Small Molecule Oligopeptides Isolated from Walnut (Juglans regia L.) and Their Anti-Fatigue Effects in Mice


Abstract
:1. Introduction
2. Results
2.1. Effects of WOPs on the Body Weight of Mice
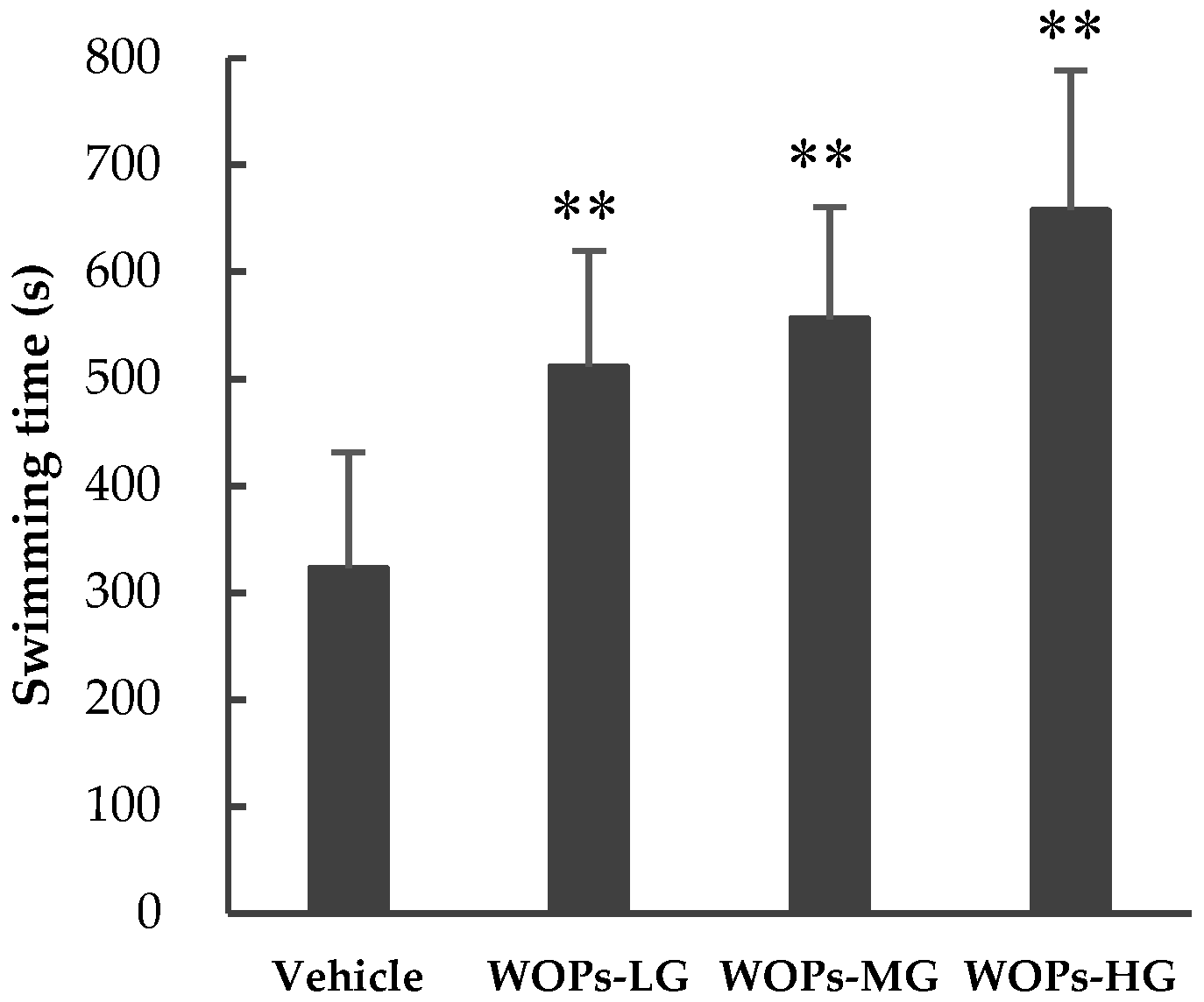
2.2. Effects of WOPs on Weight-Loaded Forced Swimming Endurance Time
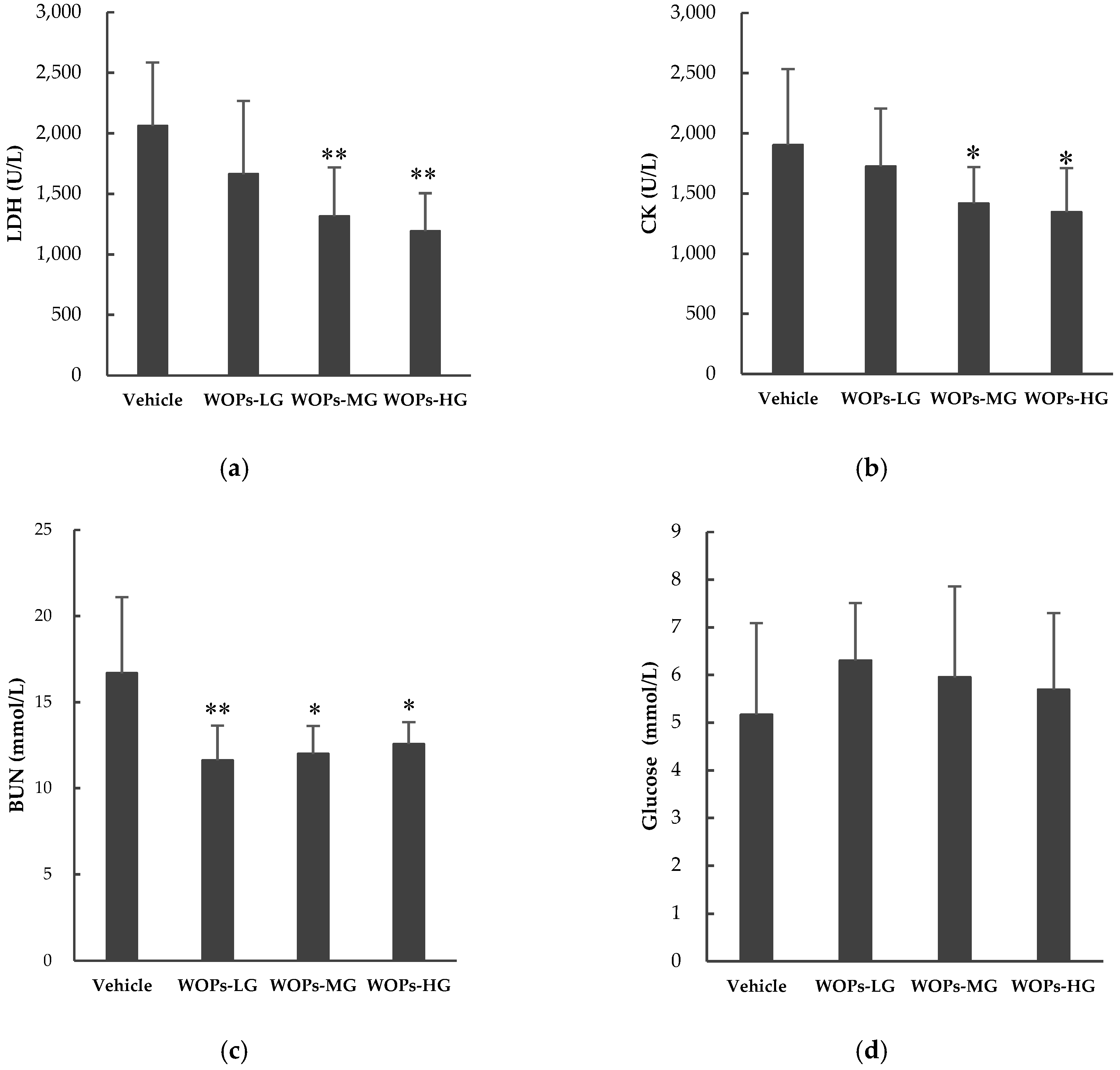
2.3. Effects of WOPs on Lactate Dehydrogenase (LDH), Blood Urea Nitrogen (BUN), Creatine Kinase (CK) and Blood Glucose Content
2.4. Effects of WOPs on Blood Lactic Acid (BLA) Levels in Mice
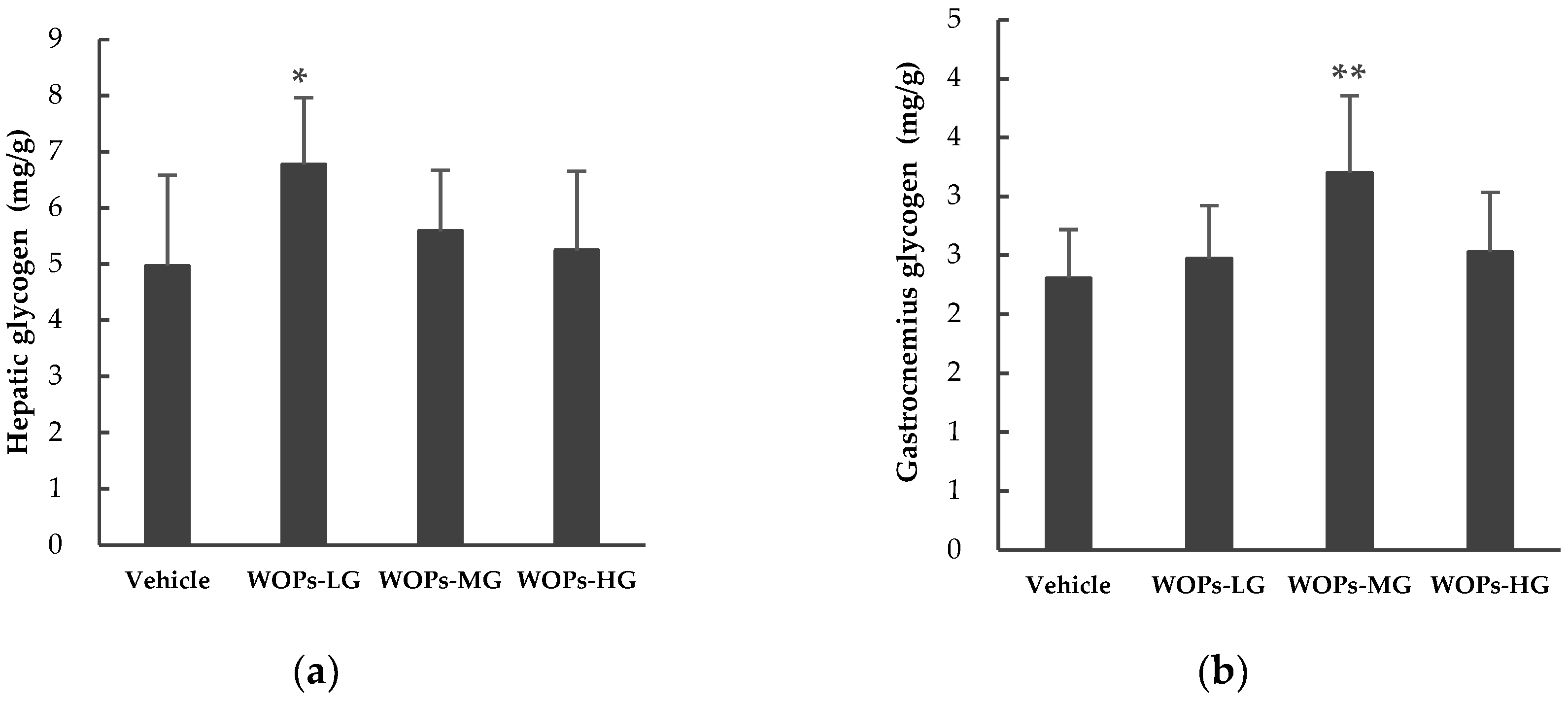
2.5. Effects of WOPs on Hepatic and Muscular Glycogen Level
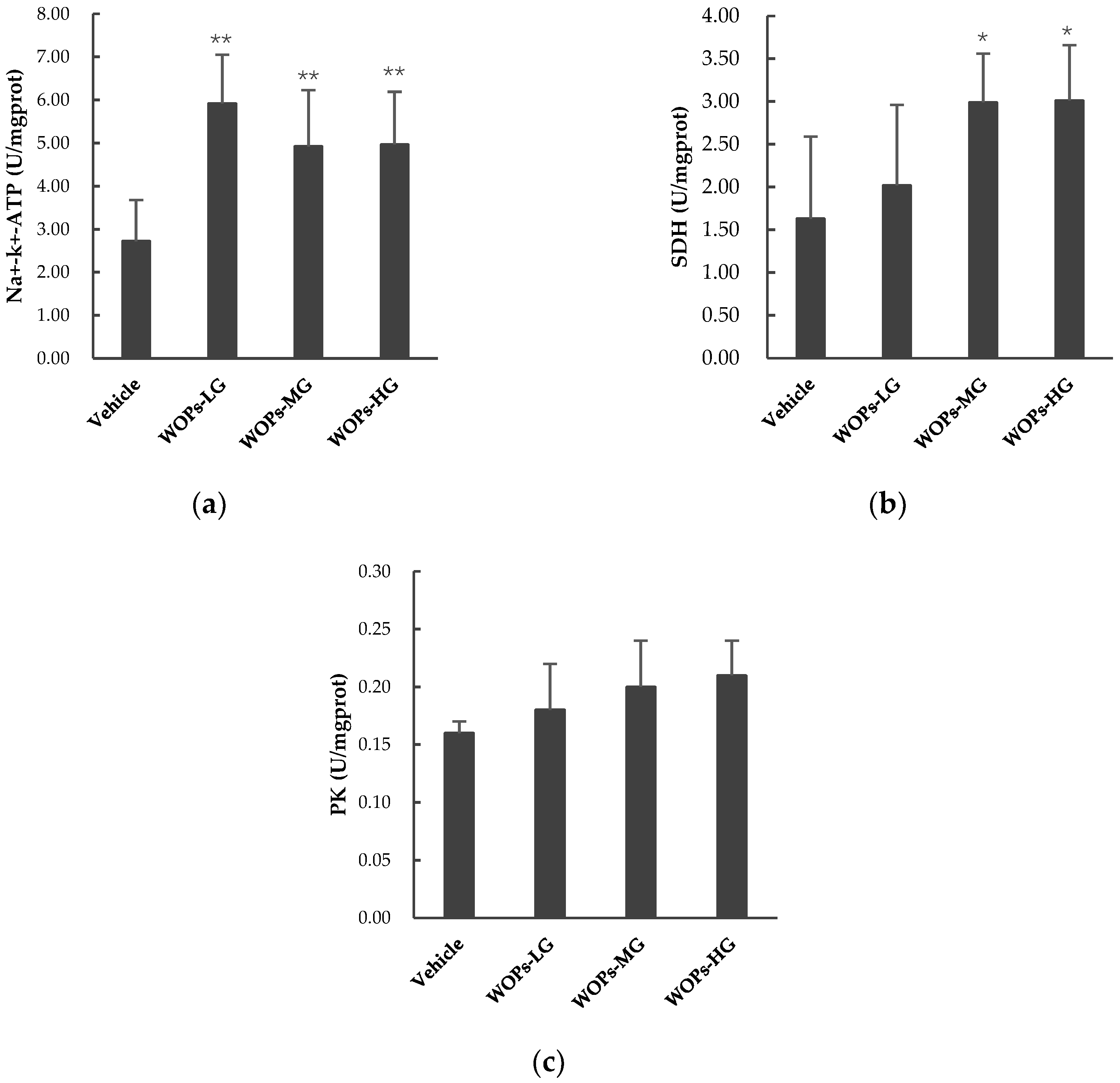
2.6. Effects of WOPs on PK, SDH, and Na+-K+-ATPase Activities in Skeletal Muscles of Mice
2.7. Effects of WOPs on Parameters of Oxidative Stress in Skeletal Muscles of Mice
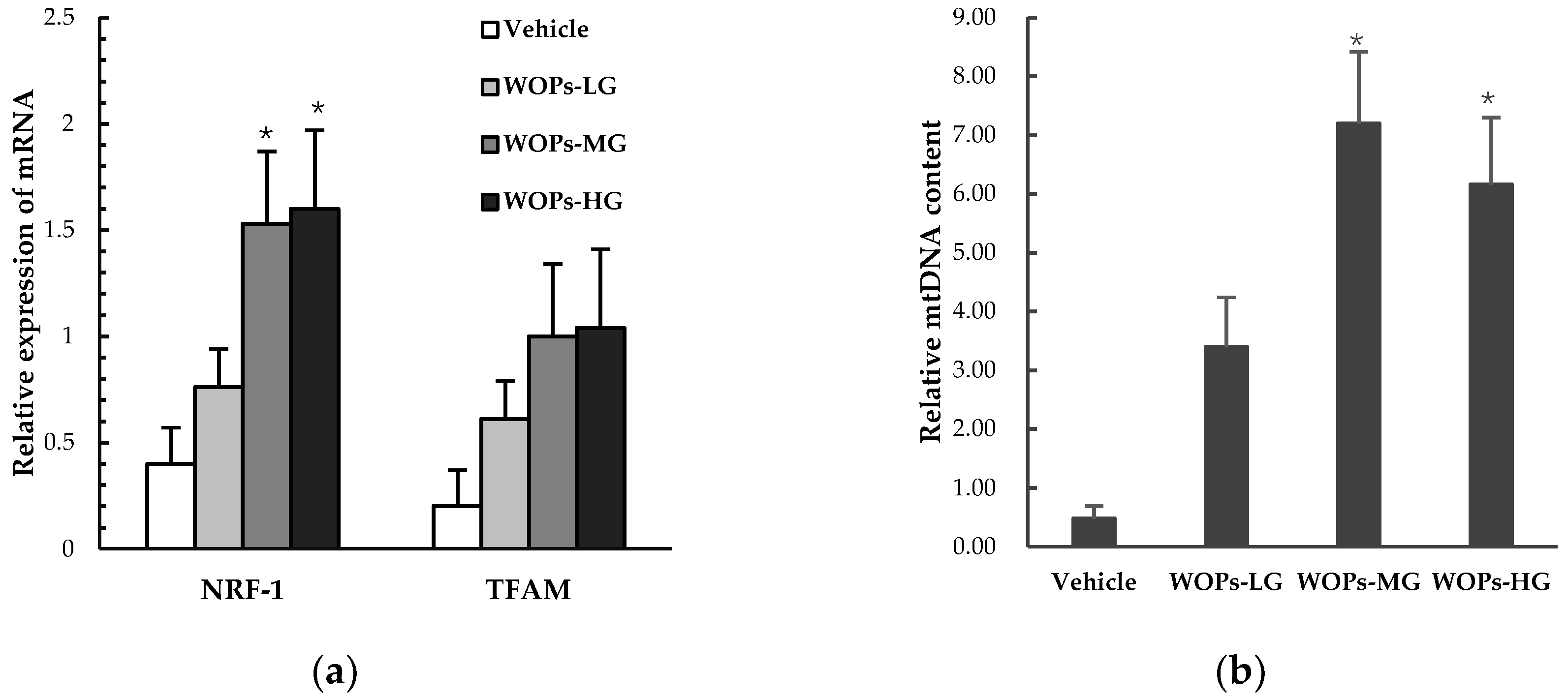
2.8. Effects of WOPs on Mitochondrial Biogenesis Factors and mtDNA Copy Number in Skeletal Muscles
3. Discussion
4. Materials and Methods
4.1. Preparation and Identification of WOPs
4.2. Chemicals and Reagents
4.3. Animals and Experimental Design
4.4. Weight-Loaded Swimming Test
4.5. Biochemical Assay
4.6. Determination of Blood Lactic Acid
4.7. Examination of Glycogen in Liver and Skeletal Muscles
4.8. Quantitative Real-Time PCR and Analyses of mtDNA Content
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Penner, I.K.; Paul, F. Fatigue as a symptom or comorbidity of neurological diseases. Nat. Rev. Neurol. 2017, 13, 662–675. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, K.; Tanaka, M.; Nozaki, S.; Mizuma, H.; Ataka, S.; Tahara, T.; Sugino, T.; Shirai, T.; Kajimoto, Y.; Kuratsune, H.; et al. Antifatigue effects of coenzyme Q10 during physical fatigue. Nutrition 2008, 24, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Li, Z.P.; Zhu, T.; Fung, H.Y.; Wong, T.L.; Wen, X.; Ma, D.L.; Leung, C.H.; Han, Q.B. Anti-fatigue effects of the unique polysaccharide marker of Dendrobium officinale on BALB/c mice. Molecules 2017, 22, 155. [Google Scholar] [CrossRef] [PubMed]
- Duan, F.F.; Guo, Y.; Li, J.W.; Yuan, K. Antifatigue effect of Luteolin-6-C-Neohesperidoside on oxidative stress injury induced by forced swimming of rats through modulation of Nrf2/ARE signaling pathways. Oxid. Med. Cell. Longev. 2017, 2017, 3159358. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.J.; Qin, Z.; Wang, P.Y.; Sun, Y.; Liu, X. Muscle fatigue: General understanding and treatment. Exp. Mol. Med. 2017, 49, 384. [Google Scholar] [CrossRef] [PubMed]
- Jin, G.; Kataoka, Y.; Tanaka, M.; Mizuma, H.; Nozaki, S.; Tahara, T.; Mizuno, K.; Yamato, M.; Watanabe, Y. Changes in plasma and tissue amino acid levels in an animal model of complex fatigue. Nutrition 2009, 25, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Potter, M.; Tomas, C.; Elson, J.L.; Morten, K.J.; Poulton, J.; Wang, N.; Jin, H.; Hou, Z.; Huang, W.E. A new approach to find biomarkers in chronic fatigue syndrome/myalgic encephalomyelitis (CFS/ME) by single-cell Raman micro-spectroscopy. Analyst 2018. [Google Scholar] [CrossRef]
- Bao, L.; Cai, X.; Wang, J.; Zhang, Y.; Sun, B.; Li, Y. Anti-fatigue effects of small molecule oligopeptides isolated from Panax ginseng C. A. Meyer in mice. Nutrients 2016, 8, 807. [Google Scholar] [CrossRef]
- Chen, J.; Wang, X.; Cai, Y.; Tang, M.; Dai, Q.; Hu, X.; Huang, M.; Sun, F.; Liu, Y.; Xia, P. Bioactivity-guided fractionation of physical fatigue-attenuating components from Rubus parvifolius L. Molecules 2013, 18, 11624–11638. [Google Scholar] [CrossRef]
- Li, D.; Ren, J.W.; Zhang, T.; Liu, R.; Wu, L.; Du, Q.; Li, Y. Anti-fatigue effects of small-molecule oligopeptides isolated from Panax quinquefolium L. in mice. Food Funct. 2018, 9, 4266–4273. [Google Scholar] [CrossRef]
- Chen, Y.M.; Lin, C.L.; Wei, L.; Hsu, Y.J.; Chen, K.N.; Huang, C.C.; Kao, C.H. Sake protein supplementation affects exercise performance and biochemical profiles in power-exercise-trained mice. Nutrients 2016, 8, 106. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.P.; Zhang, Y.; Liu, Z.; Chen, J.Y.; Zhang, S.Y.; Yang, X.D.; Zhou, H.L. Acute toxicity and anti-fatigue activity of polysaccharide-rich extract from corn silk. Biomed. Pharmacother. 2017, 90, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Yang, H.; Sun, Y.; Niu, J.; Liu, S. Purification and identification of antioxidant peptides from walnut (Juglans regia L.) protein hydrolysates. Peptides 2012, 38, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.; Huang, D.; Zhai, M.; Yang, L.; Peng, S.; Chen, C.; Feng, X.; Weng, Q.; Zhang, B.; Xu, M. Isolation of a novel bio-peptide from walnut residual protein inducing apoptosis and autophagy on cancer cells. BMC Complement. Altern. Med. 2015, 15, 413. [Google Scholar] [CrossRef] [PubMed]
- Poulose, S.M.; Bielinski, D.F.; Shukitt-Hale, B. Walnut diet reduces accumulation of polyubiquitinated proteins and inflammation in the brain of aged rats. J. Nutr. Biochem. 2013, 24, 912–919. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Du, M.; Zhang, Y.; Xu, W.; Wang, C.; Wang, K.; Zhang, L. Purification and identification of an ACE inhibitory peptide from walnut protein. J. Agric. Food Chem. 2013, 61, 4097–4100. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.; Zhang, R.; Dong, C.; Yu, Z.; Ren, J. Novel walnut peptide-selenium hybrids with enhanced anticancer synergism: Facile synthesis and mechanistic investigation of anticancer activity. Int. J. Nanomed. 2016, 11, 1305–1321. [Google Scholar] [CrossRef]
- Kim, D.I.; Kim, K.S. Walnut extract exhibits anti-fatigue action via improvement of exercise tolerance in mice. Lab Anim. Res. 2013, 29, 190–195. [Google Scholar] [CrossRef]
- He, L.X.; Wang, J.B.; Sun, B.; Zhao, J.; Li, L.; Xu, T.; Li, H.; Sun, J.Q.; Ren, J.W.; Liu, R.; et al. Suppression of TNF-alpha and free radicals reduces systematic inflammatory and metabolic disorders: Radioprotective effects of ginseng oligopeptides on intestinal barrier function and antioxidant defense. J. Nutr. Biochem. 2017, 40, 53–61. [Google Scholar] [CrossRef]
- Xu, X.; Ding, Y.; Yang, Y.; Gao, Y.; Sun, Q.; Liu, J.; Yang, X.; Wang, J.; Zhang, J. β-glucan salecan improves exercise performance and displays anti-fatigue effects through regulating energy metabolism and oxidative stress in mice. Nutrients 2018, 10, 858. [Google Scholar] [CrossRef]
- Kan, N.W.; Ho, C.S.; Chiu, Y.S.; Huang, W.C.; Chen, P.Y.; Tung, Y.T.; Huang, C.C. Effects of resveratrol supplementation and exercise training on exercise performance in middle-aged mice. Molecules 2016, 21, 661. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.Q.; Zeng, L.; Yang, Z.S.; Huang, F.F.; Ding, G.F.; Wang, B. Anti-fatigue effect by peptide fraction from protein hydrolysate of Croceine croaker (Pseudosciaena crocea) swim bladder through inhibiting the oxidative reactions including DNA damage. Mar. Drugs 2016, 14, 221. [Google Scholar] [CrossRef] [PubMed]
- Gobatto, C.A.; de Mello, M.A.; Sibuya, C.Y.; de Azevedo, J.R.; dos Santos, L.A.; Kokubun, E. Maximal lactate steady state in rats submitted to swimming exercise. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2001, 130, 21–27. [Google Scholar] [CrossRef]
- Lin, Y.; Liu, H.L.; Fang, J.; Yu, C.H.; Xiong, Y.K.; Yuan, K. Anti-fatigue and vasoprotective effects of quercetin-3-O-gentiobiose on oxidative stress and vascular endothelial dysfunction induced by endurance swimming in rats. Food Chem. Toxicol. 2014, 68, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.M.; Wei, L.; Chiu, Y.S.; Hsu, Y.J.; Tsai, T.Y.; Wang, M.F.; Huang, C.C. Lactobacillus plantarum TWK10 supplementation improves exercise performance and increases muscle mass in mice. Nutrients 2016, 8, 205. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Lin, D.; Guo, J.; Zhang, Y.; Zheng, B. In vitro antioxidant activity and in vivo anti-fatigue effect of sea horse (hippocampus) peptides. Molecules 2017, 22, 482. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Jackson, M.J. Exercise-induced oxidative stress: Cellular mechanisms and impact on muscle force production. Physiol. Rev. 2008, 88, 1243–1276. [Google Scholar] [CrossRef]
- Downs, M.L.; Semic-Jusufagic, A.; Simpson, A.; Bartra, J.; Fernandez-Rivas, M.; Rigby, N.M.; Taylor, S.L.; Baumert, J.L.; Mills, E.N. Characterization of low molecular weight allergens from English walnut (Juglans regia). J. Agric. Food Chem. 2014, 62, 11767–11775. [Google Scholar] [CrossRef]
- Allen, D.G.; Lamb, G.D.; Westerblad, H. Skeletal muscle fatigue: Cellular mechanisms. Physiol. Rev. 2008, 88, 287–332. [Google Scholar] [CrossRef]
- Paillard, T. Effects of general and local fatigue on postural control: A review. Neurosci. Biobehav. Rev. 2012, 36, 162–176. [Google Scholar] [CrossRef]
- Anand, T.; Phani Kumar, G.; Pandareesh, M.D.; Swamy, M.S.; Khanum, F.; Bawa, A.S. Effect of bacoside extract from Bacopa monniera on physical fatigue induced by forced swimming. Phytother. Res. 2012, 26, 587–593. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Qu, Y.; Zhang, Y.; Li, S.; Sun, Y.; Chen, Z.; Teng, L.; Wang, D. Antifatigue potential activity of Sarcodon imbricatus in acute excise-treated and chronic fatigue syndrome in mice via regulation of Nrf2-mediated oxidative stress. Oxid. Med. Cell. Longev. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.N.; Liang, J.L.; Chen, H.B.; Liang, Y.E.; Guo, H.Z.; Su, Z.R.; Li, Y.C.; Zeng, H.F.; Zhang, X.J. Anti-fatigue and antioxidant activity of the polysaccharides isolated from Millettiae speciosae Champ. Leguminosae. Nutrients 2015, 7, 8657–8669. [Google Scholar] [CrossRef] [PubMed]
- Robergs, R.A.; Ghiasvand, F.; Parker, D. Biochemistry of exercise-induced metabolic acidosis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, 502–516. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.J.; Huang, W.C.; Chiu, C.C.; Liu, Y.L.; Chiu, W.C.; Chiu, C.H.; Chiu, Y.S.; Huang, C.C. Capsaicin supplementation reduces physical fatigue and improves exercise performance in mice. Nutrients 2016, 8, 648. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.D.; Chiu, C.H.; Hsu, Y.J.; Hou, C.W.; Chen, Y.M.; Huang, C.C. Changbai mountain Ginseng (Panax ginseng C.A. Mey) extract supplementation improves exercise performance and energy utilization and decreases fatigue-associated parameters in mice. Molecules 2017, 22, 237. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Huang, W.C.; Liu, C.C.; Wang, M.F.; Ho, C.S.; Huang, W.P.; Hou, C.C.; Chuang, H.L.; Huang, C.C. Pumpkin (Cucurbita moschata) fruit extract improves physical fatigue and exercise performance in mice. Molecules 2012, 17, 11864–11876. [Google Scholar] [CrossRef]
- Armstrong, C.W.; McGregor, N.R.; Butt, H.L.; Gooley, P.R. Metabolism in chronic fatigue syndrome. Adv. Clin. Chem. 2014, 66, 121–172. [Google Scholar]
- Mota, M.R.; Dantas, R.A.E.; Oliveira-Silva, I.; Sales, M.M.; Sotero, R.D.C.; Venancio, P.E.M.; Teixeira Junior, J.; Chaves, S.N.; de Lima, F.D. Effect of self-paced active recovery and passive recovery on blood lactate removal following a 200 m freestyle swimming trial. J. Sports Med. 2017, 8, 155–160. [Google Scholar] [CrossRef]
- Kent, J.A.; Ortenblad, N.; Hogan, M.C.; Poole, D.C.; Musch, T.I. No muscle is an island: Integrative perspectives on muscle fatigue. Med. Sci. Sports Exerc. 2016, 48, 2281–2293. [Google Scholar] [CrossRef] [PubMed]
- Osman, W.N.W.; Mohamed, S. Standardized Morinda citrifolia L. and Morinda elliptica L. leaf extracts alleviated fatigue by improving glycogen storage and lipid/carbohydrate metabolism. Phytother. Res. 2018, 32, 2078–2085. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.J.; Li, N.; Zhou, F.; Dong, Q.T.; Zhang, X.D.; Chen, B.C.; Yu, Z. Ginsenoside Rb1 improves energy metabolism in the skeletal muscle of an animal model of postoperative fatigue syndrome. J. Surg. Res. 2014, 191, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Manoharan, P.; Radzyukevich, T.L.; Hakim Javadi, H.; Stiner, C.A.; Landero Figueroa, J.A.; Lingrel, J.B.; Heiny, J.A. Phospholemman is not required for the acute stimulation of Na(+)-K(+)-ATPase alpha(2)-activity during skeletal muscle fatigue. Am. J. Physiol. Cell Physiol. 2015, 309, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.M.; Sun, Y.Y.; Zhou, T.T.; Zhu, Z.Y.; Zhuang, J.J.; Tang, X.; Chen, J.; Hu, L.H.; Shen, X. Arctigenin enhances swimming endurance of sedentary rats partially by regulation of antioxidant pathways. Acta Pharmacol. Sin. 2014, 35, 1274–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamanian, M.; Hajizadeh, M.R.; Esmaeili Nadimi, A.; Shamsizadeh, A.; Allahtavakoli, M. Antifatigue effects of troxerutin on exercise endurance capacity, oxidative stress and matrix metalloproteinase-9 levels in trained male rats. Fundam. Clin. Pharmacol. 2017, 31, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Ostojic, S.M. Exercise-induced mitochondrial dysfunction: A myth or reality? Clin. Sci. 2016, 130, 1407–1416. [Google Scholar] [CrossRef] [PubMed]
- Milone, M.; Wong, L.J. Diagnosis of mitochondrial myopathies. Mol. Genet. Metab. 2013, 110, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Vincent, G.; Lamon, S.; Gant, N.; Vincent, P.J.; MacDonald, J.R.; Markworth, J.F.; Edge, J.A.; Hickey, A.J. Changes in mitochondrial function and mitochondria associated protein expression in response to 2-weeks of high intensity interval training. Front. Physiol. 2015, 6, 51. [Google Scholar] [CrossRef] [PubMed]
- Meeus, M.; Nijs, J.; Hermans, L.; Goubert, D.; Calders, P. The role of mitochondrial dysfunctions due to oxidative and nitrosative stress in the chronic pain or chronic fatigue syndromes and fibromyalgia patients: Peripheral and central mechanisms as therapeutic targets? Expert Opin. Ther. Targets 2013, 17, 1081–1089. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, P.F.; Liu, S.F.; Hung, T.J.; Hung, C.Y.; Liu, G.Z.; Chuang, L.Y.; Chen, M.F.; Wang, J.L.; Shi, M.D.; Hsu, C.H.; et al. Elucidation of the therapeutic role of mitochondrial biogenesis transducers NRF-1 in the regulation of renal fibrosis. Exp. Cell Res. 2016, 349, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.K.; Chang Chien, K.M.; Chang, J.H.; Huang, M.H.; Liang, Y.C.; Liu, T.H. Branched-chain amino acids and arginine improve performance in two consecutive days of simulated handball games in male and female athletes: A randomized trial. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.F.; Wu, H.J.; Chen, C.Y.; Chou, K.M.; Chang, C.K. Branched-chain amino acids, arginine, citrulline alleviate central fatigue after 3 simulated matches in taekwondo athletes: A randomized controlled trial. J. Int. Soc. Sports Nutr. 2016, 13, 28. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Yu, K.Q.; Liu, Y.Y.; Ouyang, M.Z.; Yan, M.H.; Luo, R.; Zhao, X.S. Anti-fatigue activity of polysaccharides extract from Radix Rehmanniae Preparata. Int. J. Biol. Macromol. 2012, 50, 59–62. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Weight (g) | Vehicle | WOPs-LG | WOPs-MG | WOPs-HG |
|---|---|---|---|---|
| Set 1 | ||||
| Initial body weight | 24.62 ± 2.04 | 24.88 ± 1.64 | 25.47 ± 1.98 | 24.73 ± 1.23 |
| Final body weight | 38.69 ± 3.26 | 38.75 ± 2.57 | 38.37 ± 3.19 | 39.65 ± 2.82 |
| Set 2 | ||||
| Initial bogy weight | 25.50 ± 1.43 | 25.16 ± 1.42 | 24.98 ± 1.47 | 25.14 ± 1.48 |
| Final body weight | 39.01 ± 2.52 | 39.33 ± 1.41 | 39.94 ± 2.80 | 41.12 ± 3.13 |
| Set 3 | ||||
| Initial body weight | 26.40 ± 1.42 | 25.27 ± 1.94 | 25.19 ± 1.77 | 24.73 ± 1.77 |
| Final body weight | 39.02 ± 3.00 | 39.23 ± 2.17 | 38.58 ± 2.71 | 39.07 ± 3.21 |
| Set 4 | ||||
| Initial body weight | 25.88 ± 1.43 | 25.42 ± 1.56 | 25.08 ± 1.04 | 25.19 ± 1.57 |
| Final body weight | 39.03 ± 2.12 | 40.16 ± 2.82 | 39.40 ± 3.20 | 39.73 ± 3.27 |
| BLA (g/L) | Vehicle | WOPs-LG | WOPs-MG | WOPs-HG |
|---|---|---|---|---|
| Baseline | 6.04 ± 0.93 | 6.09 ± 0.79 | 6.37 ± 2.07 | 6.46 ± 1.77 |
| 0 min after swimming | 7.65 ± 1.23 | 7.40 ± 0.59 | 8.49 ± 1.08 | 6.55 ± 1.56 * |
| 20 min after swimming | 7.99 ± 0.92 | 5.10 ± 0.44 ** | 4.83 ± 0.37 ** | 4.63 ± 0.63 ** |
| Area under BLA curve | 224.89 ± 21.48 | 192.43 ± 7.91 ** | 206.89 ± 19.98 | 173.66 ± 14.99 ** |
| Groups | SOD (U/L) | GPX (U/L) | MDA (nmol/L) |
|---|---|---|---|
| Vehicle group | 129.97 ± 12.78 | 564.79 ± 48.33 | 3.42 ± 0.32 |
| WOPs-LG | 139.29 ± 13.95 | 530.85 ± 65.48 | 3.25 ± 0.28 |
| WOPs-MG | 177.23 ± 19.14 ** | 661.47 ± 45.13 ** | 2.11 ± 0.39 ** |
| WOPs-HG | 185.75 ± 15.13 ** | 685.86 ± 57.63 ** | 2.34 ± 0.47 ** |
| Amino Acid | Amino Acid Composition of WOPs (g/100 g) |
|---|---|
| Asp | 0.09 |
| Glu | 0.12 |
| Ser | 0.11 |
| His | 0.02 |
| Gly | 0.06 |
| Thr | 0.15 |
| Arg | 1.02 |
| Ala | 0.13 |
| Tyr | 0.30 |
| Cys | 0.01 |
| Val | 0.07 |
| Met | 0.02 |
| Phe | 0.48 |
| Ile | 0.04 |
| Leu | 0.28 |
| Lys | 0.05 |
| Pro | 0.06 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, R.; Wu, L.; Du, Q.; Ren, J.-W.; Chen, Q.-H.; Li, D.; Mao, R.-X.; Liu, X.-R.; Li, Y. Small Molecule Oligopeptides Isolated from Walnut (Juglans regia L.) and Their Anti-Fatigue Effects in Mice. Molecules 2019, 24, 45. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24010045
Liu R, Wu L, Du Q, Ren J-W, Chen Q-H, Li D, Mao R-X, Liu X-R, Li Y. Small Molecule Oligopeptides Isolated from Walnut (Juglans regia L.) and Their Anti-Fatigue Effects in Mice. Molecules. 2019; 24(1):45. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24010045
Chicago/Turabian StyleLiu, Rui, Lan Wu, Qian Du, Jin-Wei Ren, Qi-He Chen, Di Li, Rui-Xue Mao, Xin-Ran Liu, and Yong Li. 2019. "Small Molecule Oligopeptides Isolated from Walnut (Juglans regia L.) and Their Anti-Fatigue Effects in Mice" Molecules 24, no. 1: 45. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24010045