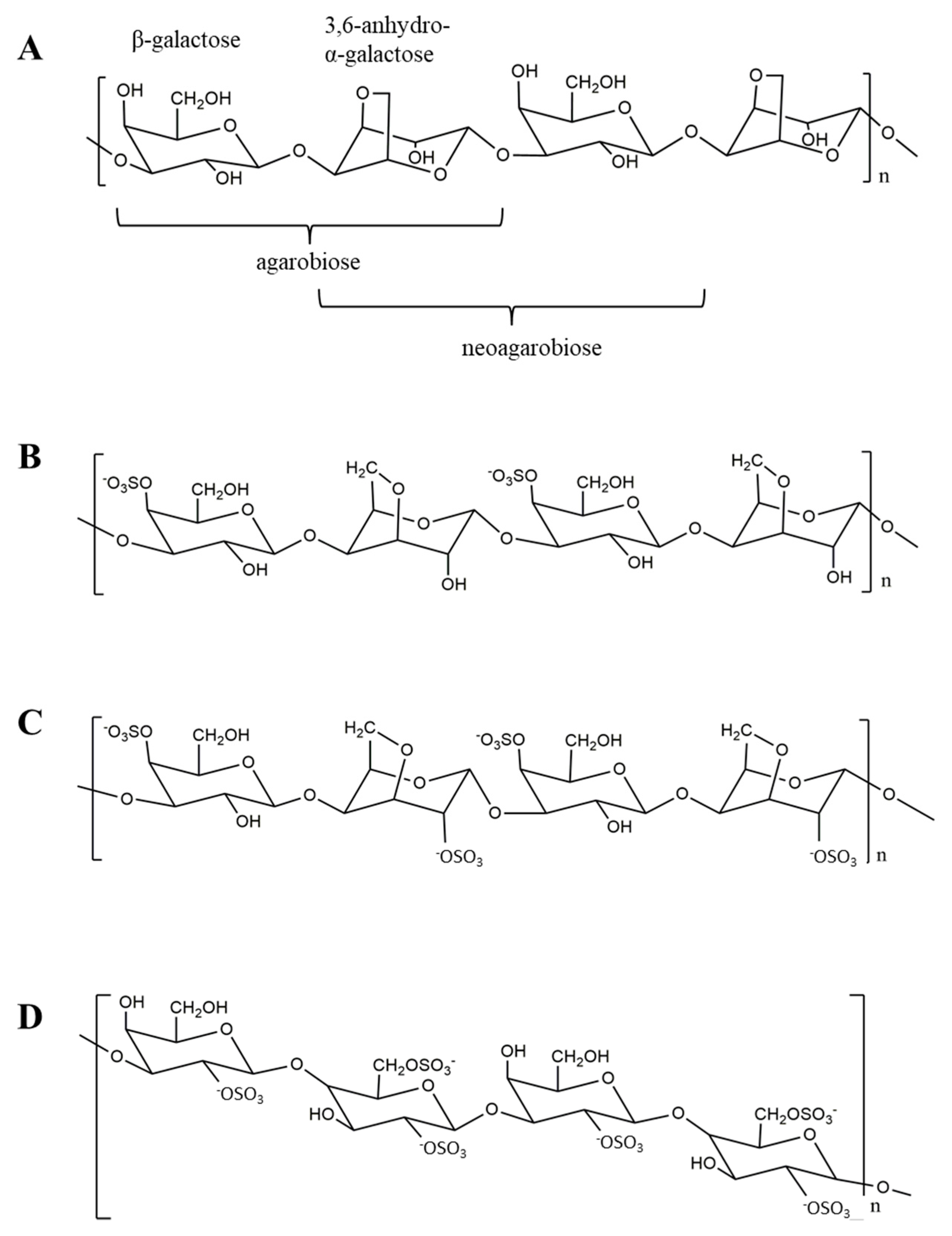
Oligosaccharides Derived from Red Seaweed: Production, Properties, and Potential Health and Cosmetic Applications


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
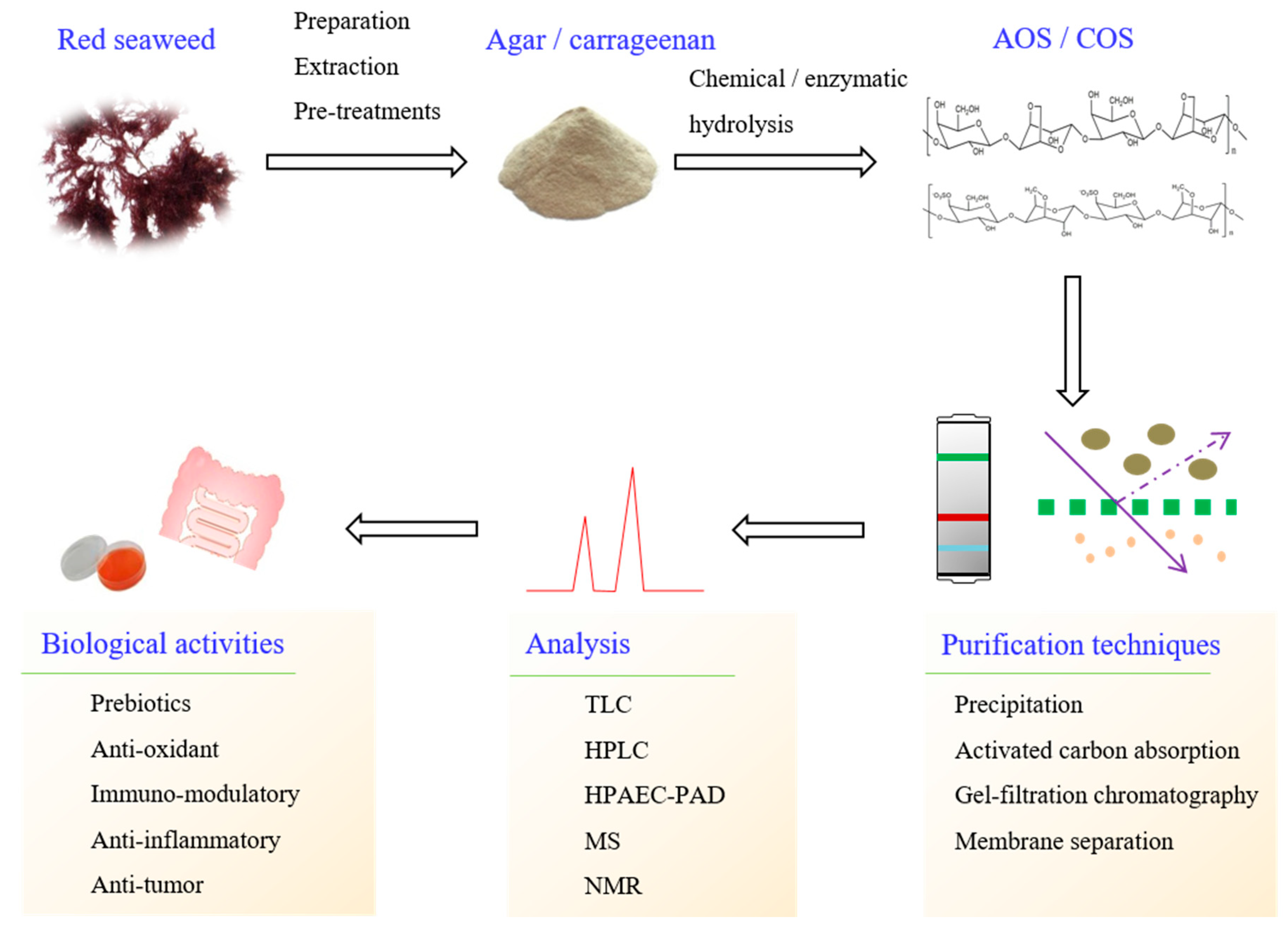
2. Production of Oligosaccharide from Red Seaweed
2.1. Pretreatment of Red Seaweed Polysaccharides
2.2. Chemical and Enzymatic Hydrolyses for Oligosaccharides Production
2.2.1. Oligosaccharides Production by Chemical Hydrolysis and De-Polymerization
2.2.2. Oligosaccharides Production by Enzymatic Hydrolysis
3. Purification of Oligosaccharides
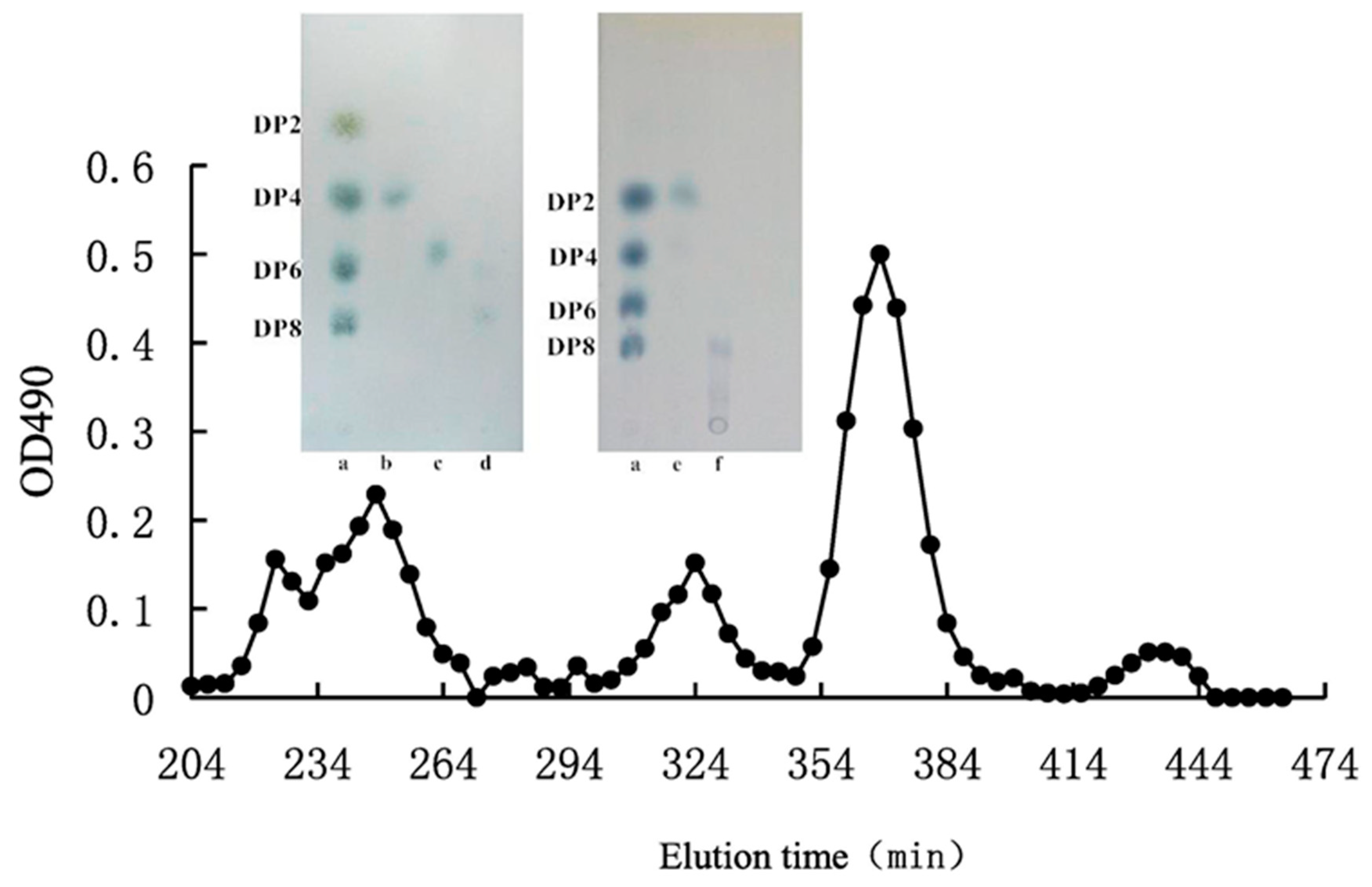
4. Chromatographic Separation of Oligosaccharides
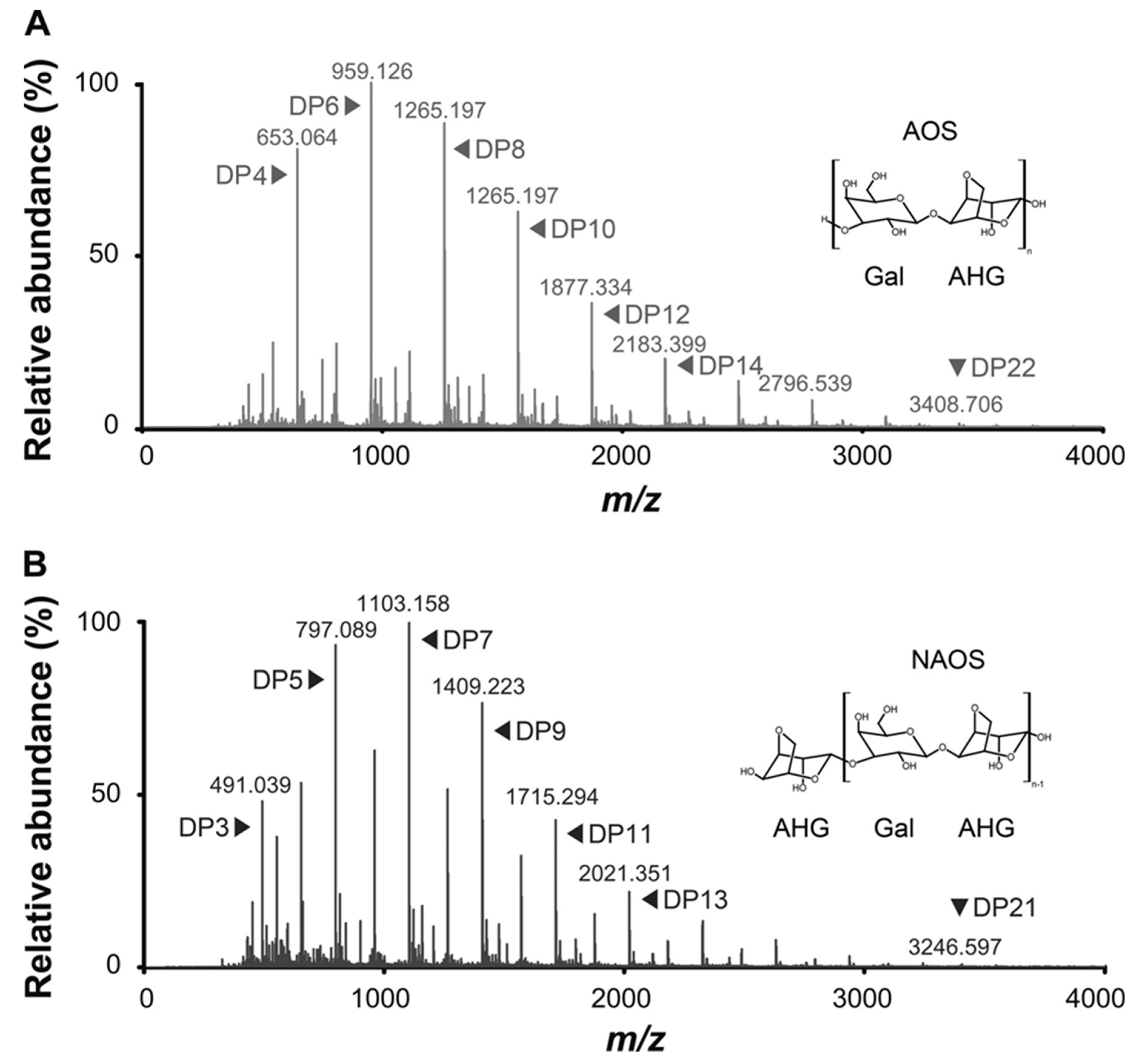
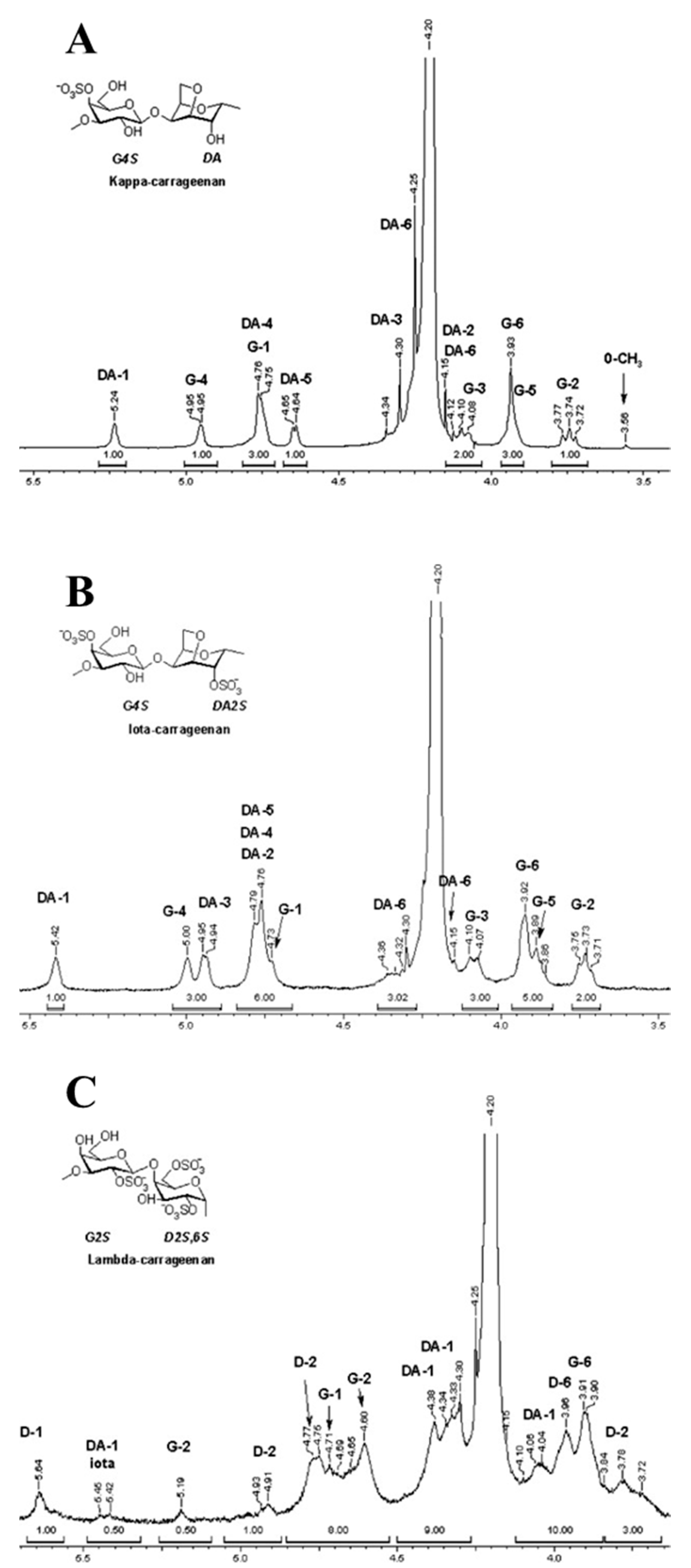
5. Physicochemical Characteristics
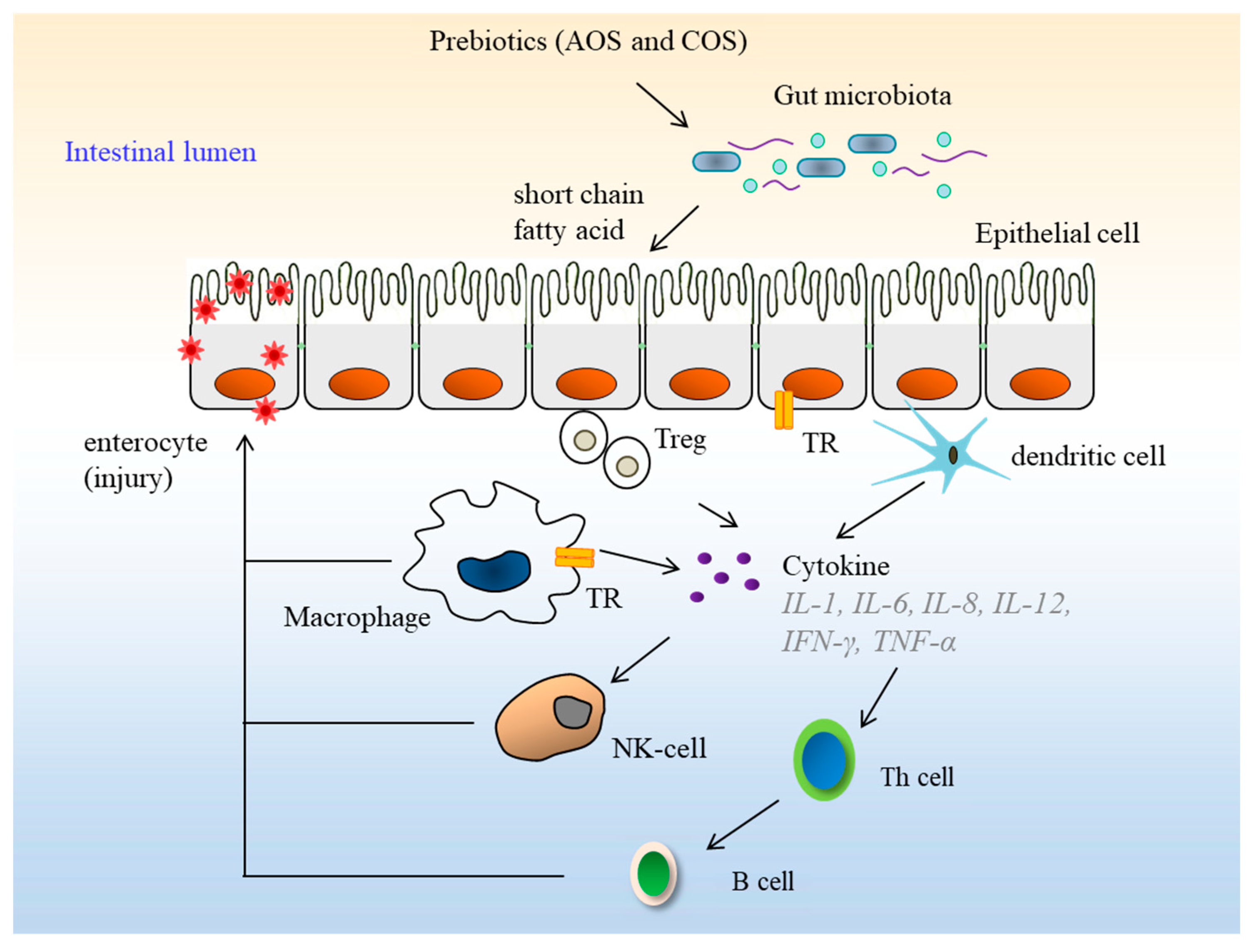
6. Biological Activities of Oligosaccharides
7. Potential Use of Oligosaccharides in Cosmetics
8. Future Perspective
Supplementary Materials
Supplementary File 1Author Contributions
Funding
Conflicts of Interest
References
- Xu, S.Y.; Huang, X.; Cheong, K.L. Recent advances in marine algae polysaccharides: Isolation, structure, and activities. Mar. Drugs 2017, 15, 388. [Google Scholar] [CrossRef] [PubMed]
- McKim, J.M. Food additive carrageenan: Part I: A critical review of carrageenan in vitro studies, potential pitfalls, and implications for human health and safety. Crit. Rev. Toxicol. 2014, 44, 211–243. [Google Scholar] [CrossRef] [PubMed]
- Pomin, V.H. Structural and functional insights into sulfated galactans: A systematic review. Glycoconj. J. 2010, 27, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Usov, A.I. Structural analysis of red seaweed galactans of agar and carrageenan groups. Food Hydrocolloids 1998, 12, 301–308. [Google Scholar] [CrossRef]
- Liu, J.; Zhan, X.; Wan, J.; Wang, Y.; Wang, C. Review for carrageenan-based pharmaceutical biomaterials: Favourable physical features versus adverse biological effects. Carbohydr. Polym. 2015, 121, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yang, B.; Wu, Y.; Liu, Y.; Gu, X.; Zhang, H.; Wang, C.; Cao, H.; Huang, L.; Wang, Z. Structural characterization and antioxidant activities of κ-carrageenan oligosaccharides degraded by different methods. Food Chem. 2015, 178, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.T.; Kim, S.M. Agarase: Review of major sources, categories, purification method, enzyme characteristics and applications. Mar. Drugs 2010, 8, 200–218. [Google Scholar] [CrossRef] [PubMed]
- Chi, W.J.; Chang, Y.K.; Hong, S.K. Agar degradation by microorganisms and agar-degrading enzymes. Appl. Microbiol. Biotechnol. 2012, 94, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Lang, Y.; Zhao, X.; Liu, L.; Yu, G. Applications of mass spectrometry to structural analysis of marine oligosaccharides. Mar. Drugs 2014, 12, 4005–4030. [Google Scholar] [CrossRef] [PubMed]
- Bhattarai, Y.; Kashyap, P.C. Agaro-oligosaccharides: A new frontier in the fight against colon cancer? Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G335. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, G.; Zhu, L.; Yin, Y.; Zhao, X.; Xiang, C.; Yu, G.; Wang, X. Isolation and characterization of an agaro-oligosaccharide (AO)-hydrolyzing bacterium from the gut microflora of Chinese individuals. PLoS ONE 2014, 9, e91106. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Xing, R.; Liu, S.; Li, P. Advances in preparation, analysis and biological activities of single chitooligosaccharides. Carbohydr. Polym. 2016, 139, 178–190. [Google Scholar] [CrossRef] [PubMed]
- McCranie, E.K.; Bachmann, B.O. Bioactive oligosaccharide natural products. Nat. Prod. Rep. 2014, 31, 1026–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nobre, C.; Teixeira, J.A.; Rodrigues, L.R. New trends and technological challenges in the industrial production and purification of fructo-oligosaccharides. Crit. Rev. Food Sci. Nutr. 2015, 55, 1444–1455. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.F.A.; de Oliva Neto, P.; da Silva, D.F.; Pastore, G.M. Xylo-oligosaccharides from lignocellulosic materials: Chemical structure, health benefits and production by chemical and enzymatic hydrolysis. Food Res. Int. 2013, 51, 75–85. [Google Scholar] [CrossRef]
- Bruno-Barcena, J.M.; Azcarate-Peril, M.A. Galacto-oligosaccharides and colorectal cancer: Feeding our intestinal probiome. J. Funct. Foods 2015, 12, 92–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamabhai, M.; Sak-Ubol, S.; Srila, W.; Haltrich, D. Mannan biotechnology: From biofuels to health. Crit. Rev. Biotechnol. 2016, 36, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Yun, E.J.; Yu, S.; Kim, K.H. Current knowledge on agarolytic enzymes and the industrial potential of agar-derived sugars. Appl. Microbiol. Biotechnol. 2017, 101, 5581–5589. [Google Scholar] [CrossRef] [PubMed]
- Fleita, D.; El-Sayed, M.; Rifaat, D. Evaluation of the antioxidant activity of enzymatically-hydrolyzed sulfated polysaccharides extracted from red algae; Pterocladia capillacea. LWT–Food Sci. Technol. 2015, 63, 1236–1244. [Google Scholar] [CrossRef]
- Boulho, R.; Marty, C.; Freile-Pelegrín, Y.; Robledo, D.; Bourgougnon, N.; Bedoux, G. Antiherpetic (HSV-1) activity of carrageenans from the red seaweed Solieria chordalis (Rhodophyta, Gigartinales) extracted by microwave-assisted extraction (MAE). J. Appl. Phycol. 2017, 29, 2219–2228. [Google Scholar] [CrossRef]
- Fidelis, G.; Camara, R.; Queiroz, M.; Costa, M.; Santos, P.; Rocha, H.; Costa, L. Proteolysis, NaOH and ultrasound-enhanced extraction of anticoagulant and antioxidant sulfated polysaccharides from the edible seaweed, Gracilaria birdiae. Molecules 2014, 19, 18511–18562. [Google Scholar] [CrossRef] [PubMed]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Application of novel extraction technologies for bioactives from marine algae. J. Agric. Food Chem. 2013, 61, 4667–4675. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Q.; Su, B.M.; Xie, J.S.; Li, R.K.; Yang, J.; Lin, J.; Ye, X.Y. Preparation of bioactive neoagaroligosaccharides through hydrolysis of Gracilaria lemaneiformis agar: A comparative study. Food Chem. 2018, 240, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Kazłowski, B.; Pan, C.L.; Ko, Y.T. Monitoring and preparation of neoagaro- and agaro-oligosaccharide products by high performance anion exchange chromatography systems. Carbohydr. Polym. 2015, 122, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Kamińska-Dwórznicka, A.; Matusiak, M.; Samborska, K.; Witrowa-Rajchert, D.; Gondek, E.; Jakubczyk, E.; Antczak, A. The influence of kappa carrageenan and its hydrolysates on the recrystallization process in sorbet. J. Food Eng. 2015, 167, 162–165. [Google Scholar] [CrossRef]
- Ducatti, D.R.; Colodi, F.G.; Goncalves, A.G.; Duarte, M.E.R.; Noseda, M.D. Production of agaro- and carra-oligosaccharides by partial acid hydrolysis of galactans. Rev. Bras. Farm. 2011, 21, 296–304. [Google Scholar] [CrossRef] [Green Version]
- Kalitnik, A.A.; Marcov, P.A.; Anastyuk, S.D.; Barabanova, A.O.B.; Glazunov, V.P.; Popov, S.V.; Ovodov, Y.S.; Yermak, I.M. Gelling polysaccharide from Chondrus armatus and its oligosaccharides: The structural peculiarities and anti-inflammatory activity. Carbohydr. Polym. 2015, 115, 768–775. [Google Scholar] [CrossRef] [PubMed]
- Kalitnik, A.A.; Byankina Barabanova, A.O.; Nagorskaya, V.P.; Reunov, A.V.; Glazunov, V.P.; Solov’eva, T.F.; Yermak, I.M. Low molecular weight derivatives of different carrageenan types and their antiviral activity. J. Appl. Phycol. 2013, 25, 65–72. [Google Scholar] [CrossRef]
- Kim, S.W.; Hong, C.H.; Jeon, S.W.; Shin, H.J. High-yield production of biosugars from Gracilaria verrucosa by acid and enzymatic hydrolysis processes. Bioresour. Technol. 2015, 196, 634–641. [Google Scholar] [CrossRef] [PubMed]
- Bian, J.; Peng, P.; Peng, F.; Xiao, X.; Xu, F.; Sun, R.C. Microwave-assisted acid hydrolysis to produce xylooligosaccharides from sugarcane bagasse hemicelluloses. Food Chem. 2014, 156, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Achour, O.; Bridiau, N.; Godhbani, A.; Le Joubioux, F.; Bordenave Juchereau, S.; Sannier, F.; Piot, J.M.; Fruitier Arnaudin, I.; Maugard, T. Ultrasonic-assisted preparation of a low molecular weight heparin (LMWH) with anticoagulant activity. Carbohydr. Polym. 2013, 97, 684–689. [Google Scholar] [CrossRef] [PubMed]
- Guangsheng, L.I.; Xia, Z.; Youjing, L.V.; Miaomiao, L.I.; Guangli, Y.U. Preparation of κ-carra-oligosaccharides with microwave assisted acid hydrolysis method. J. Ocean Univ. China 2015, 14, 345–349. [Google Scholar]
- Osamu, A.; Naoki, O.; Naomi, H.; Kiyohiko, N. Purification and characterization of α-neoagarooligosaccharide hydrolase from Cellvibrio sp. OA-2007. J. Microbiol. Biotechnol. 2014, 24, 48–51. [Google Scholar]
- Ropartz, D.; Giuliani, A.; Hervé, C.; Geairon, A.; Jam, M.; Czjzek, M.; Rogniaux, H. High-energy photon activation tandem mass spectrometry provides unprecedented insights into the structure of highly sulfated oligosaccharides extracted from macroalgal cell walls. Anal. Chem. 2015, 87, 1042–1049. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Yun, E.J.; Kim, H.T.; Choi, I.-G.; Kim, K.H. Saccharification of agar using hydrothermal pretreatment and enzymes supplemented with agarolytic β-galactosidase. Process Biochem. 2015, 50, 1629–1633. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, Y.; Jiang, K.; Wang, C.; Wang, Z.; Huang, L. Electrospray ionization mass spectrometric analysis of κ-crrageenan oligosaccharides obtained by degradation with κ-carrageenase from Pedobacter hainanensis. J. Agric. Food Chem. 2014, 62, 2398–2405. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Chang, Y.; Dong, S.; Chen, F. Cloning, expression and characterization of a ι-carrageenase from marine bacteria Wenyingzhuangia fucanilytica: A biocatalyst for producing ι-carrageenan oligosaccharides. J. Biotechnol. 2017, 259, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hu, Q.; Seswita-Zilda, D. Purification and characterization of a thermostable λ-carrageenase from a hot spring bacterium, Bacillus sp. Biotechnol. Lett. 2014, 36, 1669–1674. [Google Scholar] [CrossRef] [PubMed]
- Duan, F.; Yu, Y.; Liu, Z.; Tian, L.; Mou, H. An effective method for the preparation of carrageenan oligosaccharides directly from Eucheuma cottonii using cellulase and recombinant κ-carrageenase. Algal Res. 2016, 15, 93–99. [Google Scholar] [CrossRef]
- Kwon, O.-M.; Kim, S.K.; Jeong, G.T. Potential of phosphoric acid-catalyzed pretreatment and subsequent enzymatic hydrolysis for biosugar production from Gracilaria verrucosa. Bioproc. Biosyst. Eng. 2016, 39, 1173–1180. [Google Scholar] [CrossRef] [PubMed]
- Hehemann, J.-H.; Correc, G.; Barbeyron, T.; Helbert, W.; Czjzek, M.; Michel, G. Transfer of carbohydrate-active enzymes from marine bacteria to Japanese gut microbiota. Nature 2010, 464, 908–912. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, P.S.; Saxena, A. Bacterial carrageenases: An overview of production and biotechnological applications. 3 Biotech 2016, 6, 146. [Google Scholar] [CrossRef] [PubMed]
- Vanegas, C.H.; Hernon, A.; Bartlett, J. Enzymatic and organic acid pretreatment of seaweed: Effect on reducing sugars production and on biogas inhibition. Int. J. Ambient Energy 2015, 36, 2–7. [Google Scholar] [CrossRef]
- Kim, S.W.; Kim, Y.-W.; Hong, C.-H.; Lyo, I.-W.; Lim, H.-D.; Kim, G.-J.; Shin, H.-J. Recombinant agarase increases the production of reducing sugars from HCl-treated Gracilaria verrucosa, a red algae. Algal Res. 2017, 31, 517–524. [Google Scholar] [CrossRef]
- Cui, F.; Dong, S.; Shi, X.; Zhao, X.; Zhang, X.H. Overexpression and characterization of a novel thermostable β-agarase YM01-3, from marine bacterium Catenovulum agarivorans YM01T. Mar. Drugs 2014, 12, 2731–2747. [Google Scholar] [CrossRef] [PubMed]
- Kang, O.L.; Ghani, M.; Hassan, O.; Rahmati, S.; Ramli, N. Novel agaro-oligosaccharide production through enzymatic hydrolysis: Physicochemical properties and antioxidant activities. Food Hydrocolloidr. 2014, 42, 304–308. [Google Scholar] [CrossRef]
- Xu, S.Y.; Kan, J.; Hu, Z.; Liu, Y.; Du, H.; Pang, G.C.; Cheong, K.L. Quantification of neoagaro-oligosaccharide production through enzymatic hydrolysis and its anti-oxidant activities. Molecules 2018, 23, 1354. [Google Scholar] [CrossRef] [PubMed]
- Liese, A.; Hilterhaus, L. Evaluation of immobilized enzymes for industrial applications. Chem. Soc. Rev. 2013, 42, 6236–6249. [Google Scholar] [CrossRef] [PubMed]
- Koti, B.A.; Shinde, M.; Lalitha, J. Repeated batch production of agar-oligosaccharides from agarose by an amberlite IRA-900 immobilized agarase system. Biotechnol. Bioproc. Eng. 2013, 18, 333–341. [Google Scholar] [CrossRef]
- Xiao, A.; Xu, C.; Lin, Y.; Ni, H.; Zhu, Y.; Cai, H. Preparation and characterization of κ-carrageenase immobilized onto magnetic iron oxide nanoparticles. Electron. J. Biotechnol. 2016, 19, 1–7. [Google Scholar] [CrossRef]
- Moreno, F.J.; Corzo, N.; Montilla, A.; Villamiel, M.; Olano, A. Current state and latest advances in the concept, production and functionality of prebiotic oligosaccharides. Curr. Opin. Food Sci. 2017, 13, 50–55. [Google Scholar] [CrossRef] [Green Version]
- Ghanbarzadeh, M.; Golmoradizadeh, A.; Homaei, A. Carrageenans and carrageenases: Versatile polysaccharides and promising marine enzymes. Phytochem. Rev. 2018, 17, 535–571. [Google Scholar] [CrossRef]
- Xiao, A.; Xiao, Q.; Lin, Y.; Ni, H.; Zhu, Y.; Cai, H. Efficient immobilization of agarase using carboxyl-functionalized magnetic nanoparticles as support. Electron. J. Biotechnol. 2017, 25, 13–20. [Google Scholar] [CrossRef]
- Mei, J.; Shao, J.; Wang, Q.; Wang, H.; Yi, Y.; Ying, G. Separation and quantification of neoagaro-oligosaccharides. J. Food Sci. Technol. 2013, 50, 1217–1221. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Wu, H.; Zhang, S.; Du, Y. Enzymatic preparation of κ-carrageenan oligosaccharides and their anti-angiogenic activity. Carbohydr. Polym. 2014, 101, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Correc, G.; Hehemann, J.-H.; Czjzek, M.; Helbert, W. Structural analysis of the degradation products of porphyran digested by Zobellia galactanivorans β-porphyranase A. Carbohydr. Polym. 2011, 83, 277–283. [Google Scholar] [CrossRef]
- Hatada, Y.; Mizuno, M.; Li, Z.; Ohta, Y. Hyper-production and characterization of the ι-carrageenase useful for ι-carrageenan oligosaccharide production from a deep-sea bacterium, Microbulbifer thermotolerans JAMB-A94T, and insight into the unusual catalytic mechanism. Mar. Biotechnol. 2011, 13, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Higashimura, Y.; Naito, Y.; Takagi, T.; Mizushima, K.; Hirai, Y.; Harusato, A.; Ohnogi, H.; Yamaji, R.; Inui, H.; Nakano, Y.; et al. Oligosaccharides from agar inhibit murine intestinal inflammation through the induction of heme oxygenase-1 expression. J. Gastroenterol. 2013, 48, 897–909. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Yang, M.; Mao, X.; Mu, B.; Wei, D. Molecular cloning and expression of a new α-neoagarobiose hydrolase from Agarivorans gilvus WH0801 and enzymatic production of 3,6-anhydro-l-galactose. Biotechnol. Appl. Biochem. 2016, 63, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Han, W.J.; Gu, J.Y.; Liu, H.H.; Li, F.C.; Wu, Z.H.; Li, Y.Z. An extra peptide within the catalytic module of a β-agarase affects the agarose degradation pattern. J. Biol. Chem. 2013, 288, 9519–9531. [Google Scholar] [CrossRef] [PubMed]
- Khan, B.M.; Bakht, J. Antifungal, anti-yeast, anti-oxidant and HPLC analysis of different solvent extracted samples from Calamus aromaticus leaves. Bangladesh J. Pharmacol. 2015, 11, 91–100. [Google Scholar] [CrossRef]
- Khan, B.M.; Bakht, J.; Khan, W. Rhizome extracts of Acorus odoratus: Antifungal, anti-yeast, antioxidant and HPLC quantification. Bangladesh J. Pharmacol. 2017, 12, 44–50. [Google Scholar] [CrossRef]
- Gupta, V.; Trivedi, N.; Kumar, M.; Reddy, C.R.K.; Jha, B. Purification and characterization of exo-β-agarase from an endophytic marine bacterium and its catalytic potential in bioconversion of red algal cell wall polysaccharides into galactans. Biomass Bioenergy 2013, 49, 290–298. [Google Scholar] [CrossRef]
- Cheong, K.L.; Wu, D.T.; Zhao, J.; Li, S.P. A rapid and accurate method for the quantitative estimation of natural polysaccharides and their fractions using high performance size exclusion chromatography coupled with multi-angle laser light scattering and refractive index detector. J. Chromatogr. A 2015, 1400, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Kazłowski, B.; Kazłowska, K.; Pan, C.L.; Ko, Y.T. Evaluation of HPSEC-ELSD method for precise measurement of β-agarase activity. Biomed. Chromatogr. 2011, 25, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Leijdekkers, A.G.M.; Sanders, M.G.; Schols, H.A.; Gruppen, H. Characterizing plant cell wall derived oligosaccharides using hydrophilic interaction chromatography with mass spectrometry detection. J. Chromatogr. A 2011, 1218, 9227–9235. [Google Scholar] [CrossRef] [PubMed]
- Brokl, M.; Hernández-Hernández, O.; Soria, A.C.; Sanz, M.L. Evaluation of different operation modes of high performance liquid chromatography for the analysis of complex mixtures of neutral oligosaccharides. J. Chromatogr. A 2011, 1218, 7697–7703. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hou, Y.; Duan, D.; Zhang, Q. The structure and nephroprotective activity of oligo-porphyran on glycerol-induced acute renal failure in rats. Mar. Drugs 2017, 15, 135. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.T.; Lee, S.; Lee, D.; Kim, H.-S.; Bang, W.-G.; Kim, K.H.; Choi, I.-G. Overexpression and molecular characterization of Aga50D from Saccharophagus degradans 2-40: An exo-type β-agarase producing neoagarobiose. Appl. Microbiol. Biotechnol. 2010, 86, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.M.; Zheng, L.; Lin, W.; Yan, X.J. Product monitoring and quantitation of oligosaccharides composition in agar hydrolysates by precolumn labeling HPLC. Talanta 2004, 64, 773–777. [Google Scholar] [CrossRef] [PubMed]
- Hiltunen, S.; Sirén, H. Analysis of monosaccharides and oligosaccharides in the pulp and paper industry by use of capillary zone electrophoresis: A review. Anal. Bioanal. Chem. 2013, 405, 5773–5784. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Lin, L.; Liu, X.; Zhang, F.; Chi, L.; Xia, Q.; Linhardt, R.J. Capillary electrophoresis–mass spectrometry for the analysis of heparin oligosaccharides and low molecular weight heparin. Anal. Chem. 2016, 88, 1937–1943. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, V.; Galeotti, F.; Maccari, F.; Volpi, N. Recent advances in capillary electrophoresis separation of monosaccharides, oligosaccharides, and polysaccharides. Electrophoresis 2018, 39, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Cheong, K.L.; Wu, D.T.; Deng, Y.; Leong, F.; Zhao, J.; Zhang, W.J.; Li, S.P. Qualitation and quantification of specific polysaccharides from Panax species using GC–MS, saccharide mapping and HPSEC-RID-MALLS. Carbohydr. Polym. 2016, 153, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, P.; Yu, G.L.; Li, C.X.; Hao, C.; Qi, X.; Zhang, L.J.; Guan, H.S. Preparation and anti-influenza A virus activity of κ-carrageenan oligosaccharide and its sulphated derivatives. Food Chem. 2012, 133, 880–888. [Google Scholar] [CrossRef]
- O’Shea, M.G.; Samuel, M.S.; Konik, C.M.; Morell, M.K. Fluorophore-assisted carbohydrate electrophoresis (FACE) of oligosaccharides: Efficiency of labelling and high-resolution separation. Carbohydr. Res. 1998, 307, 1–12. [Google Scholar] [CrossRef]
- Yan, J.; Ding, J.; Liang, X. Chromatographic methods for the analysis of oligosaccharides in human milk. Anal. Methods 2017, 9, 1071–1077. [Google Scholar] [CrossRef]
- David, B.; Irague, R.; Jouanneau, D.; Daligault, F.; Czjzek, M.; Sanejouand, Y.H.; Tellier, C. Internal water dynamics control the transglycosylation/hydrolysis balance in the agarase (AgaD) of Zobellia galactanivorans. ACS Catal. 2017, 7, 3357–3367. [Google Scholar] [CrossRef]
- Jorge, P.; Abdul-Wajid, A. Sialyl-Tn-KLH, glycoconjugate analysis and stability by high-pH anion-exchange chromatography with pulsed amperometric detection (HPAEC-PAD). Glycobiology 1995, 5, 759. [Google Scholar] [CrossRef] [PubMed]
- Kailemia, M.J.; Ruhaak, L.R.; Lebrilla, C.B.; Amster, I.J. Oligosaccharide analysis by mass spectrometry: A review of recent developments. Anal. Chem. 2014, 86, 196–212. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.G.; Ducatti, D.R.B.; Grindley, T.B.; Duarte, M.E.R.; Noseda, M.D. ESI-MS differential fragmentation of positional isomers of sulfated oligosaccharides derived from carrageenans and agarans. J. Am. Soc. Mass Spectrom. 2010, 21, 1404–1416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.H.; Kim, H.T.; Yun, E.J.; Lee, A.R.; Kim, S.R.; Kim, J.-H.; Choi, I.-G.; Kim, K.H. A novel agarolytic β-galactosidase acts on agarooligosaccharides for complete hydrolysis of agarose into monomers. Appl. Environ. Microbiol. 2014, 80, 5965–5973. [Google Scholar] [CrossRef] [PubMed]
- Temuujin, U.; Chi, W.-J.; Lee, S.-Y.; Chang, Y.-K.; Hong, S.-K. Overexpression and biochemical characterization of DagA from Streptomyces coelicolor A3(2): An endo-type β-agarase producing neoagarotetraose and neoagarohexaose. Appl. Microbiol. Biotechnol. 2011, 92, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Lundborg, M.; Widmalm, G. Structural analysis of glycans by NMR chemical shift prediction. Anal. Chem. 2011, 83, 1514–1517. [Google Scholar] [CrossRef] [PubMed]
- Préchoux, A.; Helbert, W. Preparation and detailed NMR analyses of a series of oligo-α-carrageenans. Carbohydr. Polym. 2014, 101, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Campo, V.L.; Kawano, D.F.; Silva, D.B.d.; Carvalho, I. Carrageenans: Biological properties, chemical modifications and structural analysis—A review. Carbohydr. Polym. 2009, 77, 167–180. [Google Scholar] [CrossRef]
- Yao, Z.-a.; Xu, L.; Wu, H.-g. Immunomodulatory function of κ-carrageenan oligosaccharides acting on LPS-activated microglial cells. Neurochem. Res. 2014, 39, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Pei, L.; Cui, H.; Wu, L.; Wan, W.; Mao, X. Neoagaro-oligosaccharide monomers inhibit inflammation in LPS-stimulated macrophages through suppression of MAPK and NF-κB pathways. Sci. Rep. 2017, 7, 44252. [Google Scholar] [Green Version]
- Higashimura, Y.; Naito, Y.; Takagi, T.; Uchiyama, K.; Mizushima, K.; Ushiroda, C.; Ohnogi, H.; Kudo, Y.; Yasui, M.; Inui, S.; et al. Protective effect of agaro-oligosaccharides on gut dysbiosis and colon tumorigenesis in high-fat diet-fed mice. Am. J. Physiol.–Gastrointest. Liver Physiol. 2016, 310, G367. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Beatty, E.R.; Wang, X.; Cummings, J.H. Selective stimulation of bifidobacteria in the human colon by oligofructose and inulin. Gastroenterology 1995, 108, 975–982. [Google Scholar] [CrossRef]
- Li, M.; Shang, Q.; Li, G.; Wang, X.; Yu, G. Degradation of marine algae-derived carbohydrates by Bacteroidetes isolated from human gut microbiota. Mar. Drugs 2017, 15, 92. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Lin, S.; Zheng, B.; Cheung, P.C.K. Short-chain fatty acids in control of energy metabolism. Crit. Rev. Food Sci. Nutr. 2018, 58, 1243–1249. [Google Scholar] [CrossRef] [PubMed]
- Kalitnik, A.A.; Anastyuk, S.D.; Sokolova, E.V.; Kravchenko, A.O.; Khasina, E.I.; Yermak, I.M. Oligosaccharides of κ/β-carrageenan from the red alga Tichocarpus crinitus and their ability to induce interleukin 10. J. Appl. Phycol. 2016, 28, 545–553. [Google Scholar] [CrossRef]
- Liu, Y.; Jiang, L.; Li, X. κ-carrageenan-derived pentasaccharide attenuates Aβ25-35-induced apoptosis in SH-SY5Y cells via suppression of the JNK signaling pathway. Mol. Med. Rep. 2016, 15, 285. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hu, S.; Nie, S.; Yu, Q.; Xie, M. Reviews on mechanisms of in vitro antioxidant activity of polysaccharides. Oxid. Med. Cell. Longev. 2016, 2016, 13. [Google Scholar] [CrossRef] [PubMed]
- Brar, S.K.; Dhillon, G.S.; Soccol, C.R. Biotransformation of Waste Biomass into High Value biochemicals; Springer: New York, NY, USA, 2014. [Google Scholar]
- Ma, D.L.; Liu, L.J.; Leung, K.H.; Chen, Y.T.; Zhong, H.J.; Chan, D.S.; Wang, H.M.; Leung, C.H. Antagonizing STAT3 dimerization with a rhodium(III) complex. Angew. Chem. Int. Ed. Engl. 2015, 53, 9178–9182. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.M.; Chen, C.Y.; Wen, Z.H. Identifying melanogenesis inhibitors from Cinnamomum subavenium with in vitro and in vivo screening systems by targeting the human tyrosinase. Exp. Dermatol. 2015, 20, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Briganti, S.; Camera, E.; Picardo, M. Chemical and instrumental approaches to treat hyperpigmentation. Pigment. Cell Melanoma Res. 2010, 16, 101–110. [Google Scholar] [CrossRef]
- Kobayashi, R.; Takisada, M.; Suzuki, T.; Kirimura, K.; Usami, S. Neoagarobiose as a novel moisturizer with whitening effect. Biosci. Biotechnol. Biochem. 1997, 61, 162–163. [Google Scholar] [CrossRef] [PubMed]
- Yun, E.J.; Lee, S.; Kim, J.H.; Kim, B.B.; Kim, H.T.; Lee, S.H.; Pelton, J.G.; Kang, N.J.; Choi, I.-G.; Kim, K.H. Enzymatic production of 3,6-anhydro-L-galactose from agarose and its purification and in vitro skin whitening and anti-inflammatory activities. Appl. Microbiol. Biotechnol. 2013, 97, 2961–2970. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.K.; Lee, D.G.; Kim, N.Y.; Yu, K.H.; Jang, H.J.; Lee, S.W.; Jang, H.J.; Lee, Y.J.; Lee, S.H. Purification and characterization of neoagarotetraose from hydrolyzed agar. J. Microbiol. Biotechnol. 2009, 19, 1197–1200. [Google Scholar] [PubMed]
- Stephanie, B.; Eric, D.; Sophie, F.M.; Christian, B.; Yu, G. Carrageenan from Solieria chordalis (Gigartinales): Structural analysis and immunological activities of the low molecular weight fractions. Carbohydr. Polym. 2010, 81, 448–460. [Google Scholar] [CrossRef]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheong, K.-L.; Qiu, H.-M.; Du, H.; Liu, Y.; Khan, B.M. Oligosaccharides Derived from Red Seaweed: Production, Properties, and Potential Health and Cosmetic Applications. Molecules 2018, 23, 2451. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23102451
Cheong K-L, Qiu H-M, Du H, Liu Y, Khan BM. Oligosaccharides Derived from Red Seaweed: Production, Properties, and Potential Health and Cosmetic Applications. Molecules. 2018; 23(10):2451. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23102451
Chicago/Turabian StyleCheong, Kit-Leong, Hua-Mai Qiu, Hong Du, Yang Liu, and Bilal Muhammad Khan. 2018. "Oligosaccharides Derived from Red Seaweed: Production, Properties, and Potential Health and Cosmetic Applications" Molecules 23, no. 10: 2451. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23102451