Metabolomic Analysis of Pollen Grains with Different Germination Abilities from Two Clones of Chinese Fir (Cunninghamia lanceolata (Lamb) Hook)

Abstract
:1. Introduction
2. Results
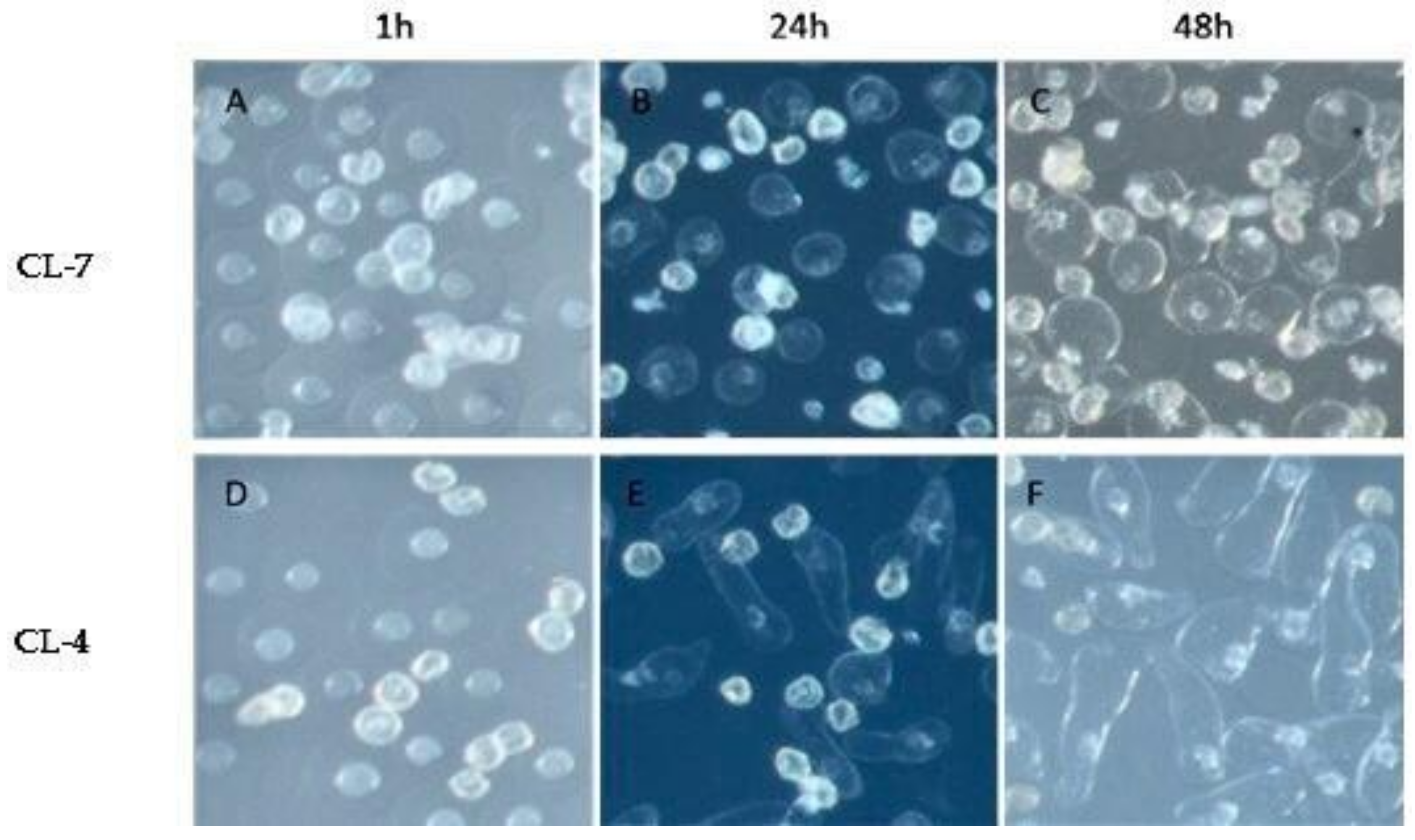
2.1. Pollen Germination Status of the Selected Clones
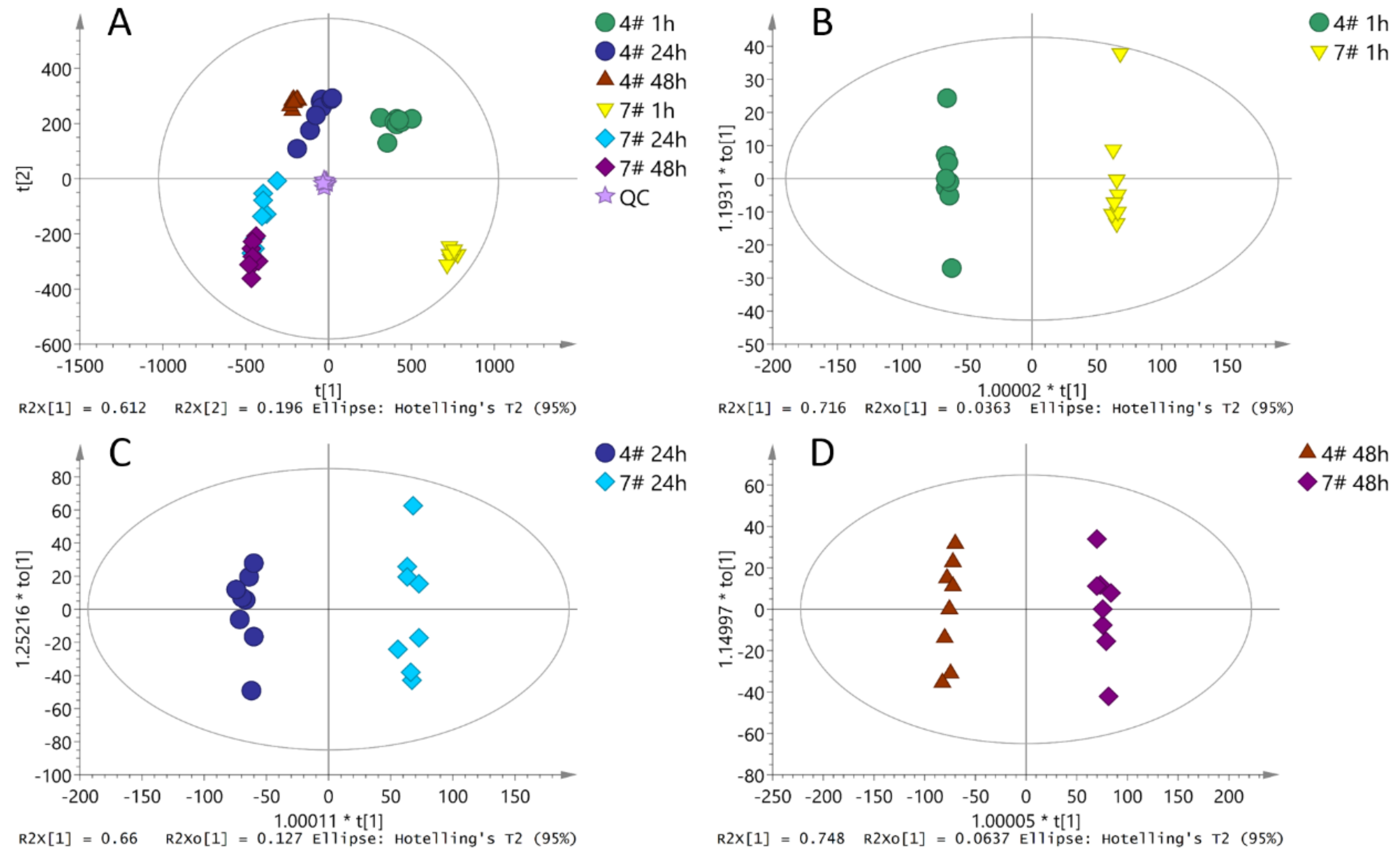
2.2. Metabolites Variations in the Two Clones (CL-4 and CL-7) in Germinating Pollen
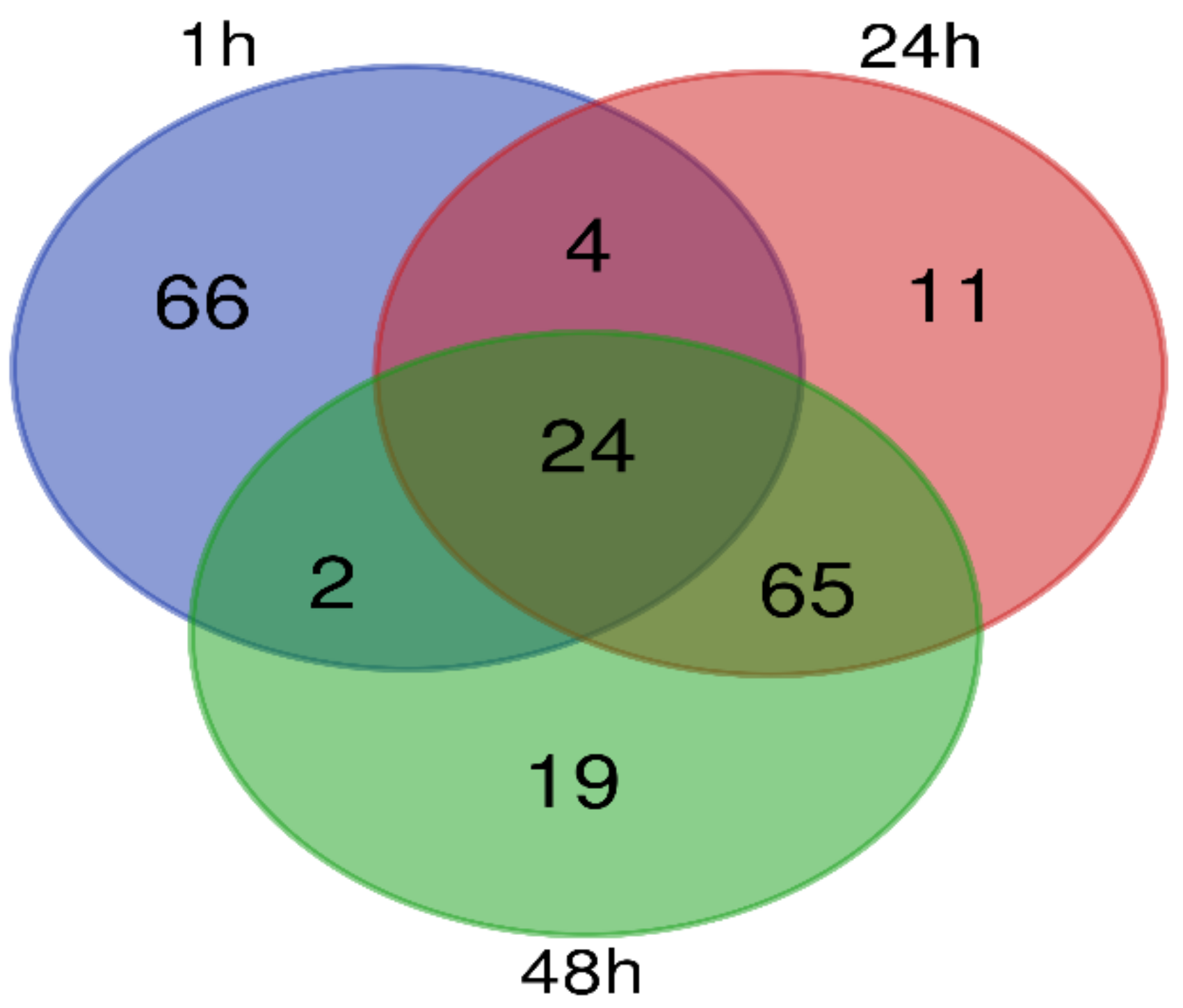
2.3. Differential Metabolites during Pollen Germination
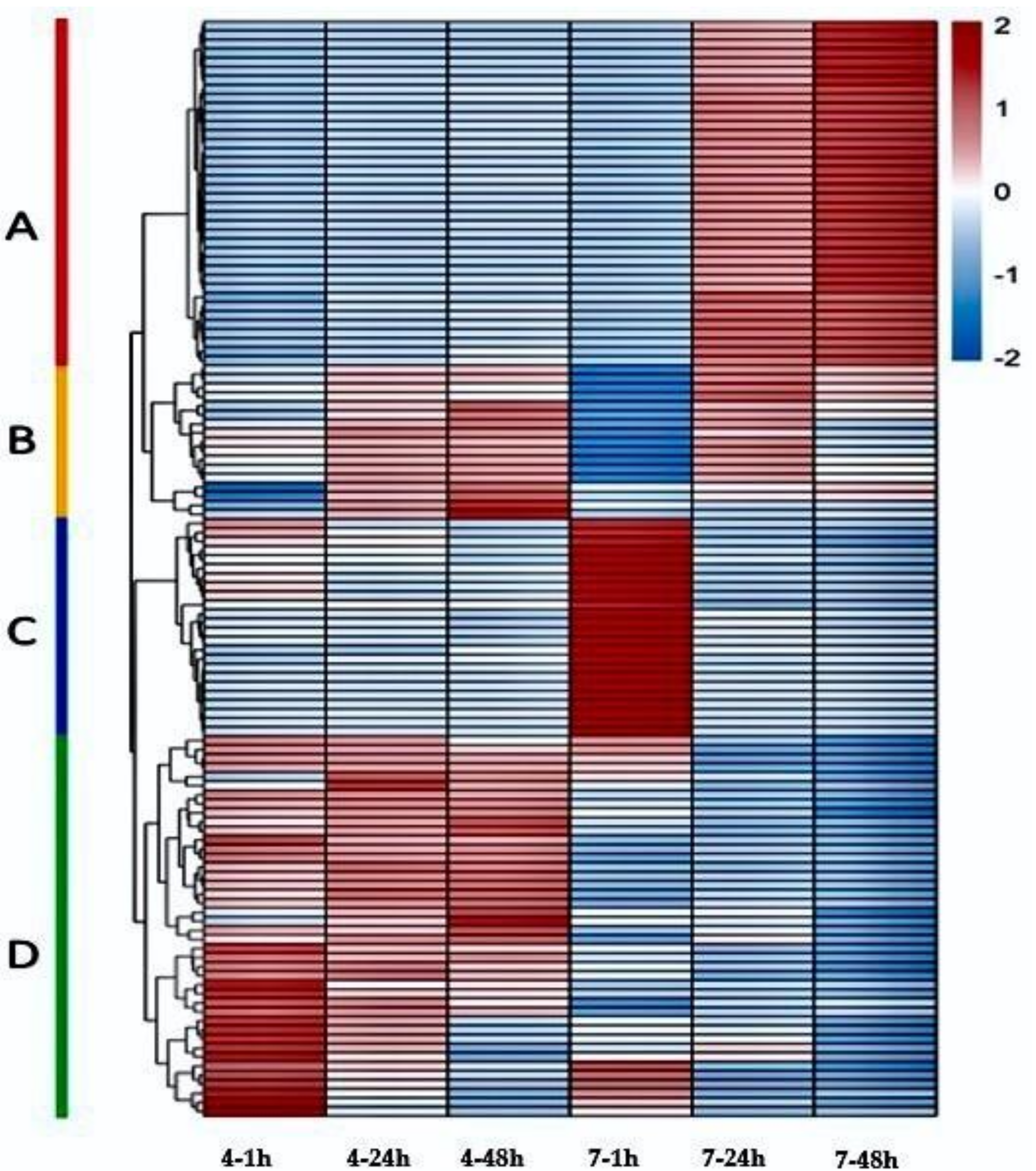
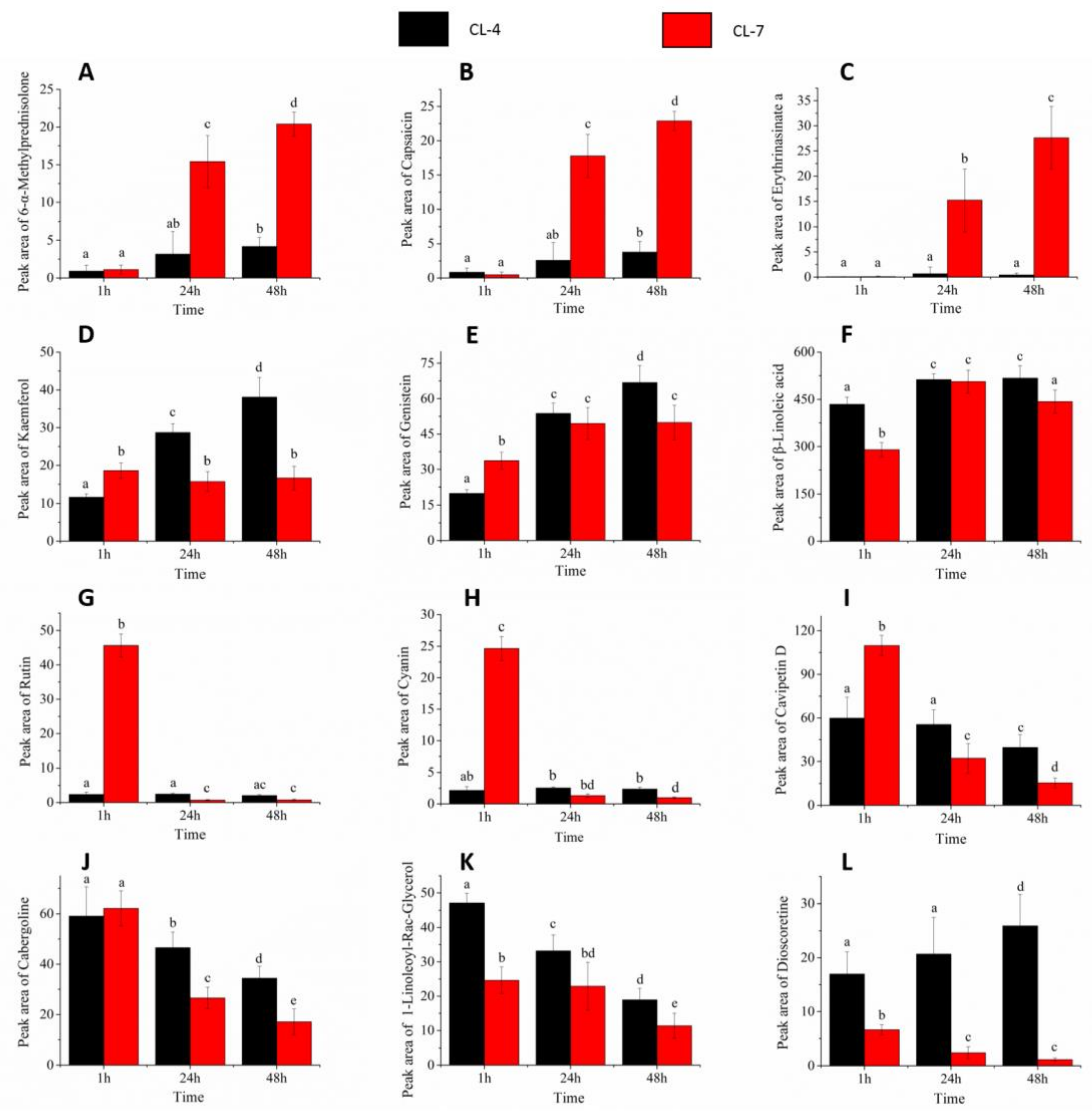
2.4. Metabolites Expression and Changes during Pollen Germination
2.5. Metabolites Content Changes during Pollen Germination
2.6. Enrichment of Differential Metabolic Pathways
3. Discussion
4. Materials and Methods
4.1. Description of the Study Area
4.2. Collection of Pollen Materials
4.3. Preliminary Pollen Germination Testing
4.4. Determination of Pollen Developmental Stages
4.5. Sample Preparation for Metabolites Extraction
4.6. Liquid Chromatography-Mass Spectrometry (LC -MS) Analysis
4.7. Data Processing and Analysis
4.7.1. Identification of Differential Metabolites
4.7.2. Metabolic Pathways Enrichment Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Yang, Z.; Liu, Y.N.; Lv, B. Rapid assessment of surface color of Chinese fir and Eucalyptus wood by near infrared spectroscopy. In Proceedings of Applied Mechanics and Materials; Trans Tech Publications: Beijing, China, 2012; pp. 304–307. [Google Scholar]
- Zheng, H.; Duan, H.; Hu, D.; Li, Y.; Hao, Y. Genotypic variation of Cunninghamia lanceolata revealed by phenotypic traits and SRAP markers. Dendrobiology 2015, 74. [Google Scholar] [CrossRef]
- Shi, H.; Chertow, M.; Song, Y. Developing country experience with eco-industrial parks: A case study of the Tianjin Economic-Technological Development Area in China. J. Clean Prod. 2010, 18, 191–199. [Google Scholar] [CrossRef]
- Zheng, H.; Hu, D.; Wang, R.; Wei, R.; Yan, S. Assessing 62 Chinese Fir (Cunninghamia lanceolata) breeding parents in a 12-year grafted clone test. Forests 2015, 6, 3799–3808. [Google Scholar] [CrossRef]
- Wu, Q.; Fan, R.; Xiao, S.; Wang, Y.; Wu, Y. Study on biology of pollination of Chinese fir. J. Nanj. For. Unive. 1995, 1. [Google Scholar]
- Shen, J.; Zhu, C.C.; Jin, X.U. Effects of mineral elements on pollen germination and pollen tube growth of Cunninghamia lanceolate (Lamb.) Hook. Chin. For. Sci. Technol. 2010, 24, 57–60. [Google Scholar]
- Soares, T.L.; Silva, S.; Costa, M.; Santos-Serejo, J.; Souza, A.d.S.; Lino, L.S.M.; Souza, E.; Jesus, O. In vitro germination and viability of pollen grains of banana diploids. Crop. Breed. Appl. Biot. 2008, 8, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Taylor, L.P.; Hepler, P.K. Pollen germination and tube growth. Annu. Rev. Plant Biol. 1997, 48, 461–491. [Google Scholar] [CrossRef] [PubMed]
- Dane, F.; Olgun, G.; Dalgiç, Ö. In vitro pollen germination of some plant species in basic culture medium. J. Cell. Mol. Biol. 2004, 3. [Google Scholar]
- Trethewey, R.N. Metabolite profiling as an aid to metabolic engineering in plants. Curr. Opin. Plant Biol. 2004, 7, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, J.K.; Wilson, I.D. Understanding ′global′ systems biology: Metabonomics and the continuum of metabolism. Nat. Rev. Drug. Discov. 2003, 2, 668. [Google Scholar] [CrossRef] [PubMed]
- Ott, K.-H.; Araníbar, N.; Singh, B.; Stockton, G.W. Metabonomics classifies pathways affected by bioactive compounds. Artificial neural network classification of NMR spectra of plant extracts. Phytochemistry 2003, 62, 971–985. [Google Scholar] [CrossRef]
- Sumner, L.W.; Mendes, P.; Dixon, R.A. Plant Metabolomics: Large-Scale Phytochemistry in the Functional Genomics Era. Phytochemistry 2003, 62, 817–836. [Google Scholar] [CrossRef]
- Weckwerth, W. Metabolomics in systems biology. Annu. Rev. Plant Biol. 2003, 54, 669–689. [Google Scholar] [CrossRef] [PubMed]
- Bjarnholt, N.; Li, B.; D’Alvise, J.; Janfelt, C. Mass spectrometry imaging of plant metabolites principles and possibilities. Nat. Prod. Rep. 2014, 31, 818–837. [Google Scholar] [CrossRef] [PubMed]
- Menino, M.R.; Carranca, C.; Varennes, A.D.E.; D’Almeida, V.V.; Baeta, J. Tree size and flowering intensity as affected by nitrogen fertilization in non-bearing orange trees grown under Mediterranean conditions. J. Plant Physiol. 2003, 160, 1435–1440. [Google Scholar] [CrossRef] [PubMed]
- Li, R.Z.; Lin, J.; Wang, X.X.; Yu, X.M.; Chen, C.L.; Guan, Y.F. Nontargeted metabolomic analysis of Anoectochilus roxburghii at different cultivation stages. Chin. J. Chin. Mater. Medica. 2017, 42, 4624. [Google Scholar]
- Dethloff, F.; Erban, A.; Orf, I.; Alpers, J.; Fehrle, I.; Beinegolovchuk, O.; Schmidt, S.; Schwachtje, J.; Kopka, J. Profiling methods to identify cold-regulated primary metabolites using gas chromatography coupled to mass spectrometry. Methods Mol. Biol. 2014, 1166, 171–197. [Google Scholar] [PubMed]
- Keurentjes, J.J.B.; Fu, J.; Vos, C.H.R.D.; Lommen, A.; Hall, R.D.; Bino, R.J.; Linus., H.W. The genetics of plant metabolism. Nat. Genet. 2006, 38, 842–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tikunov, Y.; Lommen, A.; Vos, C.H.R.D.; Verhoeven, H.A.; Bino, R.J.; Hall, R.D.; Bovy, A.G. A novel approach for nontargeted data analysis for metabolomics. Large-Scale profiling of tomato fruit volatiles. Plant Physiol. 2005, 139, 1125–1137. [Google Scholar] [CrossRef] [PubMed]
- Paupière, M.J.; Müller, F.; Li, H.; Rieu, I.; Tikunov, Y.M.; Visser, R.G.F.; Bovy, A.G. Untargeted metabolomic analysis of tomato pollen development and heat stress response. Plant Reprod. 2017, 30, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Wahyuni, Y.; Ballester, A.R.; Tikunov, Y.; Vos, R.C.H.D.; Pelgrom, K.T.B.; Maharijaya, A.; Sudarmonowati, E.; Bino, R.J.; Bovy, A.G. Metabolomics and molecular marker analysis to explore pepper (Capsicum sp.) biodiversity. Metabolomics 2013, 9, 130–144. [Google Scholar] [CrossRef] [PubMed]
- Castro, A.; Clement, C. Sucrose and starch catabolism in the anther of Lilium during its development: A comparative study among the anther wall, locular fluid and microspore/pollen fractions. Planta 2007, 225, 1573–1582. [Google Scholar] [CrossRef] [PubMed]
- Sangwan, R.S. Change in the amino-acid content during male gametophyte formation of Datura metel in Situ. Tag. Theoret. Appl. Genet. 1978, 52, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Zhi, Z.; Zhu, Z.; Shen, R.; Tang, X. Effect of artificial pollination on germination, forms and biomacromolecule of Chinese fir seed. J. Zhejiang. For. Sci. Technol. 1996, 16, 15–17. [Google Scholar]
- Lu, Y.; Jin, B.; Wang, L.; Wang, Y.; Wang, D.; Jiang, X.-X.; Chen, P. Adaptation of male reproductive structures to wind pollination in gymnosperms: Cones and pollen grains. Can. J. Plant Sci. 2011, 91, 897–906. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, J.; Xiao, F.; Li, Y.; Zhu, X.; Xu, H.; Huang, B. Study on pollen germination and preservation characteristic of Cunninghamia lanceolata. Jiangxi For. Sci. Technol. 2012, 4, 1. [Google Scholar]
- Guofeng, G.; Ye, L.; Jin, X. Microtubule organization in pollen and pollen tube of Cunninghamia lanceolata. Sci. Silv. Sinic. 2012. [Google Scholar]
- Lu, Y.; Coops, N.C.; Wang, T.; Wang, G. A process-based approach to estimate Chinese fir (Cunninghamia lanceolata) distribution and productivity in southern China under climate change. Forests 2015, 6, 360–379. [Google Scholar] [CrossRef]
- Xu, C. Study of factors affecting on physical germination of the Apple pollen. Chin. Agric. Sci. Bull. 2011, 27, 163–167. [Google Scholar]
- Portnoi, L.; Horovitz, A. Sugars in natural and artificial pollen germination substrates. Ann. Bot. 1977, 41, 21–27. [Google Scholar] [CrossRef]
- Dearnaley, J.D.; Levina, N.N.; Lew, R.R.; Heath, I.B.; Goring, D.R. Interrelationships between cytoplasmic Ca2+ peaks, pollen hydration and plasma membrane conductances during compatible and incompatible pollinations of Brassica napus papillae. Plant. Cell. Physiol. 1997, 38, 985–999. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.; Peer, W.A.; Taiz, L. Regulation of auxin transport by aminopeptidases and endogenous flavonoids. Planta 2000, 211, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.X.; Wei, X.Y.; Feng, S.X.; Alamus; Hu, T.M.; Wang, Y.Q. A study on the flavonoids of Lespedeza hedysaroides. Acta Bot. Boreali-Occidentalia Sinica 2009, 29, 1904–1908. [Google Scholar]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.W.; Yang, L.Y.; Chen, J.L.; Zhang, Y.; Wang, Z.L. Progress of flavonoids constituents on anti-free radical oxidative activity. Chi. J. Trad. Chin. Med. Pharm. 2013, 28, 2694–2697. [Google Scholar]
- Tu, W.; Geng, Y.; Zhang, J.; Zhang, L.; Zhang, Y. Analysis of fatty acid composition of eight kinds of pollens. Acta Nutrimenta Sin. 2005, 27, 351–352. [Google Scholar]
- Zhang, Q.; Chen, Y.; Huang, Q.; University, S.A. Research advances in the defense signaling pathway and transcription factors mediated by plant unsaturated fatty acid derivatives under abiotic stress. Mol. Plant Breeding. 2018, 16, 1494–1502. [Google Scholar]
- Bolat, I.; Pirlak, L. Effects of three plant growth regulators and boric acid on pollen germination and tube growth in apricot (Prunus armeniaca L.). Bangladesh. J. Bot. 2003, 32, 53–56. [Google Scholar]
- Radović, A.; Nikolić, D.; Milatović, D.; Živković, B.; Stevanović, N. The effect of plant hormones on pollen germination and pollen tube growth of almond cultivars. Acta. Hortic. 2016, 1139, 375–380. [Google Scholar] [CrossRef]
- Carlos, S.; Jorge, C.; Nicolas, V.; Roman, T. Influence of Seven Growth Regulators on Fruit Set, Pollen Germination and Pollen Tube Growth of Almonds. J. Agric. Sci. Technol. B 2012, 2, 1051–1056. [Google Scholar]
- Parish, R.W.; Phan, H.A.; Iacuone, S.; Li, S.F. Tapetal development and abiotic stress: A centre of vulnerability. Funct. PLant Biol. 2012, 39, 553–559. [Google Scholar] [CrossRef]
- Chun-Ling, H.E.; Zhang, S.X.; Bao-Zhong, J.I. Detection and function activity of 10- Hydroxy- 2- decenoic acid in the bee bread of Xylocopa tranquebarorum. J. Environ. Entomol. 2014, 36, 176–181. [Google Scholar]
- Edlund, A.F.; Swanson, R.; Preuss, D. Pollen and stigma structure and function: The role of diversity in pollination. Plant Cell 2004, 16, S84–S97. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-García, M.I.; M’Rani-Alaoui, M.; Fernández, M.C. Behavior of storage lipids during development and germination of olive (Olea europaea L.) pollen. Protoplasma 2003, 221, 237–244. [Google Scholar] [PubMed]
- Palevitch, D.; Craker, L.E. Nutritional and medical importance of red pepper (Capsicum spp.). J. Herb Spices. Medici. Plant. 1996, 3, 55–83. [Google Scholar] [CrossRef]
- Chen, J.Q.; He, L.L.; Ge, X.G.; Zhao, R.; Fu, H.D. Biosynthesis and metabolism of capsaicin in plant. Plant. Physiol. Commun. 2008, 44, 1013–1017. [Google Scholar]
- Gao, J.; Guo, Z. Recent progress in the synthesis of GPIs and GPI-anchored proteins. Sci. Sinica. Chimica. 2013, 43, 964–983. [Google Scholar] [CrossRef]
- Kinoshita, T.; Fujita, M. Thematic review series: Glycosylphosphatidylinositol (GPI) anchors: Biochemistry and cell biology: Biosynthesis of GPI-anchored proteins: Special emphasis on GPI lipid remodeling. J. Lipid. Res. 2016, 57, 6. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, J.; Schmidt, S.; Chutia, R.; Müller, J.; Böttcher, C.; Strehmel, N.; Scheel, D.; Abel, S. Non-targeted profiling of semi-polar metabolites in Arabidopsis root exudates uncovers a role for coumarin secretion and lignification during the local response to phosphate limitation. J. Exp. Bot. 2016, 67, 1421–1432. [Google Scholar] [CrossRef] [PubMed]
- Yuni, W.; Vanessa, S.H.; Ana-Rosa, B.; De, V.R.C.H.; Voorrips, R.E.; Awang, M.; Jos, M.; Viquez, Z.M.; Enny, S.; Maisonnave, A.A.C. Genetic mapping of semi-polar metabolites in pepper fruits (Capsicum sp.): Towards unravelling the molecular regulation of flavonoid quantitative trait loci. Mol. Breeding 2014, 33, 503. [Google Scholar]
- Want, E.J.; Wilson, I.D.; Gika, H.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Holmes, E.; Nicholson, J.K. Global metabolic profiling procedures for urine using UPLC-MS. Nat. Protoc. 2010, 5, 1005–1018. [Google Scholar] [CrossRef] [PubMed]
- Li, C.F.; Yao, M.Z.; Ma, C.L.; Ma, J.Q.; Jin, J.Q.; Chen, L. Differential Metabolic Profiles during the Albescent Stages of ‘Anji Baicha’ (Camellia sinensis). PLoS ONE 2015, 10, e0139996. [Google Scholar] [CrossRef] [PubMed]
- Commisso, M.; Toffali, K.; Strazzer, P.; Stocchero, M.; Ceoldo, S.; Baldan, B.; Levi, M.; Guzzo, F. Impact of Phenylpropanoid Compounds on Heat Stress Tolerance in Carrot Cell Cultures. Front. Plant Sci. 2016, 7, 1439. [Google Scholar] [CrossRef] [PubMed]
- Paupière, M.J.; Heusden, A.W.V.; Bovy, A.G. The Metabolic Basis of Pollen Thermo-Tolerance: Perspectives for Breeding. Metabolites 2014, 4, 889–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Samples of all extracted metabolites are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pollen Germination (%) | Pollen Tube Growth (µm) | |||||
|---|---|---|---|---|---|---|
| Clones | 1 h | 24 h | 48 h | 1 h | 24 h | 48 h |
| CL-4 | 82.7 | 87.02 | 78.01 | 0.28 | 0.42 | 0.93 |
| CL-7 | 50.53 | 51.99 | 50.53 | 0.27 | 0.35 | 0.51 |
| Category | Organic Acids | Fatty Acids | Hormones | Vitamins | Flavonoids | Glycosides | Lipids | Amino Acids & Peptides | Amines |
|---|---|---|---|---|---|---|---|---|---|
| A | 7 | 9 | 8 | 7 | 2 | 1 | 2 | 0 | 2 |
| B | 3 | 8 | 0 | 0 | 3 | 0 | 3 | 0 | 0 |
| C | 1 | 2 | 0 | 2 | 10 | 4 | 4 | 1 | 1 |
| D | 5 | 11 | 1 | 1 | 5 | 3 | 10 | 0 | 5 |
| Time (min) | A (%) | B (%) |
|---|---|---|
| 0 | 95 | 5 |
| 2 | 80 | 20 |
| 4 | 75 | 25 |
| 9 | 40 | 60 |
| 14 | 0 | 100 |
| 18 | 0 | 100 |
| 18.1 | 95 | 5 |
| 19.5 | 95 | 5 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fragallah, S.A.D.A.; Wang, P.; Li, N.; Chen, Y.; Lin, S. Metabolomic Analysis of Pollen Grains with Different Germination Abilities from Two Clones of Chinese Fir (Cunninghamia lanceolata (Lamb) Hook). Molecules 2018, 23, 3162. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23123162
Fragallah SADA, Wang P, Li N, Chen Y, Lin S. Metabolomic Analysis of Pollen Grains with Different Germination Abilities from Two Clones of Chinese Fir (Cunninghamia lanceolata (Lamb) Hook). Molecules. 2018; 23(12):3162. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23123162
Chicago/Turabian StyleFragallah, Seif Aldin Dawina Abdallah, Pei Wang, Nuo Li, Yu Chen, and Sizu Lin. 2018. "Metabolomic Analysis of Pollen Grains with Different Germination Abilities from Two Clones of Chinese Fir (Cunninghamia lanceolata (Lamb) Hook)" Molecules 23, no. 12: 3162. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23123162