Bioformation of Volatile and Nonvolatile Metabolites by Saccharomycopsis fibuligera KJJ81 Cultivated under Different Conditions—Carbon Sources and Cultivation Times


Abstract
:1. Introduction
2. Results and Discussion
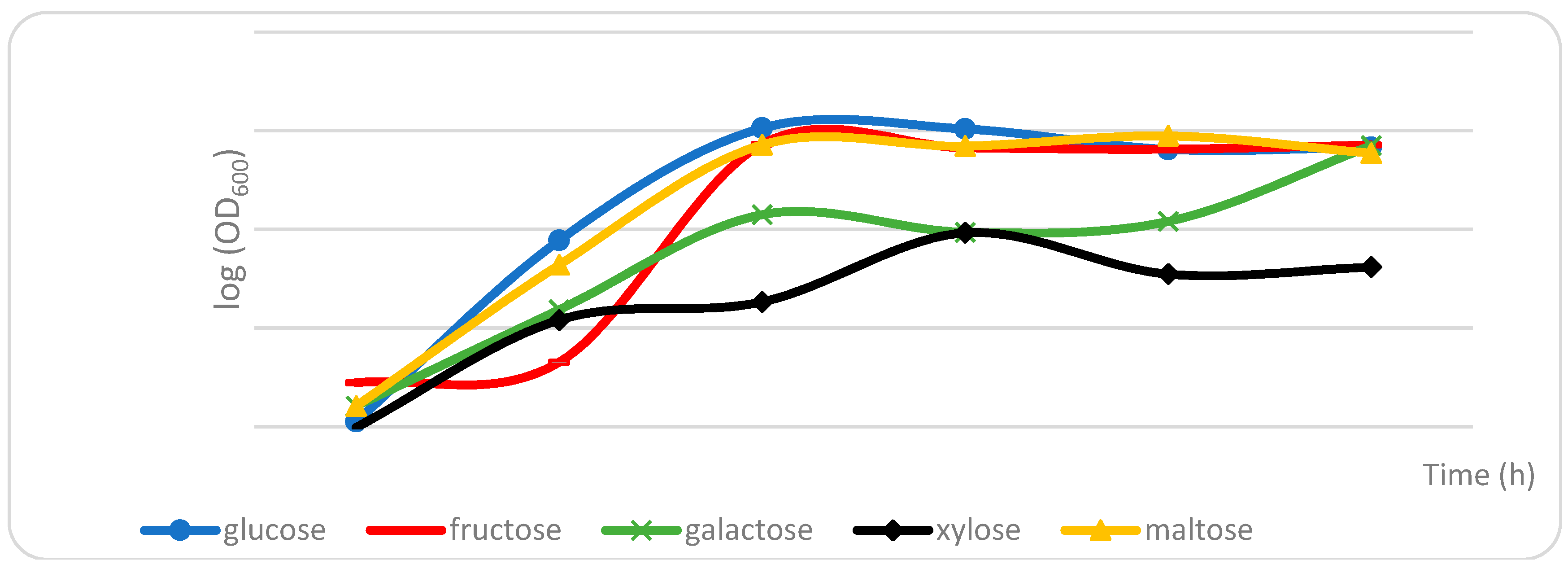
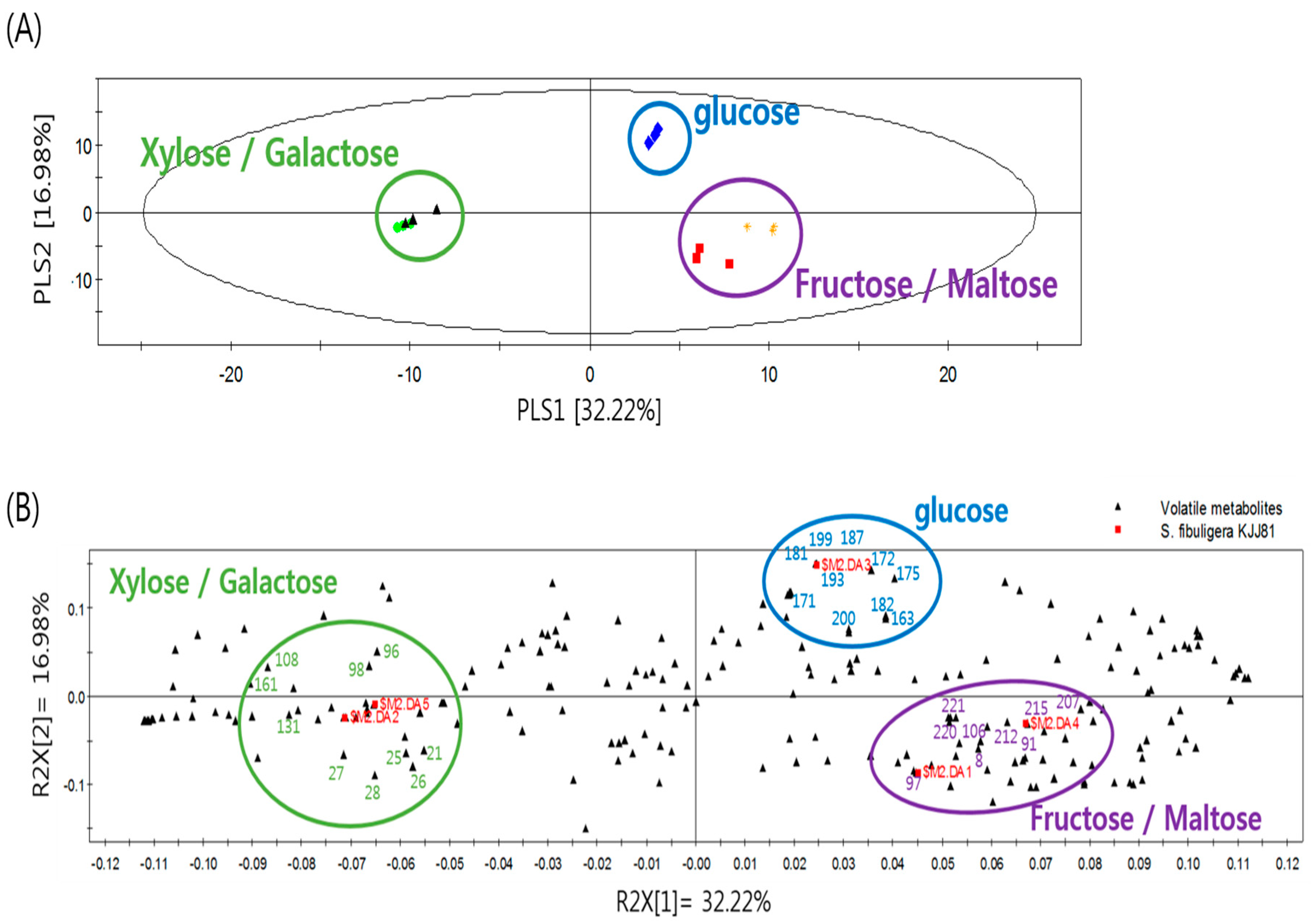
2.1. Effects of Carbon Sources on the Formation of Volatile Metabolites by S. fibuligera KJJ81
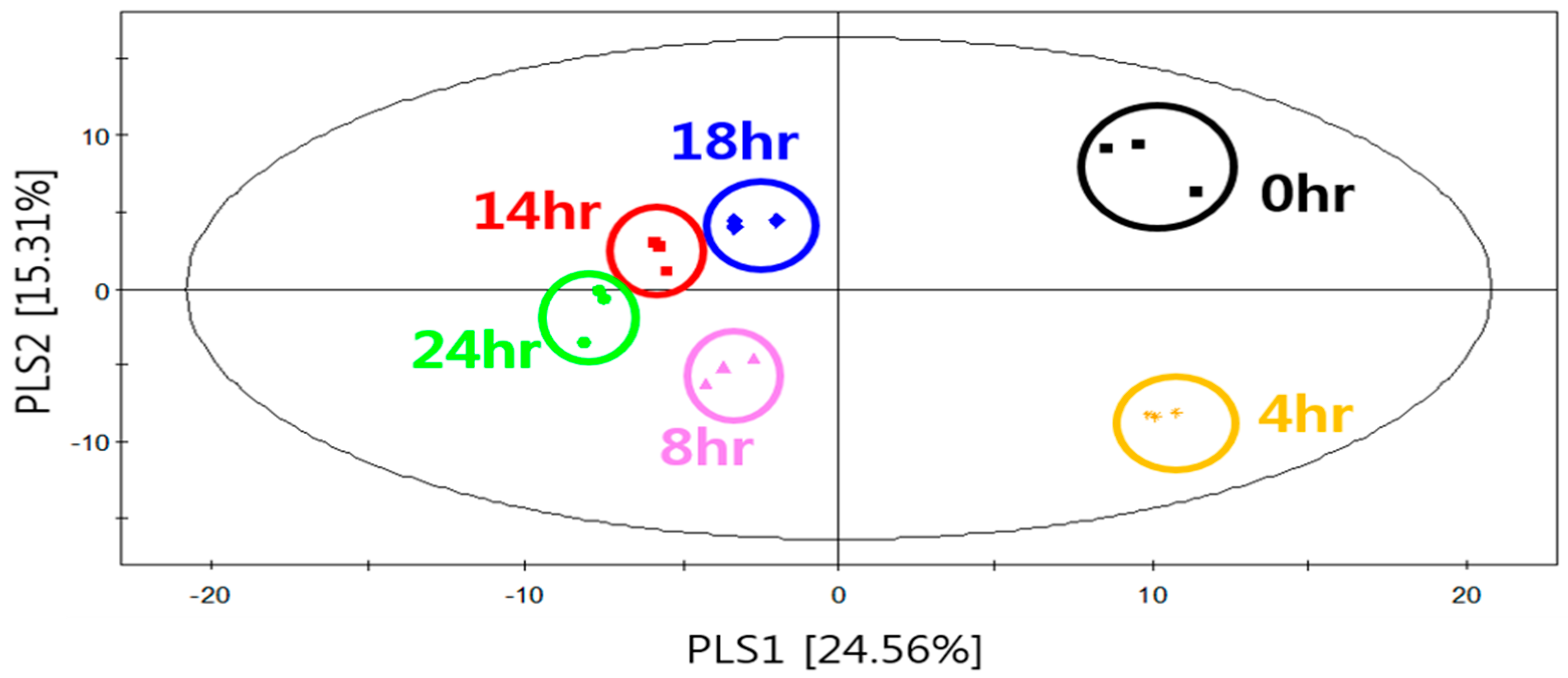
2.2. Cultivation-Time-Dependent Volatile Metabolites of S. fibuligera KJJ81 in YPD Medium
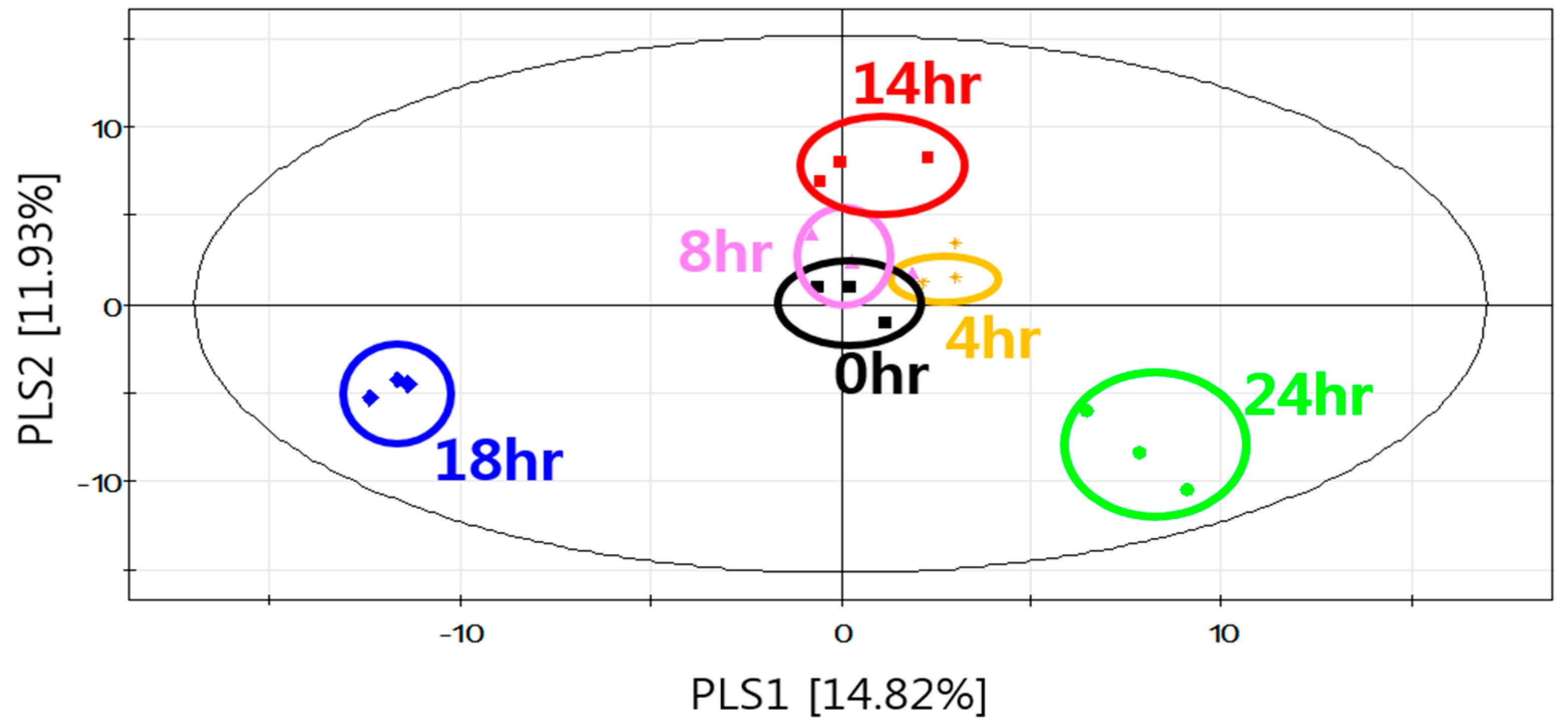
2.3. Cultivation-time-dependent Nonvolatile Metabolites of S. fibuligera KJJ81 in YPD Medium
3. Materials and Methods
3.1. Cultivation of S. fibuligera
3.2. Extraction and Analysis of Volatile Metabolites by GC-MS
3.3. Extraction and Analysis of Nonvolatile Metabolites by GC-TOF/MS
3.4. Mass Spectral Data Processing of Volatile and Nonvolatile Metabolites
3.5. Identification and Quantification of Volatile and Nonvolatile Metabolites
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Carroll, E.; Trinh, T.N.; Son, H.; Lee, Y.W.; Seo, J.A. Comprehensive analysis of fungal diversity and enzyme activity in nuruk, a Korean fermenting starter, for acquiring useful fungi. J. Microbiol. 2017, 55, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.; Lee, S.J.; Lim, J.H.; Kim, B.K.; Park, K.J. Chemical and sensory profiles of makgeolli, Korean commercial rice wine, from descriptive, chemical, and volatile compound analyses. Food Chem. 2014, 152, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.; Nam, Y.; Roh, S.W.; Bae, J. Unexpected convergence of fungal and bacterial communities during fermentation of traditional Korean alcoholic beverages inoculated with various natural starters. Food Microbiol. 2012, 30, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Kim, J.H.; Bai, D.H.; Ahn, B.H. Identification and characterization of useful fungi with α-amylase activity from the Korean traditional nuruk. Mycobiology 2011, 39, 278–282. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Lee, S.M.; Song, S.H.; Kim, Y.S. Characterization of volatile components in Makgeolli, a traditional Korean rice wine, with or without pasteurization, during storage. Molecules 2013, 18, 5317–5325. [Google Scholar] [CrossRef] [PubMed]
- Kang, B.S.; Lee, J.E.; Park, H.J. Electronic tongue-based discrimination of Korean rice wines (makgeolli) including prediction of sensory evaluation and instrumental measurements. Food Chem. 2014, 151, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Son, E.Y.; Lee, S.M.; Kim, M.; Seo, J.A.; Kim, Y.S. Comparison of volatile and non-volatile metabolites in rice wine fermented by Koji inoculated with Saccharomycopsis fibuligera and Aspergillus oryzae. Food Res. Int. 2018, 109, 596–605. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.H.; Park, E.H.; Kim, M.D. Characterization of starch-utilizing yeast Saccharomycopsis fibuligera isolated from Nuruk. J. Microbiol. Biotechnol. 2014, 42, 407–412. [Google Scholar] [CrossRef]
- Chi, Z.; Chi, Z.; Liu, G.; Wang, F.; Ju, L.; Zhang, T. Saccharomycopsis fibuligera and its applications in biotechnology. Biotechnol. Adv. 2009, 27, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Hostinová, E. Amylolytic enzymes produced by the yeast Saccharomycopsis fibuligera. Biologia 2002, 57, 247–252. [Google Scholar]
- Aidoo, K.E.; Nout, M.R.; Sarkar, P.K. Occurrence and function of yeasts in Asian indigenous fermented foods. FEMS Yeast Res. 2006, 6, 30–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, Z.; Liu, J.; Zhang, W. Trehalose accumulation from soluble starch by Saccharomycopsis fibuligera sdu. Enzyme Microb. Technol. 2001, 28, 240–245. [Google Scholar] [CrossRef]
- Lenz, E.M.; Wilson, I.D. Analytical Strategies in Metabonomics. J. Proteome Res. 2007, 6, 443–458. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.Z.; Cheng, J.S.; Xiao, W.H. Comparative metabolomic analysis on industrial continuous and batch ethanol fermentation processes by GC–TOF-MS. Metabolomics 2009, 5, 229–238. [Google Scholar] [CrossRef]
- Jonsson, P.; Johansson, A.I.; Gullberg, J. High-throughput data analysis for detecting and identifying differences between samples in GC/MS-based metabolomic analyses. Anal. Chem. 2005, 77, 5635–5642. [Google Scholar] [CrossRef] [PubMed]
- Vilanova, M.; Ugliano, M.; Varela, C.; Siebert, T.; Pretorius, I.S.; Henschke, P.A. Assimilable nitrogen utilisation and production of volatile and non-volatile compounds in chemically defined medium by Saccharomyces cerevisiae wine yeasts. Appl. Microbiol. Biotechnol. 2007, 77, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Soares, E.V.; Vroman, A.; Mortier, J.; Rijsbrack, K.; Mota, M. Carbohydrate carbon sources induce loss of flocculation of an ale-brewing yeast strain. J. Appl. Microbiol. 2004, 96, 1117–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, X.; Eksriwong, T.; Chauvatcharin, S.; Zhong, J. Optimization of carbon source and carbon/nitrogen ratio for cordycepin production by submerged cultivation of medicinal mushroom Cordyceps militaris. Process Biochem. 2005, 40, 1667–1672. [Google Scholar] [CrossRef]
- Adams, T.H.; Yu, J.H. Coordinate control of secondary metabolite production and asexual sporulation in Aspergillus nidulans. Curr. Opin. Microbiol. 1998, 1, 674–677. [Google Scholar] [CrossRef]
- Connor, M.R.; Liao, J.C. Engineering of an Escherichia coli strain for the production of 3-methyl-1-butanol. Appl. Environ. Microbiol. 2008, 74, 5769–5775. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Vadlani, P.; Harrison, M.; Bennett, G.; San, K. Aerobic production of isoamyl acetate by overexpression of the yeast alcohol acetyl-transferases AFT1 and AFT2 in Escherichia coli and using low-cost fermentation ingredients. Bioprocess Biosyst. Eng. 2008, 31, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Schoondermark-Stolk, S.A.; Jansen, M.; Veurink, J.H.; Verkleij, A.J.; Verrips, C.T.; Euverink, G.W.; Boonstra, J.; Dijkhuizen, L. Rapid identification of target genes for 3-methyl-1-butanol production in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2006, 70, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Plata, C.; Mauricio, J.C.; Millan, C.; Ortega, J.M. Influence of glucose and oxygen on the production of ethyl acetate and isoamyl acetate by a Saccharomyces cerevisiae strain during alcoholic fermentation. World J. Microbiol. Biotechnol. 2005, 21, 115–121. [Google Scholar] [CrossRef]
- Assaf, S.; Hadar, Y.; Dosoretz, C.G. 1-Octen-3-ol and 13-hydroperoxylinoleate are products of distinct pathways in the oxidative breakdown of linoleic acid by Pleurotus pulmonarius. Enzyme Microb. Technol. 1997, 21, 484–490. [Google Scholar] [CrossRef]
- Pagot, Y.; Le Clainche, A.; Nicaud, J.; Wache, Y.; Belin, J. Peroxisomal β-oxidation activities and γ-decalactone production by the yeast Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 1998, 49, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Romero-Guido, C.; Belo, I.; Ta, T.M.N.; Cao-Hoang, L.; Alchihab, M.; Gomes, N.; Wache, Y. Biochemistry of lactone formation in yeast and fungi and its utilization for the production of flavor and fragrance compounds. Appl. Microbiol. Biotechnol. 2011, 89, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Ravasio, D.; Wendland, J.; Walther, A. Major contribution of the Ehrlich pathway for 2-phenylethanol/rose flavor production in Ashbya gossypii. FEMS Yeast Res. 2014, 14, 833–844. [Google Scholar] [CrossRef] [PubMed]
- Viana, F.; Belloch, C.; Vallés, S.; Manzanares, P. Monitoring a mixed starter of Hanseniaspora vineae–Saccharomyces cerevisiae in natural must: Impact on 2-phenylethyl acetate production. Int. J. Food Microbiol. 2011, 151, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xu, Y. The influence of yeast strains on the volatile flavour compounds of Chinese rice wine. J. Inst. Brew. 2010, 116, 190–196. [Google Scholar] [CrossRef]
- Saerens, S.M.; Delvaux, F.R.; Verstrepen, K.J.; Thevelein, J.M. Production and biological function of volatile esters in Saccharomyces cerevisiae. Microb. Biotechnol. 2010, 3, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Xu, E.; Long, J.; Wu, Z.; Li, H.; Wang, F.; Xu, X.; Jiao, A. Characterization of volatile flavor compounds in Chinese rice wine fermented from enzymatic extruded rice. J. Food Sci. 2015, 80, C1476–C1489. [Google Scholar] [CrossRef] [PubMed]
- Aoki, M.A.; Pastore, G.M.; Park, Y.K. Microbial transformation of sucrose and glucose to erythritol. Biotechnol. Lett. 1993, 15, 383–388. [Google Scholar] [CrossRef]
- Tomaszewska, L.; Rywińska, A.; Gładkowski, W. Production of erythritol and mannitol by Yarrowia lipolytica yeast in media containing glycerol. J. Ind. Microbiol. Biotechnol. 2012, 39, 1333–1343. [Google Scholar] [CrossRef] [PubMed]
- Onishi, H.; Suzuki, T. Production of D-mannitol and glycerol by yeasts. Appl. Microbiol. 1968, 16, 1847–1852. [Google Scholar] [PubMed]
- Lee, D.E.; Lee, S.; Jang, E.S.; Shin, H.W.; Moon, B.S.; Lee, C.H. Metabolomic profiles of Aspergillus oryzae and Bacillus amyloliquefaciens during Rice Koji fermentation. Molecules 2016, 21, 773. [Google Scholar] [CrossRef] [PubMed]
- Lomakin, I.B.; Xiong, Y.; Steitz, T.A. The crystal structure of yeast fatty acid synthase, a cellular machine with eight active sites working together. Cell 2007, 129, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Dai, X.; Zhu, J.; Yu, H. Classification of Chinese Rice wine according to geographic origin and wine age based on chemometric methods and SBSE-TD-GC-MS analysis of volatile compounds. Food Sci. Technol. Res. 2015, 21, 371–380. [Google Scholar] [CrossRef]
- Foster, J.; Nakata, P.A. An oxalyl-CoA synthetase is important for oxalate metabolism in Saccharomyces cerevisiae. FEBS Lett. 2014, 588, 160–166. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Dong, J.; Yin, H.; Zhao, Y.; Chen, R.; Wan, X.; Chen, P.; Hou, X.; Liu, J.; Chen, L. Wort composition and its impact on the flavour-active higher alcohol and ester formation of beer—A review. J. Inst. Brew. 2014, 120, 157–163. [Google Scholar] [CrossRef]
- Swiegers, J.; Bartowsky, E.; Henschke, P.; Pretorius, I. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Hazelwood, L.A.; Daran, J.M.; van Maris, A.J.; Pronk, J.T.; Dickinson, J.R. The Ehrlich pathway for fusel alcohol production: A century of research on Saccharomyces cerevisiae metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef] [PubMed]
- Choo, J.H.; Hong, C.P.; Lim, J.Y.; Seo, J.A.; Kim, Y.S.; Lee, D.W.; Kang, H.A. Whole-genome de novo sequencing, combined with RNA-Seq analysis, reveals unique genome and physiological features of the amylolytic yeast Saccharomycopsis fibuligera and its interspecies hybrid. Biotechnol. Biofuels 2016, 9, 246–268. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, D.Y.; Wohlgemuth, G.; Park, H.S.; Fiehn, O.; Kim, K.H. Evaluation and optimization of metabolome sample preparation methods for Saccharomyces cerevisiae. Anal. Chem. 2013, 85, 2169–2176. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples are not available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | RI 1 | Major Volatile Metabolites 2 | VIP 3 | ID 4 |
|---|---|---|---|---|
| Media Containing Xylose and Galactose | ||||
| Alcohols | ||||
| 98 | 1560 | 1-Octanol | 0.80 | A |
| Benzenes & Benzene derivatives | ||||
| 96 | 1429 | 1,3-Bis(2-methyl-2-propanyl)benzene | 1.11 | B |
| Esters | ||||
| 161 | 2261 | (2E,6E)-3,7,11-Trimethyl-2,6,10-dodecatrien-1-yl acetate | 0.79 | B |
| 131 | 1680 | 4-(2-Methyl-2-propanyl)cyclohexyl acetate | 0.74 | B |
| Furans | ||||
| 108 | 2394 | 5-[(2E)-2-Octen-1-yl]dihydro-2(3H)-furanone | 0.74 | B |
| Media Containing Glucose | ||||
| Alcohols | ||||
| 187 | 1910 | 2-Phenylethanol | 1.42 | A |
| 175 | 1709 | (2E)-3,7-Dimethyl-2,6-octadien-1-ol | 1.25 | B |
| Aldehydes | ||||
| 181 | 1750 | 3-Dodecenal | 1.42 | B |
| 199 | 1287 | Octanal | 1.42 | B |
| 172 | 1429 | 2-Octenal | 1.33 | B |
| Esters | ||||
| 183 | 1813 | 2-Phenylethyl acetate | 1.12 | A |
| 182 | 1782 | Ethyl phenylacetate | 0.90 | A |
| 163 | <1100 | Ethyl acetate | 0.87 | A |
| Lactones | ||||
| 193 | 2424 | 6-Heptyltetrahydro-2H-pyran-2-one | 1.43 | A |
| Media Containing Fructose and Maltose | ||||
| Alcohols | ||||
| 97 | 1452 | 1-Octen-3-ol | 1.59 | A |
| 212 | 1659 | 2-Furylmethanol | 1.51 | A |
| 106 | 2003 | Phenol | 1.25 | A |
| 91 | 1218 | 3-Methyl-1-butanol | 0.86 | A |
| Esters | ||||
| 220 | 1128 | 3-Methylbutyl acetate | 1.26 | A |
| Furans | ||||
| 83 | 2375 | 5-Hexyldihydro-2(3H)-furanone | 0.87 | A |
| Sulfur-containing compounds | ||||
| 8 | 1059 | (Methyldisulfanyl)methane | 0.84 | A |
| VIP 1 | Volatile Metabolites 2 | RI 3 | Relative Peak Area (%) 4 | |||||
|---|---|---|---|---|---|---|---|---|
| 0 h 5 | 4 h | 8 h | 14 h | 18 h | 24 h | |||
| Acids | ||||||||
| 1.17 | Butyric acid | 1625 | N.D. 6 a 7 | 0.02 ± 0.01 a | 0.09 ± 0.04 ab | 0.27 ± 0.02 d | 0.14 ± 0.03 bc | 0.24 ± 0.14 cd |
| 1.36 | 2-Methylpropanoic acid | 1568 | N.D. a | N.D. a | 0.01 ± 0.00 a | 0.02 ± 0.00 b | N.D. a | 0.04 ± 0.01 c |
| Alcohols | ||||||||
| 1.10 | 2-Methyl-1-propanol | 1113 | N.D. a | 0.04 ± 0.01 a | 0.43 ± 0.06 b | 0.59 ± 0.07 c | 0.44 ± 0.01 b | 0.46 ± 0.00 b |
| 1.17 | (2E)-3,7-Dimethyl-2,6-octadien-1-ol | 1847 | N.D. a | N.D. a | 0.01 ± 0.01 a | 0.04 ± 0.00 b | 0.03 ± 0.00 b | 0.03 ± 0.01 b |
| 1.17 | Phenol | 1997 | 0.03 ± 0.01 bc | 0.01 ± 0.00 a | N.D. a | 0.03 ± 0.01 bc | 0.02 ± 0.00 b | 0.04 ± 0.01 c |
| 1.19 | 1-Butanol | 1160 | 2.22 ± 0.93 b | 0.91 ± 0.04 a | 0.32 ± 0.03 a | 0.41 ± 0.06 a | 0.25 ± 0.01 a | 0.32 ± 0.02 a |
| 1.27 | 2,4-Bis(2-methyl-2-propanyl)phenol | 2307 | 0.18 ± 0.01 d | 0.13 ± 0.01 ab | 0.14 ± 0.01 bc | 0.15 ± 0.01 bc | 0.12 ± 0.00 a | 0.16 ± 0.02 c |
| 1.29 | 3-Methyl-1-butanol | 1220 | 0.18 ± 0.05 a | 2.24 ± 0.12 b | 4.25 ± 0.21 c | 5.02 ± 0.48 d | 4.67 ± 0.26 cd | 4.85 ± 0.28 d |
| 1.40 | 2-Phenylethanol | 1908 | 0.21 ± 0.04 a | 2.88 ± 0.12 b | 4.63 ± 0.35 c | 5.40 ± 0.40 d | 5.38 ± 0.32 d | 6.18 ± 0.41 e |
| Aldehydes | ||||||||
| 1.23 | 2-Furaldehyde | 1458 | 0.04 ± 0.01 a | 0.01 ± 0.00 a | 0.02 ± 0.00 a | 0.07 ± 0.02 bc | 0.10 ± 0.05 c | 0.05 ± 0.01 ab |
| 1.47 | Octanal | 1287 | N.D. a | 0.01 ± 0.00 b | N.D. a | 0.06 ± 0.01 d | 0.07 ± 0.0037 c | 0.03 ± 0.00 c |
| Benzen and benzene derivatives | ||||||||
| 1.25 | Benzaldehyde | 1518 | 0.16 ± 0.05 b | 0.04 ± 0.01 a | 0.05 ± 0.05 a | 0.03 ± 0.00 a | 0.03 ± 0.00 a | 0.03 ± 0.00 a |
| 1.31 | 1-(2-Aminophenyl)ethanone | 2210 | 0.01 ± 0.00 a | 0.02 ± 0.00 b | 0.02 ± 0.00 b | 0.02 ± 0.00 ab | 0.02 ± 0.00 a | 0.02 ± 0.00 a |
| 1.35 | 3-Methylbenzaldehyde | 1642 | 0.05 ± 0.02 c | N.D. a | N.D. a | 0.06 ± 0.01 cd | 0.03 ± 0.00 b | 0.07 ± 0.01 d |
| Esters | ||||||||
| 1.11 | Ethyl butyrate | <1100 | N.D. a | N.D. a | 0.44 ± 0.07 b | 1.15 ± 0.13 d | 0.91 ± 0.05 c | 0.79 ± 0.08 c |
| 1.14 | (2E)-3,7-Dimethyl-2,6-octadien-1-yl acetate | 1708 | N.D. a | N.D. a | 0.03 ± 0.00 c | 0.03 ± 0.01 c | 0.03 ± 0.00 c | 0.02 ± 0.00 b |
| 1.15 | 3-Methylbutyl acetate | 1130 | 0.08 ± 0.01 a | 0.30 ± 0.01 a | 2.29 ± 0.16 c | 4.14 ± 0.36 e | 3.48 ± 0.17 d | 1.59 ± 0.06 b |
| 1.17 | 2-Phenylethyl acetate | 1812 | 0.05 ± 0.01 a | 0.25 ± 0.01 b | 1.27 ± 0.12 c | 1.63 ± 0.14 d | 1.27 ± 0.08 c | 1.56 ± 0.08 d |
| 1.27 | (2E,6E)-3,7,11-Trimethyl-2,6,10-dodecatrien-1-yl acetate | 2260 | 0.02 ± 0.01 a | 0.02 ± 0.00 a | 0.09 ± 0.01 c | 0.07 ± 0.02 b | 0.09 ± 0.01 c | 0.07 ± 0.00 b |
| 1.37 | Ethyl 3-phenylacrylate | 2125 | N.D. a | 0.16 ± 0.01 b | 0.35 ± 0.01 d | 0.47 ± 0.05 e | 0.22 ± 0.01 c | 0.33 ± 0.04 d |
| 1.45 | Butyl 3-phenylacrylate | 2333 | N.D. a | 0.07 ± 0.01 b | 0.09 ± 0.01 b | 0.09 ± 0.01 b | 0.08 ± 0.01 b | 0.07 ± 0.02 b |
| Hydrocarbons | ||||||||
| 1.18 | Hexadecane | 1598 | 0.02 ± 0.00 ab | 0.01 ± 0.00 a | 0.01 ± 0.01 a | 0.02 ± 0.00 bc | 0.02 ± 0.01 c | 0.02 ± 0.00 bc |
| Lactones | ||||||||
| 1.21 | 6-Heptyltetrahydro-2H-pyran-2-one | 2423 | N.D. a | 0.02 ± 0.00 b | 0.06 ± 0.00 c | 0.14 ± 0.02 e | 0.09 ± 0.01 d | 0.10 ± 0.01 d |
| 1.22 | 6-Pentyltetrahydro-2H-pyran-2-one | 2189 | N.D. a | 0.02 ± 0.00 b | 0.05 ± 0.00 c | 0.06 ± 0.01 c | 0.06 ± 0.00 c | 0.06 ± 0.00 c |
| 1.25 | 5-Hexyldihydro-2(3H)-furanone | 2139 | 0.05 ± 0.00 a | 0.27 ± 0.03 b | 0.66 ± 0.01 cd | 0.74 ± 0.09 d | 0.61 ± 0.02 c | 0.65 ± 0.06 cd |
| 1.27 | 5-[(2Z)-2-Octen-1-yl]dihydro-2(3H)-furanone | 2393 | N.D. a | 0.03 ± 0.01 b | 0.16 ± 0.01 c | 0.23 ± 0.03 d | 0.21 ± 0.01 d | 0.17 ± 0.02 c |
| 1.41 | 5-Pentyldihydro-2(3H)-furanone | 2023 | 0.01 ± 0.00 a | 0.07 ± 0.00 b | 0.08 ± 0.02 bc | 0.10 ± 0.02 c | 0.08 ± 0.00 b | 0.09 ± 0.01 bc |
| Pyrazines | ||||||||
| 1.16 | 2-Isopropyl-5-methylpyrazine | 1413 | 0.01 ± 0.00 a | 0.02 ± 0.00 b | 0.01 ± 0.01 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a |
| 1.17 | 3-Ethyl-2,5-dimethylpyrazine | 1446 | 0.03 ± 0.01 a | 0.03 ± 0.00 a | 0.04 ± 0.00 b | 0.03 ± 0.01 a | 0.03 ± 0.00 a | 0.04 ± 0.00 b |
| 1.21 | 2-Butyl-3,5-dimethylpyrazine | 1610 | 0.02 ± 0.00 a | 0.02 ± 0.00 a | 0.02 ± 0.00 a | 0.03 ± 0.00 c | 0.03 ± 0.01 c | 0.02 ± 0.00 b |
| 1.22 | 2,5-Dimethylpyrazine | 1320 | 0.08 ± 0.01 ab | 0.11 ± 0.01 c | 0.09 ± 0.01 b | 0.07 ± 0.01 a | 0.07 ± 0.01 a | 0.07 ± 0.01 a |
| 1.25 | 3-Butyl-2,5-dimethylpyrazine | 1658 | N.D. a | N.D. a | 0.01 ± 0.00 a | 0.10 ± 0.05 b | 0.01 ± 0.00 a | 0.01 ± 0.00 a |
| VIP 1 | Non-Volatile Metabolite 2 | MS 3 | Relative Peak Area (%) 4 | |||||
|---|---|---|---|---|---|---|---|---|
| 0 h 5 | 4 h | 8 h | 14 h | 18 h | 24 h | |||
| Carbohydrates | ||||||||
| 1.46 | Mannose | 147 | 0.56 ± 0.03 d 6 | 0.11 ± 0.09 b | 0.01 ± 0.00 a | 0.02 ± 0.00 a | 0.09 ± 0.01 b | 0.48 ± 0.01 c |
| 1.19 | Arabitol | 217 | 0.35 ± 0.31 b | 1.07 ± 0.84 c | 0.24 ± 0.02 a | 0.04 ± 0.00 a | 0.01 ± 0.00 a | 0.28 ± 0.01 a |
| 0.73 | Mannitol | 147 | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.01 a | 0.01 ± 0.00 a | N.D. 7 a | 0.03 ± 0.00 b |
| Lipids | ||||||||
| 1.87 | Propionic acid | 147 | 0.22 ± 0.01 c | 0.09 ± 0.01 b | 0.23 ± 0.04 c | 0.01 ± 0.00 a | 0.50 ± 0.04 d | 0.16 ± 0.01 b |
| 1.37 | Palmitic acid | 117 | 3.90 ± 0.22 b | 4.87 ± 0.16 c | 3.75 ± 0.40 b | 3.06 ± 0.34 a | 3.74 ± 0.43 b | 3.03 ± 0.37 a |
| 1.30 | Stearic acid | 117 | 4.19 ± 0.23 b | 5.26 ± 0.14 c | 4.19 ± 0.38 b | 3.38 ± 0.43 a | 4.26 ± 0.50 b | 3.31 ± 0.39 a |
| 1.01 | Myristic acid | 117 | 0.12 ± 0.01 ab | 0.18 ± 0.03 d | 0.14 ± 0.01 bc | 0.14 ± 0.01 bc | 0.10 ± 0.01 a | 0.11 ± 0.02 ab |
| Organic acids | ||||||||
| 1.53 | Oxalic acid1 | 147 | 0.01 ± 0.00 a | 1.19 ± 2.02 b | 1.76 ± 2.99 c | 0.01 ± 0.01 a | N.D. a | 0.01 ± 0.00 a |
| 1.48 | Oxalic acid2 | 147 | 9.95 ± 1.78 b | 5.30 ± 1.70 a | 5.12 ± 1.87 a | 28.11 ± 2.14 d | 10.22 ± 1.66 b | 18.37 ± 1.69 c |
| 0.92 | Succinic acid | 147 | 0.01 ± 0.01 a | 0.01 ± 0.00 a | 0.01 ± 0.01 a | 0.02 ± 0.03 a | N.D. a | 0.03 ± 0.06 a |
| Amino acids | ||||||||
| 1.40 | Isoleucine | 158 | 0.01 ± 0.00 a | 0.01 ± 0.001 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.03 ± 0.00 b | 0.15 ± 0.00 c |
| 1.38 | Serine | 204 | 0.01 ± 0.00 b | 0.06 ± 0.00 e | 0.01 ± 0.00 b | 0.03 ± 0.00 c | 0.00 ± 0.00 a | 0.05 ± 0.00 d |
| 1.20 | Alanine | 116 | 0.27 ± 0.01 c | 0.40 ± 0.03 d | 0.17 ± 0.07 b | 0.30 ± 0.03 c | 0.01 ± 0.00 a | 0.26 ± 0.02 c |
| 1.03 | Glutamic acid | 246 | 0.01 ± 0.00 a | 0.10 ± 0.01 b | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.01 a |
| 0.96 | Glycine | 102 | 0.05 ± 0.01 b | 0.15 ± 0.03 c | 0.02 ± 0.02 ab | 0.04 ± 0.01 b | N.D. a | 0.05 ± 0.01 b |
| 0.96 | Proline | 142 | 0.20 ± 0.00 c | 0.04 ± 0.03 ab | 0.08 ± 0.07 b | 0.24 ± 0.01 c | N.D. a | 0.25 ± 0.01 c |
| 0.85 | Phenylalanine | 218 | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.11 ± 0.01 b |
| 0.81 | Threonine | 117 | 0.01 ± 0.00 b | 0.03 ± 0.00 c | 0.01 ± 0.00 ab | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.03 ± 0.00 b |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.M.; Jung, J.H.; Seo, J.-A.; Kim, Y.-S. Bioformation of Volatile and Nonvolatile Metabolites by Saccharomycopsis fibuligera KJJ81 Cultivated under Different Conditions—Carbon Sources and Cultivation Times. Molecules 2018, 23, 2762. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23112762
Lee SM, Jung JH, Seo J-A, Kim Y-S. Bioformation of Volatile and Nonvolatile Metabolites by Saccharomycopsis fibuligera KJJ81 Cultivated under Different Conditions—Carbon Sources and Cultivation Times. Molecules. 2018; 23(11):2762. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23112762
Chicago/Turabian StyleLee, Sang Mi, Ji Hye Jung, Jeong-Ah Seo, and Young-Suk Kim. 2018. "Bioformation of Volatile and Nonvolatile Metabolites by Saccharomycopsis fibuligera KJJ81 Cultivated under Different Conditions—Carbon Sources and Cultivation Times" Molecules 23, no. 11: 2762. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23112762