1. Introduction
Hemp (
Cannabis sativa L.) is often indicated as the oldest known, cultivated, and used plant by humanity. Its wide development was facilitated by broad soil and climate adaptation, allowing a large geographical distribution, as well as by crop flexibility, giving textile, fiber, food, feed, and biosolvents as final products or intermediates for manufacturing industries. A parallel history involves relative plants with secondary metabolism producing cannabinoids, and particularly tetrahydrocannabinoids (THCs), which characterize crops for medicinal or illegal recreational use due to psychotropic effects and addiction. As a consequence of the longtime debate on the correct taxonomic level of distinction between THC-producing or non-THC-producing plants, for most of the 20th century, a general prohibition of hemp cultivation existed. Currently, the non-THC-producing species of
Cannabis sativa, or at least those producing a concentration lower than 0.2%, are usually referred to as fiber hemp, industrial hemp, and seed-oil hemp. Formerly, the maximum THC limit was imposed on plant cultivation in the open field and was not fixed by law with regards to final products such as food or nutraceuticals. The Italian Government promoted industrial hemp cultivation as a strategy to implement environmentally friendly crops and to contrast the loss of agricultural lands and green belts, as well as the desertification and the decrement of biodiversity [
1].
In the Abruzzo region (central Italy), hemp cultivation is the object of renewed interest; both the surface dedicated to hemp crops and the volume of plant material produced are increasing. The economy of hemp crops is mainly sustained by female flower collection. This represents an innovative and high-value product that is used for tea infusion, to aromatize beer and other food products, and for nutraceutical purposes, involving extraction of important secondary metabolites [
2]. Futura 75 cultivar was selected in France and is characterized by low THC content (<0.2%) and is widely cultivated for the production of fiber and seeds, while floral buds and leaves are considered as waste or by-products [
3,
4]. In this context, the essential oil (EO) obtained via hydro-distillation could represent a solution for the novelty and the multipurpose applications in medicinal, cosmetic, and food industries, because it could be obtained as primary product from female flowers or aerial parts, as well as from crop by-products, such as residual green parts (leaves and stems) after flower collection or from female inflorescences.
A previous study compared the composition and the antimicrobial activities of EOs obtained from three varieties of industrial hemp (Carmagnola, Fibranova, and Futura). Futura cultivar gave the highest essential oil yield and resulted in the most promising for application as an antimicrobial agent with a broad spectrum of activity [
5]. Another comparative study was conducted on five hemp cultivars (Felina 34, Fedrina 74, SwissMix, Kompolti, and Secuemi) for chemical composition and antimicrobial activity. The most abundant compounds were found to be α-pinene, myrcene,
trans-β-ocimene, γ-terpinolene, (
E)-caryophyllene, and α-humulene. The disc diffusion test revealed moderate antimicrobial activity which different among the cultivars [
6].
More recently, the EO obtained from inflorescences or leaves of hemp cv. Futura 75 was proposed as a potential source of botanical insecticides against larvae of
Spodoptera littoralis and
Culex quinquefasciatus and against adults of
Musca domestica [
4]. The essential oil was also tested in a field trial to control the rosy apple aphid pest and was determined as an ecological alternative to the acetamiprid [
7]. An aqueous emulsion of the essential oil (0.02–0.1%), mainly characterized by (
E)-caryophyllene (35%), β-myrcene (18%), and α-pinene (9%), induced high mortality rate when tested on the foxglove aphid (
Aulacorthum solani Kalt.) and the two-spotted spider mite (
Tetranychus urticae Koch) [
8]. Hemp EO was also shown to fight the human-biting Asian tiger mosquito (
Aedes albopictus) and the rice-crop invasive snail
Physella acuta [
9]. Increasing interest for the essential oil is confirmed by the increasing literature data, which could be related to its limited toxicity for mammals [
10], as well as its low ecological impact on the environment [
11].
Due to the increasing attention in the valorization of locally produced plants to support their use in the pharmaceutic, cosmetic, and food industry and to valorize the cultivation by-products, the EO obtained via hydro-distillation of aerial parts from
C. sativa cv. Futura 75, cultivated in Italy, and its aromatic water were studied herein. Phytochemical investigation was performed on the terpenoid fraction (GC/MS), and the phenolic and flavonoid fingerprint (spectrophotometric and chromatographic methods), in addition to color analysis (CIELAB parameters; see
Section 2.2). The potential applications of the EO were investigated in a wide multidisciplinary approach for different biological activities, such as antioxidant and antiradical, enzyme inhibition, antimicrobial activities on bacteria and fungi, and a comparative cytotoxic activity on tumor and non-tumor cell lines. A preliminary in vivo evaluation of the toxicity was also carried out on
Galleria mellonella larvae. The broad-spectrum investigation provided a general characterization of the EO of THC-free
C. sativa and could be used to support its use and application in innovative and multi-target ingredients for medicinal, cosmetic, veterinary, agronomic, or food use.
2. Materials and Methods
2.1. Experimental Farm and Plant Extraction
Two experimental fields were selected in the surroundings of Chieti, Italy (42°21′46.5′′ N 14°05′49.2′′ E, 100 m above sea level) in the old alluvial plain of the Pescara River Valley. The climate is Mediterranean, with an average yearly temperature around 15 °C and mean yearly rainfall around 750 mm. Summer can be variably dry. Total extension of the experimental crops comprises 15,000 square meters on level ground (slope 0–5%). During the two years preceding, the farm field was uncultured and mowed to obtain hay. The sowings were scheduled on the last week of March 2016 and were planned to favor the plant development during a long photoperiod and to stimulate full blooming in late summer.
The selection of Futura 75 was done considering the local climatic and soil characteristics in relation to the evidence of the high adaptive capabilities of this variety which are useful for multi-purpose cultivation. Selected seeds were purchased from South Hemp Tecno (Taranto, Italy) and certified by SOC France (Service officiel de contrôle et de certification des semences et plants) according to the plant variety database of the European Commission in the agricultural species list (position A-85, FR 8194). Soil tillage consisted of superficial ploughing followed by disc harrowing and final milling to prepare the sowing bed. Sowing was performed using a mechanic seeder for wheat, settled to obtain a final density of 50 kg of seeds/ha. The sowing was in line every 30 cm with 10 cm between lines. No fertilizer was used and, during growth, three irrigation cycles were programmed (automatic irrigation system, each cycle 24–36 h, depending on soil water availability). Plants development was monitored weekly until the beginning of flowering, when manual collection of aerial parts from flowering plants was done. The collection was carried out every three days, starting on the first week of September up to October 2016. Immediately after harvest, the plant material was transferred to the laboratory for extraction (about 100 m away). Random samples were tested for macroscopic characteristics in order to confirm their botanical identity (done by Prof. L. Menghini).
Fresh aerial parts consisting of leaves, inflorescences, and thinner residues of stem were roughly cropped to uniform size and accurately distributed inside the distillation chamber. The distillatory was a stainless-steel 12-L Clevenger-type apparatus (Albrigiluigi S.r.l., Stallavena, Italy). The distillation started within 5 h of plant collection in the field and was performed for no longer than 3 h. Straightaway, the collected EOs were passed through septa with anhydrous sodium sulfate in order to remove water residues, before being transferred to a sealed blue glass bottle and stored in the dark at 4 °C until used for phytochemical and biological assays. The EO yields (%, v/w) were determined each time and were expressed as mean volume obtained from plant fresh weight. Moreover, as a by-product, the aromatic water was also characterized in our experiments to assess its potential economic value in the framework of a lower-waste agri-food chain. In agreement with “circular economy” principles, our approach could further provide innovative results for reducing waste load.
2.2. Color Analysis
CIELAB parameters (L*, a*, b*,
C*
ab, and
hab), according to the “Commission Internationale de l′Eclairage”, were determined using a colorimeter X-Rite SP-62 (X-Rite Europe GmbH, Regensdorf, Switzerland), equipped with a D65 illuminant and an observer angle of 10°. Color interpretation was based on the lightness L* (between 0, absolute black, and 100, absolute white), the greenness (negative) or redness (positive) a*, and the blueness (negative) or yellowness (positive) b*. The chroma, or saturation (
C*
ab) quantitatively describes the color intensity, and the hue or color angle (
hab) is the attribute of appearance by which a color is identified according to its resemblance to red, yellow, green, blue, or a combination of two of these [
12]. The results are reported as the mean value ± standard deviation (SD).
2.3. Gas Chromatography/Mass Spectrometry (GC/MS) Analysis
The volatile component analysis was carried out using an Agilent Technologies 6850 gas chromatograph (Santa Clara, CA, USA) coupled with an Agilent Technologies 5975 mass spectrometer (Santa Clara, CA, USA), equipped with an HP-5MS capillary column (5% phenyl 95% methylpolysiloxane, 30 m × 0.25 mm inner diameter, film thickness 0.25 µm; Hewlett-Packard, Palo Alto, CA, USA). GC parameters were adjusted as follows: injector temperature, 250 °C; flow rate of the helium carrier gas (99.995% purity), 1.0 mL/min. The oven temperature was set at 40 °C (5 min), then raised to 200 °C at 5 °C/min, and maintained at this temperature for 60 min. MS parameters were set as follows: energy of electron ionization, 70 eV; solvent delay, 6 min; source temperature, 230 °C; quadrupole temperature, 150 °C; and mass scan was carried out over the 50–350 m/z range.
The eluted compounds were identified by matching the relative mass spectra with those available from both a commercial database (FFNSC 3, Chromaleont srl, Messina, Italy) and online libraries (NIST 11, Flavor2, Scientific Instrument Services, Ringoes, NJ, USA) [
13]. Kovats index (KI) was used as a second parameter to confirm the analyte identification. KIs were measured using a mixture of
n-alkanes (C8–C24) in the same analytic conditions, and then compared with values reported in the literature and in the FFNSC 3 database. The identity of several compounds was confirmed through injection of standard samples available from commercial sources. The relative abundance of oil components was obtained by integrating the GC/MS peak areas without any further correction.
2.4. Total Phenolic Acid Content
The samples (1.0 mL) were added to 0.5 M hydrochloric acid (2 mL), 8.5% sodium hydroxide (2 mL), and Arnow reagent (10% aqueous solution of sodium nitrite and sodium molybdate, 2 mL), and diluted to 10.0 mL with water. The absorbance of the test solution was measured against the blank at 505 nm. Obtained results are reported as caffeic acid equivalents (CAE/g sample) [
14].
2.5. Total Phenolic Content
The total phenolic quantification was determined using the Folin–Ciocâlteu method [
15]. Test solutions (0.25 mL) were mixed with Folin–Ciocâlteu reagent (1 mL, 1:9
v/v). After 3 min, Na
2CO
3 solution (0.75 mL, 1%) was added and, after 2 h incubation at room temperature, the absorbance was measured at 760 nm. Results are expressed as milligrams of gallic acid equivalent per gram of sample (GAE/g sample).
2.6. Total Flavonoid Content
The total flavonoid quantification was measured using the AlCl
3 method by Berk at al. (2011) [
16]. Test solutions (1 mL) were mixed with aluminum trichloride (2% in methanol). Similarly, a blank was prepared (sample solution (1 mL) and methanol (1 mL)). The absorbances were recorded at 415 nm after a 10-min incubation at room temperature. Results are expressed as milligrams of rutin equivalent per gram of sample (RE/g sample) [
17].
2.7. HPLC Analysis
All chemicals were purchased from Sigma-Aldrich (Milan, Italy), whereas HPLC-grade solvents were obtained from Carlo Erba Reagenti (Milan, Italy). Double-distilled water was obtained using a Millipore Milli-Q Plus water treatment system (Millipore Bedford Corp., Bedford, MA, USA). HPLC photo diode array (PDA) phenolic patterns were established using the validated method reported in the literature [
18]. Data are reported as means ± standard deviation of three independent measurements.
4. Results and Discussion
The hemp crop produced approximatively 10,000–12,000 kg/ha of fresh biomass. The hydro-distillation of flowering aerial parts gave 0.28% essential oil yield (mL/kg fresh weight), expressed as the mean value of separate distillations from 2 kg of fresh weight each. Minimum and maximum ranges of EO were 0.19% and 0.31%, respectively. The highest yield value was obtained in the early-September distillation, while the lowest was from the late-September/early-October collection (full blooming plants) (
Figure S1, Supplementary Materials). As expected, the difference in EO yield was not evident in day-by-day analysis, while there was a significant difference (*
p < 0.05) in the data comparison between the collections on the first and last days (0.29 ± 0.02% and 0.19 ± 0.01% mean yield ± SEM, 1 September and 2 October, respectively). The trend of yield variation was also evident upon analyzing data grouped by decades, confirming the early collection as the preferable practice for optimal yield (0.29 ± 0.004%, 0.25 ± 0.005%, and 0.21 ± 0.003% mean yield ± SEM; 1–11 September, 12–22 September, and 23 September–2 October, respectively (ANOVA,
p < 0.0001; ***
p < 0.001). The quantitative production of hemp EO was monitored for one month. Fresh plant material was collected daily, manually selecting plants only in full bloom, and three or four separated distillations were immediately performed. Experimental data confirm that plant harvest is preferable at the early flowering stage due to the evident reduction in EO content in proximity to fruiting. From the weekly data analysis, the reduction (−30%) of EO production was evident in late flowering stages, while all yield data within the same week were more homogenous. At the end of the production process, all collected hemp EO samples were pooled to provide a single batch available for the market and for the successive analyses. The aromatic water, separated from the EO but enriched by fragrant molecules, was further investigated for its chemical composition, antioxidant activity, color characteristics, and enzyme inhibitory activity with the aim of giving value to this important by-product.
4.1. Phytochemical Analyses
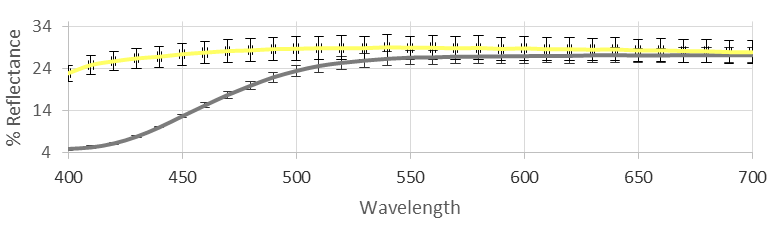
4.1.1. Color Analysis
The tristimulus colorimetry was not yet reported in the literature to evaluate the color properties of hemp EO and its aromatic water. The EO, obtained from the fresh inflorescences via steam distillation and separated by water, mainly contains a mixture of monoterpene and sesquiterpene hydrocarbons. Our analyses, as subsequently described, account for twelve components belonging to this class, in which (
E)-caryophyllene and caryophyllene oxide represent the principal impact compounds. As a counterpart in the aromatic water, a polyphenolic fraction mainly represented by catechins and phenolic acids was found. The color of the EO, which could vary between pale yellow and light green was found to show a significant yellow parameter (b*) along with a weak green value (a*), which, together with the quite bright L* value, account for the light yellow-green nuance characterizing our sample. The aromatic water showed a very clear yellow color associated with a relatively high L* value and particularly low a* and b* parameters. If this is related to the catechin content, the more intense yellow-green color of the separated oil could be maybe attributed to other pigments, weakly represented but intensely colored, such as chlorophylls, carotenoids, or others [
37]. The colorimetric data, complete with standard deviations, are reported in
Table 1, and the reflectance curves are shown in
Figure 1.
4.1.2. GC Flame Ionization Detector Analysis of the Tetrahydrocannabinol (THC) Content
The THC content was routinely checked on C. sativa var. Futura 75 aerial parts and on the processed final product (hemp EO) and was found to be below the limits of Italian government law (THC < 0.2%).
4.1.3. GC/MS Analysis of the Volatile Components of the EO
The GC/MS analysis of hemp EO exhibited several peaks that were related to different classes of phytochemicals. The most abundant components belonged to the class of sesquiterpenes and sesquiterpenoids (28% (
E)-caryophyllene, 4%
trans-α-bergamotene, 13% humulene, 7% α- and β-selinene, and 15% caryophyllene oxide) representing about 67% of the total composition (
Table 2). Monoterpenoids were also present in an appreciable amount (11% total pinene fraction and 11% β-myrcene), together with lower-abundance components ranging from 6% (α-terpinolene) to 2% (
d-limonene).
4.1.4. Total Phenolic, Flavonoid, and Phenolic Acid Content
The amounts of total bioactive compounds in the aromatic water were detected by colorimetric methods, and the results are summarized in
Table 3 (hemp EO was not soluble in the assay reagents). Total phenolics, flavonoids, and phenolic acids were found to be 28.04 mg GAE/g, 4.04 mg RE/g, and 1.76 mg CE/g extract, respectively. Apparently, the total amount of flavonoids and phenolic acids was approximately 20% of total phenolics. Several researchers reported different levels of total bioactive compounds in
Cannabis sativa extracts [
38,
39,
40,
41,
42] and, generally, the extracts were found to be rich sources of these compounds. These differences in the literature may also be linked to geographical and climatic conditions, or genetic structures.
4.1.5. HPLC-PDA Analysis of the Phenolic Fraction
Both hemp EO and its aromatic water, studied for the presence of the most important 22 phenolics, displayed an interesting profile in terms of richness and metabolite-oriented amount (
Table 4) [
43].
Naringenin (706 µg/mL) and its glycosylated derivative naringin (83 µg/mL) emerged, among the ones studied, as the two most important metabolites which could further characterize this EO, along with the presence of catechin (60 µg/mL) and epicatechin (56 µg/mL) (see
Supplementary Materials for the chromatogram of a real sample). Considering the mean concentration of naringenin, hemp EO seems to be relevant as a source of this important flavanone for cosmetic and pharmaceutical purposes. Moreover, naringenin was shown to act as an antioxidant, anti-inflammatory, anti-cancer, anti-diabetic, and anti-obesity agent [
44]. Other compounds such as benzoic acid, 2,3-dimethoxybenzoic acid, and syringic acid were also detected in discrete quantities. Conversely, the number and the relative amounts of these phytochemicals were scarcely found in the aromatic water (rich in catechin), despite the unique presence of a small percentage of rutin (0.18 µg/mL). Chlorogenic acid, vanillic acid, 3-hydroxy-4-methoxybenzaldehyde,
p-coumaric acid, sinapinic acid,
o-coumaric acid, harpagoside,
t-cinnamic acid, and carvacrol were absent in this EO and in its aromatic water.
4.2. Antioxidant Properties
Antioxidative effects of natural products could be considered as a first insight in detecting their ethnopharmacological relevance and potential. To this end, a certain antioxidant profile could be provided by comparing different chemical assays representative of alternative mechanisms. For this reason, the antioxidant ability of
C. sativa samples was evaluated using six complementary in vitro tests: free-radical scavenging (DPPH and ABTS), reducing power (CUPRAC and FRAP), phosphomolybdenum, and metal chelating assays. The results are tabulated in
Table 5. The aromatic water exhibited stronger free-radical scavenging and ferric reducing potential when compared to the EO. In addition, the EO was not active on ABTS radicals. However, the best activity in phosphomolybdenum and metal chelating assays was obtained by the EO. These findings may be linked to the presence of a large amount of naringenin and catechin, which act as good chelators [
45,
46]. Also, the phosphomolybdenum assay is known as a total antioxidant test, and different phytochemicals, including vitamin C, tocopherol, or carotenoids, may be active in this assay. CUPRAC and FRAP tests showed good results with these products, which, along with metal chelating activity, could account for the potential of hemp EO in limiting the generation and the accumulation of harmful free radicals via the Fenton reaction. Similarly, some researchers [
39,
40,
47,
48] reported that
C. sativa has great potential as a source of natural antioxidants for developing novel functional applications.
4.3. Antimicrobial and Antibiofilm Activity of the Hemp EO versus Staphylococcus aureus
Staphylococcus aureus is a Gram-positive bacterium which permanently or transiently colonizes up to 60% of the human population and, for this reason, is also considered a component of the human microbiota [
49].
S. aureus, however, is associated with nosocomial infections including those involving orthopedic implants [
50], and may be responsible for many infections such as endocarditis, osteomyelitis, prostatitis, and infections of skin and soft tissue [
51]. Its persistence and recalcitrance are due to the capability of
S. aureus to develop biofilms as a complex structure formed by bacterial cells embedded in an extracellular polymeric substance (EPS), a mixture of macromolecules such as exopolysaccharides, proteins, and extracellular DNA. The EPS composition may differ with bacterial strain, culture conditions, and biofilm age. Biofilm formation guarantees antibiotic tolerance and protection from the host immune system, making microbial biofilms difficult to eradicate [
52].
The inhibitory effect of hemp EO against Gram-positive opportunistic/pathogenic microorganisms such as
Clostridium spp. and
Enterococcus spp. was demonstrated [
5], suggesting its use against nosocomial, spoilage, and food-borne pathogens. In our paper, the antimicrobial and antibiofilm effects of hemp EO were evaluated against five
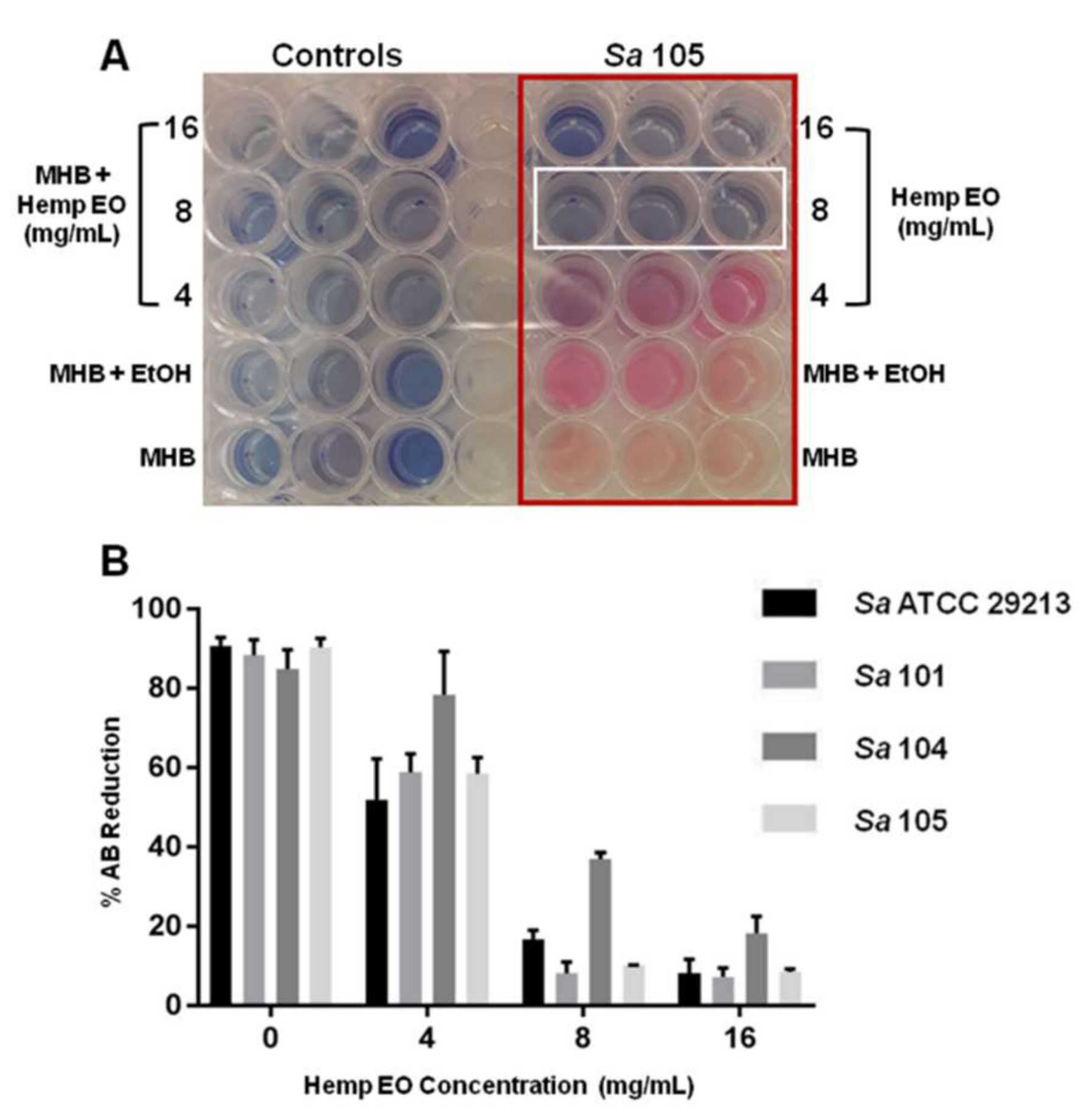
S. aureus strains, one reference and four clinical strains, isolated from different areas of the body and also characterized by a different antimicrobial susceptibility pattern (
Table 6;
Table S1, Supplementary Materials).
The hemp EO showed MIC values corresponding to 8 mg/mL versus all
S. aureus strains, including
S. aureus 104, a multi-drug resistant strain isolated from a pharyngeal swab of a male patient (
Table 6). With regards to MBC, the hemp EO was shown to be effective at 16 mg/mL versus all
S. aureus strains (
Table 6). The results obtained were confirmed using the AB reduction assay (
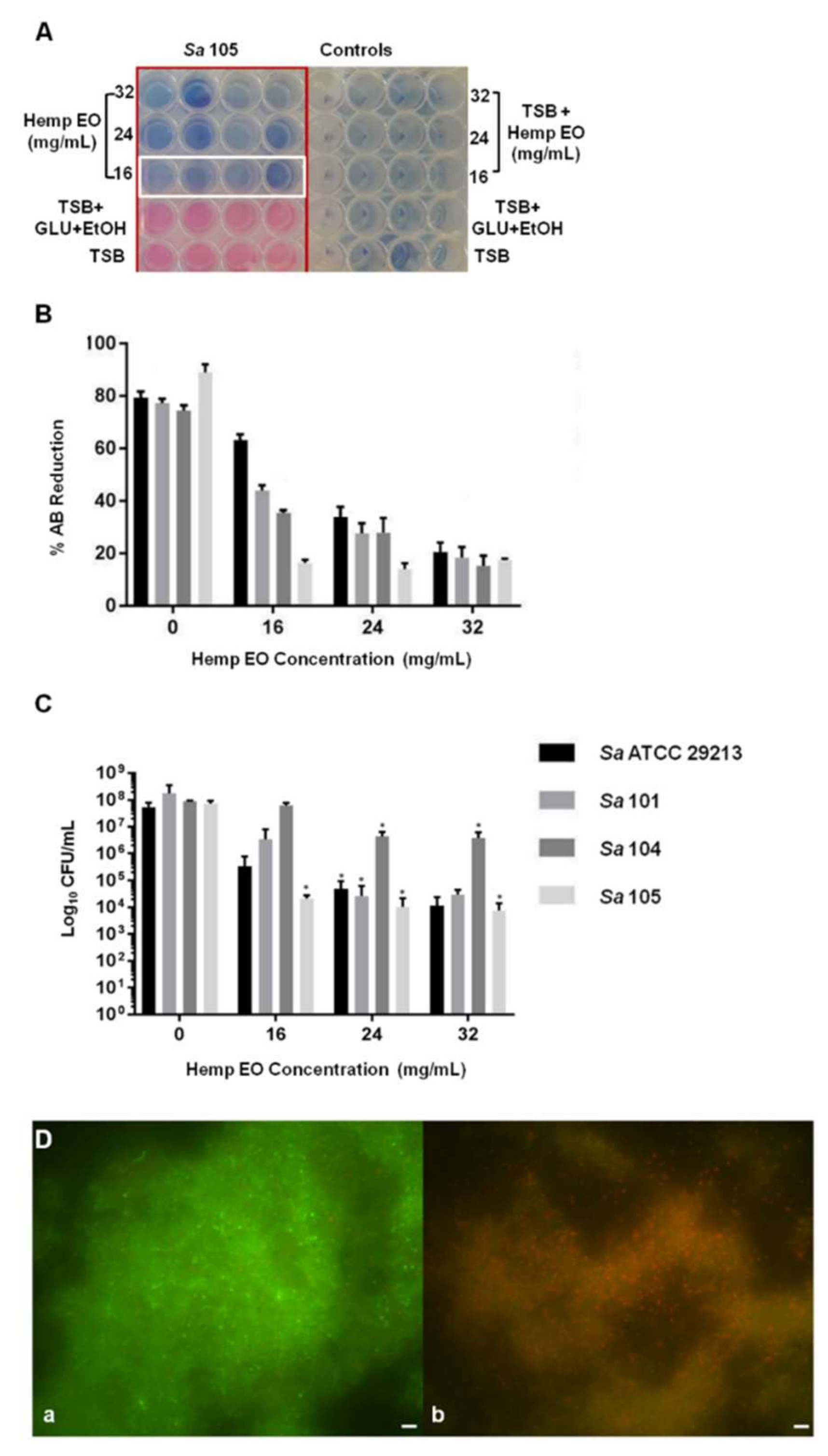
Figure 2A,B).
Furthermore, the hemp EO showed its capability to eradicate a mature biofilm developed by
S. aureus; specifically, the minimum biofilm eradication concentration (MBEC) was determined using the AB assay, CV staining, CFU counting, and live/dead staining, followed by fluorescent microscopy analysis. The Hemp EO displayed an MBEC value of 24 mg/mL toward all strains of
S. aureus except for
S. aureus 105, which showed an MBEC value corresponding to 16 mg/mL (
Table 6 and
Figure 3A,B). The effect of the hemp EO on the eradication of
S. aureus pre-formed biofilms was confirmed by the statistically significant reduction in CFU count, as shown in
Figure 3C. The live/dead staining and fluorescent microscopy evaluation showed a well-structured biofilm constituting a great amount of live cells, as indicated by the green fluorescence (
Figure 3D(a)). On the contrary,
S. aureus mature biofilms treated with 24 mg/mL hemp EO did not show a reduction in biofilm biomass, as confirmed by the CV staining (data not shown); however, the biofilms treated with hemp EO were characterized by a multitude of dead cells as indicated by the red fluorescence (
Figure 3D(b)). As mentioned above, the CV staining did not display any differences between the biomasses of treated and untreated biofilms (data not shown).
Due to the high amount of naringenin in this EO, we aimed to evaluate the impact of this secondary metabolite on the antimicrobial activity. We also studied the MIC, MBC, and MBEC of naringenin against
S. aureus 105. The data indicated an MIC of 512 μg/mL and an MBEC corresponding to 2048 μg/mL. The results demonstrated the antimicrobial and antibiofilm effects of naringenin contained in the hemp EO versus
S. aureus. These results also confirmed the data of Yue et al. [
53] who reported the effect of naringenin on the formation of the biofilm of
Streptococcus mutans. The authors demonstrated that the MIC of naringenin was between 100 and 200 μg/mL, and that 200 μg/mL naringenin was capable of inhibiting the biofilm formation after 4 and 24 h of incubation. Conversely, in the present study, we focalized the attention on the capability of naringenin to eradicate a mature biofilm developed by
S. aureus after 24 h of incubation to evaluate an application of hemp EO in the treatment of pre-formed biofilms.
Moreover, the antimicrobial effect of naringenin was also previously investigated on several bacterial species. In particular, the mechanism of action of naringenin was studied on
Escherichia coli ATCC 25922 and
S. aureus ATCC 6538. The authors demonstrated that the presence of naringenin induced modifications of cell-membrane fluidity, fatty-acid composition, and fatty-acid biosynthesis-associated genes. Furthermore, the growth of
S. aureus was significantly inhibited by low concentrations of naringenin [
54,
55].
4.4. Anti-Helicobacter Pylori Activity
The antimicrobial potency of hemp EO and its main component naringenin was also explored against 14
H. pylori clinical isolates and the reference strain ATCC 43629, using the microdilution method at pH 7.5 [
56]. As shown in
Table 7, the activity of the two test samples was not related to the different antibiotic susceptibility of the strains; the EO showed an MIC
50 of 32 µg/mL and an MIC
90 of 64 µg/mL, whereas naringenin showed an MIC
50 of 16 µg/mL and an MIC
90 of 32 µg/mL. By considering the strains resistant to the traditional antibiotics (MNZ and CLR), hemp EO was 2–4-fold more active than MNZ, and 4–16-fold more active than CLR. The potency of naringenin was 2–8- and 4–16-fold higher than MNZ and CLR, respectively. MBC values were 64 µg/mL for hemp EO and 32 µg/mL for naringenin. The MIC and MBC concentrations of all strains were the same for both compounds, except for the strains E34, 68, and F40/442, which had MBC values corresponding to two times the MIC. Because a drug is defined “bactericidal” when the MBC concentration does not exceed four-fold MIC value [
57], we can conclude that hemp EO and naringenin have very good bactericidal effects (1 < MBC/MIC < 2).
Nariman and coworkers [
58] reported a study about the anti-
Helicobacter pylori effect of
C. sativa and other 19 plants extracts, collected from Iran areas. Of the 10 strains tested, six were defined susceptible by disc diffusion method. The MIC values of the 10 most effective extracts (
C. sativa not included) against
Helicobacter pylori ranged from 31.25 to 500 µg/mL. Following these studies and the threshold values reported by Kuete (2010) [
59], our results demonstrate that hemp EO and naringenin have very good activity against
H. pylori. Furthermore, it was reported that cannabidiol prevents intestinal inflammation damage induced by an alteration of cytokine levels [
60]. Because the pathogenic effect of
H. pylori infection is supposed to also be mediated by cytokine production of inflammatory cells present in the infected gastric mucosa [
61], the antimicrobial activity of
C. sativa extracts demonstrated in this study, associated with their anti-inflammatory effect, could represent a very interesting approach against
Helicobacter pylori infection.
4.5. Antifungal Activity of Hemp EO
The antifungal activity of hemp EO against
Candida spp. and
Malassezia spp. was calculated as MIC value >12,460 µg/mL with no differences observed among
Candida spp. and
Malassezia spp. Hemp EO was not active against yeasts following the defined threshold activity values for plant extracts proposed as follows: MIC below 100 μg/mL (significant activity), 100 ≤ MIC ≤ 625 μg/mL (moderate activity), and MIC > 625 μg/mL (low activity) [
59]. These results reinforced the selective antibacterial activity of hemp EO.
4.6. Cytotoxicity
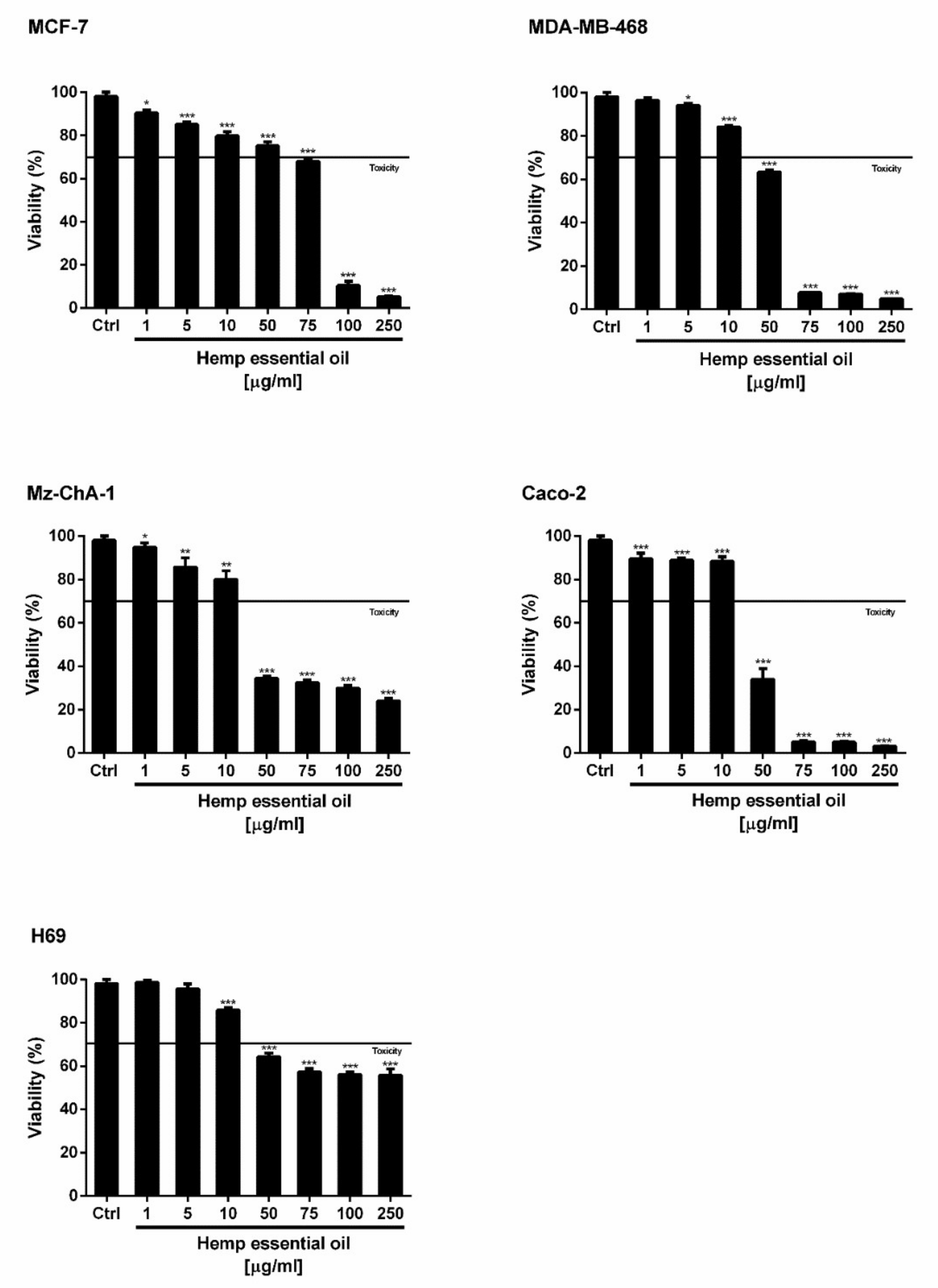
To the best of our knowledge, the cytotoxicity of hemp EO in cancer cells is yet to be investigated. Under our experimental conditions, the hemp EO induced a statistically significant and concentration-dependent reduction of cancer cell viability, reaching almost a 75% inhibition of cell proliferation at the highest concentration of 250 µg/mL (
Figure 4). A different anti-proliferative potency was found in the tested cell models, with Caco-2 and Mz-ChA-1 cells as the most sensitive biological systems. In fact, after 24 h of treatment with 50 µg/mL hemp EO, the cell viability was inhibited by about 66% in both cell lines (
Figure 4). Accordingly, similar half maximal inhibitory concentration (IC
50) values were calculated for Caco-2 and Mz-ChA-1 cells (
Table 8).
Next, hemp EO significantly inhibited the growth of both breast cancer cell lines characterized by different biological features and heterogeneous susceptibility to anticancer agents, with more potency against MDA-MB-468 cells. In particular, the treatment with 50 and 75 µg/mL produced an inhibition of cell viability of 25% and 32% in MCF7 cells, and of 37% and 92% in MDA-MB-468 cells; 90% proliferation inhibition was reached only at 100 µg/mL in MCF7 cells. As expected, the resulting IC
50 value of the Hemp EO in MCF7 cells was about 1.6-fold higher than that obtained for MDA-MB-468 cells, and almost threefold higher than that calculated for Caco-2 and Mz-ChA-1 cells. In contrast, slight cytotoxicity was produced in the nonmalignant cholangiocytes (H69), reaching a maximum of 44% inhibition of cell proliferation at the highest concentration of 250 µg/mL; as a consequence, the IC
50 was not evaluable. On the basis of these results, the cytotoxicity of hemp EO appeared to be selective for cancer cells, particularly cholangiocarcinoma and coloncarcinoma, with a higher tolerability in normal cells. The cytotoxicity was similar to that of the positive control doxorubicin in Caco-2 and Mz-ChA-1 cells, while about 10-fold lower in breast cancer MCF7 and MDA-MB-468 cells (
Table 8). Of note, this behavior suggests that hemp EO can affect specific cancer targets, which deserves to be deeply investigated.
Upon phytochemical analysis, several terpenoids, including monoterpenes and sesquiterpenes, along with flavonoids and tannins, were identified in this phytocomplex. Interestingly, α-pinene and β-caryophyllene exhibited high anti-proliferative activity on human erythroleukemic K562 cells [
62]; moreover, β-caryophyllene and linalool exhibited interesting anti-proliferative activity on human amelanotic melanoma C32 cells and on renal cell adenocarcinoma cells [
63]. β-Caryophyllene was also reported to possess chemopreventive properties and to interfere with liver cancer cell proliferation by inhibiting the activation of signal transducer and activator of transcription 3 (STAT3) signaling [
64], whereas β-caryophyllene oxide was found to possess anti-proliferative activity, although at high concentrations, and chemosensitizing properties, thus restoring the sensitivity of resistant cancer cells to chemotherapy [
65]. β-Myrcene was reported to possess anti-invasive properties in metastatic MDA-MB-231 human breast cancer cells through the inhibition of nuclear factor kappa B (NF-κB) activity [
66].
d-Limonene is widely known to possess chemopreventive and antitumor properties; in a previous study, it induced tumor regression and cancer cell apoptosis, and inhibited inflammation and oxidative stress [
67].
Among phenolics, naringin was reported to possess antitumor properties and to inhibit the growth of gastric cancer cells through the induction of autophagy [
68]. The anti-proliferative and pro-apoptotic activity of naringenin is also well documented [
69,
70]. This evidence suggests that several phytoconstituents can represent the possible bioactive agents of hemp EO, although the role of unique interactions of all the compounds in the phytocomplex cannot be excluded.
4.7. Enzyme Inhibition
Enzyme inhibitory effects of the tested
C. sativa samples were investigated toward a panel of important enzymes such as acetylcholinesterase (AChE), butyrylcholinesterase (BChE), α-amylase, α-glucosidase, tyrosinase, and lipase. The results are presented in
Table 9. Generally, from the enzymatic standpoint, the highest values were observed in the EO, except for the lack of inhibitory effect on AChE and α-amylase. These variations observed can be explained by the different phytochemical compositions of the
C. sativa samples. For example, the EO is richer in terms of volatile components, such as (
E)-caryophyllene, caryophyllene oxide, and humulene, which are known as anti-tyrosinase agents [
71]. Similarly, naringenin, the major compound in the EO, is very active on α-glucosidase and pancreatic lipase [
72,
73]. In particular, lipase inhibition was the most promising effect after treatment with hemp EO.
In addition, the complex nature of phytochemicals or their interactions could be responsible for the observed inhibitory activities. Up to now, few studies were carried out on the enzyme inhibitory effect of
C. sativa [
42,
74,
75]. With this regard, the present study could provide a starting contribution for the scientific audience, and this plant may serve to prepare functional ingredients as a source of natural inhibitors.
4.8. In Vivo Toxicity Studies
Larvae of
Galleria mellonella are widely used as an invertebrate wax moth larva model to evaluate the virulence of microbial pathogens, measure the efficacy/safety of biologically active antimicrobial agents, and produce results comparable to those that can be obtained using mammals [
76]. As a consequence, no ethical approval was required for this study because there was no use of mammal or animal models. In this work, the viability of
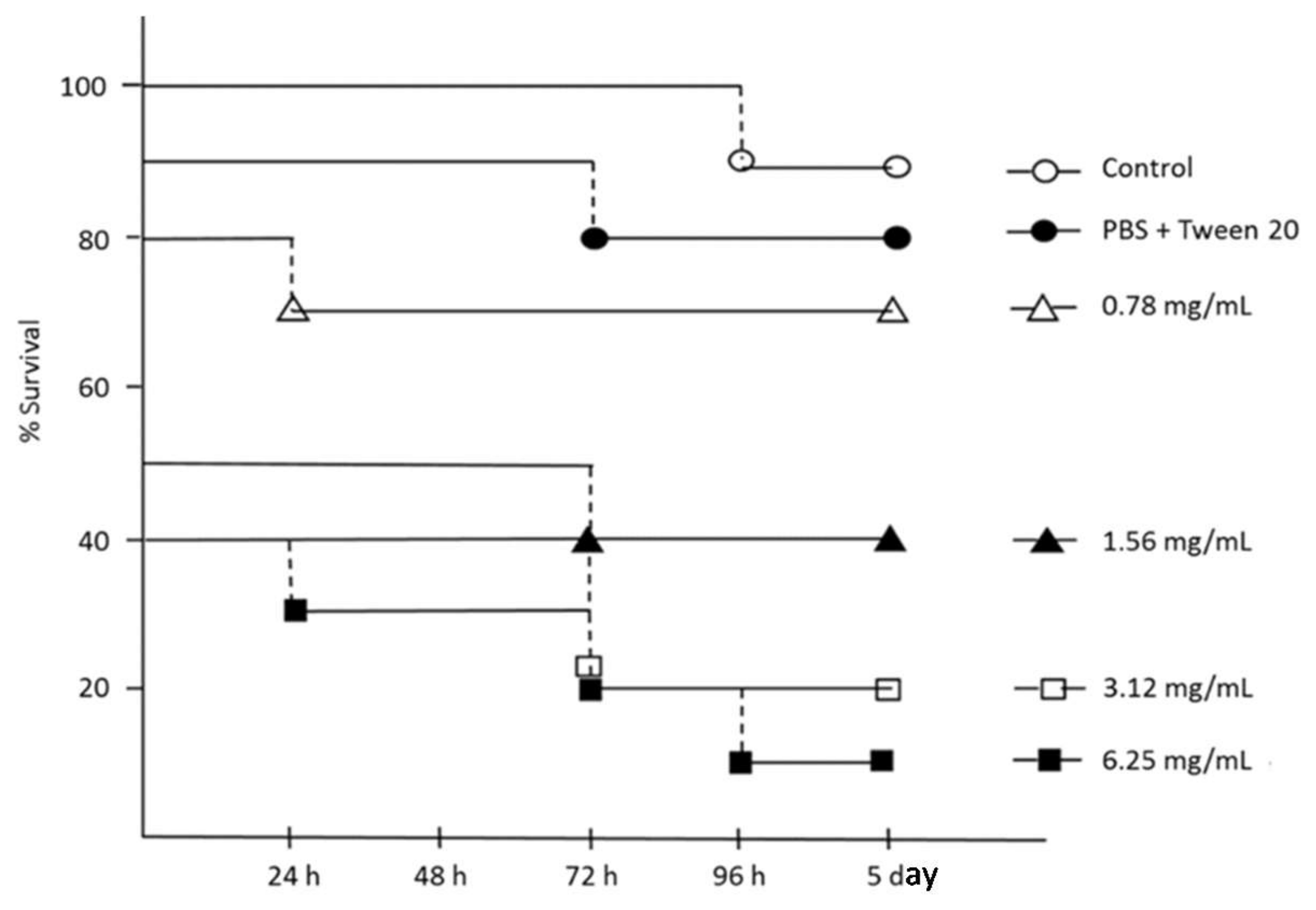
G. mellonella larvae, to measure the relative toxicity of different concentrations of hemp EO, was evaluated over the study period of five days. As reported in
Figure 5, no evident effects on survival were registered with 0.78 mg/mL hemp EO up to five days after treatment. The lethal dose (LD
50) of Hemp EO was 1.56 mg/mL in the first 48 h of treatment, probably due to the lipophilic nature of the EO with respect to the larval biological fluids and the invasive administration procedure (
Figure 5).
The LD50 of 1.56 mg/mL was higher than the effective concentrations reported by us against H. pylori (8–64 μg/mL) and cell lines (22.3–250 μg/mL), but in the same range as those found active against S. aureus strains (8–24 mg/mL). In this case, the putative topical administration of the hemp EO to combat S. aureus wound infections guarantees a discrete safety index with respect to the parental administration in G. mellonella larvae.
5. Conclusions
The study of traditional crops could explain their ethnobotanical relevance by means of a deep analysis of the quali-quantitative phytochemical profile. We devoted our attention to the EO of the aerial parts of C. sativa var. Futura 75 cultivated in Italy, due to an increment in this crop’s cultivation. After the evaluation of tetrahydrocannabinol content, volatile fraction, phenolic and flavonoid pattern, and color parameters, we tested the EO of C. sativa aerial parts and its aromatic water against several targets of pharmaceutical interest (S. aureus, H. pylori, Candida and Malassezia spp., enzymes, and cancer cell lines), along with the preliminary evaluation of its safety toward a non-cancer cell line and G. mellonella larvae in vivo. In addition, due to the high content of naringenin, we further explored the inhibitory effects of this secondary metabolite against H. pylori and S. aureus to correlate the biological activity with the chemical composition. In particular, the antibacterial and antibiofilm activities of hemp EO suggested it could be a possible candidate for the treatment of infections related to those abovementioned microorganisms. Currently, reinvigorating the antimicrobial drug pipeline is mandatory, as increasingly resistant species continue to emerge or organize themselves in accessible communities. Secondary metabolites display drug-like and metabolite-like properties, acting synergistically with validated drugs, and possessing new mechanisms of action; nevertheless, the evaluation of their safety/tolerability is strongly requested.

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}