
Effect of Steaming Processing on Phenolic Profiles and Cellular Antioxidant Activities of Castanea mollissima


Abstract
:1. Introduction
2. Results
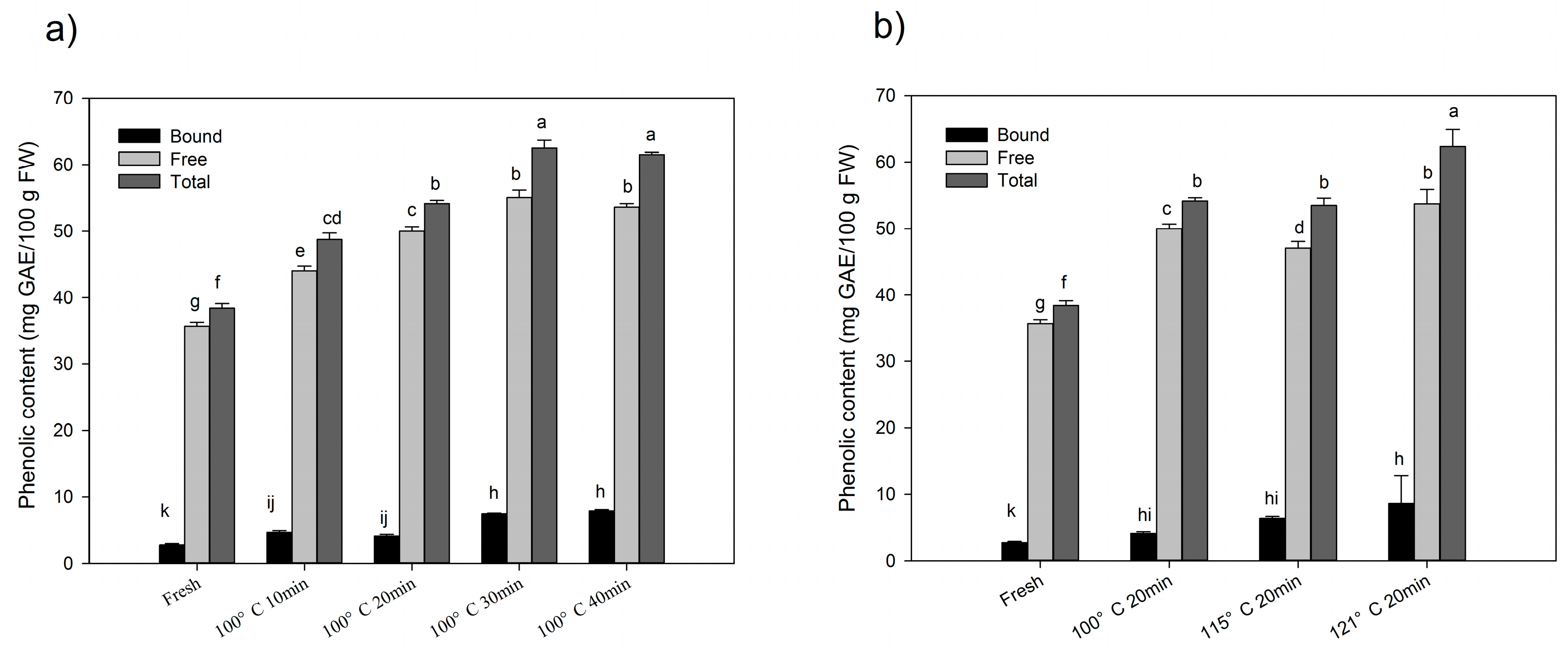
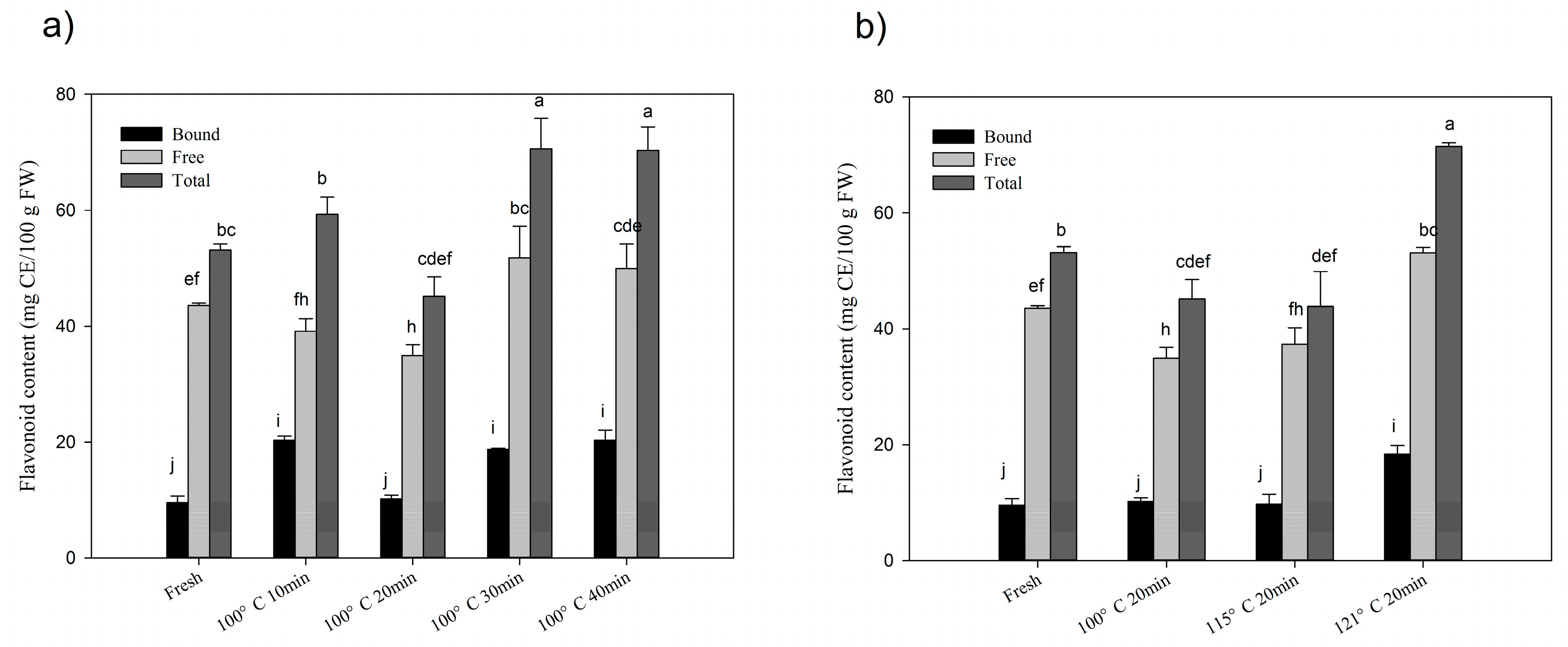
2.1. Changes of Total Phenolics and Flavonoids after Processing
2.2. Changes of Phenolic Components after Processing
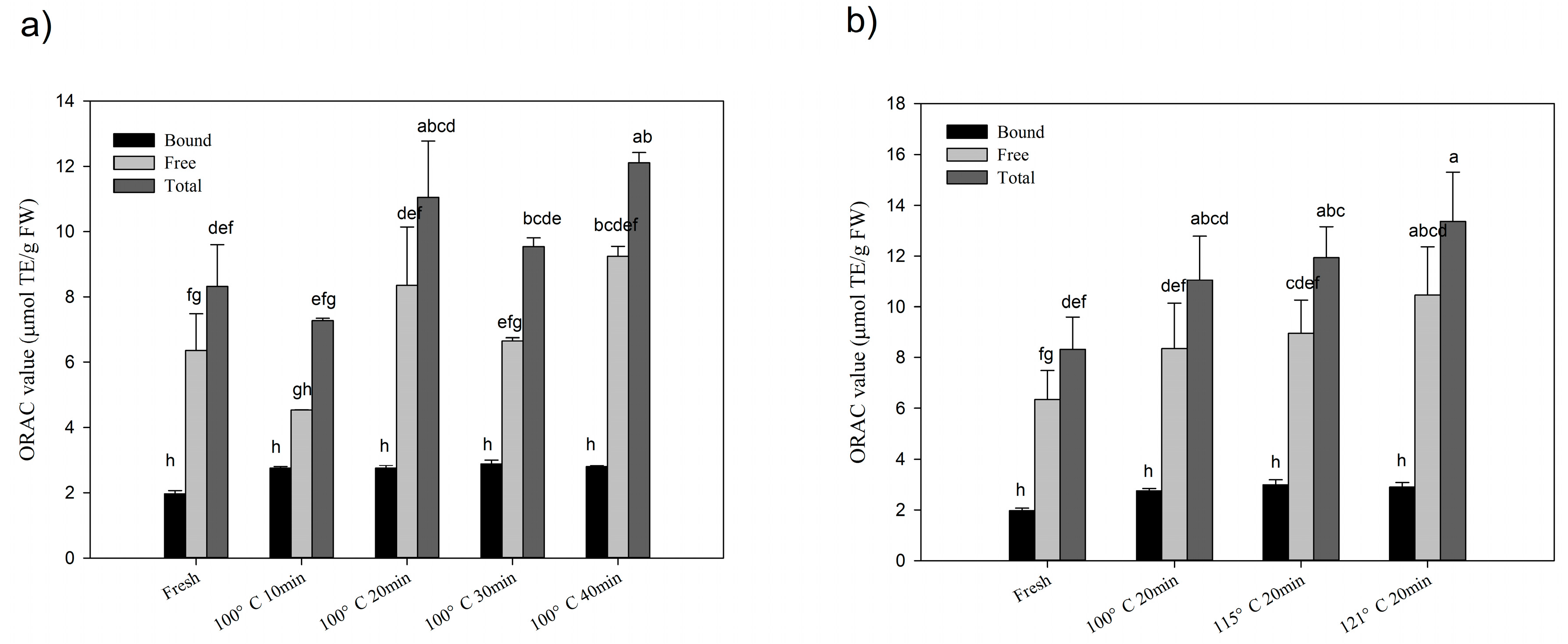
2.3. Changes of Total Antioxidant Activities after Processing
2.4. Changes of Cellular Antioxidant Activities after Processing
2.5. Correlation Analysis
3. Discussion
3.1. Influence on Phenolics and Flavonoids
3.2. Revelation from Phytochemical Profiling by HPLC
3.3. Effect of Steaming on Cellular Antioxidant Activity (CAA)
4. Materials and Methods
4.1. Sampling
4.2. Extraction
4.3. Determination of Total Phenolics and Flavonoids
4.4. Phenolic Acids and Flavonoids Profiling
4.5. Oxygen Radical Scavenging Capacity (ORAC) Assay
4.6. Cellular Antioxidant Activity (CAA) Assay
4.7. Statistics Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lv, J.; Zhao, Y.; Wang, J.Z.; Ouyang, J.; Wang, F.J. Effects of environmental factors on functional properties of Chinese chestnut (Castanea mollissima) protein isolates. Eur. Food Res. Technol. 2015, 240, 463–469. [Google Scholar] [CrossRef]
- Seo, K.H.; Lee, J.Y.; Debnath, T.; Kim, Y.M.; Park, J.Y.; Kim, Y.O.; Park, S.J.; Lim, B.O. DNA Protection and Antioxidant Potential of Chestnut Shell Extracts. J. Food Biochem. 2016, 40, 20–30. [Google Scholar] [CrossRef]
- Erturk, U.; Mert, C.; Soylu, A. Chemical composition of fruits of some important chestnut cultivars. Braz. Arch. Biol. Techn. 2006, 49, 183–188. [Google Scholar] [CrossRef]
- Abe, L.T.; Lajolo, F.M.; Genovese, M.I. Comparison of phenol content and antioxidant capacity of nuts. Cienc. Tecnol. Aliment. 2010, 30, 254–259. [Google Scholar] [CrossRef] [Green Version]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: Chemistry, bioavailability and effects on health. Nat. Prod. Rep. 2009, 26, 1001–1043. [Google Scholar] [CrossRef] [PubMed]
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouysegu, L. Plant Polyphenols: Chemical Properties, Biological Activities, and Synthesis. Angew. Chem. Int. Edit. 2011, 50, 586–621. [Google Scholar] [CrossRef]
- Barreira, J.C.M.; Ferreira, I.C.F.R.; Oliveira, M.B.P.P.; Pereira, J.A. Antioxidant activities of the extracts from chestnut flower, leaf, skins and fruit. Food Chem. 2008, 107, 1106–1113. [Google Scholar] [CrossRef] [Green Version]
- Pinto, D.; Rodrigues, F.; Braga, N.; Santos, J.; Pimentel, F.B.; Palmeira-de-Oliveira, A.; Oliveira, M.B.P.P. The Castanea sativa bur as a new potential ingredient for nutraceutical and cosmetic outcomes: Preliminary studies. Food Funct. 2017, 8, 201–208. [Google Scholar] [CrossRef]
- Gong, Z.Q.; Li, D.J.; Liu, C.Q.; Cheng, A.W.; Wang, W.L. Partial purification and characterization of polyphenol oxidase and peroxidase from chestnut kernel. Lwt Food Sci. Technol. 2015, 60, 1095–1099. [Google Scholar] [CrossRef]
- Zhou, D.; Li, L.; Wu, Y.W.; Fan, J.F.; Ouyang, J. Salicylic acid inhibits enzymatic browning of fresh-cut Chinese chestnut (Castanea mollissima) by competitively inhibiting polyphenol oxidase. Food Chem. 2015, 171, 19–25. [Google Scholar] [CrossRef]
- Nayak, B.; Liu, R.H.; Tang, J.M. Effect of Processing on Phenolic Antioxidants of Fruits, Vegetables, and Grains-A Review. Crit. Rev. Food Sci. 2015, 55, 887–918. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F. Effect of Processing on Quality Attributes of Chestnut. Food Bioprocess Technol. 2016, 9, 1429–1443. [Google Scholar] [CrossRef]
- Hou, F.; Shi, X.H.; Li, Q.; Xie, S.S.; Wu, Y.W.; Ouyang, J. Nutritional Quality of Chinese Chestnut and Effect of Cooking on its Bioactive Compounds and Antioxidant Activity. J. Food Process. Preserv. 2016, 40, 1383–1390. [Google Scholar] [CrossRef]
- Li, Q.; Shi, X.H.; Zhao, Q.J.; Cui, Y.H.; Ouyang, J.; Xu, F. Effect of cooking methods on nutritional quality and volatile compounds of Chinese chestnut (Castanea mollissima Blume). Food Chem. 2016, 201, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.D.; De Vasconcelos, M.; Bennett, R.N.; Rosa, E.A.S.; Cardoso, J.V.F. Primary and secondary metabolite composition of kernels from three cultivars of Portuguese chestnut (Castanea sativa Mill.) at different stages of industrial transformation. J. Agric. Food Chem. 2007, 55, 3508–3516. [Google Scholar]
- De Vasconcelos, M.C.B.M.; Bennett, R.N.; Rosa, E.A.S.; Ferreira-Cardoso, J.V. Composition of European chestnut (Castanea sativa Mill.) and association with health effects: Fresh and processed products. J. Sci. Food Agric. 2010, 90, 1578–1589. [Google Scholar] [CrossRef]
- Palermo, M.; Pellegrini, N.; Fogliano, V. The effect of cooking on the phytochemical content of vegetables. J. Sci. Food Agric. 2014, 94, 1057–1070. [Google Scholar] [CrossRef]
- Liu, F.Y.; Chang, X.X.; Hu, X.D.; Brennan, C.S.; Guo, X.B. Effect of thermal processing on phenolic profiles and antioxidant activities in Castanea mollissima. Int. J. Food Sci. Technol. 2017, 52, 439–447. [Google Scholar] [CrossRef]
- Barros, A.I.R.N.A.; Nunes, F.M.; Goncalves, B.; Bennett, R.N.; Silva, A.P. Effect of cooking on total vitamin C contents and antioxidant activity of sweet chestnuts (Castanea sativa Mill.). Food Chem. 2011, 128, 165–172. [Google Scholar] [CrossRef] [Green Version]
- Dewanto, V.; Wu, X.Z.; Liu, R.H. Processed sweet corn has higher antioxidant activity. J. Agric. Food Chem. 2002, 50, 4959–4964. [Google Scholar] [CrossRef]
- Czubinski, J.; Dwiecki, K. A review of methods used for investigation of protein-phenolic compound interactions. Int. J. Food Sci. Technol. 2017, 52, 573–585. [Google Scholar] [CrossRef]
- Ozdal, T.; Capanoglu, E.; Altay, F. A review on protein-phenolic interactions and associated changes. Food Res. Int. 2013, 51, 954–970. [Google Scholar] [CrossRef]
- Wolfe, K.L.; Liu, R.H. Cellular antioxidant activity (CAA) assay for assessing antioxidants, foods, and dietary supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, B.; Borges, O.; Costa, H.S.; Bennett, R.; Santos, M.; Silva, A.P. Metabolite composition of chestnut (Castanea sativa Mill.) upon cooking: Proximate analysis, fibre, organic acids and phenolics. Food Chem. 2010, 122, 154–160. [Google Scholar] [CrossRef] [Green Version]
- Nazzaro, M.; Barbarisi, C.; La Cara, F.; Volpe, M.G. Chemical and biochemical characterisation of an IGP ecotype chestnut subjected to different treatments. Food Chem. 2011, 128, 930–936. [Google Scholar] [CrossRef]
- Li, G.P.; Zhou, D.; Kan, L.N.; Wu, Y.W.; Fan, J.F.; Ouyang, J. Competitive inhibition of phytic acid on enzymatic browning of chestnut (Castanea mollissima Blume). Acta Aliment. 2017, 46, 100–108. [Google Scholar] [CrossRef] [Green Version]
- Hwang, S.H.; Zuo, G.L.; Wang, Z.Q.; Lim, S.S. Novel aldose reductase inhibitory and antioxidant chlorogenic acid derivatives obtained by heat treatment of chlorogenic acid and amino acids. Food Chem. 2018, 266, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Dawidowicz, A.L.; Typek, R. Thermal Stability of 5-O-Caffeoylquinic Acid in Aqueous Solutions at Different Heating Conditions. J. Agric. Food Chem. 2010, 58, 12578–12584. [Google Scholar] [CrossRef] [PubMed]
- Gullon, B.; Eibes, G.; Davila, I.; Moreira, M.T.; Labidi, J.; Gullon, P. Hydrothermal treatment of chestnut shells (Castanea sativa) to produce oligosaccharides and antioxidant compounds. Carbohydr. Polym. 2018, 192, 75–83. [Google Scholar] [CrossRef]
- Gu, H.H.; Yao, H.; Wang, F.J. Structural and physicochemical properties of resistant starch from Chinese chestnut (Castanea mollissima) prepared by autoclaving treatment and pullulanase hydrolysis. J. Food Process. Preserv. 2018, 42. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, T.; Fu, X.; Brennan, M.; Abbasi, A.M.; Zheng, B.S.; Liu, R.H. The use of an enzymatic extraction procedure for the enhancement of highland barley (Hordeum vulgare L.) phenolic and antioxidant compounds. Int. J. Food Sci. Technol. 2016, 51, 1916–1924. [Google Scholar] [CrossRef]
- Zhang, L.Z.; Liu, R.H. Phenolic and carotenoid profiles and antiproliferative activity of foxtail millet. Food Chem. 2015, 174, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Nayak, B.; Liu, R.H.; Berrios, J.D.; Tang, J.; Derito, C. Bioactivity of Antioxidants in Extruded Products Prepared from Purple Potato and Dry Pea Flours. J. Agric. Food Chem. 2011, 59, 8233–8243. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.B.; Li, T.; Tang, K.X.; Liu, R.H. Effect of Germination on Phytochemical Profiles and Antioxidant Activity of Mung Bean Sprouts (Vigna radiata). J. Agric. Food Chem. 2012, 60, 11050–11055. [Google Scholar] [CrossRef]
- He, X.J.; Liu, D.; Liu, R.H. Sodium Borohydride/Chloranil-Based Assay for Quantifying Total Flavonoids. J. Agric. Food Chem. 2008, 56, 9337–9344. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cao, G.H.; Prior, R.L. Oxygen radical absorbing capacity of anthocyanins. J. Agric. Food Chem. 1997, 45, 304–309. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| Compound | Conjugation | Fresh | 100 °C 10 min | 100 °C 20 min | 100 °C 30 min | 100 °C 40 min | 115 °C 20 min | 121 °C 20 min |
|---|---|---|---|---|---|---|---|---|
| Ferulic acid | free | 2.98 ± 0.01 a | 2.91 ± 0.01 b | 2.93 ± 0.01 b | 2.83 ± 0.01 c | 2.83 ± 0.01 c | 2.92 ± 0.03 b | 2.73 ± 0.01 d |
| bound | 2.92 ± 0.43 e | 3.57 ± 0.01 bc | 3.24 ± 0.02 c,d | 4.16 ± 0.05 a | 4.10 ± 0.01 a | 3.88 ± 0.01 a,b | 3.58 ± 0.01 b,c | |
| Chlorogenic acid | free | 18.28 ± 0.14 d | 15.16 ± 0.13 e | 18.80 ± 0.12 c | 18.81 ± 0.08 c | 22.42 ± 0.01 a | 21.13 ± 0.28 b | 21.17 ± 0.16 b |
| bound | ND | ND | ND | ND | ND | ND | ND | |
| Gallic acid | free | 9.76 ± 0.30 d | 9.61 ± 0.09 d | 12.93 ± 0.06 a | 11.72 ± 0.03 c | 12.54 ± 0.02 b | 4.99 ± 0.11 e | 5.32 ± 0.01 e |
| bound | 5.55 ± 0.01 a | 2.86 ± 0.01 e | 2.92 ± 0.01 d | 2.85 ± 0.01 e | 2.86 ± 0.01 e | 3.94 ± 0.01 c | 5.47 ± 0.01 b | |
| Vanillic acid | free | 6.38 ± 0.14 b | 6.47 ± 0.01 a,b | 6.38 ± 0.01 b | 6.54 ± 0.01 a,b | 6.65 ± 0.01 a | 6.47 ± 0.11 a,b | 6.52 ± 0.04 a,b |
| bound | ND | ND | ND | ND | ND | ND | ND | |
| Syringate | free | 3.95 ± 0.06 a | 2.74 ± 0.02 e | 3.43 ± 0.02 b | 3.26 ± 0.09 c | 2.97 ± 0.05 d | 3.19 ± 0.07 c | 2.72 ± 0.01 e |
| bound | ND | ND | ND | ND | ND | ND | ND | |
| Quercetin | free | 1.70 ± 0.11 a | 0.40 ± 0.01 d | 0.53 ± 0.01 c,d | 0.59 ± 0.04 c,d | 0.37 ± 0.01 d | 1.39 ± 0.19 b | 0.63 ± 0.01 c |
| bound | 0.90 ± 0.01 c | 1.62 ± 0.01 a | 1.13 ± 0.10 b | 0.75 ± 0.06 d | 0.99 ± 0.05 b,c | 0.84 ± 0.01 c,d | 1.58 ± 0.07 a | |
| p-Coumaric acid | free | 2.38 ± 0.01 c,d | 2.39 ± 0.01 c | 2.39 ± 0.01 c | 2.39 ± 0.01 c | 2.37± 0.01 d | 2.43± 0.01 b | 2.52± 0.01 a |
| bound | 2.41 ± 0.01 d,e | 2.77 ± 0.01 a | 2.42 ± 0.01 c,d | 2.43 ± 0.01 c | 2.41± 0.01 d,e | 2.43± 0.01 c | 2.46 ± 0.01 b |
| Sample Group | CAA Value (nmol QE/g FW) | Cellular Uptake (%) | ||||
|---|---|---|---|---|---|---|
| No-PBS Wash | PBS Wash | |||||
| Free | Bound | Free | Bound | Free | Bound | |
| Fresh | 1.69 ± 0.51 b | 0.19 ± 0.01 a,b | 0.24 ± 0.01 d,e | 0.17 ± 0.04 a | 14.3% d | 88.1% a |
| 100 °C 10 min | 1.61 ± 0.16 b | ND | 0.20 ± 0.05 e | ND | 12.3% d | ND |
| 100 °C 20 min | 1.52 ± 0.07 b | 0.19 ± 0.01 a,b | 0.28 ± 0.01 c,d,e | ND | 18.6% c | ND |
| 100 °C 30 min | 1.69 ± 0.07 b | ND | 0.45 ± 0.06 a | ND | 26.3% a | ND |
| 100 °C 40 min | 2.27 ± 0.10 a | 0.11 ± 0.04 c | 0.31 ± 0.01 b,c | ND | 13.4% d | ND |
| 115 °C 20 min | 1.81 ± 0.05 b | 0.24 ± 0.01 a | 0.34 ± 0.01 b | 0.06 ± 0.02 b | 18.7% c | 26.8% c |
| 121 °C 20 min | 1.61 ± 0.02 b | 0.14 ± 0.04 b,c | 0.33 ± 0.03 b,c | 0.07 ± 0.02 b | 20.3% b | 48.2% b |
| Correlation | ORAC | CAA No-PBS Wash | CAA PBS Wash | TPC | TFC | Thermal Processes |
|---|---|---|---|---|---|---|
| ORAC | - | 0.40 | 0.23 | 0.59 * | 0.17 | 0.78 ** |
| CAA No-PBS wash | - | - | 0.12 | 0.16 | 0.11 | 0.21 |
| CAA PBS wash | - | - | - | 0.10 | 0.16 | −0.44 * |
| TPC | - | - | - | - | 0.61 ** | 0.56 * |
| TFC | - | - | - | - | - | 0.18 |
| TFA | 0.24 | 0.36 | 0.07 | 0.65 ** | 0.42 | 0.41 |
| TCA | 0.82 ** | 0.70 ** | 0.42 | 0.55 ** | 0.25 | 0.66 ** |
| TGA | −0.30 | 0.05 | −0.15 | −0.13 | 0.06 | −0.84 ** |
| TVA | 0.29 | 0.35 | 0.06 | 0.63 ** | 0.68 ** | 0.40 |
| TS | −0.29 | 0.05 | 0.39 | −0.62 ** | −0.51 * | −0.57 * |
| TQ | −0.19 | −0.14 | 0.17 | −0.69 ** | −0.39 | −0.01 |
| TPA | −0.29 | −0.49 * | −0.55 * | −0.08 | 0.12 | 0.19 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, F.; Liu, F.; Abbasi, A.M.; Chang, X.; Guo, X. Effect of Steaming Processing on Phenolic Profiles and Cellular Antioxidant Activities of Castanea mollissima. Molecules 2019, 24, 703. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24040703
Zhang F, Liu F, Abbasi AM, Chang X, Guo X. Effect of Steaming Processing on Phenolic Profiles and Cellular Antioxidant Activities of Castanea mollissima. Molecules. 2019; 24(4):703. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24040703
Chicago/Turabian StyleZhang, Fangyuan, Fengyuan Liu, Arshad Mehmood Abbasi, Xiaoxiao Chang, and Xinbo Guo. 2019. "Effect of Steaming Processing on Phenolic Profiles and Cellular Antioxidant Activities of Castanea mollissima" Molecules 24, no. 4: 703. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24040703