
Production of Added-Value Chemical Compounds through Bioconversions of Olive-Mill Wastewaters Blended with Crude Glycerol by a Yarrowia lipolytica Strain

 ,
,
Abstract
:1. Introduction
2. Results
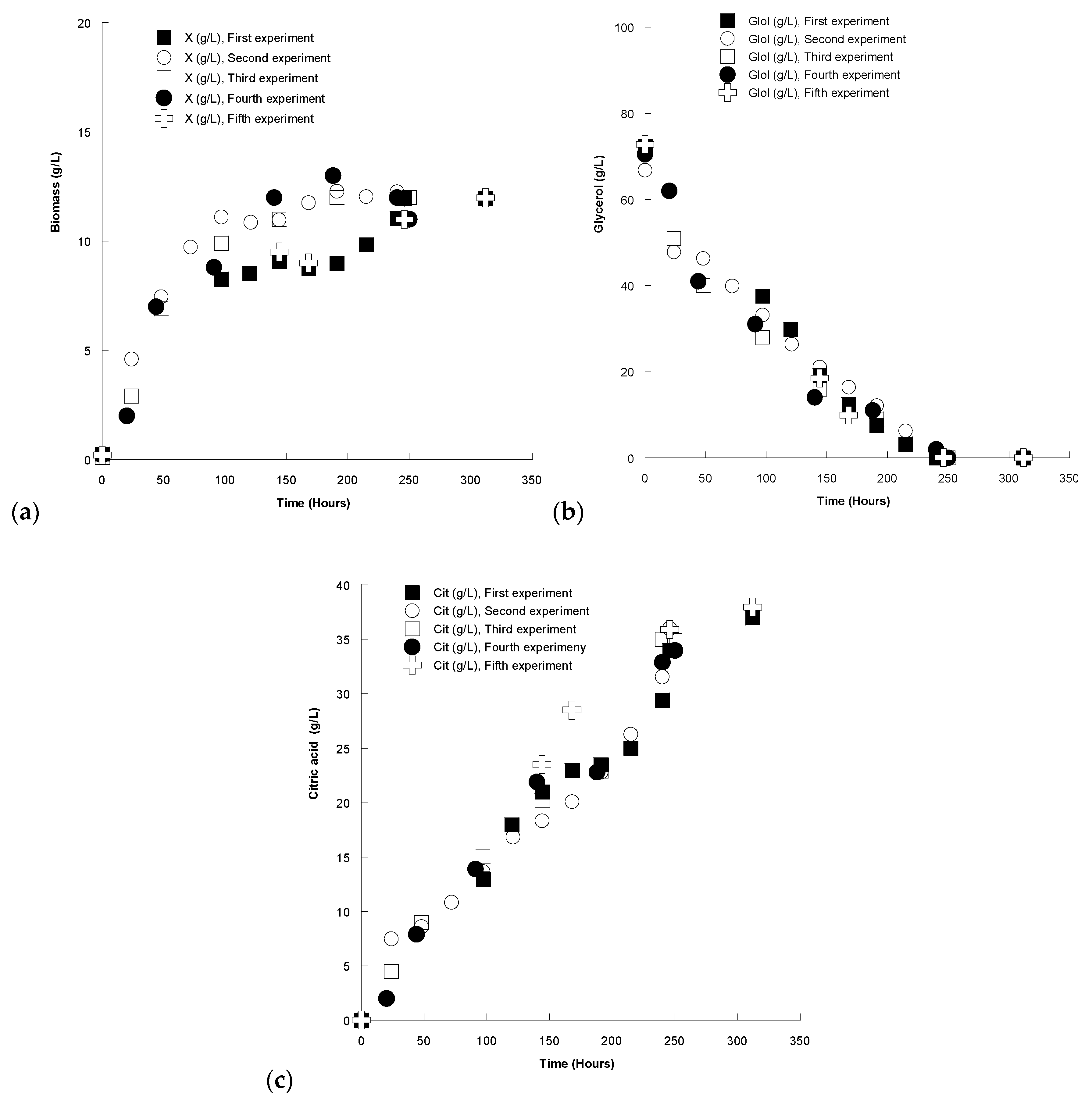
2.1. Growth of Y. lipolytica Strain ACA-DC 5029 in Media Containing Blends of OMW and Crude Glycerol with Initial Concentration of ~70 g/L
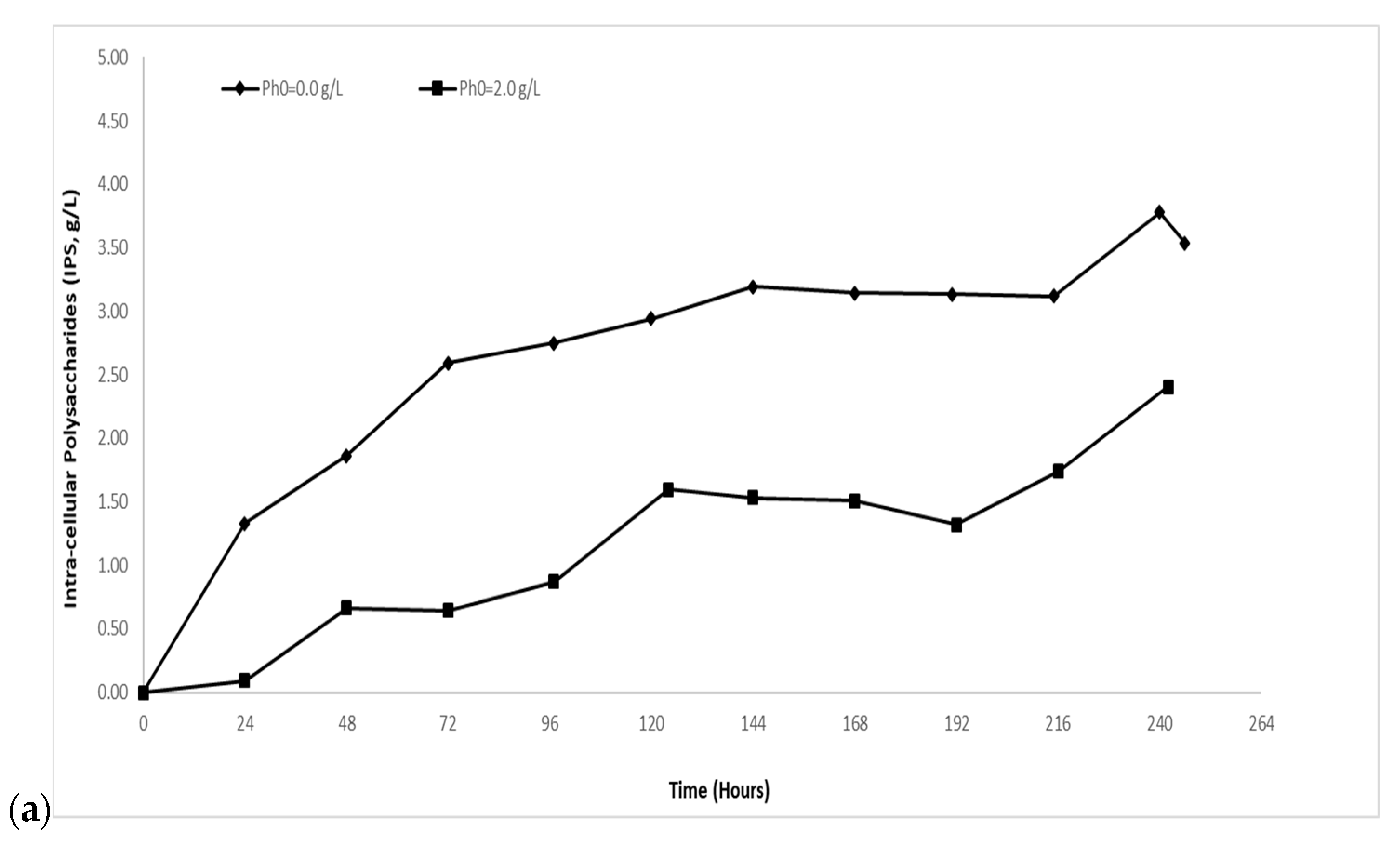
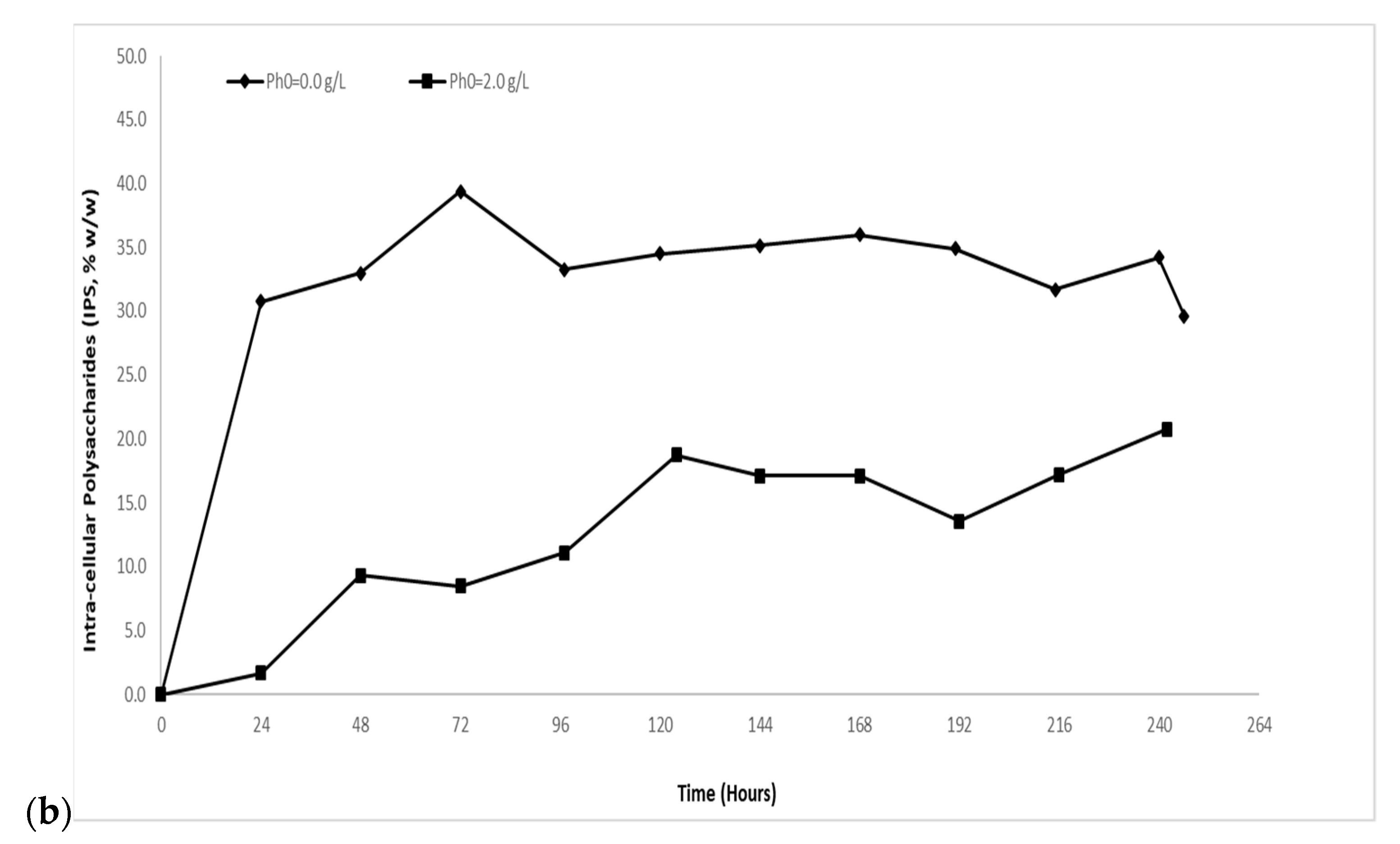
2.1.1. Biomass and Intra-Cellular Polysaccharides (IPS) Evolution
2.1.2. Citric Acid and Polyols Production
2.1.3. Biogenesis of Cellular lipids and Fatty Acid (FA) Composition
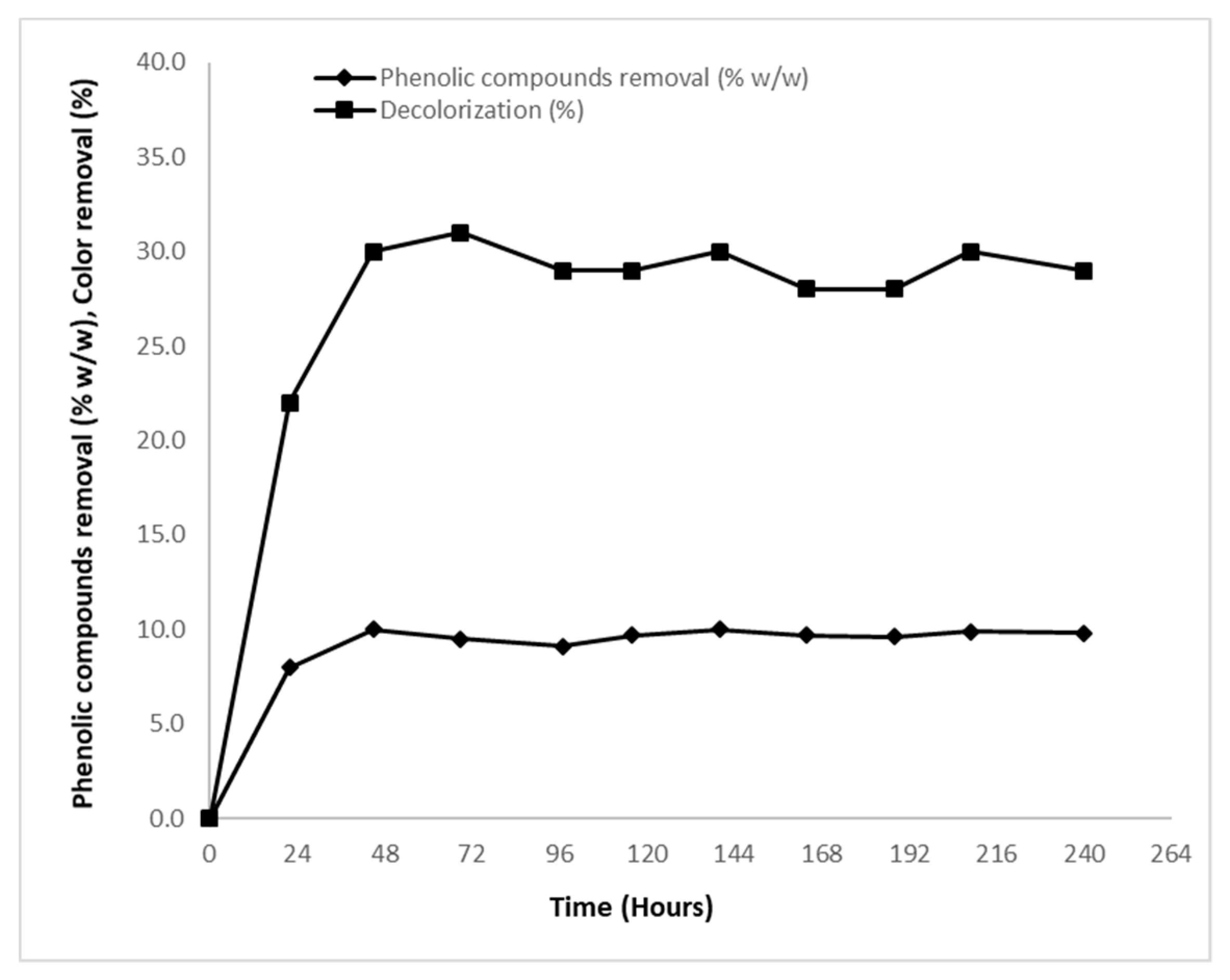
2.1.4. Decolorization—Removal of Phenolic Compounds
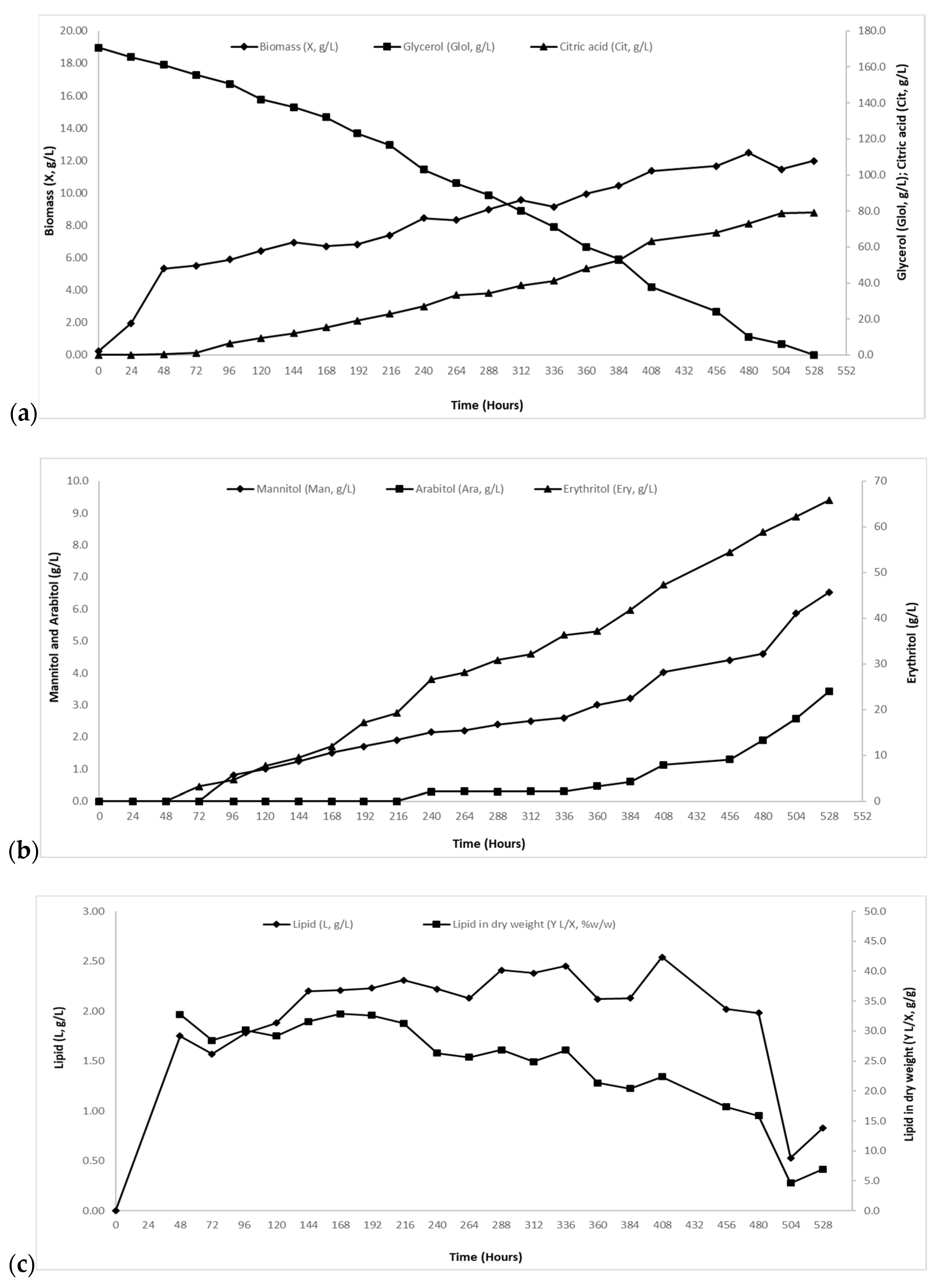
2.2. Growth of Y. lipolytica Strain ACA-DC 5029 in Media with Initial Crude Glycerol Concentration of ~120 g/L and of ~170 g/L.
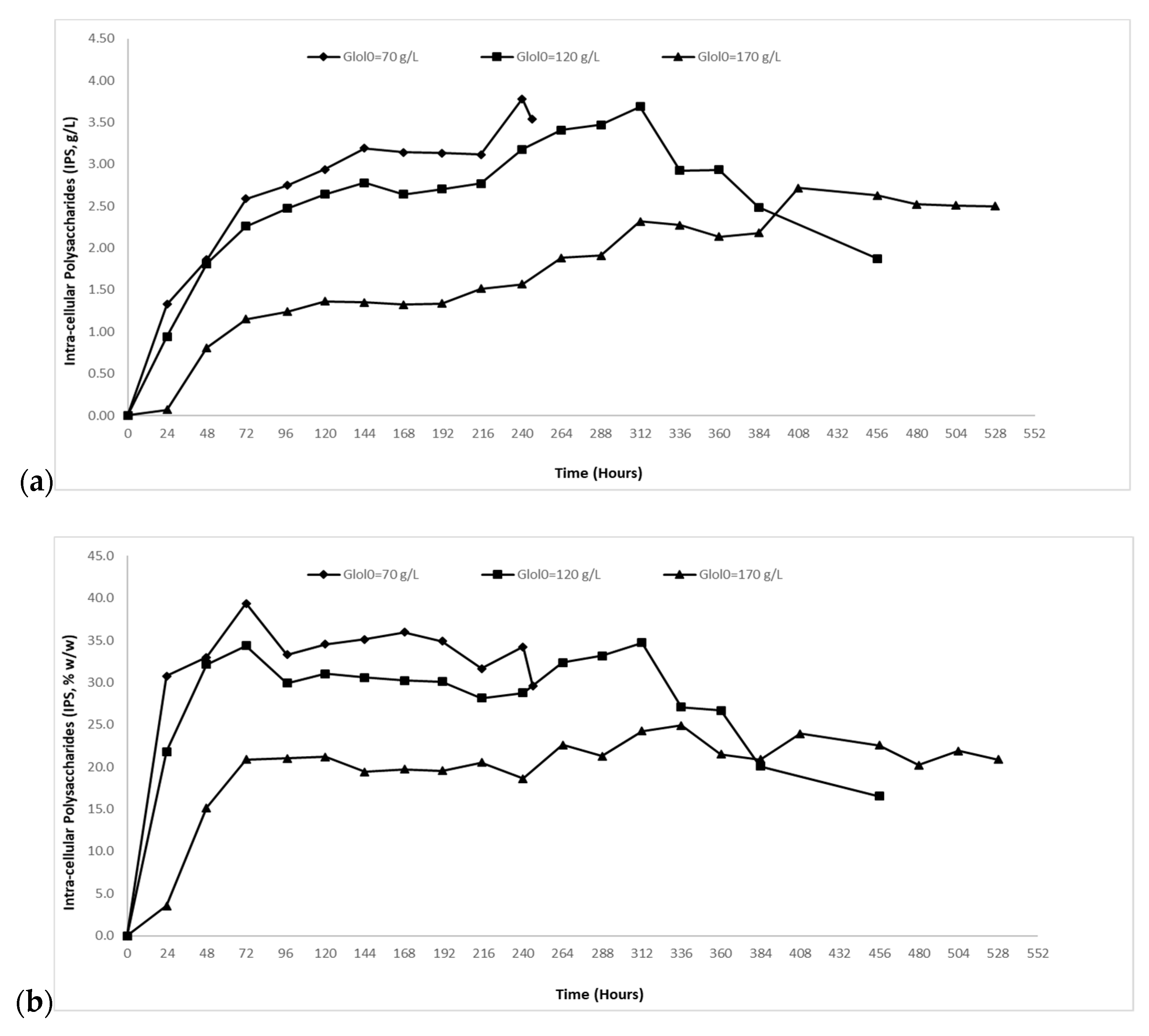
2.2.1. Biomass and Intra-Cellular Polysaccharides (IPS) Evolution
2.2.2. Citric Acid and Polyols Production
2.2.3. Biogenesis of Cellular Lipids and Fatty Acid (FA) Composition
3. Discussion
4. Materials and Methods
4.1. Microorganism and Growth Media
4.2. Culture Conditions
4.3. Analytical Methods
4.4. Repeatability—Statistical Treatment
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Nomenclature
| OMW | Olive mill wastewaters |
| X (g/L) | Biomass |
| Glol (g/L) | Glycerol |
| Ara (g/L) | Arabitol |
| Ery (g/L) | Erythritol |
| Man (g/L) | Mannitol |
| Cit (g/L) | Citric acid |
| L (g/L) | Total lipid |
| Ph (g/L) | Phenolic compounds |
| YX/Glol (g formed per g of glycerol consumed) | Biomass yield on glycerol consumed |
| YAra/Glol (g formed per g of glycerol consumed) | Arabitol yield on glycerol consumed |
| YEry/Glol (g formed per g of glycerol consumed) | Erythritol yield on glycerol consumed |
| YMan/Glol (g formed per g of glycerol consumed) | Mannitol yield on glycerol consumed |
| YCit/Glol (g formed per g of glycerol consumed) | Citric acid yield on glycerol consumed |
| YL/X (% w/w) | Total lipid on biomass |
| UI | Unsaturation index |
| FA | Fatty acid |
| IPS (g formed per g of biomass formed; % w/w) | Endopolysaccharides |
| Subscripts 0, cons and max indicate the initial, consumed and maximum quantity of the components in the kinetics performed | |
References
- Lanciotti, R.; Gianotti, A.; Baldi, D.; Angrisani, R.; Suzzi, G.; Mastrocola, D.; Guerzoni, M.E. Use of Yarrowia lipolytica strains for the treatment of olive mill wastewater. Bioresour. Technol. 2005, 96, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Madrona, A.; Pereira-Caro, G.; Mateos, R.; Rodríguez, G.; Trujillo, M.; Fernández-Bolaños, J.; Espartero, J.L. Synthesis of hydroxytyrosyl alkyl ethers from olive oil waste waters. Molecules 2009, 14, 1762–1772. [Google Scholar] [CrossRef]
- Ergül, F.E.; Sargın, S.; Öngen, G.; Sukan, F.V. Dephenolization and decolorization of olive mill wastewater through sequential batch and co-culture applications. World J. Microbiol. Biotechnol. 2011, 27, 107–114. [Google Scholar] [CrossRef]
- Aggelis, G.; Iconomou, D.; Christou, M.; Bokas, D.; Kotzailias, S.; Christou, G.; Tsagou, V.; Papanikolaou, S. Phenolic removal in a model olive oil mill wastewater using Pleurotus ostreatus in bioreactor cultures and biological evaluation of the process. Water Res. 2003, 37, 3897–3904. [Google Scholar] [CrossRef]
- Tsioulpas, A.; Dimou, D.; Iconomou, D.; Aggelis, G. Phenolic removal in olive oil mill wastewater by strains of Pleurotus spp. in respect to their phenol oxidase (laccase) activity. Bioresour. Technol. 2002, 84, 251–257. [Google Scholar]
- Sayadi, S.; Ellouz, R. Decolourization of olive mill waste-waters by the white-rot fungus Phanerochaete chrysosporium: Involvement of the lignin-degrading system. Appl. Microbiol. Biotechnol. 1992, 37, 813–817. [Google Scholar] [CrossRef]
- Massadeh, M.; Modallal, N. Ethanol production from olive mill wastewater (OMW) pretreated with Pleurotus sajor-caju. Energy Fuels 2007, 22, 150–154. [Google Scholar] [CrossRef]
- Bellou, S.; Makri, A.; Sarris, D.; Michos, K.; Rentoumi, P.; Celik, A.; Papanikolaou, S.; Aggelis, G. The olive mill wastewater as substrate for single cell oil production by Zygomycetes. J. Biotechnol. 2014, 170, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Ginos, A.; Manios, T.; Mantzavinos, D. Treatment of olive mill effluents by coagulation-flocculation-hydrogen peroxide oxidation and effect on phytotoxicity. J. Hazard. Mater. 2006, 133, 135–142. [Google Scholar] [CrossRef]
- Mantzavinos, D.; Kalogerakis, N. Treatment of olive mill effluents, Part I organic matter degradation by chemical and biological processes-an overview. Environ. Int. 2005, 31, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Sarika, R.; Kalogerakis, N.; Mantzavinos, D. Treatment of olive mill effluents Part II. Complete removal of solids by direct flocculation with poly-electrolytes. Environ. Int. 2005, 31, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Crognale, S.; D’ Annibale, A.; Federici, F.; Fenice, M.; Quaratino, D.; Petruccioli, M. Olive oil mill wastewater valorization by fungi. J. Chem. Technol. Biotechnol. 2006, 81, 1547–1555. [Google Scholar] [CrossRef]
- Sarris, D.; Galiotou-Panayotou, M.; Koutinas, A.A.; Komaitis, M.; Papanikolaou, S. Citric acid, biomass and cellular lipid production by Yarrowia lipolytica strains cultivated on olive mill wastewater-based media. J. Chem. Technol. Biotechnol. 2011, 86, 1439–1448. [Google Scholar]
- Ben Sassi, A.; Ouazzani, N.; Walker, G.M.; Ibnsouda, S.; El Mzibri, M.; Boussaid, A. Detoxification of olive mill wastewaters by Moroccan yeast isolates. Biodegrad. 2008, 19, 337–346. [Google Scholar]
- Scioli, C.; Vollaro, L. The use of Yarrowia lipolytica to reduce pollution in olive mill wastewaters. Water Res. 1997, 31, 2520–2524. [Google Scholar]
- Papanikolaou, S.; Galiotou-Panayotou, M.; Fakas, S.; Komaitis, M.; Aggelis, G. Citric acid production by Yarrowia lipolytica cultivated on olive-mill wastewater-based media. Bioresour. Technol. 2008, 99, 2419–2428. [Google Scholar] [CrossRef] [PubMed]
- Angenent, L.T.; Karim, K.; Al-Dahhan, M.H.; Wrenn, B.A.; Domíguez-Espinosa, R. Production of bioenergy and biochemicals from industrial and agricultural wastewater. Trends Biotechnol. 2004, 22, 477–485. [Google Scholar] [CrossRef]
- Dionisi, D.; Carucci, G.; Papini, M.P.; Riccardi, C.; Majone, M.; Carrasco, F. Olive oil mill effluents as a feedstock for production of biodegradable polymers. Water Res. 2005, 39, 2076–2084. [Google Scholar] [CrossRef]
- Sarris, D.; Matsakas, L.; Aggelis, G.; Koutinas, A.A.; Papanikolaou, S. Aerated vs non-aerated conversions of molasses and olive mill wastewaters blends into bioethanol by Saccharomyces cerevisiae under non-aseptic conditions. Ind. Crops. Prod. 2014, 56, 83–93. [Google Scholar]
- Sarris, D.; Giannakis, M.; Philippoussis, A.; Komaitis, M.; Koutinas, A.A.; Papanikolaou, S. Conversions of olive mill wastewater-based media by Saccharomyces cerevisiae through sterile and non-sterile bioprocesses. J. Chem. Technol. Biotechnol. 2013, 88, 958–969. [Google Scholar]
- Zanichelli, D.; Carloni, F.; Hasanaj, E.; D’Andrea, N.; Filippini, A.; Setti, L. Production of ethanol by an integrated valorization of olive oil byproducts. The role of phenolic inhibition. Env. Sci. Pollut. Res. 2007, 14, 5–6. [Google Scholar]
- Bernal, J.M.; Lozano, P.; García-Verdugo, E.; Burguete, M.I.; Sánchez-Gómez, G.; López-López, G.; Pucheault, M.; Vaultier, M.; Luis, S.V. Supercritical synthesis of biodiesel. Molecules 2012, 17, 8696–8719. [Google Scholar] [CrossRef] [PubMed]
- André, A.; Chatzifragkou, A.; Diamantopoulou, P.; Sarris, D.; Philippoussis, A.; Galiotou-Panayotou, M.; Komaitis, M.; Papanikolaou, S. Biotechnological conversions of bio-diesel-derived crude glycerol by Yarrowia lipolytica strains. Eng. Life Sci. 2009, 9, 468–478. [Google Scholar] [CrossRef]
- André, A.; Diamantopoulou, P.; Philippoussis, A.; Sarris, D.; Komaitis, M.; Papanikolaou, S. Biotechnological conversions of bio-diesel derived waste glycerol into added-value compounds by higher fungi: Production of biomass, single cell oil and oxalic acid. Ind. Crops Prod. 2010, 31, 407–416. [Google Scholar] [CrossRef]
- Juszczyk, P.; Tomaszewska, L.; Kita, A.; Rymowicz, W. Biomass production by novel strains of Yarrowia lipolytica using raw glycerol, derived from biodiesel production. Bioresour. Technol. 2013, 137, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Rywińska, A.; Juszczyk, P.; Wojtatowicz, M.; Robak, M.; Lazar, Z.; Tomaszewska, L.; Rymowicz, W. Glycerol as a promising substrate for Yarrowia lipolytica biotechnological applications. Biomass Bioenergy 2013, 48, 148–166. [Google Scholar] [CrossRef]
- Garlapati, V.K.; Shankar, U.; Budhiraja, A. Bioconversion technologies of crude glycerol to value added industrial products. Biotechnol. Rep. 2016, 9, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Morgunov, I.G.; Kamzolova, S.V.; Lunina, J.N. The citric acid production from raw glycerol by Yarrowia lipolytica yeast and its regulation. Appl. Microbiol. Biotechnol. 2013, 97, 7387–7397. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Biotechnological valorization of biodiesel derived glycerol waste through production of single cell oil and citric acid by Yarrowia lipolytica. Lipid Technol. 2009, 21, 83–87. [Google Scholar] [CrossRef]
- Yazdani, S.S.; Gonzalez, R. Anaerobic fermentation of glycerol: A path to economic viability for the biofuels industry. Curr. Opin. Biotechnol. 2007, 18, 213–219. [Google Scholar] [CrossRef]
- Santos, E.O.; Michelon, M.; Gallas, J.A.; Kalil, S.J.; Burkert, C.A.V. Raw glycerol as substrate for the production of yeast biomass. Int. J. Food Eng. 2013, 9, 413–420. [Google Scholar] [CrossRef]
- Papanikolaou, S. 1,3-Propanediol and citric acid production from glycerol containing waste discharged after bio-diesel manufacturing process. In Current Topics on Bioprocesses in Food Industry; Koutinas, A., Pandey, A., Christian, L., Eds.; AsiaTech Publishers Inc.: New Delhi, India, 2008; Volume 2, pp. 381–399. [Google Scholar]
- Dourou, M.; Kancelista, A.; Juszczyk, P.; Sarris, D.; Bellou, S.; Triantaphyllidou, I.-E.; Rywinska, A.; Papanikolaou, S.; Aggelis, G. Bioconversion of olive mill wastewater into high-added value products. J. Clean. Prod. 2016, 139, 957–969. [Google Scholar] [CrossRef]
- Sarris, D.; Stoforos, N.G.; Mallouchos, A.; Kookos, I.K.; Koutinas, A.A.; Aggelis, G.; Papanikolaou, S. Production of added-value metabolites by Yarrowia lipolytica growing in olive mill wastewater-based media under aseptic and non-aseptic conditions. Eng. Life Sci. 2017, 17, 695–709. [Google Scholar] [CrossRef]
- Morgunov, I.G.; Kamzolova, S.V.; Lunina, J.N. Citric Acid Production by Yarrowia lipolytica Yeast on Different Renewable Raw Materials. Ferment. 2018, 4, 36. [Google Scholar] [CrossRef]
- Soccol, C.R.; Vandenberghe, L.P.; Rodrigues, C.; Pandey, A. New perspectives for citric acid production and application. Food Technol. Biotechnol. 2006, 44, 141–149. [Google Scholar]
- Rakicka, M.; Rywińska, A.; Cybulski, K.; Rymowicz, W. Enhanced production of erythritol and mannitol by Yarrowia lipolytica in media containing surfactants. Braz. J. Microbiol. 2016, 47, 417–423. [Google Scholar] [CrossRef]
- Rakicka, M.; Rukowicz, B.; Rywińska, A.; Lazar, Z.; Rymowicz, W. Technology of efficient continuous erythritol production from glycerol. J. Cleaner Prod. 2016, 139, 905–913. [Google Scholar] [CrossRef]
- Park, Y.-C.; Oh, E.J.; Jo, J.-H.; Jin, Y.-S.; Seo, J.-H. Recent advances in biological production of sugar alcohols. Curr. Opin. Biotechnol. 2016, 37, 105–113. [Google Scholar] [CrossRef]
- Tomaszewska, L.; Rakicka, M.; Rymowicz, W.; Rywińska, A. A comparative study on glycerol metabolism to erythritol and citric acid in Yarrowia lipolytica yeast cells. FEMS Yeast Res. 2014, 14, 966–976. [Google Scholar] [CrossRef]
- Rywińska, A.; Tomaszewska, L.; Rymowicz, W. Erythritol biosynthesis by Yarrowia lipolytica yeast under various culture conditions. Afr. J. Microbiol. Res. 2013, 7, 3511–3516. [Google Scholar]
- Almeida, J.R.; Fávaro, L.C.; Quirino, B.F. Biodiesel biorefinery: Opportunities and challenges for microbial production of fuels and chemicals from glycerol waste. Biotechnol. Biofuels 2012, 5, 48. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Aggelis, G. Yarrowia lipolytica: A model microorganism used for the production of tailor-made lipids. Eur. J. Lipid Sci. Technol. 2010, 112, 639–654. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Modeling lipid accumulation and degradation in Yarrowia lipolytica cultivated on industrial fats. Curr. Microbiol. 2003, 46, 0398–0402. [Google Scholar] [CrossRef] [PubMed]
- Athenaki, M.; Gardeli, C.; Diamantopoulou, P.; Tchakouteu, S.S.; Sarris, D.; Philippoussis, A.; Papanikolaou, S. Lipids from yeasts and fungi: Physiology, production and analytical considerations. J. Appl. Microbiol. 2018, 124, 336–367. [Google Scholar] [CrossRef] [PubMed]
- Rzechonek, D.A.; Dobrowolski, A.; Rymowicz, W.; Mirończuk, A.M. Recent advances in biological production of erythritol. Crit. Rev. Biotechnol. 2018, 38, 620–633. [Google Scholar] [CrossRef] [PubMed]
- Carsanba, E.; Papanikolaou, S.; Erten, H. Production of oils and fats by oleaginous microorganisms with an emphasis given to the potential of the nonconventional yeast Yarrowia lipolytica. Crit. Rev. Biotechnol. 2018, 38, 1230–1243. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, M.; Ventrice, D.; Varone, M.; Sidari, R.; Caridi, A. HPLC determination of phenolics adsorbed on yeasts. J. Pharm. Biomed. Anal. 2006, 42, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Chtourou, M.; Ammar, E.; Nasri, M.; Medhioub, K. Isolation of a yeast, Trichosporon cutaneum, able to use low molecular weight phenolic compounds: Application to olive mill waste water treatment. J. Chem. Technol. Biotechnol. 2004, 79, 869–878. [Google Scholar] [CrossRef]
- Koganti, S.; Kuo, T.M.; Kurtzman, C.P.; Smith, N.; Ju, L.-K. Production of arabitol from glycerol: Strain screening and study of factors affecting production yield. Appl. Microbiol. Biotechnol. 2011, 90, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Fakas, S.; Fick, M.; Chevalot, I.; Galiotou-Panayotou, M.; Komaitis, M.; Marc, I.; Aggelis, G. Biotechnological valorisation of raw glycerol discharged after bio-diesel (fatty acid methyl esters) manufacturing process: Production of 1, 3-propanediol, citric acid and single cell oil. Biomass Bioenergy 2008, 32, 60–71. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Gortzi, O.; Margeli, E.; Chinou, I.; Galiotou-Panayotou, M.; Lalas, S. Effect of Citrus essential oil addition upon growth and cellular lipids of Yarrowia lipolytica yeast. Eur. J. Lipid Sci. Technol. 2008, 110, 997–1006. [Google Scholar] [CrossRef]
- Kamzolova, S.V.; Morgunov, I.G.; Aurich, A.; Perevoznikova, O.A.; Shishkanova, N.V.; Stottmeister, U.; Finogenova, T.V. Lipase secretion and citric acid production in Yarrowia lipolytica yeast grown on animal and vegetable fat. Food Technol. Biotechnol. 2005, 43, 113–122. [Google Scholar]
- Anastassiadis, S.; Rehm, H.-J. Continuous citric acid secretion by a high specific pH dependent active transport system in yeast Candida oleophila ATCC 20177. Electron. J. Biotechnol. 2005, 8, 26–42. [Google Scholar] [CrossRef]
- Kamzolova, S.V.; Shishkanova, N.V.; Morgunov, I.G.; Finogenova, T.V. Oxygen requirements for growth and citric acid production of Yarrowia lipolytica. FEMS Yeast Res. 2003, 3, 217–222. [Google Scholar] [CrossRef]
- Aggelis, G.; Komaitis, M. Enhancement of single cell oil production by Yarrowia lipolytica growing in the presence of Teucrium polium L. aqueous extract. Biotechnol. Lett. 1999, 21, 747–749. [Google Scholar] [CrossRef]
- Chatzifragkou, A.; Petrou, I.; Gardeli, C.; Komaitis, M.; Papanikolaou, S. Effect of Origanum vulgare L. essential oil on growth and lipid profile of Yarrowia lipolytica cultivated on glycerol-based media. J. Am. Oil Chem. Soc. 2011, 88, 1955–1964. [Google Scholar] [CrossRef]
- D’Annibale, A.; Sermanni, G.G.; Federici, F.; Petruccioli, M. Olive-mill wastewaters: A promising substrate for microbial lipase production. Bioresour. Technol. 2006, 97, 1828–1833. [Google Scholar] [CrossRef] [PubMed]
- De Felice, B.; Pontecorvo, G.; Carfagna, M. Degradation of waste waters from olive oil mills by Yarrowia lipolytica ATCC 20255 and Pseudomonas putida. Acta Biotechnol. 1997, 17, 231–239. [Google Scholar] [CrossRef]
- Aissam, H.; Penninckx, M.J.; Benlemlih, M. Reduction of phenolics content and COD in olive oil mill wastewaters by indigenous yeasts and fungi. World J. Microbiol. Biotechnol. 2007, 23, 1203–1208. [Google Scholar] [CrossRef]
- Assas, N.; Ayed, L.; Marouani, L.; Hamdi, M. Decolorization of fresh and stored-black olive mill wastewaters by Geotrichum candidum. Process Biochem. 2002, 38, 361–365. [Google Scholar] [CrossRef]
- Garcıa, I.G.; Pena, P.J.; Venceslada, J.B.; Martın, A.M.; Santos, M.M.; Gomez, E.R. Removal of phenol compounds from olive mill wastewater using Phanerochaete chrysosporium, Aspergillus niger, Aspergillus terreus and Geotrichum candidum. Process Biochem. 2000, 35, 751–758. [Google Scholar] [CrossRef]
- Fountoulakis, M.S.; Dokianakis, S.N.; Kornaros, M.; Aggelis, G.; Lyberatos, G. Removal of phenolics in olive mill wastewaters using the white-rot fungus Pleurotus ostreatus. Water Res. 2002, 36, 4735–4744. [Google Scholar] [CrossRef]
- Zerva, A.; Zervakis, G.I.; Christakopoulos, P.; Topakas, E. Degradation of olive mill wastewater by the induced extracellular ligninolytic enzymes of two wood-rot fungi. J. Environ. Manag. 2017, 203, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Lakhtar, H.; Ismaili-Alaoui, M.; Philippoussis, A.; Perraud-Gaime, I.; Roussos, S. Screening of strains of Lentinula edodes grown on model olive mill wastewater in solid and liquid state culture for polyphenol biodegradation. Int. Biodeterior. Biodegrad. 2010, 64, 167–172. [Google Scholar] [CrossRef]
- Jolivalt, C.; Madzak, C.; Brault, A.; Caminade, E.; Malosse, C.; Mougin, C. Expression of laccase IIIb from the white-rot fungus Trametes versicolor in the yeast Yarrowia lipolytica for environmental applications. Appl. Microbiol. Biotechnol. 2005, 66, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.; Araújo, C.; Aguedo, M.; Gomes, N.; Gonçalves, C.; Teixeira, J.A.; Belo, I. The use of olive mill wastewater by wild type Yarrowia lipolytica strains: Medium supplementation and surfactant presence effect. J. Chem. Technol. Biotechnol. 2009, 84, 533–537. [Google Scholar] [CrossRef]
- Makri, A.; Fakas, S.; Aggelis, G. Metabolic activities of biotechnological interest in Yarrowia lipolytica grown on glycerol in repeated batch cultures. Bioresour. Technol. 2010, 101, 2351–2358. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Beopoulos, A.; Koletti, A.; Thevenieau, F.; Koutinas, A.A.; Nicaud, J.-M.; Aggelis, G. Importance of the methyl-citrate cycle on glycerol metabolism in the yeast Yarrowia lipolytica. J. Biotechnol. 2013, 168, 303–314. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Kampisopoulou, E.; Blanchard, F.; Rondags, E.; Gardeli, C.; Koutinas, A.A.; Chevalot, I.; Aggelis, G. Production of secondary metabolites through glycerol fermentation under carbon-excess conditions by the yeasts Yarrowia lipolytica and Rhodosporidium toruloides. Eur. J. Lipid Sci. Technol. 2017, 119. [Google Scholar] [CrossRef]
- Tchakouteu, S.S.; Kalantzi, O.; Gardeli, C.; Koutinas, A.A.; Aggelis, G.; Papanikolaou, S. Lipid production by yeasts growing on biodiesel-derived crude glycerol: Strain selection and impact of substrate concentration on the fermentation efficiency. J. Appl. Microbiol. 2015, 118, 911–927. [Google Scholar] [CrossRef] [PubMed]
- Rywińska, A.; Juszczyk, P.; Wojtatowicz, M.; Rymowicz, W. Chemostat study of citric acid production from glycerol by Yarrowia lipolytica. J. Biotechnol. 2011, 152, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Kamzolova, S.V.; Fatykhova, A.R.; Dedyukhina, E.G.; Anastassiadis, S.G.; Golovchenko, N.P.; Morgunov, I.G. Citric acid production by yeast grown on glycerol-containing waste from biodiesel industry. Food Technol. Biotechnol. 2011, 49, 65–74. [Google Scholar]
- Rywinska, A.; Rymowicz, W.; Marcinkiewicz, M. Valorization of raw glycerol for citric acid production by Yarrowia lipolytica yeast. Electron. J. Biotechnol. 2010, 13, 9–10. [Google Scholar] [CrossRef]
- Rywińska, A.; Rymowicz, W. High-yield production of citric acid by Yarrowia lipolytica on glycerol in repeated-batch bioreactors. J. Ind. Microbiol. Biotechnol. 2010, 37, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Rymowicz, W.; Fatykhova, A.R.; Kamzolova, S.V.; Rywińska, A.; Morgunov, I.G. Citric acid production from glycerol-containing waste of biodiesel industry by Yarrowia lipolytica in batch, repeated batch, and cell recycle regimes. Appl. Microbiol. Biotechnol. 2010, 87, 971–979. [Google Scholar] [CrossRef] [PubMed]
- Rymowicz, W.; Rywińska, A.; Żarowska, B.; Juszczyk, P. Citric acid production from raw glycerol by acetate mutants of Yarrowia lipolytica. Chem. Pap. 2006, 60, 391–394. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Muniglia, L.; Chevalot, I.; Aggelis, G.; Marc, I. Yarrowia lipolytica as a potential producer of citric acid from raw glycerol. J. Appl. Microbiol. 2002, 92, 737–744. [Google Scholar] [CrossRef]
- Rymowicz, W.; Rywińska, A.; Gładkowski, W. Simultaneous production of citric acid and erythritol from crude glycerol by Yarrowia lipolytica Wratislavia K1. Chem. Pap. 2008, 62, 239–246. [Google Scholar] [CrossRef]
- Rymowicz, W.; Rywińska, A.; Marcinkiewicz, M. High-yield production of erythritol from raw glycerol in fed-batch cultures of Yarrowia lipolytica. Biotechnol. Lett. 2009, 31, 377–380. [Google Scholar] [CrossRef]
- Chatzifragkou, A.; Makri, A.; Belka, A.; Bellou, S.; Mavrou, M.; Mastoridou, M.; Mystrioti, P.; Onjaro, G.; Aggelis, G.; Papanikolaou, S. Biotechnological conversions of biodiesel derived waste glycerol by yeast and fungal species. Energy 2011, 36, 1097–1108. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Rontou, M.; Belka, A.; Athenaki, M.; Gardeli, C.; Mallouchos, A.; Kalantzi, O.; Koutinas, A.A.; Kookos, I.K.; Zeng, A.P. Conversion of biodiesel-derived glycerol into biotechnological products of industrial significance by yeast and fungal strains. Eng. Life Sci. 2017, 17, 262–281. [Google Scholar] [CrossRef]
- Mirończuk, A.M.; Furgała, J.; Rakicka, M.; Rymowicz, W. Enhanced production of erythritol by Yarrowia lipolytica on glycerol in repeated batch cultures. J. Ind. Microbiol. Biotechnol. 2014, 41, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Tomaszewska, L.; Rywińska, A.; Gładkowski, W. Production of erythritol and mannitol by Yarrowia lipolytica yeast in media containing glycerol. J. Ind. Microbiol. Biotechnol. 2012, 39, 1333–1343. [Google Scholar] [CrossRef] [PubMed]
- Beopoulos, A.; Cescut, J.; Haddouche, R.; Uribelarrea, J.-L.; Molina-Jouve, C.; Nicaud, J.-M. Yarrowia lipolytica as a model for bio-oil production. Prog. Lipid Res. 2009, 48, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Aggelis, G. Lipids of oleaginous yeasts. Part I: Biochemistry of single cell oil production. Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Lipids of oleaginous yeasts. Part II: Technology and potential applications. Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073. [Google Scholar] [CrossRef]
- Beopoulos, A.; Chardot, T.; Nicaud, J.-M. Yarrowia lipolytica: A model and a tool to understand the mechanisms implicated in lipid accumulation. Biochimie 2009, 91, 692–696. [Google Scholar] [CrossRef]
- Tsigie, Y.A.; Wang, C.-Y.; Kasim, N.S.; Diem, Q.-D.; Huynh, L.-H.; Ho, Q.-P.; Truong, C.-T.; Ju, Y.-H. Oil production from Yarrowia lipolytica Po1g using rice bran hydrolysate. J. Biomed. Biotechnol. 2012, 2012, 378–384. [Google Scholar] [CrossRef]
- Tsigie, Y.A.; Wang, C.-Y.; Truong, C.-T.; Ju, Y.-H. Lipid production from Yarrowia lipolytica Po1g grown in sugarcane bagasse hydrolysate. Bioresour. Technol. 2011, 102, 9216–9222. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Lipid production by Yarrowia lipolytica growing on industrial glycerol in a single-stage continuous culture. Bioresour. Technol. 2002, 82, 43–49. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Chatzifragkou, A.; Fakas, S.; Galiotou-Panayotou, M.; Komaitis, M.; Nicaud, J.M.; Aggelis, G. Biosynthesis of lipids and organic acids by Yarrowia lipolytica strains cultivated on glucose. Eur. J. Lipid Sci. Technol. 2009, 111, 1221–1232. [Google Scholar] [CrossRef]
- Dobrowolski, A.; Mituła, P.; Rymowicz, W.; Mirończuk, A.M. Efficient conversion of crude glycerol from various industrial wastes into single cell oil by yeast Yarrowia lipolytica. Bioresour. Technol. 2016, 207, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Kamzolova, S.V.; Morgunov, I.G. Biosynthesis of pyruvic acid from glycerol-containing substrates and its regulation in the yeast Yarrowia lipolytica. Bioresour. Technol. 2018, 266, 125–133. [Google Scholar] [CrossRef]
- Finogenova, T.; Kamzolova, S.; Dedyukhina, E.; Shishkanova, N.; Il’chenko, A.; Morgunov, I.; Chernyavskaya, O.; Sokolov, A. Biosynthesis of citric and isocitric acids from ethanol by mutant Yarrowia lipolytica N 1 under continuous cultivation. Appl. Microbiol. Biotechnol. 2002, 59, 493–500. [Google Scholar] [PubMed]
- Fakas, S.; Papanikolaou, S.; Galiotou-Panayotou, M.; Komaitis, M.; Aggelis, G. Lipids of Cunninghamella echinulata with emphasis to γ-linolenic acid distribution among lipid classes. Appl. Microbiol. Biotechnol. 2006, 73, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Komaitis, M.; Aggelis, G. Single cell oil (SCO) production by Mortierella isabellina grown on high-sugar content media. Bioresour. Technol. 2004, 95, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Tsakona, S.; Kopsahelis, N.; Chatzifragkou, A.; Papanikolaou, S.; Kookos, I.K.; Koutinas, A.A. Formulation of fermentation media from flour-rich waste streams for microbial lipid production by Lipomyces starkeyi. J. Biotechnol. 2014, 189, 36–45. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Initial Phenolics (g/L) | Hours | X (g/L) | L (g/L) | Glolcons (g/L) | Cit (g/L) | Man (g/L) | Ara (g/L) | Ery (g/L) | YX/Glol (g/g) | YL/X % (w/w) | YCit/Glol (g/g) | YMan/Glol (g/g) | YAra/Glol (g/g) | YEry/Glol (g/g) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0.0 | 191 f | 8.98 | 0.60 | 64.9 | 17.3 | 9.9 | 3.1 | 14.9 | 0.14 | 6.7 | 0.27 | 0.15 | 0.05 | 0.23 |
| 240 d | 11.04 | 0.76 | 72.4 | 29.4 | 10.1 | 2.9 | 14.5 | 0.15 | 6.9 | 0.41 | 0.14 | 0.04 | 0.20 | |
| 246 a,b,e | 11.96 | 1.27 | 72.4 | 34.0 | 8.1 | 3.2 | 9.1 | 0.17 | 10.6 | 0.47 | 0.11 | 0.04 | 0.13 | |
| 312 c | 11.95 | 0.62 | 72.4 | 42.5 | 6.8 | 3.0 | 4.1 | 0.17 | 5.2 | 0.59 | 0.09 | 0.04 | 0.06 | |
| ~1.0 | 73 b | 9.26 | 1.65 | 25.4 | 2.6 | 3.4 | 0.8 | 3.7 | 0.36 | 17.8 | 0.10 | 0.13 | 0.03 | 0.15 |
| 124 f | 10.48 | 1.25 | 50.6 | 12.0 | 6.6 | 2.0 | 8.9 | 0.21 | 11.9 | 0.24 | 0.13 | 0.04 | 0.18 | |
| 193 a | 11.56 | 0.63 | 66.2 | 25.1 | 8.8 | 2.0 | 7.8 | 0.17 | 5.5 | 0.38 | 0.13 | 0.03 | 0.12 | |
| 241 c,d,e | 11.41 | 0.28 | 70.0 | 28.8 | 10.9 | 2.5 | 5.6 | 0.16 | 2.4 | 0.41 | 0.16 | 0.04 | 0.08 | |
| ~2.0 | 192 b | 9.74 | 1.95 | 48.8 | 9.5 | 3.9 | 1.2 | 13.1 | 0.20 | 20.0 | 0.19 | 0.08 | 0.03 | 0.27 |
| 242 a,c,d,e,f | 11.58 | 1.60 | 70.0 | 31.5 | 5.3 | 2.0 | 13.5 | 0.17 | 13.8 | 0.45 | 0.08 | 0.03 | 0.19 | |
| ~3.5 | 140 b,d,f | 8.30 | 1.32 | 61.4 | 26.2 | 13.1 | 1.2 | 2.4 | 0.14 | 15.9 | 0.43 | 0.21 | 0.02 | 0.04 |
| 240 a,c,e | 8.72 | 0.81 | 68.1 | 37.4 | 10.1 | 3.1 | 0.0 | 0.13 | 9.3 | 0.55 | 0.15 | 0.05 | 0.00 |
| Initial Phenolics (g/L) | Time (h) | C16:0 | Δ9C16:1 | C18:0 | Δ9C18:1 | Δ9,12C18:2 | UI |
|---|---|---|---|---|---|---|---|
| 0.0 | 72 | 21.9 | 9.9 | 12.4 | 43.3 | 12.3 | 0.778 |
| 144 | 16.9 | 13.9 | 6.5 | 48.3 | 14.2 | 0.906 | |
| 216 | 17.1 | 13.5 | 8.1 | 46.6 | 14.4 | 0.889 | |
| ~1.0 | 48 | 19.8 | 10.8 | 11.3 | 46.0 | 12.1 | 0.810 |
| 145 | 16.0 | 11.4 | 8.0 | 50.9 | 13.7 | 0.897 | |
| 217 | 17.2 | 11.0 | 9.2 | 51.1 | 11.5 | 0.851 | |
| ~2.0 | 48 | 20.1 | 10.7 | 8.1 | 48.4 | 12.7 | 0.845 |
| 144 | 11.5 | 4.5 | 5.4 | 61.3 | 17.3 | 1.004 | |
| 216 | 16.2 | 13.6 | 7.0 | 50.4 | 12.8 | 0.896 | |
| ~3.5 | 45 | 20.0 | 4.4 | 6.6 | 53.9 | 15.1 | 0.885 |
| 140 | 18.3 | 7.8 | 7.4 | 50.6 | 15.9 | 0.902 | |
| 209 | 16.7 | 3.0 | 6.8 | 56.4 | 17.1 | 0.937 |
| Glol0 (g/L) | Hours | X (g/L) | L (g/L) | Glolcons (g/L) | Cit (g/L) | Man (g/L) | Ara (g/L) | Ery (g/L) | YX/Glol (g/g) | YL/X % (w/w) | YCit/Glol (g/g) | YMan/Glol (g/g) | YAra/Glol (g/g) | YEry/Glol (g/g) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ~70.0 | 191 f | 8.98 | 0.60 | 64.9 | 17.3 | 9.9 | 3.1 | 14.9 | 0.14 | 6.7 | 0.27 | 0.15 | 0.05 | 0.23 |
| 240 d | 11.04 | 0.76 | 72.4 | 29.4 | 10.1 | 2.9 | 14.5 | 0.15 | 6.9 | 0.41 | 0.14 | 0.04 | 0.20 | |
| 246 a,b,e | 11.96 | 1.27 | 72.4 | 34.0 | 8.1 | 3.2 | 9.1 | 0.17 | 10.6 | 0.47 | 0.11 | 0.04 | 0.13 | |
| 312 c | 11.95 | 0.62 | 72.4 | 42.5 | 6.8 | 3.0 | 4.1 | 0.17 | 5.2 | 0.59 | 0.09 | 0.04 | 0.06 | |
| ~120.0 | 264 b,d,,e,f | 10.52 | 2.07 | 98.5 | 33.5 | 8.0 | 4.5 | 38.4 | 0.11 | 19.7 | 0.34 | 0.08 | 0.05 | 0.39 |
| 384 a,c | 12.40 | 1.11 | 122.9 | 63.8 | 7.2 | 4.2 | 31.4 | 0.10 | 9.0 | 0.52 | 0.06 | 0.03 | 0.26 | |
| ~170.0 | 408 b | 11.36 | 2.54 | 132.8 | 63.3 | 4.0 | 1.1 | 47.3 | 0.09 | 22.4 | 0.48 | 0.03 | 0.01 | 0.36 |
| 480 a | 12.48 | 2.52 | 160.5 | 72.9 | 4.6 | 1.9 | 58.8 | 0.08 | 15.9 | 0.45 | 0.03 | 0.01 | 0.37 | |
| 528 c,d,e,f | 11.98 | 0.83 | 170.6 | 79.0 | 6.5 | 3.4 | 65.8 | 0.07 | 6.9 | 0.46 | 0.04 | 0.02 | 0.39 |
| Glol0 (g/L) | Time (h) | C16:0 | Δ9C16:1 | C18:0 | Δ9C18:1 | Δ9,12C18:2 | UI |
|---|---|---|---|---|---|---|---|
| ~70 | 72 | 21.9 | 9.9 | 12.4 | 43.3 | 12.3 | 0.778 |
| 144 | 16.9 | 13.9 | 6.5 | 48.3 | 14.2 | 0.906 | |
| 216 | 17.1 | 13.5 | 8.1 | 46.6 | 14.4 | 0.889 | |
| ~120 | 48 | 15.3 | 16.3 | 5.0 | 50.1 | 13.2 | 0.928 |
| 120 | 15.7 | 22.3 | 4.8 | 47.7 | 9.3 | 0.886 | |
| 216 | 16.4 | 24.1 | 4.2 | 48.1 | 7.1 | 0.864 | |
| 288 | 14.4 | 23.4 | 4.3 | 49.5 | 8.3 | 0.895 | |
| 336 | 16.9 | 29.9 | 5.2 | 63.0 | 11.8 | 1.165 | |
| 456 | 18.0 | 17.5 | 6.2 | 6.2 | 52.0 | 1.278 | |
| ~170 | 72 | 36.1 | 14.8 | 16.2 | 26.6 | 6.3 | 0.540 |
| 168 | 18.2 | 28.9 | 5.0 | 41.9 | 6.1 | 0.830 | |
| 264 | 14.7 | 22.5 | 2.9 | 52.6 | 7.3 | 0.897 | |
| 360 | 12.9 | 32.0 | 2.9 | 45.9 | 6.4 | 0.906 | |
| 432 | 12.2 | 31.7 | 3.3 | 45.1 | 7.7 | 0.922 | |
| 504 | 14.1 | 28.7 | 5.0 | 44.8 | 7.3 | 0.881 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarris, D.; Rapti, A.; Papafotis, N.; Koutinas, A.A.; Papanikolaou, S. Production of Added-Value Chemical Compounds through Bioconversions of Olive-Mill Wastewaters Blended with Crude Glycerol by a Yarrowia lipolytica Strain. Molecules 2019, 24, 222. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24020222
Sarris D, Rapti A, Papafotis N, Koutinas AA, Papanikolaou S. Production of Added-Value Chemical Compounds through Bioconversions of Olive-Mill Wastewaters Blended with Crude Glycerol by a Yarrowia lipolytica Strain. Molecules. 2019; 24(2):222. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24020222
Chicago/Turabian StyleSarris, Dimitris, Anna Rapti, Nikolaos Papafotis, Apostolis A. Koutinas, and Seraphim Papanikolaou. 2019. "Production of Added-Value Chemical Compounds through Bioconversions of Olive-Mill Wastewaters Blended with Crude Glycerol by a Yarrowia lipolytica Strain" Molecules 24, no. 2: 222. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24020222