Effects of Rich-Polyphenols Extract of Dendrobium loddigesii on Anti-Diabetic, Anti-Inflammatory, Anti-Oxidant, and Gut Microbiota Modulation in db/db Mice

Abstract
:1. Introduction
2. Results
2.1. Main Compounds in DJP
2.2. Anti-Diabetic Effects of DJP
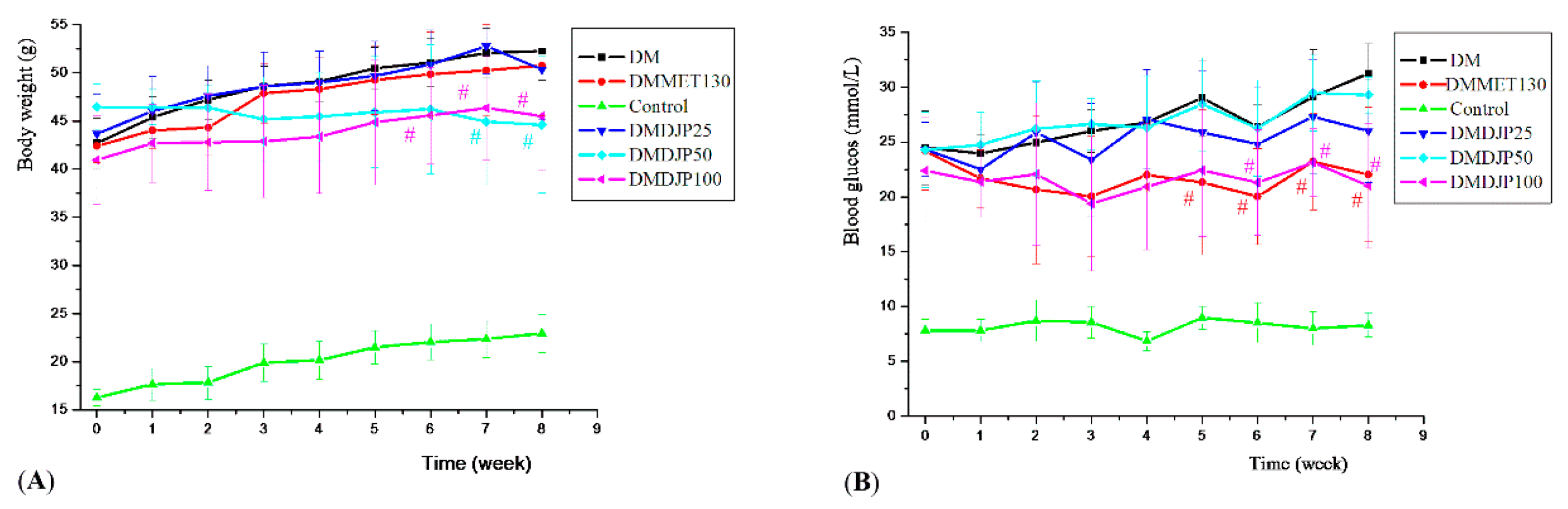
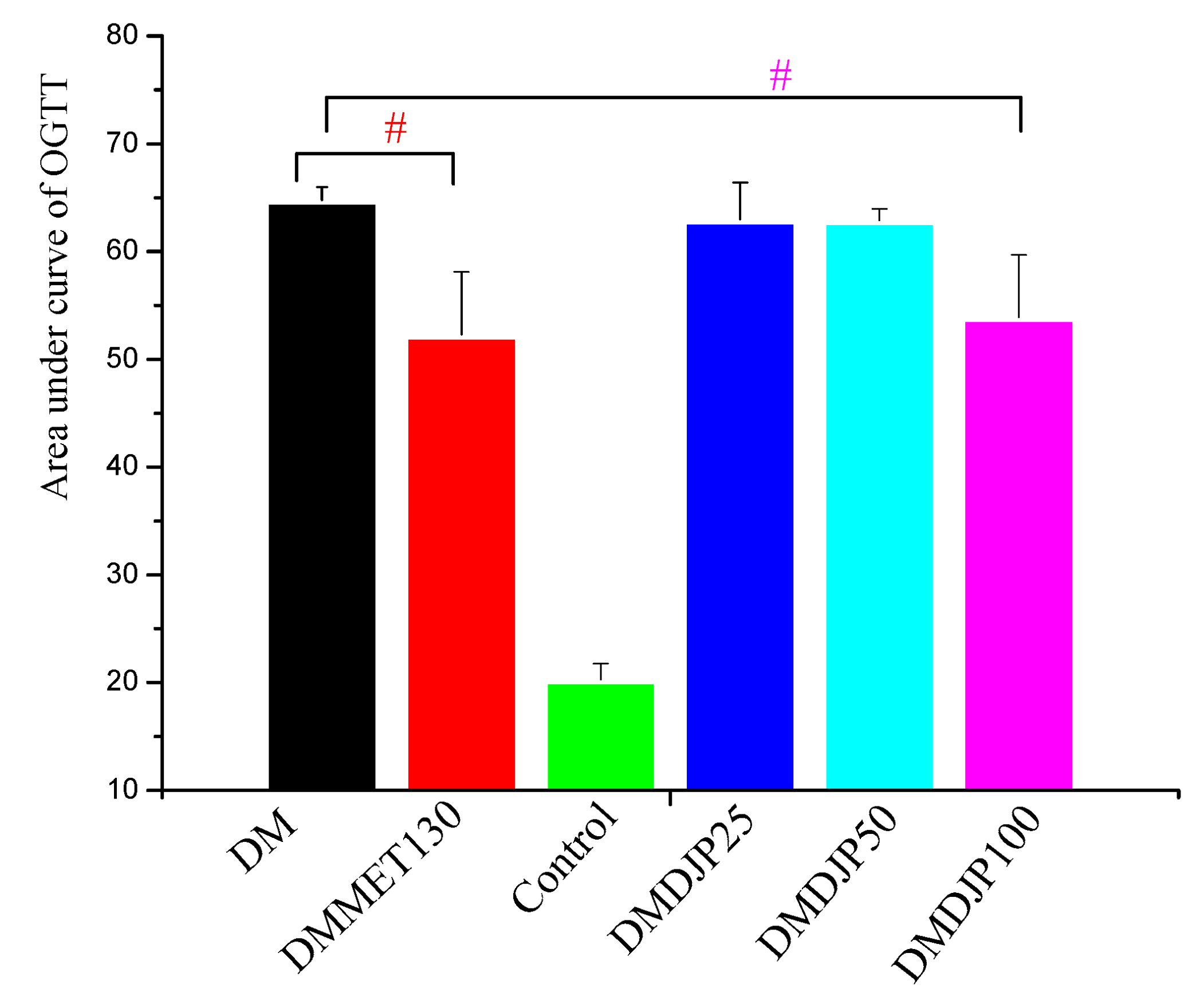
2.2.1. Effects of DJP on the Body Weight, Blood Glucose Level, and Oral Glucose Tolerance (OGTT)
2.2.2. Effect of DJP on Serum Insulin and Lipid Levels
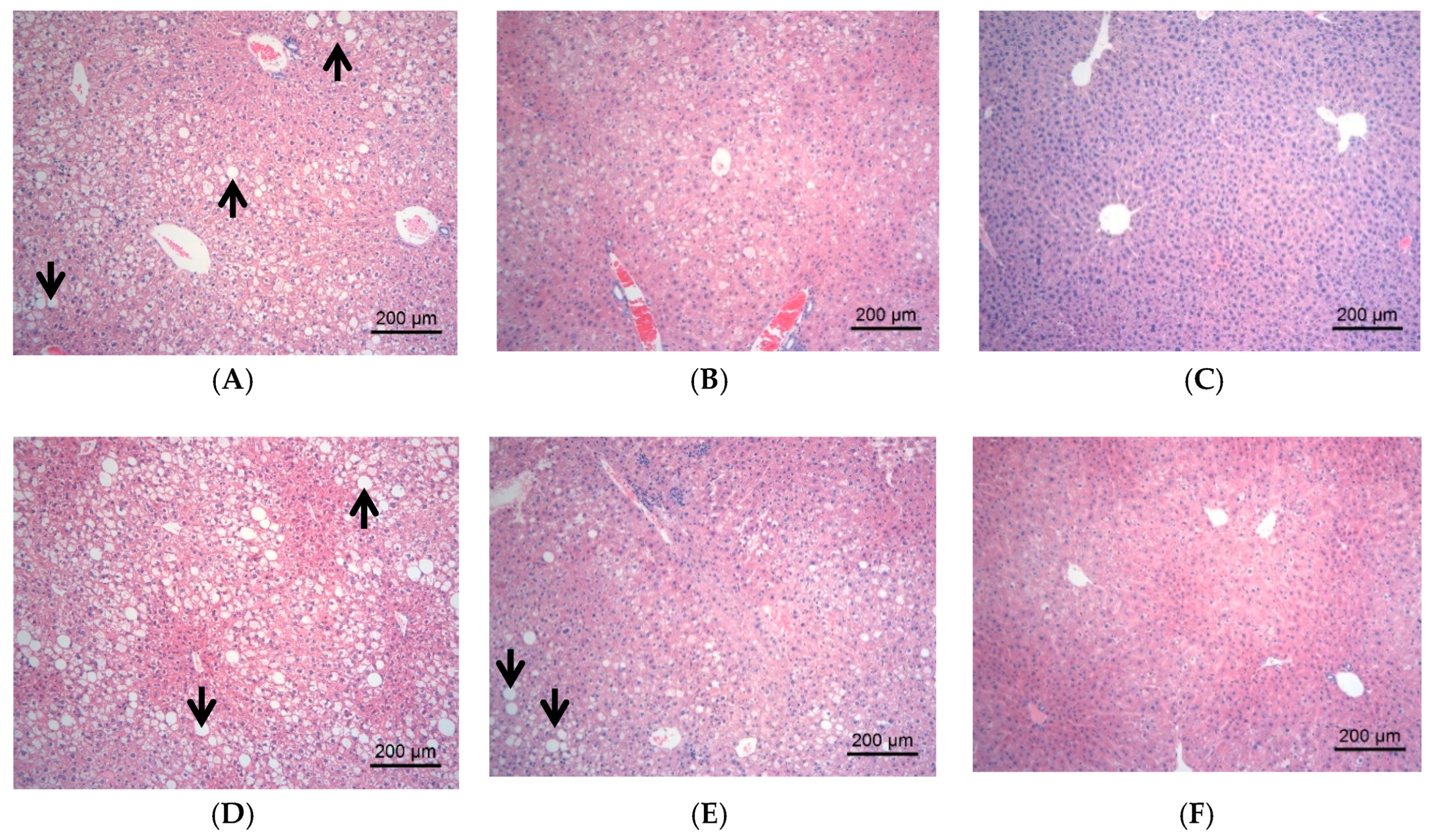
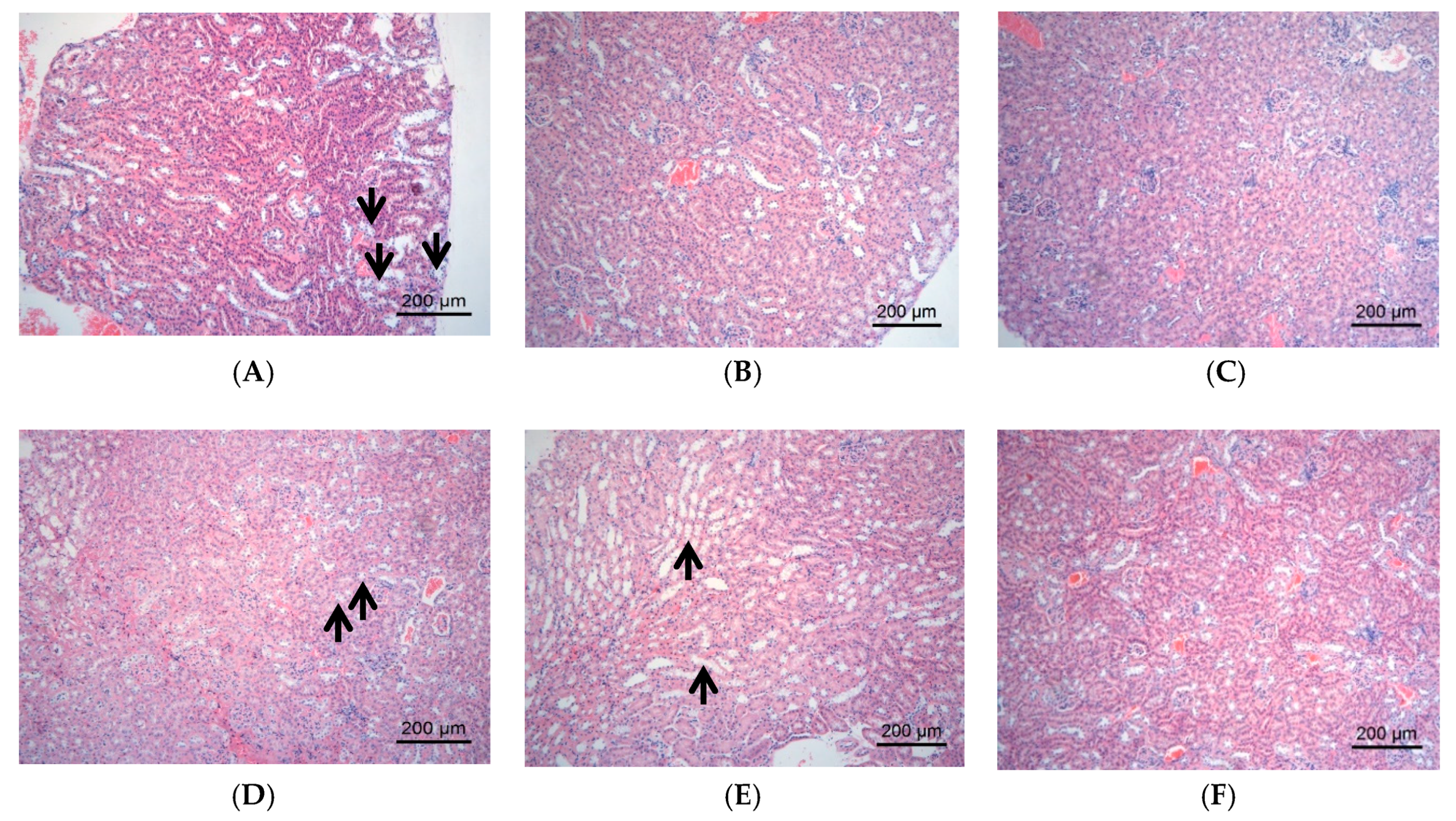
2.2.3. Effect of DJP on Tissue Forms of Liver/Kidney
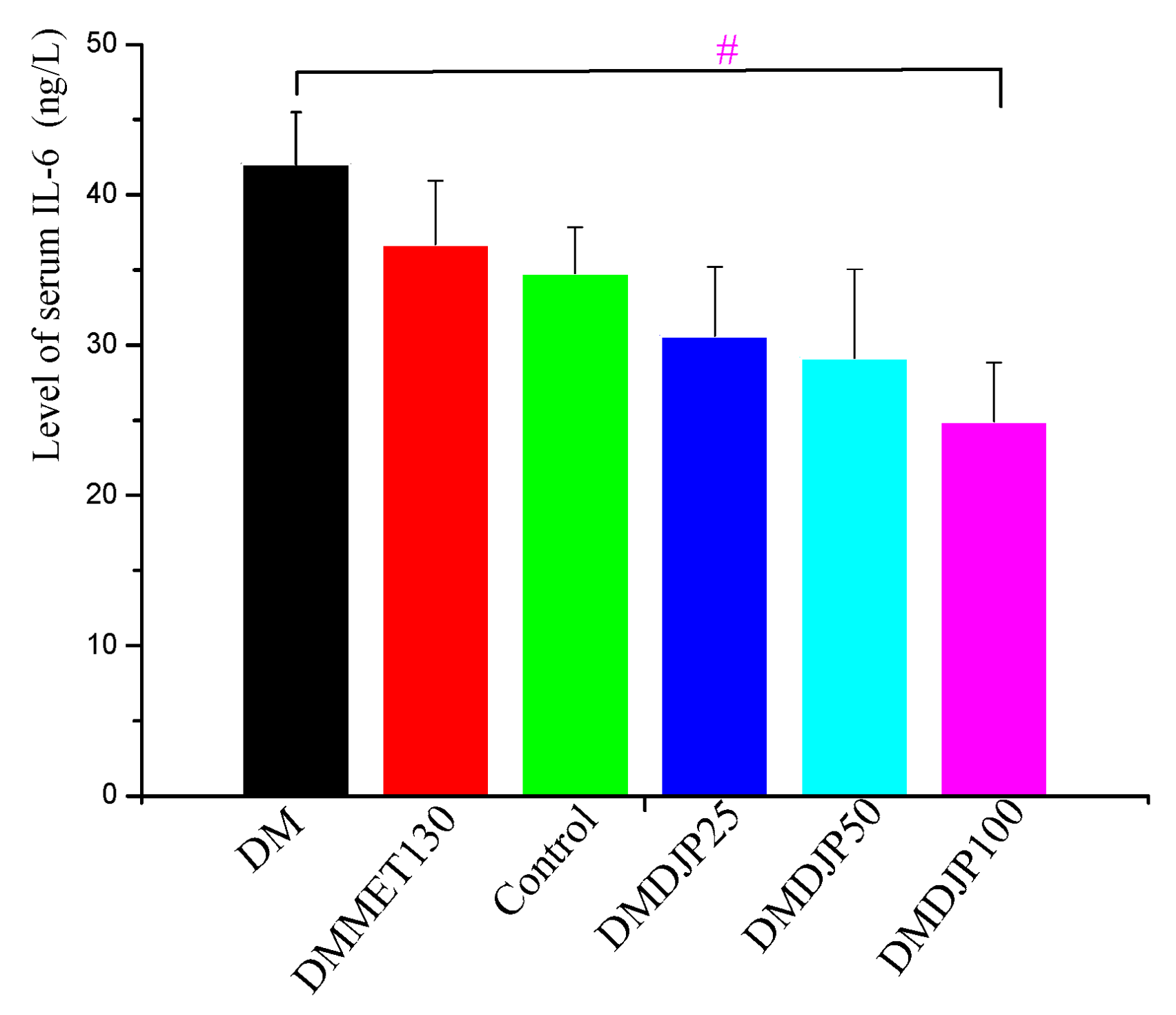
2.3. Effect of DJP on Inflammatory Factors
2.4. Effect of DJP on Oxidative Stress Index
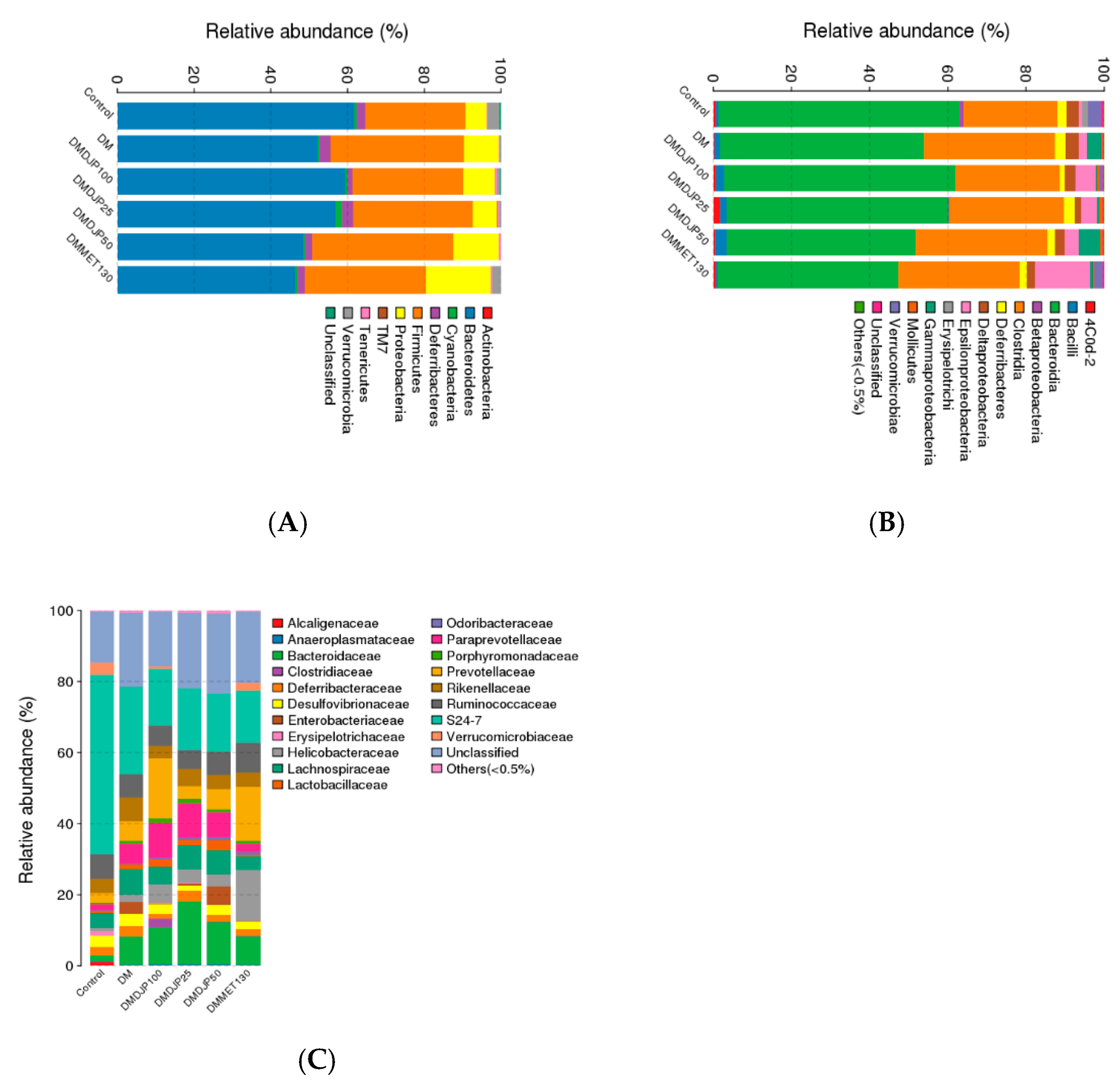
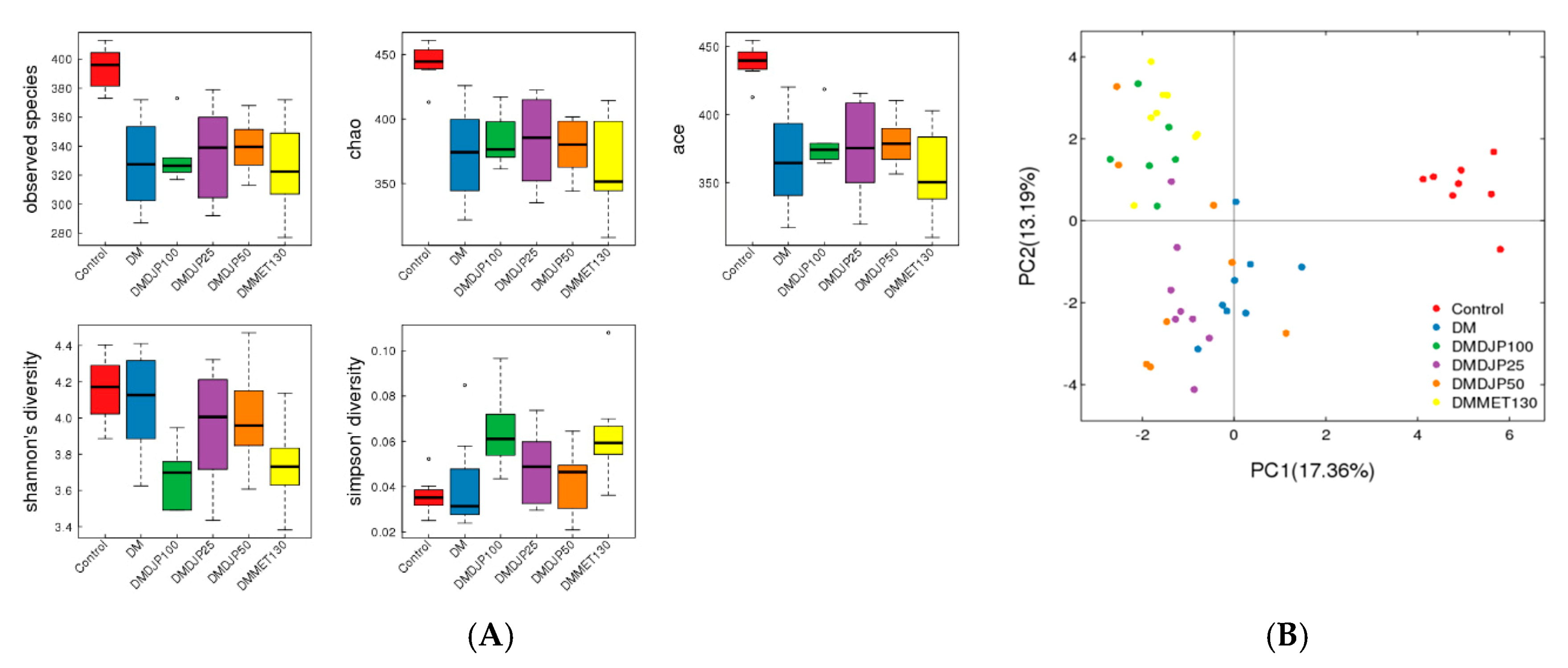
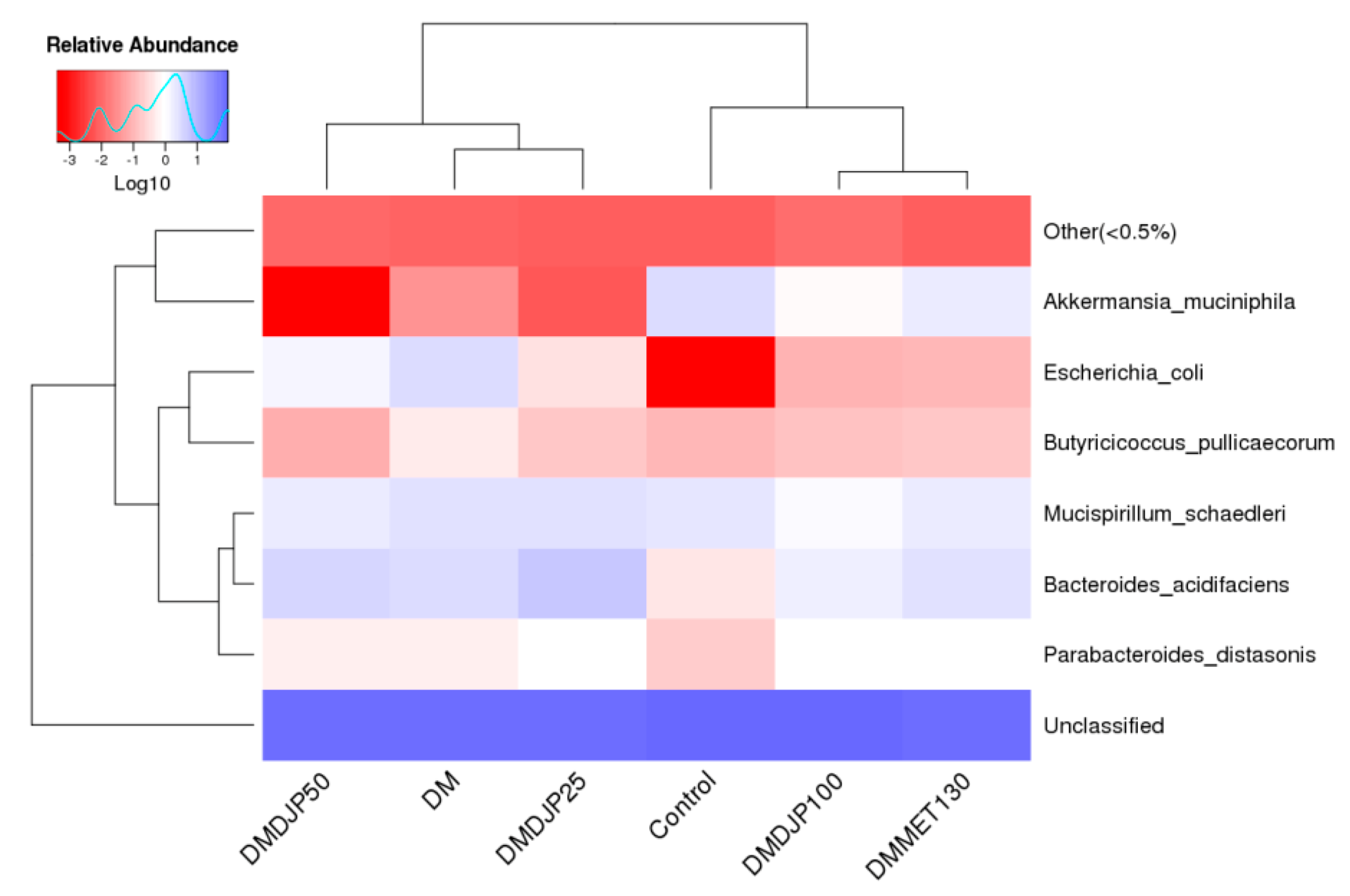
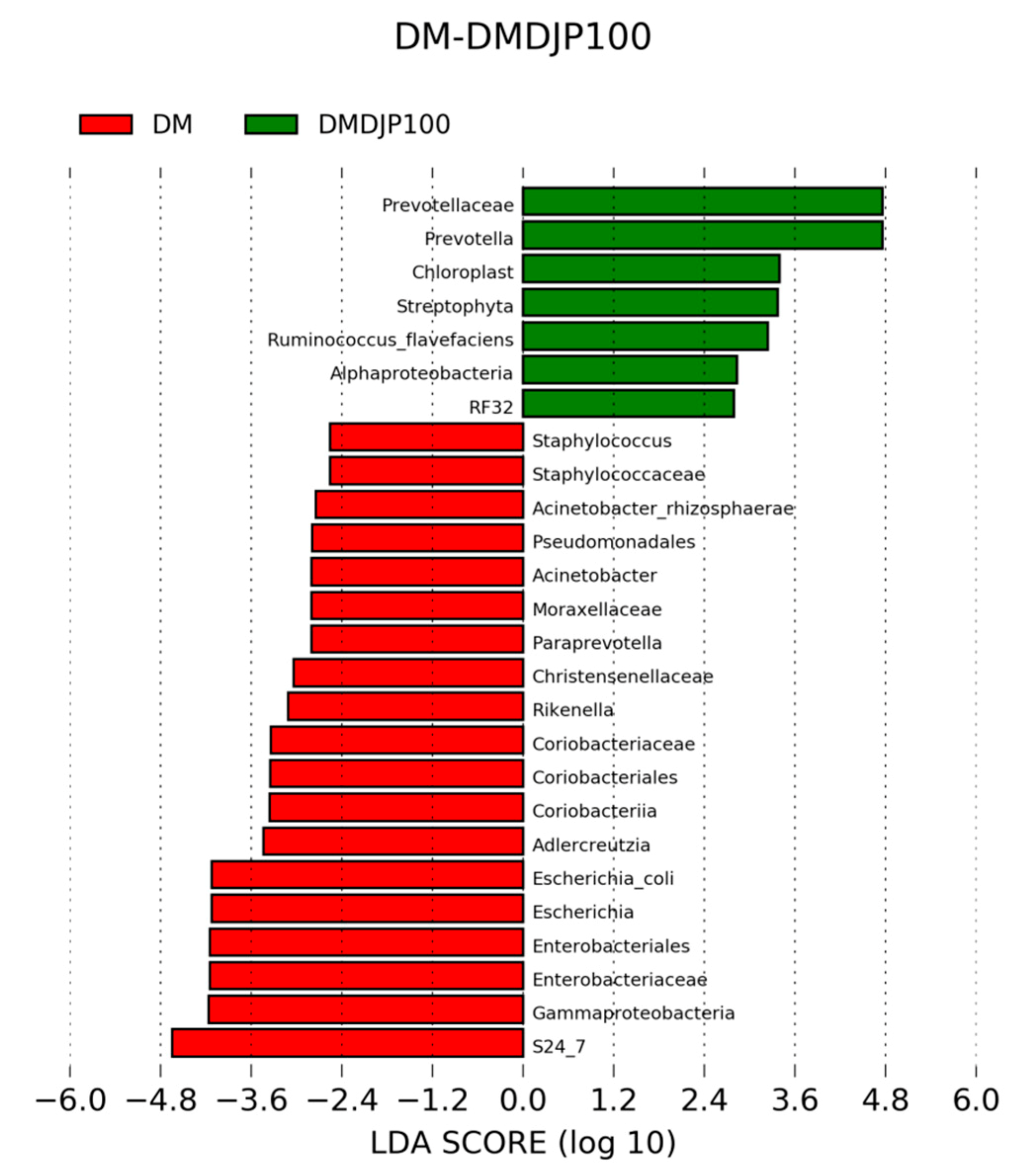
2.5. DJP on Modulation of Intestinal Microbiome
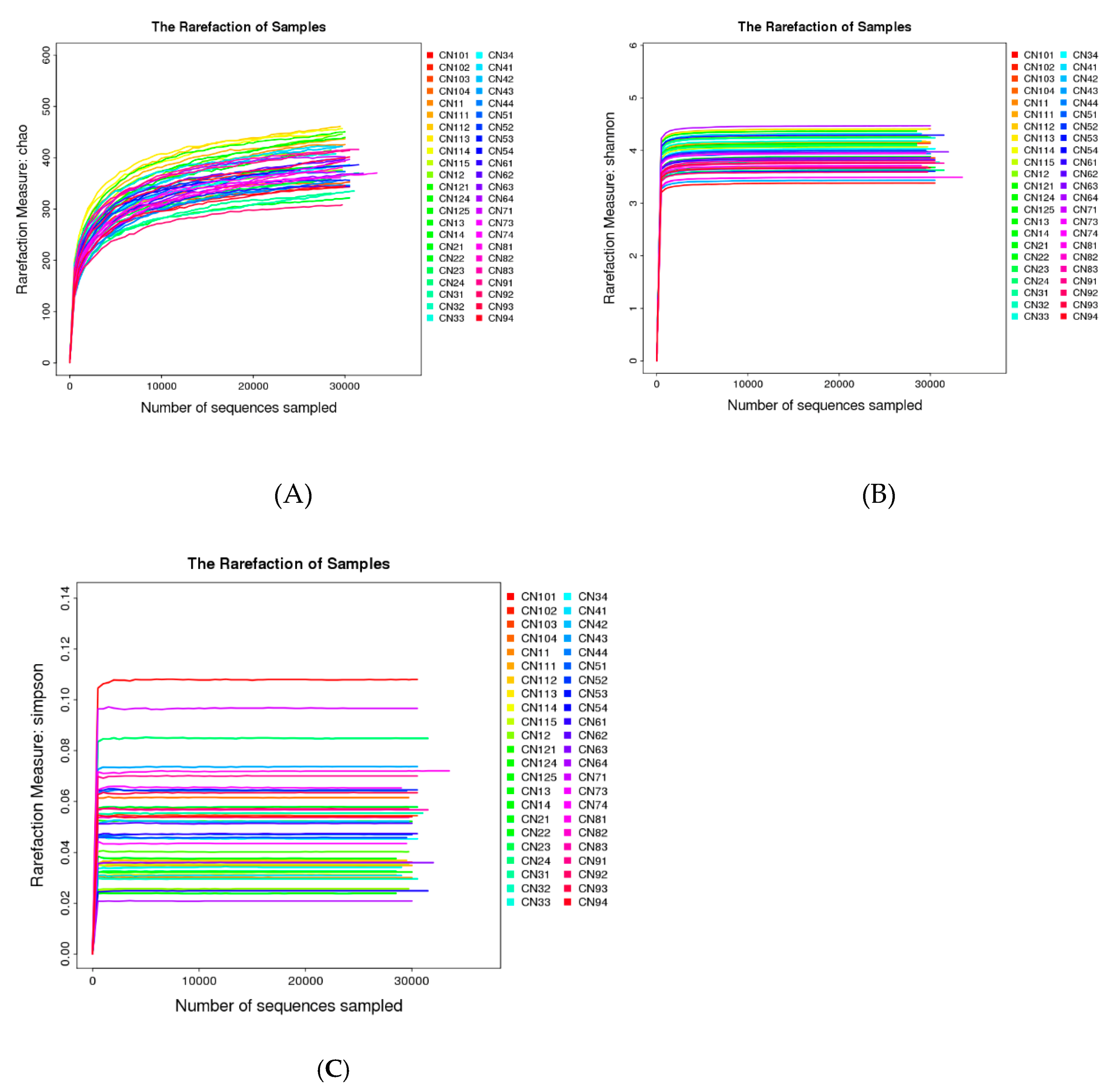
2.5.1. Taxonomic Characterization of Intestinal Microbiome
2.5.2. Distinct Microbial Composition
2.5.3. Functional Predictions of Gut Microbiome
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Preparation Rich-Polyphenols Extract of D. Loddigesii
4.3. Animal Experiments
4.4. Blood Glucose and ELISA Analysis
4.5. Histologic Analysis
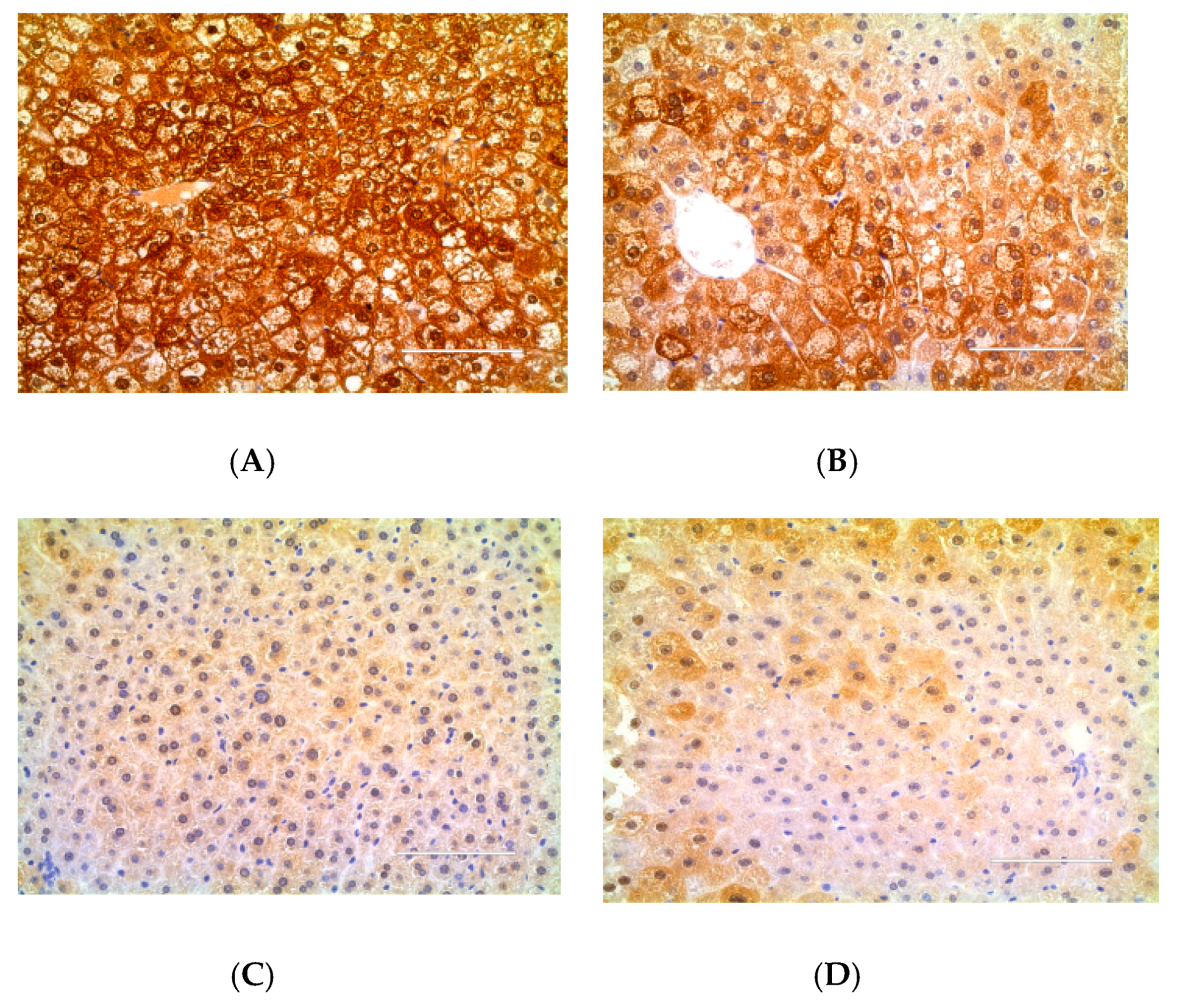
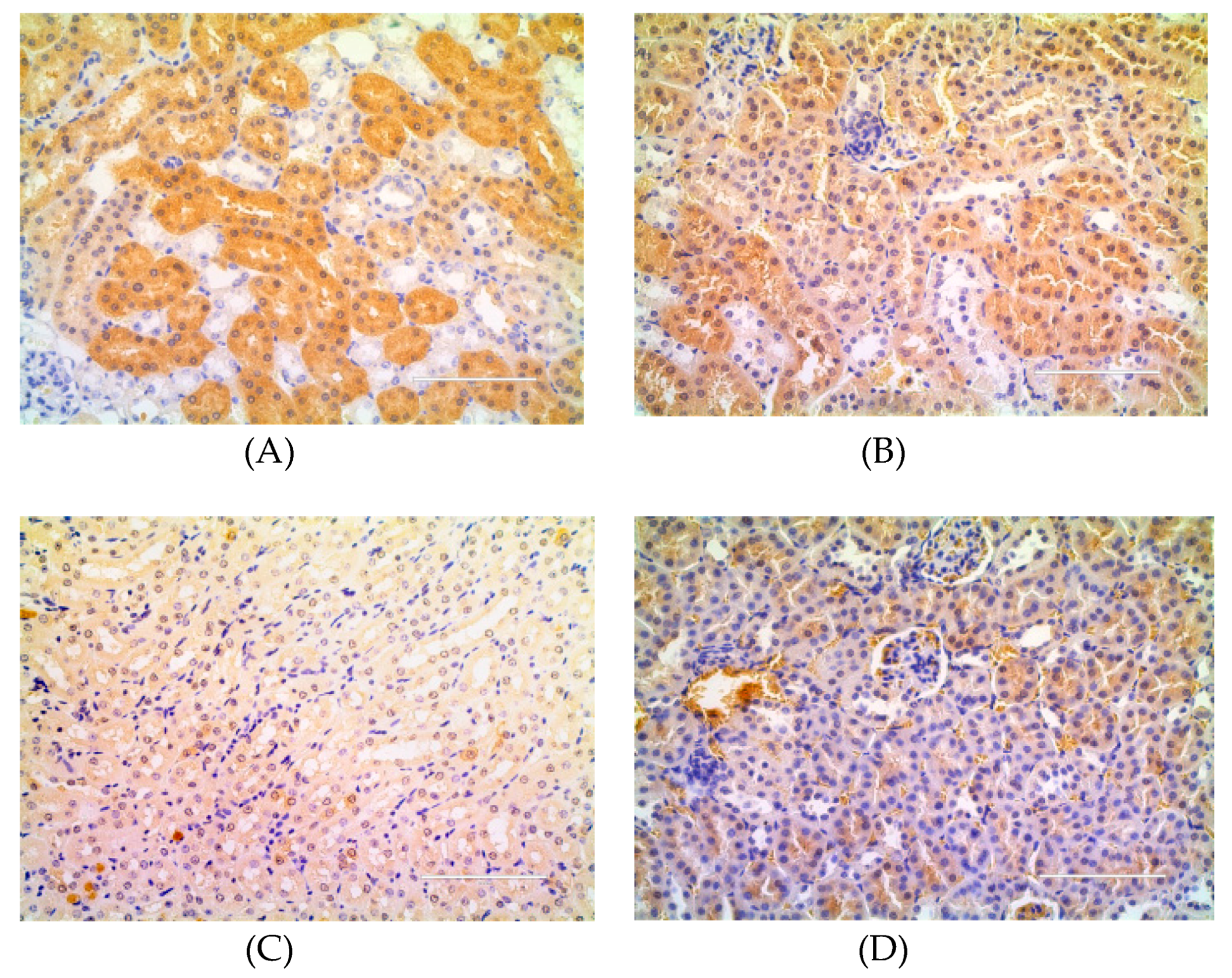
4.6. Immunohistochemical Analysis of TNF-α
4.7. 16S rRNA Gene Extraction, Amplification and Sequencing
4.8. Statistical and Bioinformatics Analysis
4.9. Predicted Metabolic Profile
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| C57 control | no-treatment C57 mice |
| CAT | catalase |
| DJP | rich-polyphenols extract of D. loddigesii |
| DM | no-treatment diabetes db/db mice |
| DMDJP100 | DJP-treatment diabetes db/db mice, in a dose of 100 mg/kg |
| DMDJP25 | DJP-treatment diabetes db/db mice, in a dose of 25 mg/kg |
| DMDJP50 | DJP-treatment diabetes db/db mice, in a dose of 50 mg/kg |
| DMMET130 | MET-treatment diabetes db/db mice, in a dose of 130 mg/kg |
| GSH | glutathione |
| HDL-C | high density lipoprotein cholesterol |
| IL-6 | interleukin-6 |
| INS | insulin |
| LDL-C | low density lipoprotein cholesterol |
| MDA | malondialdehyde |
| MET | metformin |
| OGTT | oral glucose tolerance |
| OTUs | operational taxonomic units |
| PCA | principal component analysis |
| SCFA | short chain fatty acid |
| SOD | superoxide dismutase |
| T2DM | Type 2 diabetes mellitus |
| TC | total cholesterol |
| TG | triglyceride |
| TNF-a | tumor necrosis factor-a |
References
- Ogurtsova, K.; da Rocha Fernandes, J.D.; Huang, Y.; Linnenkamp, U.; Guariguata, L.; Cho, N.H.; Cavan, D.; Shaw, J.E.; Makaroff, L.E. IDF Diabetes Atlas: Global estimates for the prevalence of diabetes for 2015 and 2040. Diabetes Res. Clin. Pract. 2017, 128, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Palem, S.P.; Abraham, P. A Study on the level of oxidative stress and inflammatory markers in type 2 diabetes mellitus patients with different treatment modalities. J. Clin. Diagn. Res. 2015, 9, BC04-7. [Google Scholar] [CrossRef]
- Brownlee, M. The pathobiology of diabetic complications: A unifying mechanism. Diabetes 2005, 54, 1615–1625. [Google Scholar] [CrossRef] [PubMed]
- Salabei, J.K.; Lorkiewicz, P.K.; Mehra, P.; Gibb, A.A.; Haberzettl, P.; Hong, K.U.; Wei, X.; Zhang, X.; Li, Q.; Wysoczynski, M.; et al. Type 2 diabetes dysregulates glucose metabolism in cardiac progenitor cells. J. Biol. Chem. 2016, 291, 13634–13648. [Google Scholar] [CrossRef] [PubMed]
- Coomer, M.; Essop, M.F. Differential hexosamine biosynthetic pathway gene expression with type 2 diabetes. Mol. Genet. Metab. Rep. 2014, 1, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, X.C.; Zhao, L.; Zhang, Y.; Yao, L.L.; Lin, Y.; Peng, Y.D.; Hu, R.M. Oligonucleotide microarray analysis reveals dysregulation of energy-related metabolism in insulin-sensitive tissues of type 2 diabetes patients. Genet. Mol. Res. 2014, 13, 4494–4504. [Google Scholar] [CrossRef] [PubMed]
- Mizukami, H.; Takahashi, K.; Inaba, W.; Tsuboi, K.; Osonoi, S.; Yoshida, T.; Yagihashi, S. Involvement of oxidative stress-induced DNA damage, endoplasmic reticulum stress, and autophagy deficits in the decline of β-cell mass in Japanese type 2 diabetic patients. Diabetes Care 2014, 37, 1966–1974. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Feng, X.; Li, Q.; Wang, Y.; Li, Q.; Hua, M. Adiponectin; TNF-α and inflammatory cytokines and risk of type 2 diabetes: A systematic review and meta-analysis. Cytokine 2016, 86, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Fathy, S.A.; Mohamed, M.R.; Ali, M.A.M.; Elhelaly, A.E.; Alattar, A.T. Influence of IL-6, IL-10, IFN-γ and TNF-α genetic variants on susceptibility to diabetic kidney disease in type 2 diabetes mellitus patients. Biomarkers 2018, 31, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Esteve, E.; Ricart, W.; Fernández-Real, J.M. Gut microbiota interactions with obesity, insulin resistance and type 2 diabetes: Did gut microbiote co-evolve with insulin resistance? Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 483–490. [Google Scholar] [CrossRef]
- Diamant, M.; Blaak, E.E.; De Vos, W.M. Do nutrient-gut microbiota interactions play a role in human obesity, insulin resistance and type 2 diabetes? Obes. Rev. 2011, 12, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Bäckhed, F.; Fulton, L.; Gordon, J.I. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell. Host. Microbe 2008, 3, 213–223. [Google Scholar] [CrossRef]
- Neacsu, O.; Cleveland, K.; Xu, H.; Tchkonia, T.T.; Kirkland, J.L.; Boney, C.M. IGF-I attenuates FFA-induced activation of JNK1 phosphorylation and TNF-α expression in human subcutaneous preadipocytes. Obesity 2013, 21, 1843–1849. [Google Scholar] [CrossRef] [PubMed]
- Compare, D.; Rocco, A.; Sanduzzi Zamparelli, M.; Nardone, G. The gut bacteria-driven obesity development. Dig. Dis. 2016, 34, 221–229. [Google Scholar] [CrossRef] [PubMed]
- China Pharmacopoeia Committee. Chinese Pharmacopoeia; China Medical Science Press: Beijing, China, 2010; pp. 85–86. [Google Scholar]
- Xu, J.; Han, Q.B.; Li, S.L.; Chen, X.J.; Wang, X.N.; Zhao, Z.Z.; Chen, H.B. Chemistry, bioactivity and quality control of Dendrobium, a commonly used tonic herb in traditional Chinese medicine. Phytochem. Rev. 2013, 12, 341–367. [Google Scholar] [CrossRef]
- Qu, X.Y.; Wu, Y.X.; Zhang, J.L. Research advances on chemical constituents and pharmacological activities of Dendrobium. Tianjin Agric. Sci. 2015, 21, 19–32. [Google Scholar]
- Miyazawa, M.; Shimamura, H.; Nakamura, S.; Kameoka, H. Antimutagenic activity of gigantol from Dendrobium nobile. J. Agric. Food Chem. 1997, 45, 2849–2953. [Google Scholar] [CrossRef]
- Miyazawa, M.; Shimamura, H.; Nakamura, S.; Sugiura, W.; Kosaka, H.; Kameoka, H. Moscatilin from Dendrobium nobile, a naturally occurring bibenzyl compound with potential antimutagenic activity. J. Agric. Food Chem. 1999, 47, 2163–2167. [Google Scholar] [CrossRef]
- Chen, T.H.; Pan, S.L.; Guh, J.H.; Liao, C.H.; Huang, D.Y.; Chen, C.C.; Teng, C.M. Moscatilin induces apoptosis in human colorectal cancer cells: A crucial role of c-Jun NH2-terminal protein kinase activation caused by tubulin depolymerization and DNA damage. Clin. Cancer Res. 2008, 14, 4250–4258. [Google Scholar] [CrossRef]
- Chen, C.C.; Wu, L.G.; Ko, F.N.; Teng, C.M. Antiplatelet aggregation principles of Dendrobium loddigesii. J. Nat. Prod. 1994, 57, 1271–1274. [Google Scholar] [CrossRef]
- Liu, Y.N.; Pan, S.L.; Peng, C.Y.; Huang, D.Y.; Guh, J.H.; Chen, C.C.; Shen, C.C.; Teng, C.M. Moscatilin repressed lipopolysaccharide-induced HIF-1α accumulation and NF-κB activation in murine raw264.7 cells. Shock 2010, 33, 70–75. [Google Scholar] [CrossRef]
- Lu, Y.; Kuang, M.; Hu, G.P.; Wu, R.B.; Wang, J.; Liu, L.; Lin, Y.C. Loddigesiinols G–J: α-glucosidase inhibitors from Dendrobium loddigesii. Molecules 2014, 19, 8544–8555. [Google Scholar] [CrossRef]
- Zhang, J.P.; Zheng, X.L.; Hong, J.Z.; Chen, J.C.; Zheng, Y.Y.; Xin, J.Z.; Wang, Q.Y.; Zhu, K.D.; Wang, X.N.; Shi, H. Dendrobium compound in treating 90 case of type 2 diabetes memtus. J. Fujian Univ. TCM 2011, 21, 6–10. [Google Scholar]
- Wu, H.Z.; Xu, J.H.; Chen, L.Z.; Sun, J.J. Studies on anti-hyperglycemic effect and its mechanism of Dendrobium candidum. Chin. J. Chin. Mat. Med. 2004, 29, 160–163. [Google Scholar]
- Gong, C.Y.; Yu, Z.Y.; Lu, B.; Yang, L.; Sheng, Y.C.; Fan, Y.M.; Ji, L.L.; Wang, Z.T. Ethanol extract of Dendrobium chrysotoxum Lindl ameliorates diabetic retinopathy and its mechanism. Vascul. Pharmacol. 2014, 62, 134–142. [Google Scholar] [CrossRef]
- Li, C.Y.; Lu, Y.; Chen, Y.; Zheng, J.W.; Wang, J. Chemical components of Dendrobium loddigesii. Acta Sci. Nat. Univ. Sunyatseni. 2013, 52, 73–76. [Google Scholar]
- Ito, M.; Matsuzaki, K.; Wang, J.; Daikonya, A.; Wang, N.L.; Yao, X.S.; Kitanaka, S. New phenanthrenes and stilbenes from Dendrobium loddigesii. Chem. Pharm. Bull. 2010, 58, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Li, X.W.; Chen, H.P.; He, W.B.; Yang, W.L.; Ni, F.Y.; Huang, Z.W.; Hu, H.Y.; Wang, J. Polyphenols from Dendrobium loddigesii and their biological activities. Acta Sci. Nat. Univ. Sunyatseni 2018. accepted. [Google Scholar]
- Xu, C.J.; Wang, H.B. Research progress of Shihu Yeguang Pill. Chin. J. Chin. Ophthalmol. 2016, 4, 266–268. [Google Scholar]
- Lee, G.H.; Proenca, R.; Montez, J.M.; Carroll, K.M.; Darvishzadeh, J.G.; Lee, J.I.; Friedman, J.M. Abnormal splicing of the leptin receptor in diabetic mice. Nature 1996, 379, 632–635. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Chen, D.; Zhang, J.N.; Lv, X.; Wang, K.; Duan, L.P.; Nie, Y.; Wu, X.L. Bacterial community mapping of the mouse gastrointestinal tract. PLoS ONE 2013, 8, e74957. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Mazcorro, J.F.; Ivanov, I.; Mills, D.A.; Noratto, G. Influence of whole-wheat consumption on fecal microbial community structure of obese diabeticmice. Peer J. 2016, 4, e1702. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Shi, H. Research progress on Dendrobium single agent in the treatment of diabetes. Chin. Med. Mod. Dis Edu Chin. 2016, 17, 148–150. [Google Scholar]
- Zheng, H.; Whitman, S.A.; Wu, W.; Wondrak, G.T.; Wong, P.K.; Fang, D.; Zhang, D.D. Therapeutic potential of Nrf2 activators in streptozotocin-induced diabetic nephropathy. Diabetes 2011, 60, 3055–3066. [Google Scholar] [CrossRef] [PubMed]
- Brasnyó, P.; Molnár, G.A.; Mohás, M.; Markó, L.; Laczy, B.; Cseh, J.; Mikolás, E.; Szijártó, I.A.; Mérei, A.; Halmai, R.; et al. Resveratrol improves insulin sensitivity, reduces oxidative stress and activates the Akt pathway in type 2 diabetic patients. Br. J. Nutr. 2011, 106, 383–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.H.; Lee, E.S.; Choi, R.; Nawaboot, J.; Lee, M.Y.; Lee, E.Y.; Kim, H.S.; Chung, C.H. Protective effects of curcumin on renal oxidative stress and lipid metabolism in a rat model of type 2 diabetic nephropathy. Yonsei Med. J. 2016, 57, 664–673. [Google Scholar] [CrossRef] [PubMed]
- Verdam, F.J.; Fuentes, S.; de Jonge, C.; Zoetendal, E.G.; Erbil, R.; Greve, J.W.; Buurman, W.A.; de Vos, W.M.; Rensen, S.S. Human intestinal microbiota composition is associated with local and systemic inflammation in obesity. Obesity 2013, 21, E607–E615. [Google Scholar] [CrossRef]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef]
- Larsen, N.; Vogensen, F.K.; van den Berg, F.W.J.; Nielsen, D.S.; Andreasen, A.S.; Pedersen, B.K.; Al-Soud, W.A.; Sørensen, S.J.; Hansen, L.H.; Jakobsen, M. Gut microbiota in human adults with type 2 diabetes differs from non-diabetic adults. PLoS ONE 2010, 5, e9085. [Google Scholar] [CrossRef]
- Carmody, R.N.; Gerber, G.K.; Luevano, J.M., Jr.; Gatti, D.M.; Somes, L.; Svenson, K.L.; Turnbaugh, P.J. Diet dominates host genotype in shaping the murine gut microbiota. Cell. Host Microbe. 2015, 17, 72–84. [Google Scholar] [CrossRef]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [Green Version]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed]
- Forslund, K.; Hildebrand, F.; Nielsen, T.; Falony, G.; Le Chatelier, E.; Sunagawa, S.; Prifti, E.; Vieira-Silva, S.; Gudmundsdottir, V.; Pedersen, H.; et al. Disentangling the effects of type 2 diabetes and metformin on the human gut microbiota. Nature 2015, 528, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.R.; Lee, J.C.; Lee, H.Y.; Kim, M.S.; Whon, T.W.; Lee, M.S.; Bae, J.W. An increase in the Akkermansia spp. population induced by metformin treatment improves glucose homeostasis in diet-induced obese mice. Gut 2014, 63, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef] [PubMed]
- LeRoith, D.; Shiloach, J.; Roth, J.; Lesniak, M.A. Insulin or a closely related molecule is native to Escherichia coli. J. Biol. Chem. 1981, 256, 6533–6536. [Google Scholar]
- Amar, J.; Chabo, C.; Waget, A.; Klopp, P.; Vachoux, C.; Bermúdez-Humarán, L.G.; Smirnova, N.; Bergé, M.; Sulpice, T.; Lahtinen, S.; et al. Intestinal mucosal adherence and translocation of commensal bacteria at the early onset of type2 diabetes: Molecular mechanisms and probiotic treatment. EMBO Mol. Med. 2011, 3, 559–572. [Google Scholar] [CrossRef]
- Monk, J.M.; Lepp, D.; Wu, W.; Pauls, K.P.; Robinson, L.E.; Power, K.A. Navy and black bean supplementation primes the colonic mucosal microenvironment to improve gut health. J. Nutr. Biochem. 2017, 49, 89–100. [Google Scholar] [CrossRef]
- Suzuki, T.; Yoshida, S.; Hara, H. Physiological concentrations of short-chain fatty acids immediately suppress colonic epithelial permeability. Br. J. Nutr. 2008, 100, 297–305. [Google Scholar] [CrossRef]
- Liao, T.; Chen, Y.P.; Huang, S.Q.; Tan, L.L.; Li, C.Q.; Huang, X.A.; Xu, Q.; Wang, Q.; Zeng, Q.P. Chondroitin sulfate elicits systemic pathogenesis in mice by interfering with gut microbiota homeostasis. BioRxiv 2017. [Google Scholar] [CrossRef]
- Liao, T.; Chen, Y.P.; Tan, L.L.; Li, C.Q.; Wang, Q.; Huang, S.Q.; Huang, X.A.; Xu, Q.; Zeng, Q.P. Chondroitin sulfate flourishes gut sulfatase-secreting bacteria to damage mucus layers, leak bacterial debris, and trigger inflammatory lesions in mice. BioRxiv 2017. [Google Scholar] [CrossRef]
- Sinclair, L.; Osman, O.A.; Bertilsson, S.; Eiler, A. Microbial community composition and diversity via 16S rRNA gene amplicons: Evaluating the illumina platform. PLoS ONE 2015, 10, e0116955. [Google Scholar] [CrossRef] [PubMed]
- Fadrosh, D.W.; Ma, B.; Gajer, P.; Sengamalay, N.; Ott, S.; Brotman, R.M.; Ravel, J. An improved dual-indexing approach for multiplexed 16S rRNA gene sequencing on the Illumina MiSeq platform. Microbiome 2014, 2, 6. [Google Scholar] [CrossRef] [PubMed]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods. 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.R.; Wang, Q.; Fish, J.A.; Chai, B.; McGarrell, D.M.; Sun, Y.; Brown, C.T.; Porras-Alfaro, A.; Kuske, C.R.; Tiedje, J.M. Ribosomal Database Project: Data and tools for high throughput rRNA analysis. Nucl. Acid. Res. 2014, 42, D633–642. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; Sahl, J.W.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- White, J.R.; Nagarajan, N.; Pop, M. Statistical methods for detecting differentially abundant features in clinical metagenomic samples. PLoS Comput. Biol. 2009, 5, e1000352. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucl. Acid. Res. 2004, 32, D277–280. [Google Scholar] [CrossRef]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Samples of the rich-polyphenols extract of D. Loddigesii or marker compounds (1–4) are not available from the authors. |













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | TG | TC | HDL-C | LDL-C | INS |
|---|---|---|---|---|---|
| mmol/L | mIU/L | ||||
| DM | 1.45 ± 0.21 | 5.06 ± 0.49 | 0.99 ± 0.18 | 1.19 ± 0.21 | 7.06 ± 1.63 |
| DMMET130 | 1.32 ± 0.29 | 4.69 ± 0.59 | 1.01 ± 0.12 | 1.20 ± 0.12 | 11.18 ± 1.82 #, 1 |
| Control | 1.01 ± 0.27 | 2.51 ± 0.58 | 1.10 ± 0.21 | 0.65 ± 0.10 | 10.64 ± 3.74 |
| DMDJP25 | 1.50 ± 0.31 | 4.98 ± 0.69 | 0.93 ± 0.12 | 1.25 ± 0.11 | 8.22 ± 3.80 |
| DMDJP50 | 1.48 ± 0.19 | 4.93 ± 0.59 | 0.96 ± 0.19 | 0.69 ± 0.20 # | 10.93 ± 3.48 # |
| DMDJP100 | 1.49 ± 0.33 | 4.48 ± 0.25 | 1.03 ± 0.12 | 0.65 ± 0.11 # | 11.43 ± 3.37 # |
| Group | N | Grading of Liver Fatty Degeneration | Grading of Vacuolar Degeneration of Renal Tubule Epithelium Cells | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | + | ++ | +++ | ++++ | +++++ | 0 | ++ | +++ | ||
| DM | 8 | 0 | 0 | 0 | 0 | 5 | 3 | 0 | 6 | 2 |
| DMMET130 *, 1 | 8 | 3 | 0 | 0 | 1 | 2 | 2 | 8 | 0 | 0 |
| Control | 8 | 8 | 0 | 0 | 0 | 0 | 0 | 8 | 0 | 0 |
| DMDJP25 | 8 | 1 | 0 | 0 | 2 | 4 | 1 | 1 | 4 | 3 |
| DMDJP50 #, 2 | 8 | 3 | 2 | 0 | 1 | 2 | 0 | 2 | 4 | 2 |
| DMDJP100 ##, 3, * | 6 | 1 | 1 | 2 | 2 | 0 | 0 | 6 | 0 | 0 |
| Group | Expression of TNF-α in Liver Tissue | Expression of TNF-α in Kidney Tissue |
|---|---|---|
| DM | 0.21 ± 0.03 | 0.077 ± 0.005 |
| DMMET130 | 0.18 ± 0.05 | 0.110 ± 0.002 |
| Control | 0.069 ± 0.006 | 0.028 ± 0.003 |
| DMDJP100 | 0.088 ± 0.005 ##, 1 | 0.034 ± 0.003 #, 2 |
| Tissue | Oxidative Stress Index | Group | ||||
|---|---|---|---|---|---|---|
| DM | DMMET130 | Control | DMDJP100 | |||
| Liver | GSH | μmol/gprot | 37.06 ± 2.87 | 40.58 ± 2.61 #, 1 | 44.51 ± 3.70 | 41.86 ± 3.43 # |
| MDA | 0.66 ± 0.03 | 0.34 ± 0.05 ##, 2 | 0.59 ± 0.08 | 0.44 ± 0.05 ## | ||
| CAT | U/mgprot | 45.92 ± 4.41 | 57.48 ± 4.27 ## | 47.21 ± 1.59 | 70.35 ± 5.07 ## | |
| SOD | 161.5 ± 5.9 | 161.7 ± 7.5 | 205.8 ± 8.7 | 177.4 ± 6.4 # | ||
| Kidney | GSH | μmol/gprot | 27.88 ± 2.26 | 37.82 ± 2.72 # | 36.27 ± 3.88 | 33.38 ± 3.09 # |
| MDA | 1.79 ± 0.08 | 1.38 ± 0.13 ## | 1.31 ± 0.15 | 1.51 ± 0.11 # | ||
| CAT | U/mgprot | 12.43 ± 5.23 | 30.48 ± 5.49 ## | 30.11 ± 3.34 | 32.21 ± 4.53 ## | |
| SOD | 83.65 ± 6.8 | 115.2 ± 6.3 ## | 131.1 ± 5.8 | 111.0 ± 5.6 # | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.-W.; Chen, H.-P.; He, Y.-Y.; Chen, W.-L.; Chen, J.-W.; Gao, L.; Hu, H.-Y.; Wang, J. Effects of Rich-Polyphenols Extract of Dendrobium loddigesii on Anti-Diabetic, Anti-Inflammatory, Anti-Oxidant, and Gut Microbiota Modulation in db/db Mice. Molecules 2018, 23, 3245. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23123245
Li X-W, Chen H-P, He Y-Y, Chen W-L, Chen J-W, Gao L, Hu H-Y, Wang J. Effects of Rich-Polyphenols Extract of Dendrobium loddigesii on Anti-Diabetic, Anti-Inflammatory, Anti-Oxidant, and Gut Microbiota Modulation in db/db Mice. Molecules. 2018; 23(12):3245. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23123245
Chicago/Turabian StyleLi, Xue-Wen, Hui-Ping Chen, Ying-Yan He, Wei-Li Chen, Jian-Wen Chen, Lu Gao, Hai-Yan Hu, and Jun Wang. 2018. "Effects of Rich-Polyphenols Extract of Dendrobium loddigesii on Anti-Diabetic, Anti-Inflammatory, Anti-Oxidant, and Gut Microbiota Modulation in db/db Mice" Molecules 23, no. 12: 3245. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23123245