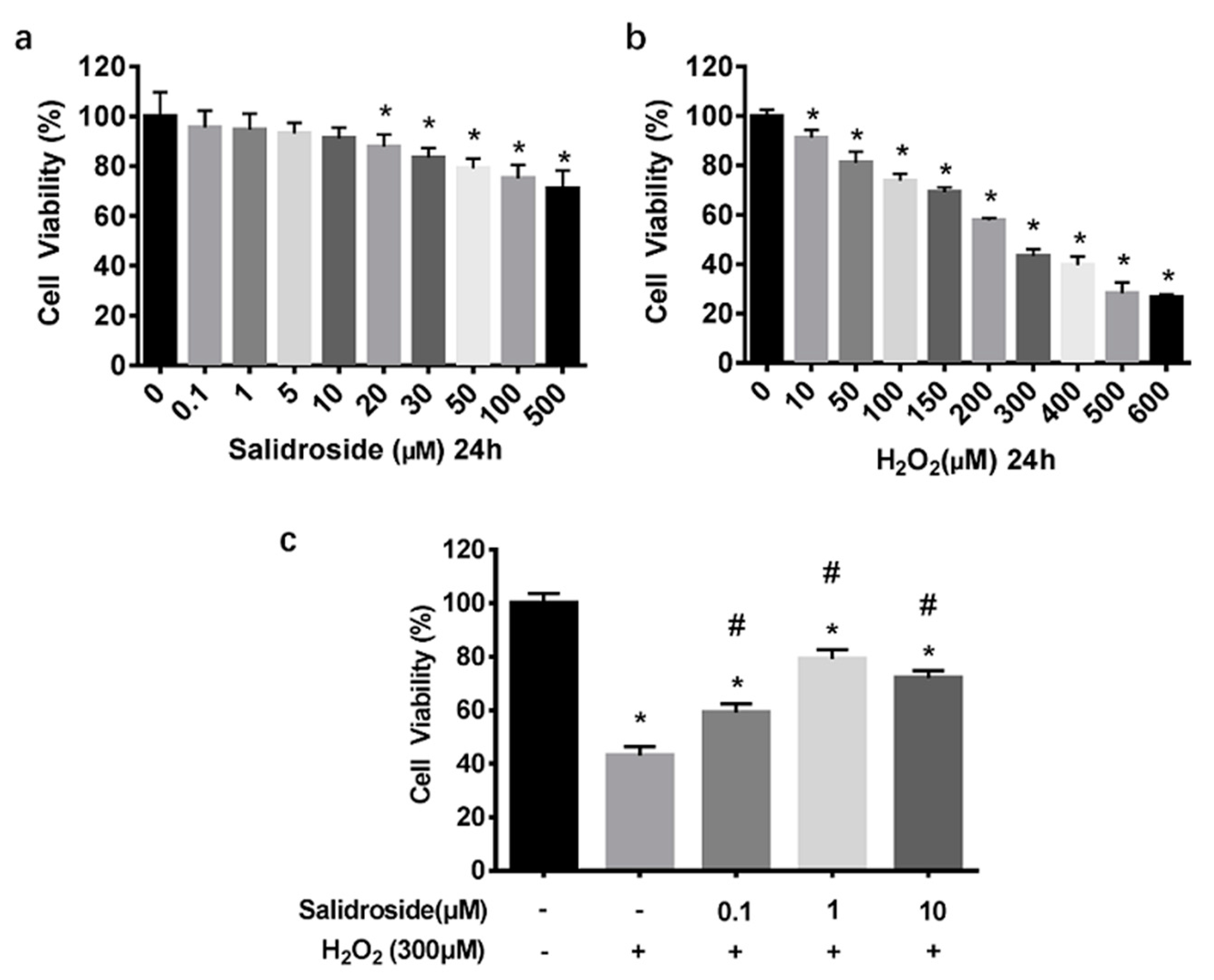
2.1. SAL Prevents H2O2-Induced Cell Injury
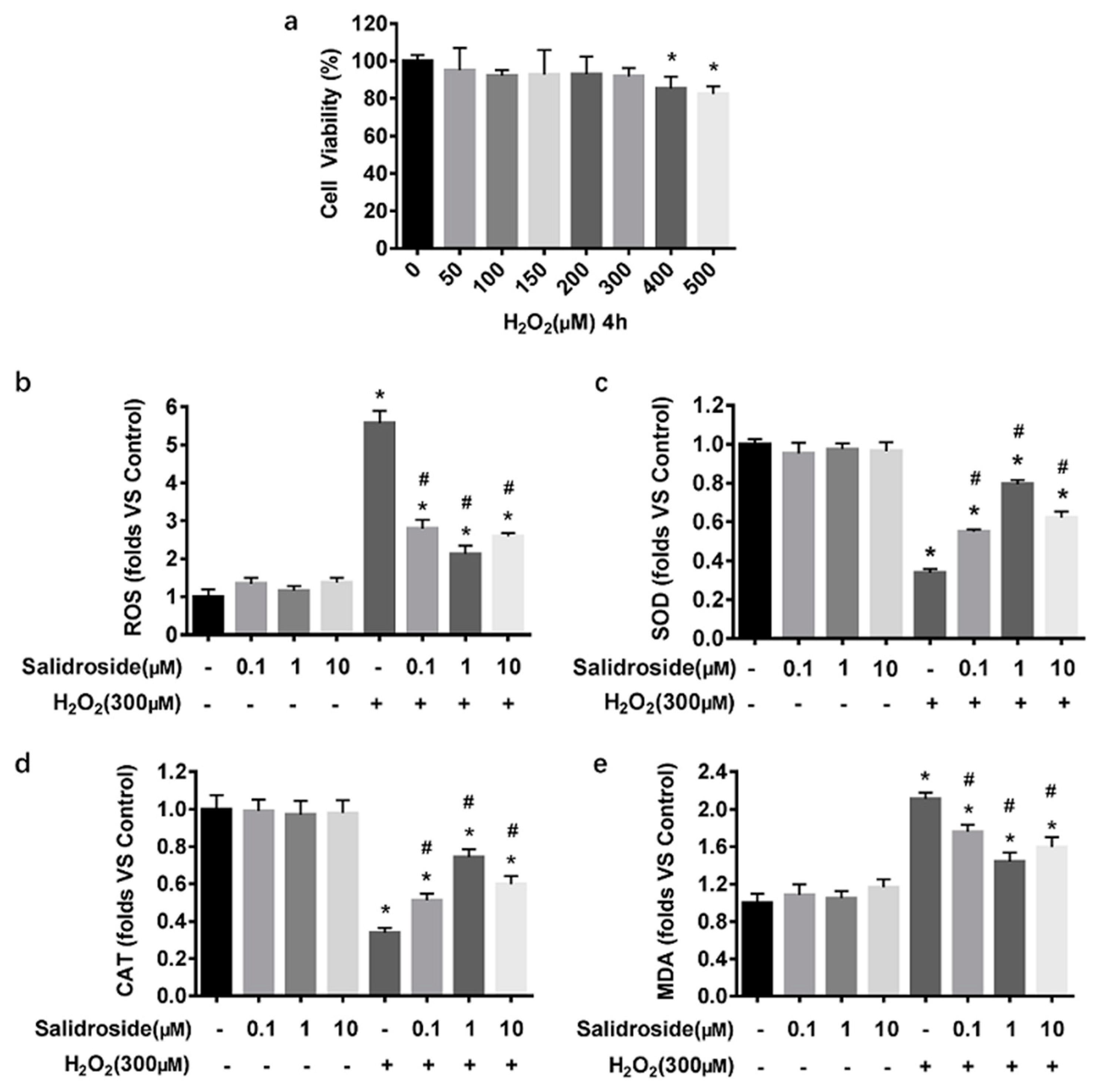
2.2. SAL Inhibits H2O2-Induced Oxidative Stress in HUVECs
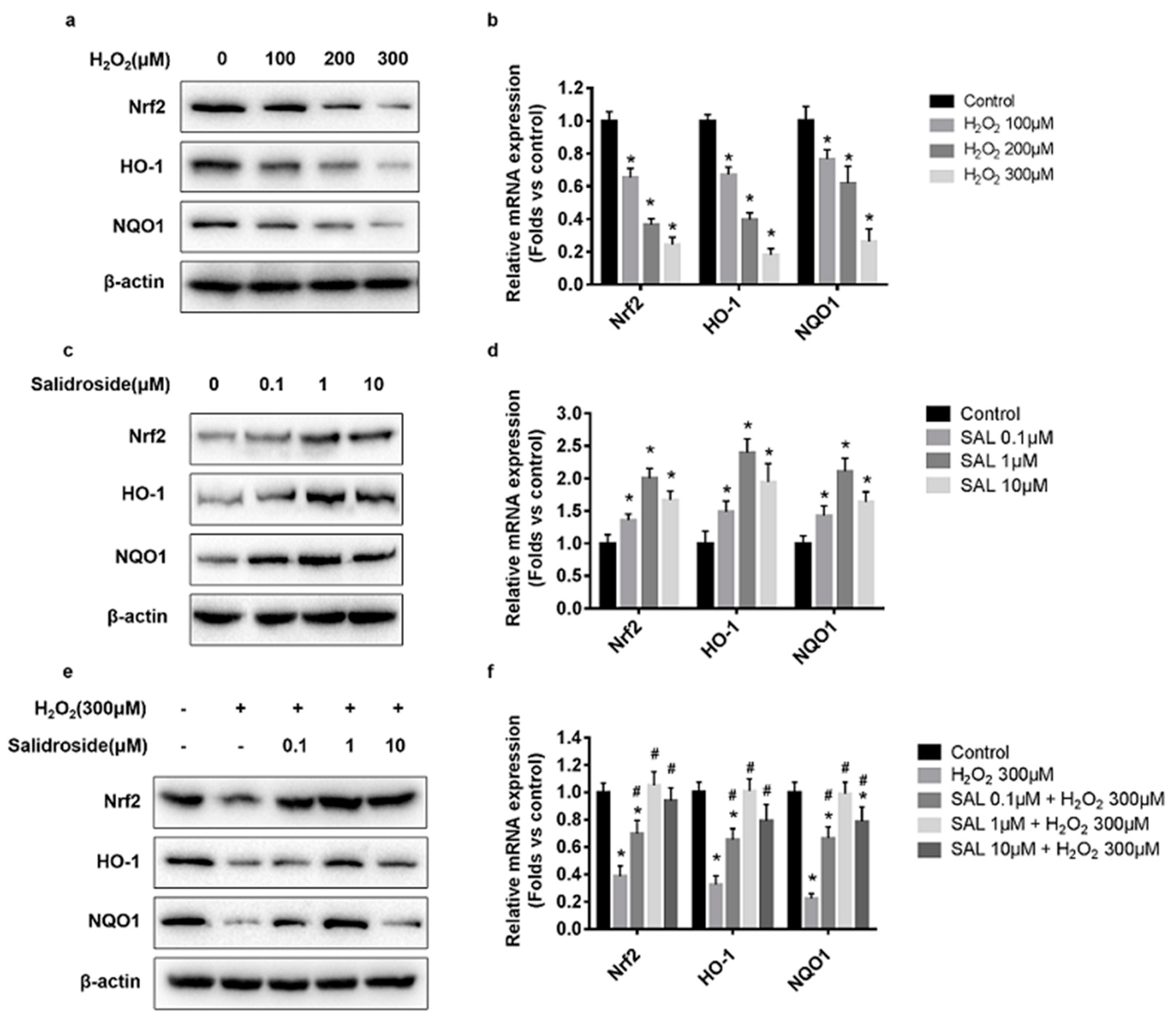
2.3. SAL Stimulated Nrf2, HO-1, and NQO1 Expression in H2O2-Treated HUVECs
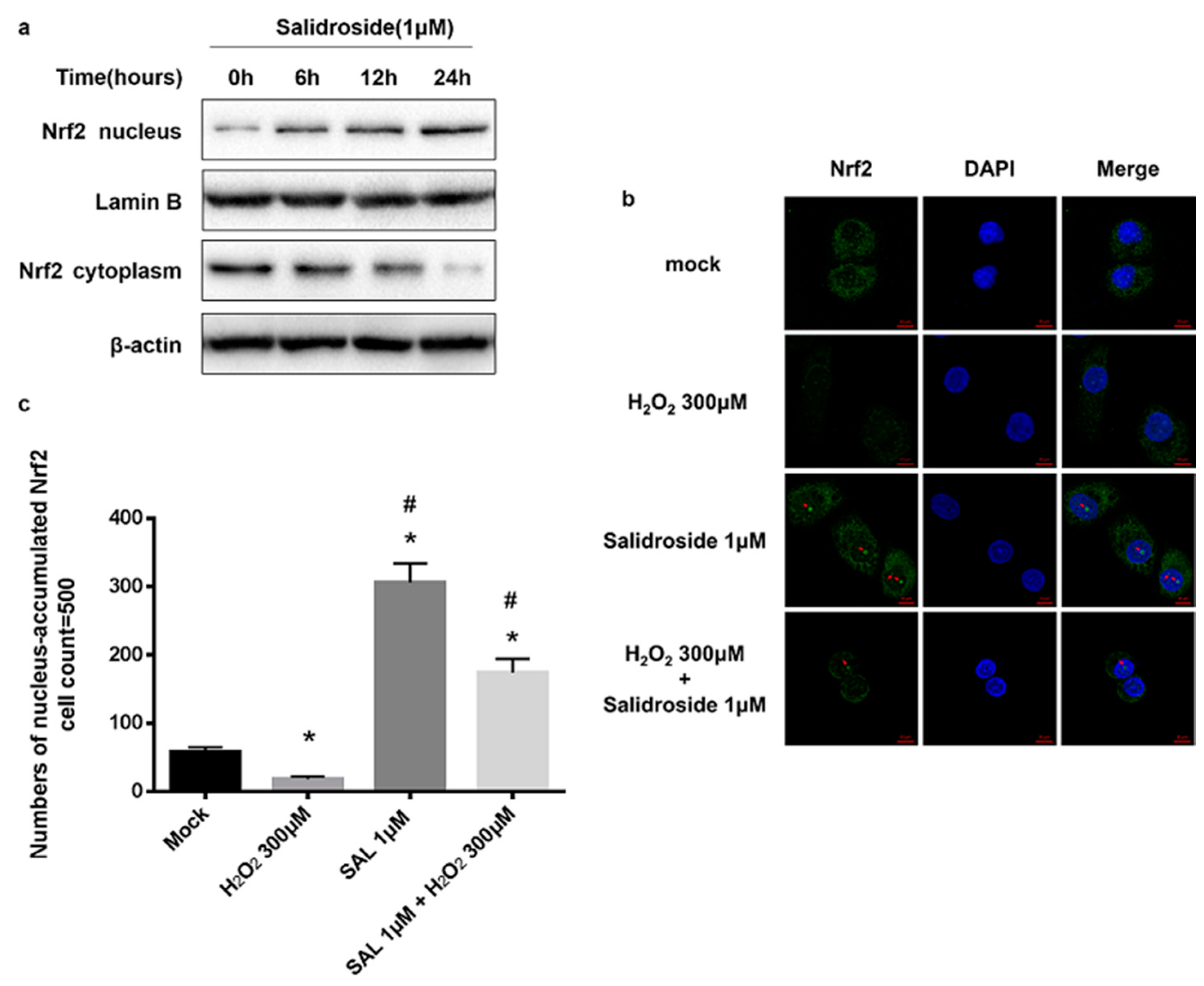
2.4. SAL Induced Nucleus Accumulation of Nrf2 in HUVECs
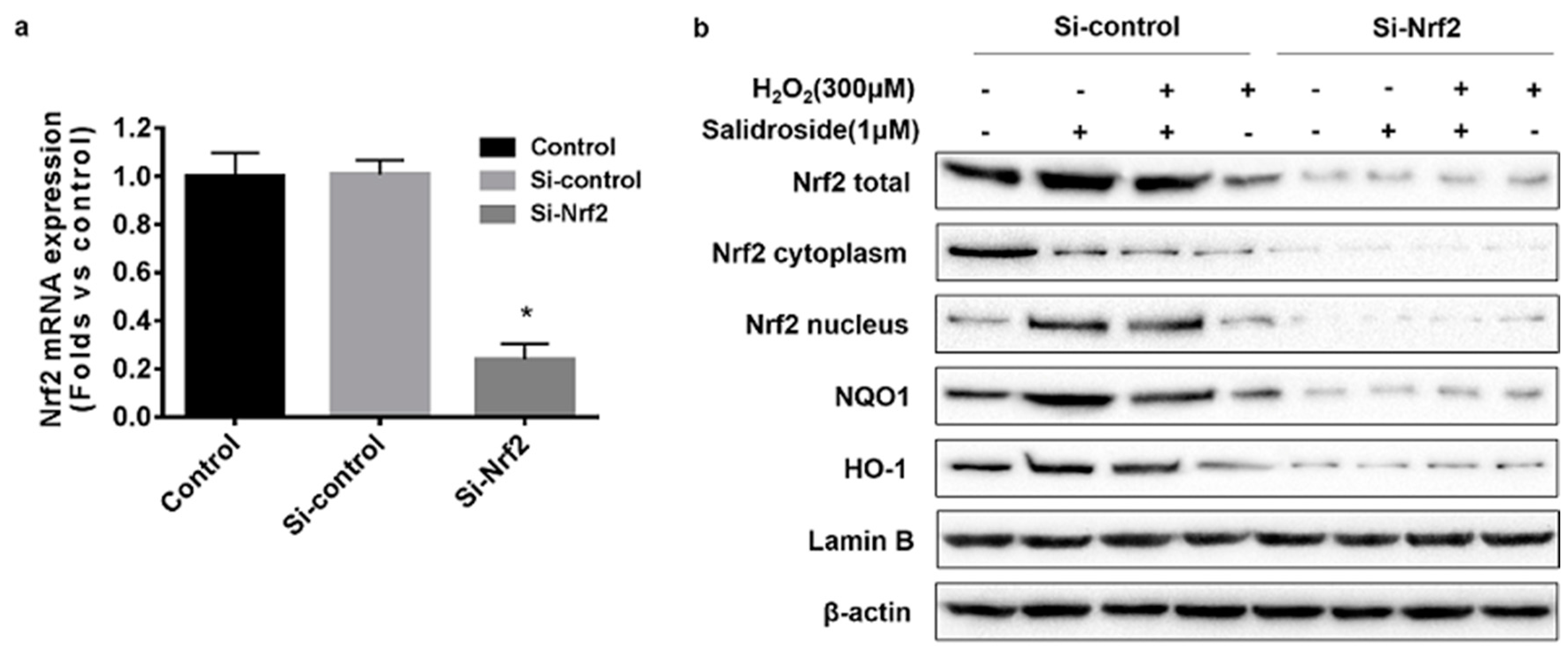
2.5. Knockdown of Nrf2 with siRNA Decreased the Expression of Nrf2, HO-1, and NQO1, and Inhibited the Nucleus Translocation of Nrf2 in HUVECs
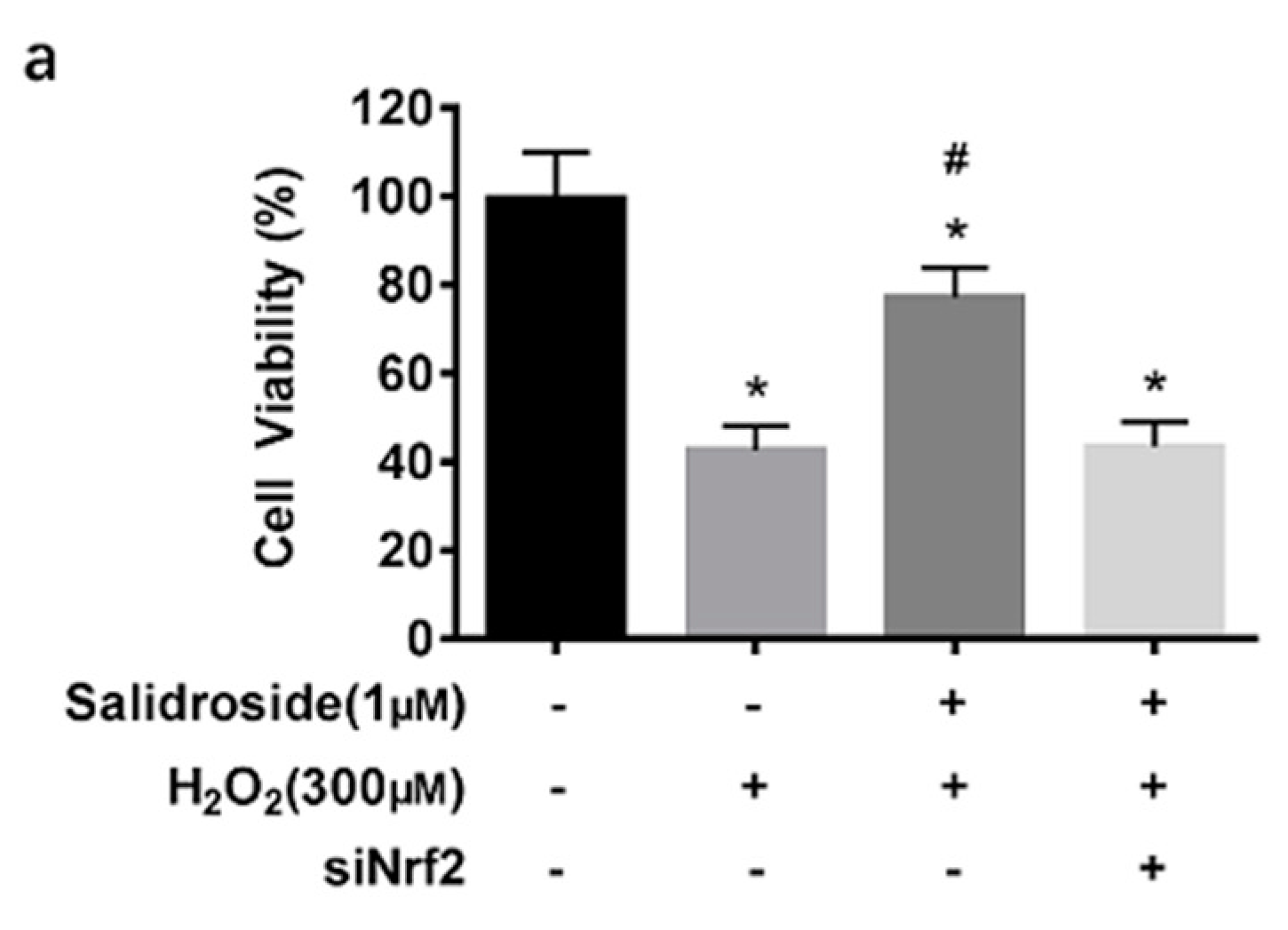
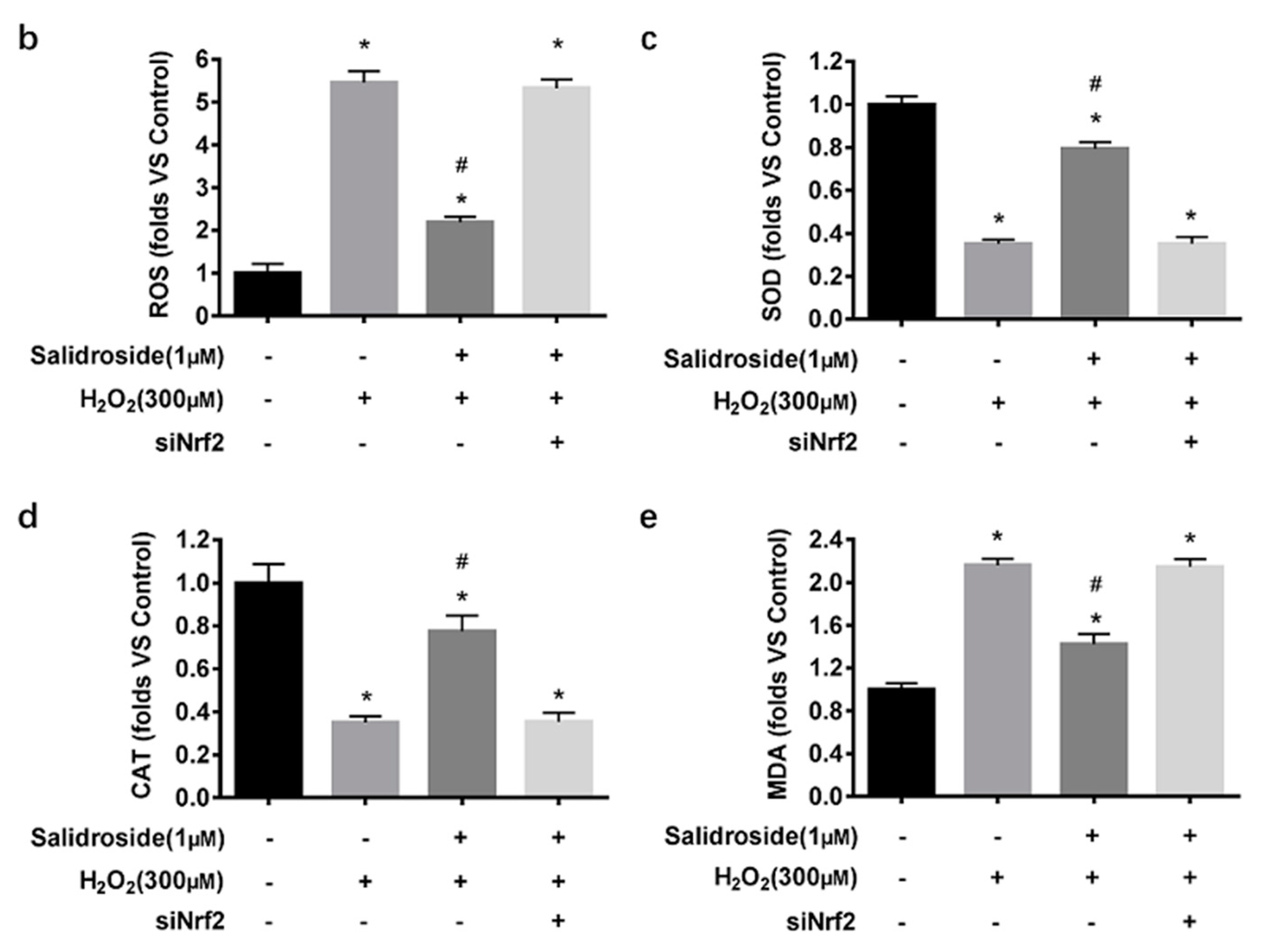
2.6. SAL-Induced Cytoprotective Effects against Oxidative Stress Induced by H2O2 in HUVECs Are Dependent on Activation of Nrf2
3. Discussion
4. Experimental Section
4.1. Reagents
4.2. Cell Culture
4.3. MTT Assay
4.4. ROS Measurement
4.5. SOD, CAT and MDA Measurements
4.6. Western Blotting
4.7. Real-Time Polymerase Chain Reaction (RT-PCR)
- Nrf2: Forward primer, 5′-CATCCAGTCAGAAACCAGTGG-3′;
- Reverse primer, 5′-GCAGTCATCAAAGTACAAAGCAT-3′;
- HO-1: Forward primer, 5′-CTTCTTCACCTTCCCCAACA-3′;
- Reverse primer, 5′-ATTGCCTGGATGTGCTTTTC-3′;
- NQO1: Forward primer, 5′- GGGATCCACGGGGACATGAATG-3′;
- Reverse primer, 5′-ATTTGAATTCGGGCGTCTGCTG-3′;
- β-actin: Forward primer, 5′-GGAAATCGTGCGTGACATTA-3′;
- Reverse primer, 5′-GGAGCAATGATCTTGATCTTC-3′;
4.8. Nrf2-siRNA Transient Transfection
4.9. Preparation of Nuclear and Cytosolic Fractions
4.10. Immunofluorescence Staining
4.11. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Ann. Rev. Plant Biol. 2004, 55, 728–749. [Google Scholar] [CrossRef] [PubMed]
- D’Autréaux, B.; Toledano, M.B. Ros as signalling molecules: Mechanisms that generate specificity in ros homeostasis. Nat. Rev. Mol. Cell Biol. 2007, 8, 813–824. [Google Scholar] [CrossRef] [PubMed]
- Chandra, K.; Salman, A.S.; Mohd, A.; Sweety, R.; Ali, K.N. Protection against fca induced oxidative stress induced DNA damage as a model of arthritis and in vitro anti-arthritic potential of costus speciosus rhizome extract. Int. J. Pharm. Phytochem. Res. 2015, 7, 383–389. [Google Scholar]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Stark, J. Oxidative stress and atherosclerosis. Orvosi Hetilap 2015, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pober, J.S.; Min, W. Endothelial cell dysfunction, injury and death. Radiother. Oncol. 2006, 111, S264–S265. [Google Scholar]
- Huet, O.; Harrois, A.; Duranteau, J. Oxidative stress and endothelial dysfunction during sepsis. Front. Biosci. 2011, 16, 1986–1995. [Google Scholar] [CrossRef]
- Heitzer, T.; Schlinzig, T.; Krohn, K.; Meinertz, T. Endothelial dysfunction, oxidative stress, and risk of cardiovascular events in patients with coronary artery disease. Circulation 2001, 104, 2673–2678. [Google Scholar] [CrossRef] [PubMed]
- Gold, R.; Kappos, L.; Arnold, D.L.; Bar-Or, A.; Giovannoni, G.; Selmaj, K.; Tornatore, C.; Sweetser, M.T.; Yang, M.; Sheikh, S.I. Placebo-controlled phase 3 study of oral bg-12 for relapsing multiple sclerosis. N. Engl. J. Med. 2012, 367, 1098–1107. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Kong, A.N. Molecular mechanisms of nrf2-mediated antioxidant response. Mol. Carcinog. 2009, 48, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Suzuki, T.; Kobayashi, A.; Wakabayashi, J.; Maher, J.; Motohashi, H.; Yamamoto, M. Physiological significance of reactive cysteine residues of Keap1 in determining nrf2 activity. Mol. Cell. Biol. 2008, 28, 2758–2770. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Chanas, S.A.; Henderson, C.J.; McMahon, M.; Sun, C.; Moffat, G.J.; Wolf, C.R.; Yamamoto, M. The nrf2 transcription factor contributes both to the basal expression of glutathione s-transferases in mouse liver and to their induction by the chemopreventive synthetic antioxidants, butylated hydroxyanisole and ethoxyquin. Biochem. Soc. Trans. 2000, 28, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Jarmi, T.; Agarwal, A. Heme oxygenase and renal disease. Curr. Hypertens. Rep. 2009, 11, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, R.; Jaiswal, A.K. Nrf1 and nrf2 positively and c-fos and fra1 negatively regulate the human antioxidant response element-mediated expression of nad(p)h:Quinone oxidoreductase1 gene. Proc. Natl. Acad. Sci. USA 1996, 93, 14960–14965. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.H.; Chen, H.W.; Liu, C.T.; Tsai, C.W.; Lii, C.K. Activation of nrf2 is required for up-regulation of the pi class of glutathione s-transferase in rat primary hepatocytes with l-methionine starvation. J. Agric. Food Chem. 2012, 60, 6537–6545. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.C.; Cui, J.Y.; Klaassen, C.D. Effect of graded nrf2 activation on phase-i and -ii drug metabolizing enzymes and transporters in mouse liver. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Morin, P., Jr.; Ni, Z.; McMullen, D.C.; Storey, K.B. Expression of nrf2 and its downstream gene targets in hibernating 13-lined ground squirrels, spermophilus tridecemlineatus. Mol. Cell Biochem. 2008, 312, 121–129. [Google Scholar] [CrossRef] [PubMed]
- McCord, J.M.; Fridovich, I. Superoxide dismutase: The first twenty years (1968–1988). Free Radical Biol. Med. 1988, 5, 363–369. [Google Scholar] [CrossRef]
- Chelikani, P.; Fita, I.; Loewen, P.C. Diversity of structures and properties among catalases. Cell. Mol. Life Sci. Cmls 2004, 61, 192–208. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Feng, L.; Jiang, W.D.; Wu, P.; Kuang, S.Y.; Jiang, J.; Tang, L.; Tang, W.N.; Zhang, Y.A.; Liu, Y.; et al. Copper-induced tight junction mrna expression changes, apoptosis and antioxidant responses via nf-kappab, tor and nrf2 signaling molecules in the gills of fish: Preventive role of arginine. Aquat. Toxicol. 2015, 158, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wang, Y.; Wan, X.; Yang, C.S.; Zhang, J. Green tea polyphenol (−)-epigallocatechin-3-gallate triggered hepatotoxicity in mice: Responses of major antioxidant enzymes and the nrf2 rescue pathway. Toxicol. Appl. Pharm. 2015, 283, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Xiu-Wen, W.U.; Peng, Y.S.; Wang, R.F. Research progress of alternative production approaches of salidroside. Chin. J. Chin. Mat. Med. 2013, 38, 3656–3660. [Google Scholar]
- Qi, Z.; Qi, S.; Ling, L.; Lv, J.; Feng, Z. Salidroside attenuates inflammatory response via suppressing jak2-stat3 pathway activation and preventing stat3 transfer into nucleus. Int. Immunopharm. 2016, 35, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Si, P.P.; Zhen, J.L.; Cai, Y.L.; Wang, W.J.; Wang, W.P. Salidroside protects against kainic acid-induced status epilepticus via suppressing oxidative stress. Neurosci. Lett. 2016, 618, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Zhang, K.; Zhou, R.; Luo, F.; Zhu, L.; Gao, J.; He, H.; Wei, T.; Yan, T.; Ma, C. Cardioprotective effects of salidroside on myocardial ischemia–reperfusion injury in coronary artery occlusion-induced rats and langendorff-perfused rat hearts. Int. J. Cardiol. 2016, 215, 532–544. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Shi, A.; Fan, Z.; Du, Y. Salidroside inhibits the growth of human breast cancer in vitro and in vivo. Oncol. Rep. 2015, 33, 2553–2560. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wang, Y.; Li, H.; Xiong, R.; Zhao, Z.; Chu, X.; Li, Q.; Sun, S.; Chen, S. Neuroprotective effects of salidroside through pi3k/akt pathway activation in alzheimer’s disease models. Drug Des. Develop. Therap. 2016, 10, 1335–1343. [Google Scholar]
- Chang, X.; Luo, F.; Jiang, W.; Zhu, L.; Gao, J.; He, H.; Wei, T.; Gong, S.; Yan, T. Protective activity of salidroside against ethanol-induced gastric ulcer via the mapk/nf-kappab pathway in vivo and in vitro. Int. Immunopharm. 2015, 28, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Liu, X.; Han, T.; Hu, D.; Wang, Y.; Yuan, Y.; Gu, J.; Bian, J.; Zhu, J.; Liu, Z.P. Salidroside protects against cadmium-induced hepatotoxicity in rats via gjic and mapk pathways. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Vater, C.; Jacobi, A.; Liebers, C.; Zou, X.; Stiehler, M. Salidroside exerts angiogenic and cytoprotective effects on human bone marrow-derived endothelial progenitor cells via akt/mtor/p70s6k and mapk signalling pathways. Br. J. Pharmacol. 2014, 171, 2440–2456. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, N.R.; Hank, N.C.; Zhang, J. Detecting presence of cardiovascular disease through mitochondria respiration as depicted through biophotonic emission. Red. Biol. 2015, 8, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Andrew, O.K.; Andreas, M.B.; Karima, A.-A.; David, D.G. Mitochondrial signaling in the vascular endothelium: Beyond reactive oxygen species. Basic Res. Cardiol. 2016, 111, 1–12. [Google Scholar] [CrossRef]
- Hamilton, C.A.; Miller, W.H.; Al-Benna, S.; Brosnan, M.J.; Drummond, R.D.; McBride, M.W.; Dominiczak, A.F. Strategies to reduce oxidative stress in cardiovascular disease. Clin. Sci. 2004, 106, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Lu, Y.; Chen, Y.; Cheng, J. The role of nrf2 in oxidative stress-induced endothelial injuries. J. Endocrinol. 2015, 225, R83–R99. [Google Scholar] [CrossRef] [PubMed]
- Ishikado, A.; Sono, Y.; Matsumoto, M.; Robida-Stubbs, S.; Okuno, A.; Goto, M.; King, G.L.; Blackwell, T.K.; Makino, T. Willow bark extract increases antioxidant enzymes and reduces oxidative stress through activation of nrf2 in vascular endothelial cells and caenorhabditis elegans. Free Radic. Biol. Med. 2013, 65, 1506–1515. [Google Scholar] [CrossRef] [PubMed]
- Yama, K.; Sato, K.; Abe, N.; Murao, Y.; Tatsunami, R.; Tampo, Y. Epalrestat increases glutathione, thioredoxin, and heme oxygenase-1 by stimulating nrf2 pathway in endothelial cells. Redox Biol. 2015, 4, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Kang, S.M.; Lee, W.T.; Park, K.A.; Lee, K.M.; Lee, J.E. Glutathione protects brain endothelial cells from hydrogen peroxide-induced oxidative stress by increasing nrf2 expression. Exp. Neurobiol. 2014, 23, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.Z.; Wang, Z.J.; Bai, F.; Qin, X.J.; Cao, J.; Lv, J.Y.; Zhang, M.S. Epigallocatechin-3-gallate protects huvecs from pm2.5-induced oxidative stress injury by activating critical antioxidant pathways. Molecules 2015, 20, 6626–6639. [Google Scholar] [CrossRef] [PubMed]
- Shi, K.; Wang, X.; Zhu, J.; Cao, G.; Zhang, K.; Su, Z. Salidroside protects retinal endothelial cells against hydrogen peroxide-induced injury via modulating oxidative status and apoptosis. Biosci. Biotechnol. Biochem. 2015, 79, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.C.; Shi, H.M.; Wang, H.; Gao, X.F. Salidroside protects against hydrogen peroxide-induced injury in huvecs via the regulation of redd1 and mtor activation. Mol. Med. Rep. 2013, 8, 147–153. [Google Scholar] [PubMed]
- Xing, S.; Yang, X.; Li, W.; Fang, B.; Dan, W.; Chi, J.; Gao, X.; Zhang, Y.; Si, J. Salidroside stimulates mitochondrial biogenesis and protects against H2O2-induced endothelial dysfunction. Oxidative Med. Cell. Longevity 2014, 2014, 904834. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Gao, L.; Mao, J.; He, H.; Jia, L.; Xin, C.; Lin, H.; Wu, T. Salidroside protects against bleomycin-induced pulmonary fibrosis: Activation of nrf2-antioxidant signaling, and inhibition of nf-κb and tgf-β1/smad-2/-3 pathways. Cell Stress Chaperones 2015, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Sheng, Z.; Li, Y.; Lu, H. Salidroside inhibits oxygen glucose deprivation (ogd)/re-oxygenation-induced h9c2 cell necrosis through activating of akt–nrf2 signaling. Biochem. Biophys. Res. Commun. 2014, 451, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Ping, Z.; Zhang, L.F.; Cui, Y.J.; Chang, Y.M.; Jiang, C.W.; Meng, Z.Z.; Xu, P.; Liu, H.Y.; Wang, D.Y.; Cao, X.B. The protective effects of salidroside from exhaustive exercise-induced heart injury by enhancing the pgc-1α–nrf1/nrf2 pathway and mitochondrial respiratory function in rats. Oxidative Med. Cell. Longevity 2014, 2015. [Google Scholar] [CrossRef]
- Wang, S.; He, H.; Li, X.; Wang, L.; He, G.; Chen, J. Salidroside attenuated mpp~+-induced apoptosis in pc12 cells by the activation of nrf2. Chin. J. Neuroanat. 2013, 29, 651–655. [Google Scholar]
- Han, J.; Xiao, Q.; Lin, Y.H.; Zheng, Z.Z.; He, Z.D.; Hu, J.; Chen, L.D. Neuroprotective effects of salidroside on focal cerebral ischemia/reperfusion injury involve the nuclear erythroid 2-related factor 2 pathway. Neur. Regenerat. Res. 2015, 10, 1989–1996. [Google Scholar]
- Cai, H.; Harrison, D.G. Endothelial dysfunction in cardiovascular diseases: The role of oxidant stress. Circ. Res. 2000, 87, 840–844. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhao, H.; Li, H.; Kalyanaraman, B.; Nicolosi, A.C.; Gutterman, D.D. Mitochondrial sources of H2O2 generation play a key role in flow-mediated dilation in human coronary resistance arteries. Circ. Res. 2003, 93, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Cai, H. Hydrogen peroxide regulation of endothelial function: Origins, mechanisms, and consequences. Cardiovasc. Res. 2005, 68, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Covas, G.; Marinho, H.S.; Cyrne, L.; Antunes, F. Activation of nrf2 by H2O2: De novo synthesis versus nuclear translocation. Meth. Enzymol. 2013, 528, 157–171. [Google Scholar] [PubMed]
- Fourquet, S.; Guerois, R.; Biard, D.; Toledano, M.B. Activation of nrf2 by nitrosative agents and H2O2 involves Keap1 disulfide formation. J. Biol. Chem. 2010, 285, 8463–8471. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of the salidroside are available from the authors.








© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.; Zhang, Y.-J.; Liu, W.-W.; Shi, A.-W.; Gu, N. Salidroside Suppresses HUVECs Cell Injury Induced by Oxidative Stress through Activating the Nrf2 Signaling Pathway. Molecules 2016, 21, 1033. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21081033
Zhu Y, Zhang Y-J, Liu W-W, Shi A-W, Gu N. Salidroside Suppresses HUVECs Cell Injury Induced by Oxidative Stress through Activating the Nrf2 Signaling Pathway. Molecules. 2016; 21(8):1033. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21081033
Chicago/Turabian StyleZhu, Yao, Ya-Jie Zhang, Wei-Wei Liu, Ai-Wu Shi, and Ning Gu. 2016. "Salidroside Suppresses HUVECs Cell Injury Induced by Oxidative Stress through Activating the Nrf2 Signaling Pathway" Molecules 21, no. 8: 1033. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21081033