1. Background
Dermatophytosis is a common and relevant zoonotic disease of public health concern [
1]. It is caused by various species of dermatophytes that damage superficial keratinized tissues in both humans and animals [
2,
3]. One of the common dermatophyte species in this regard is
Trichophyton mentagrophytes, which is equally important for humans and animals. Rabbits are frequent hosts of
T. mentagrophytes [
4], which causes serious infections and inflicts substantial economic loss on rabbit farmers [
5]. Various antifungal compounds, including clotrimazole, terbinafine, and ketoconazole have been reported for the treatment of dermatophytosis [
6]. However, drug resistance, toxicity, and drug-drug interactions limit their application [
7]. To identify new classes of antifungal agents to overcome multi-drug resistance mechanisms, many research groups have assessed medicinal plants worldwide [
8]. Antimicrobial and antifungal properties of medicinal plant extracts have been widely reported [
9]. A previous study has demonstrated that the ethanolic extract of
Phellodendron amurense has significant antifungal effects both in vitro and in vivo against
T. mentagrophytes [
10]. Berberine hydrochloride constitutes the most important bioactive compound of
Cortex phellodendri, accounting for approximately 0.6% of the extract from
Berberidis Radix [
11,
12].
Phellodendron amurense mainly contains about 1.6%
Berberidis Radix.
Berberidis Radix is a kind of alkaloid, which is not easy to dissolve in water. It is formed after salt formation with hydrochloric acid. The solubility of berberine hydrochloride is improved and it is easy to dissolve in water. Berberine was shown to inhibit bacteria and other microbes [
13,
14,
15]. Some studies have found that Berberine can inhibit the formation of bacterial extracellular amyloid peptide, thereby interfering with the formation and stability of biofilm [
16]. In addition, Berberine can also affect the integrity and permeability of bacterial cell membrane, and bind to some proteins on the membrane, thus affecting the structure and function of proteins. It also affects the expression of bacterial DNA and binds to DNA [
17]. High-throughput RNA sequencing (RNA-Seq) constitutes a new potent and affordable method for transcriptomic studies [
18]. Indeed, the data produced by this technology include sufficient read coverage for
de novo transcriptome assembly, gene expression assessment, and gene discovery [
19]. Sterol 14α-demethylation as a general part of sterol biosynthetic pathways in eukaryotes has been known and studied for more than 30 years [
20]. The enzyme catalyzing this reaction was first purified from Saccharomyces cerevisiae in 1984 [
21]. Sterol 14α-demethylase participates in sterol biosynthesis and is an essential requirement for yeast viability. Sterol 14α-demethylase is the target for azole antifungal compounds, and resistance to these drugs and agrochemicals is of significant practical importance [
22]. In the present study, berberine hydrochloride displayed antifungal activity against
T. mentagrophytes dermatophytosis, and Illumina Sequencing for transcriptome analysis was used to explore the mechanism of this antifungal activity.
3. Discussion
According to the U.S. Environmental Protection Agency (EPA), in 1997 approximately 244,000 tons and 37,000 tons of fungicides were sold worldwide and in the United States, respectively [
23]. A broad spectrum antifungal activity has been demonstrated for azole fungicides, which are employed both in prevention and cure of fungal infections. Similar to the emerging antibiotic resistance, there is a possibility of developing antifungal resistance as a result of increased use of antifungal products in humans, animals, and plants. Antifungal resistance to azoles, and some of its underlying molecular mechanisms have been reported in agricultural research works [
24]. Therefore, identifying new medicinal drugs against fungi with reduced side effects is a necessity.
Despite the reported antifungal properties of several plants, their underlying mechanisms remain largely unknown. Berberine hydrochloride, a major
Cortex Phellodendri constituent, shows various biological properties [
25] including antifungal activity [
26,
27].
T. mentagrophytes is one of the common species of dermatophytes, and is equally important for humans and animals. It is frequently found in rabbits [
28] and causes serious infections, with subsequent economic loss to rabbit farmers [
5]. In the present study, we found antifungal activity for berberine hydrochloride against
T. mentagrophytes in animal experiments.
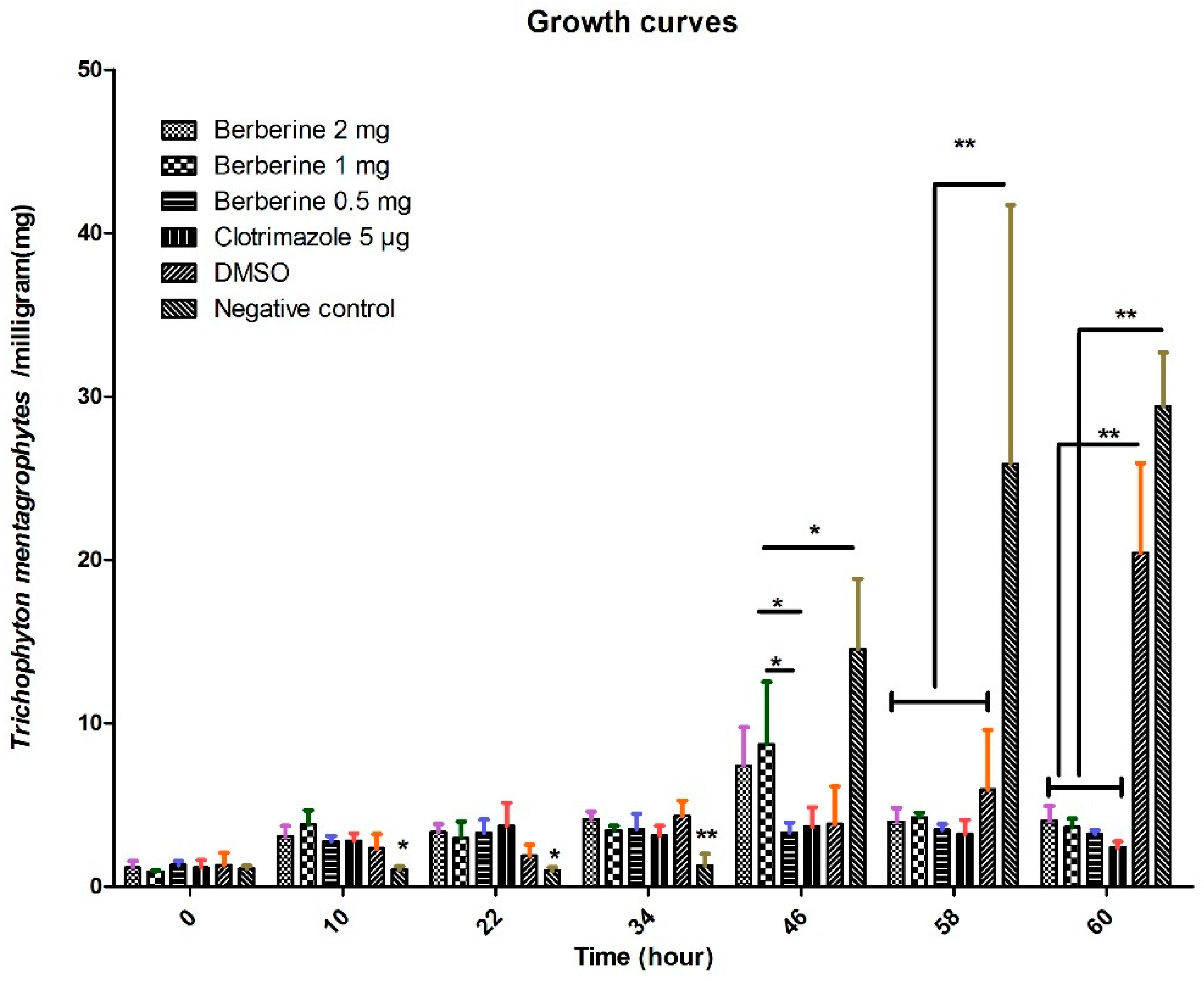
The growing rate of
T. mentagrophytes treated with untreated control was significantly higher than those obtained in the berberine hydrochloride and clotrimazole groups at 60 h, demonstrating that it was fungicidal at a relatively high berberine hydrochloride dose or clotrimazole dose. Interestingly, we noticed that berberine hydrochloride 0.5 mg is more effective than 1 mg (46 h point), it might be in the early stage of action, low dose treatment may have better effect on fungi. The potential flaws of our present study include lacking an azole resistant
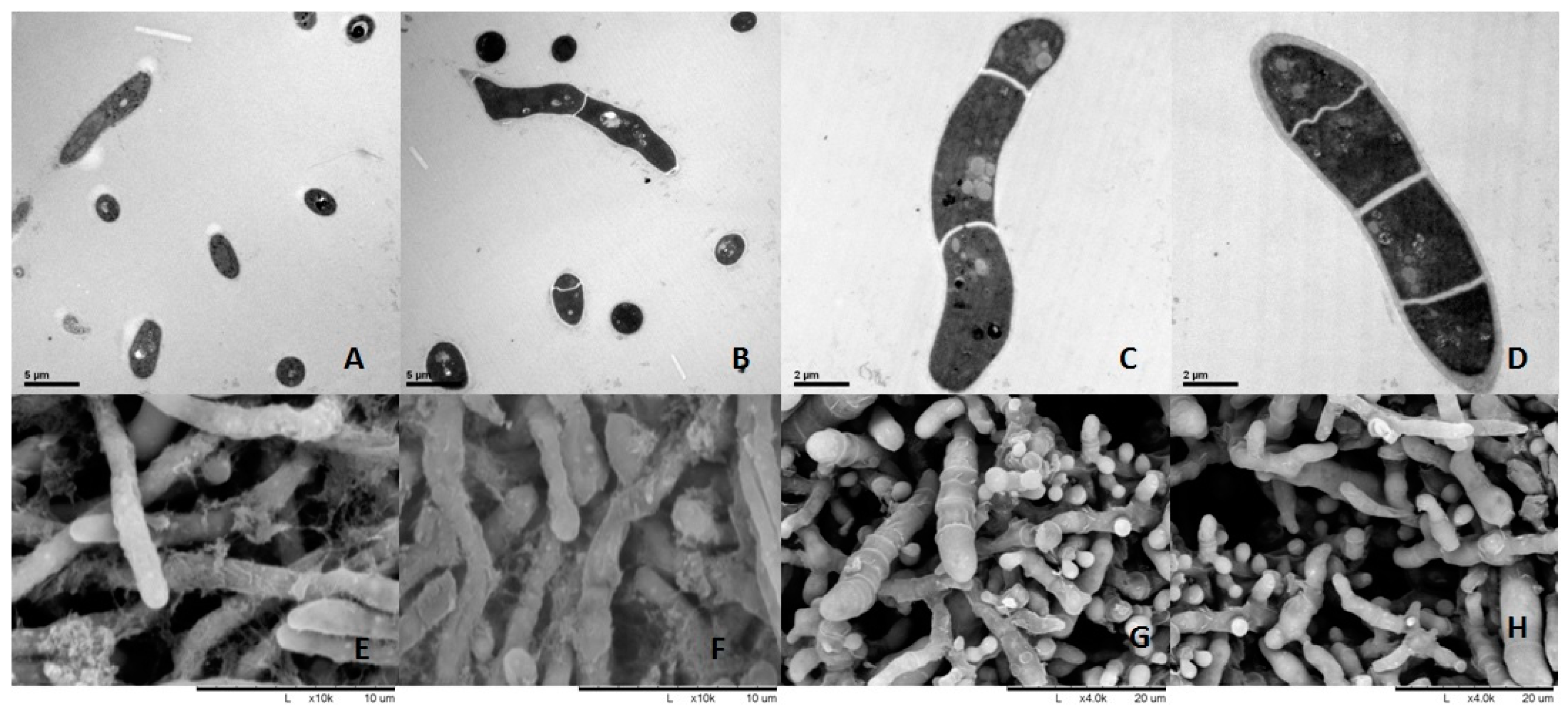
T. mentagrophytes strain which could be used in vitro inhibition assay using berberine hydrochloride at an inhibitory dose in order to unveil if it is also resistant to berberine hydrochloride at certain concentration. Berberine or clotrimazole altered the shape and cell membrane of
Trichophyton mentagrophytes, as examined by TEM and SEM, respectively, confirming the antifungal activity of berberine chloride against
T. mentagrophytes. Similar results were obtained in previously reported studies [
10]. Thus, clinical and PAS experiments demonstrated that berberine chloride could cure fungal disease in rabbits effectively.
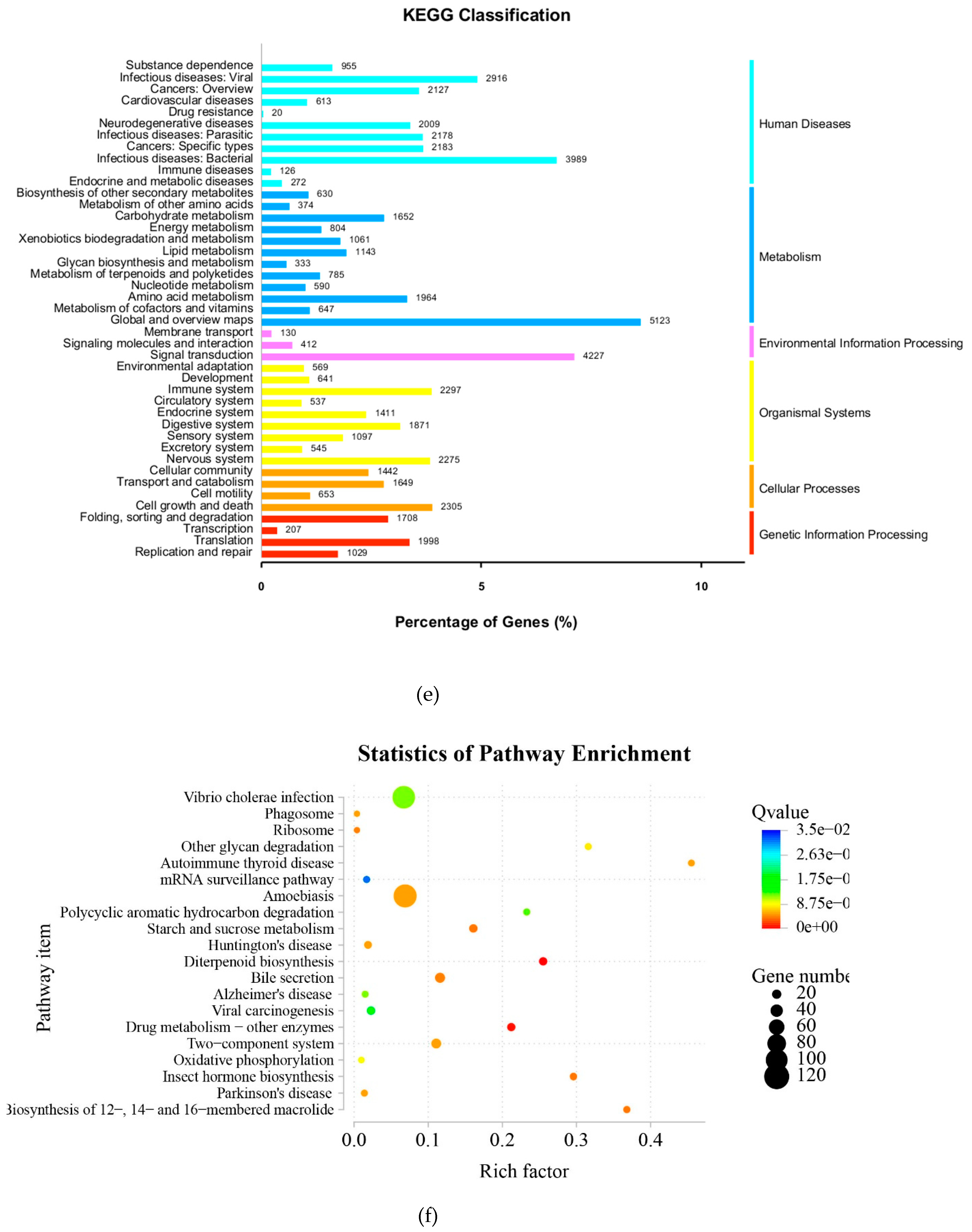
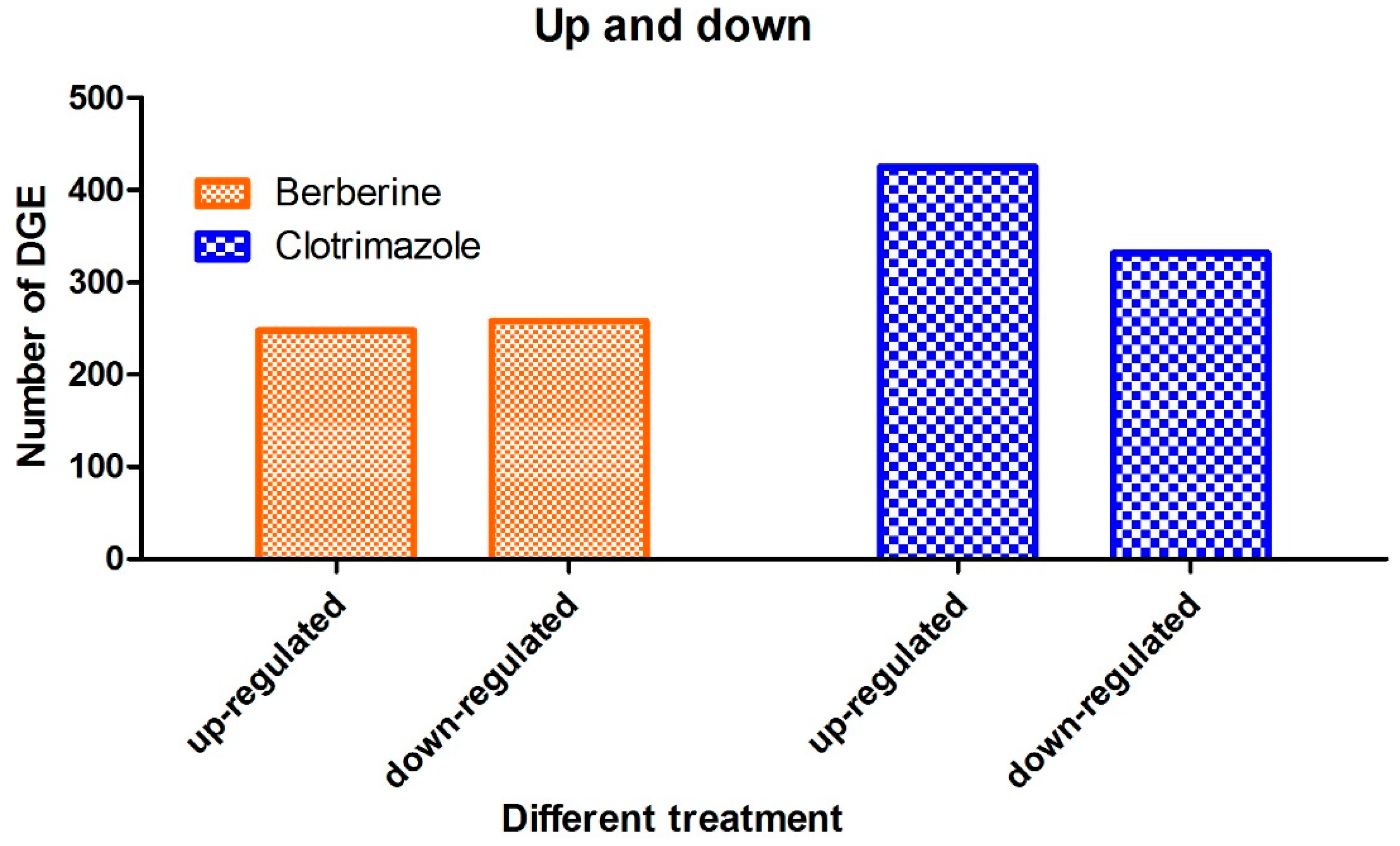
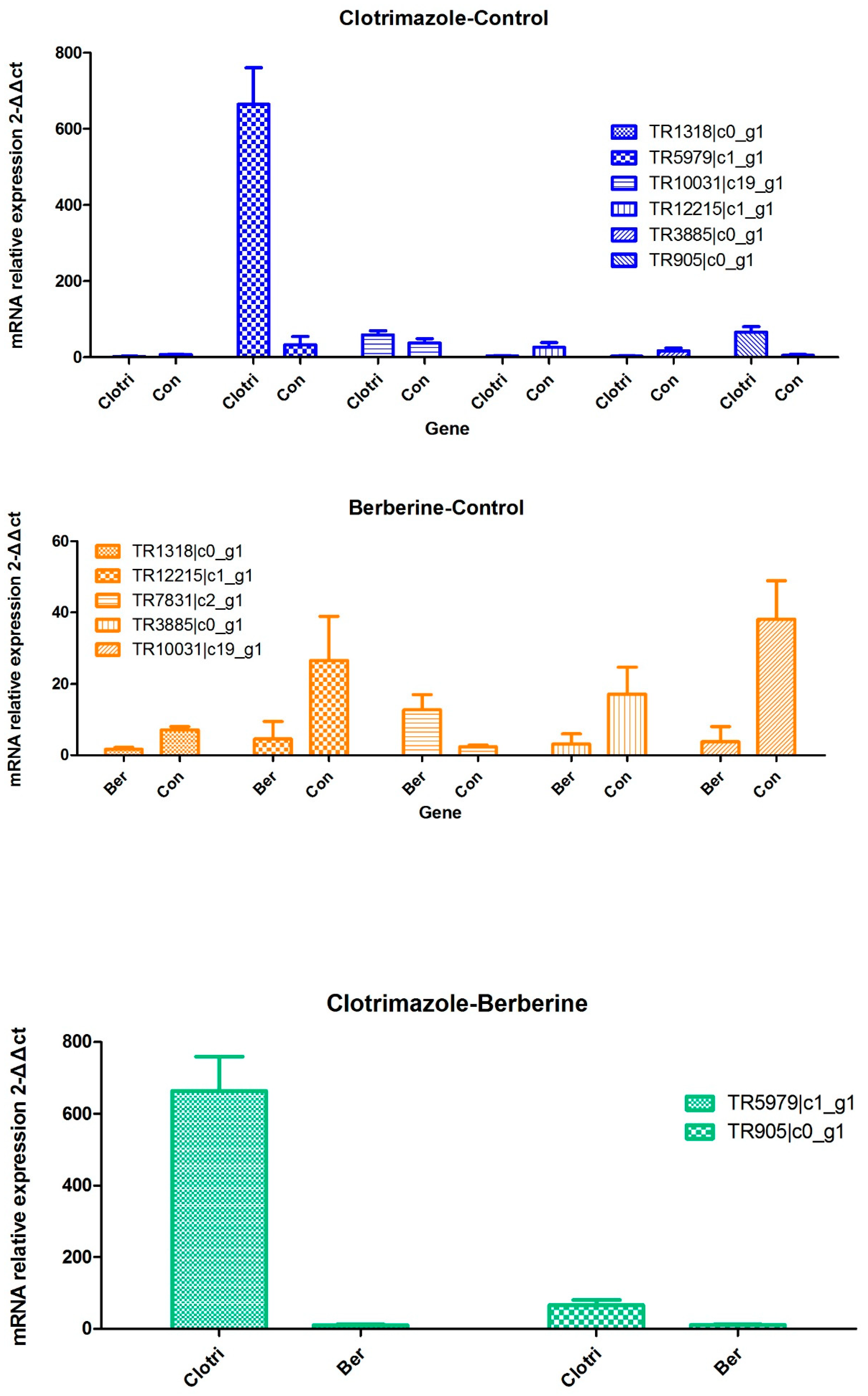
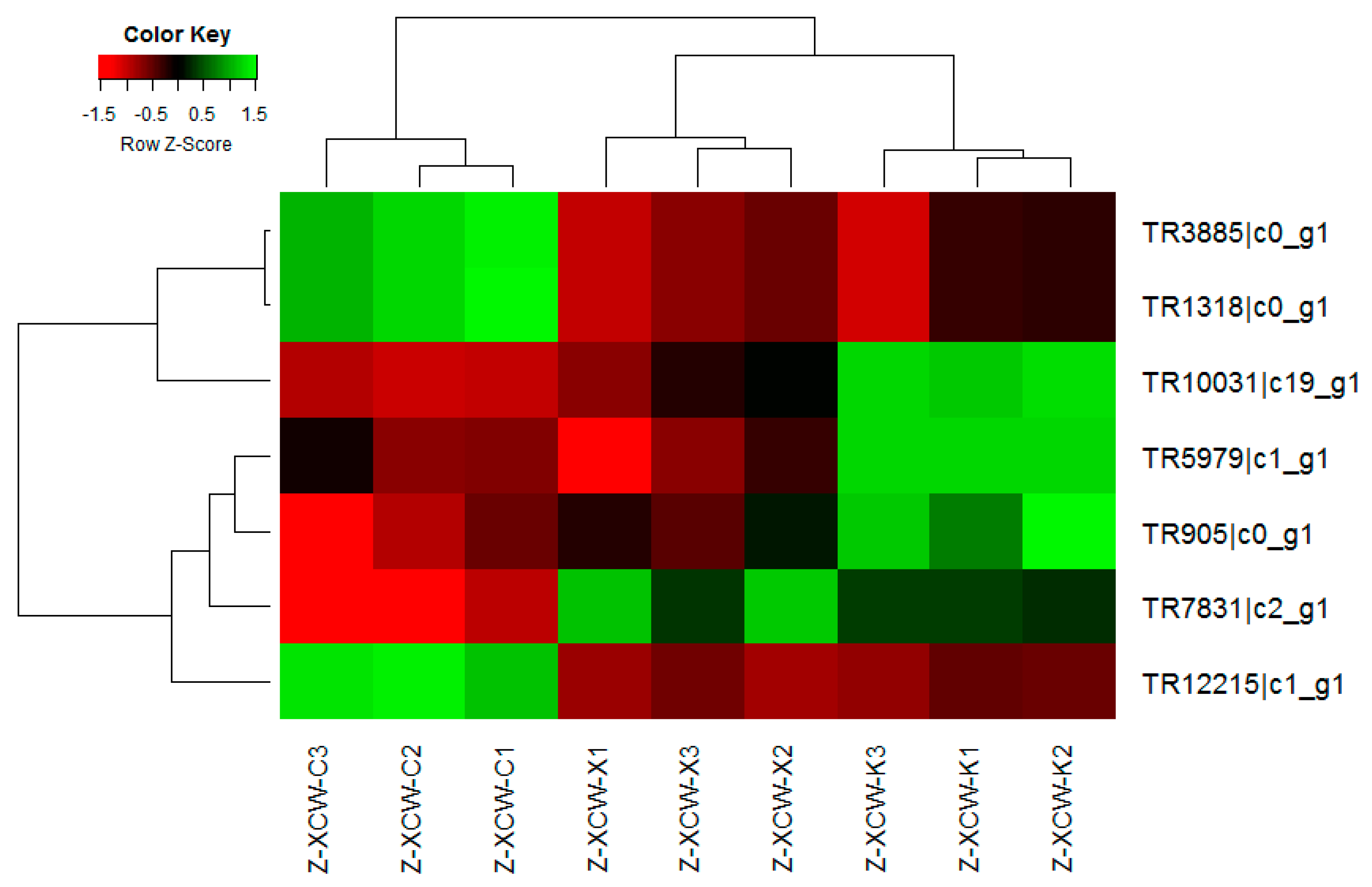
A comprehensive evaluation of gene expression was carried out with the DEGs method. As shown above, multiple signaling pathways were altered in T. mentagrophytes treated with berberine chloride, including metabolic pathways, biosynthesis of secondary metabolites, microbial metabolism. These data were validated by qRT-PCR.
SSRs (simple sequence repeats) represent an informative class of genetic markers (
Table 3). They are distributed throughout the coding and non-coding regions of all eukaryotic genomes [
29], widely use to characterize genetic diversity, construct linkage maps and even tag genes for the purpose of marker-assisted breeding [
30]. SSR markers acquired by our transcriptome analysis are potentially useful for genetic analysis in the
T. mentagrophytes genome. The genomic variable affecting genetic function may have an evolutionary role. It has reported that the rapid evolution of SSR sequences, specifically for those gains or losses of repeats at certain locus, may provide a molecular basis for adaptation to various environments [
31].
The fungal sterol 14α-demethylase is an important ergosterol precursor. Azole compounds exert inhibit yeast and fungal sterol 14α-demethylase, suppressing the synthesis of ergosterol, which is a critical constituent of the membrane in these organisms [
32]; blocking the synthesis of ergosterol causes disturbance in cell membrane assembly. Sterol 14α-demethylase belongs to the heme-containing cytochrome P450 (CYP51) superfamily of metabolic proteins. The reaction closely resembles demethylation at C-10 by aromatase [
33]; sterol 14α-demethylase oxidatively demethylates C-14 of sterols. In the present study, 14α-demethylase was downregulated upon exposure to either berberine hydrochloride or clotrimazole (Please
Table 6). This indicates that both medicines potentially might inhibit the growth of
T. mentagrophytes by blocking the biosynthesis of ergosterol.
Lathosterol transformation into 7-dehydrocholesterol downstream the squalene pathway of cholesterol biosynthesis involves lathosterol oxidase (5-DES, Δ7-sterol-C5(6)-desaturase), a membrane microsomal enzyme [
34], requiring molecular oxygen and remarkably induced by NAD(P)H [
35]. This enzyme was proposed to have non-heme iron in its catalytic center, and is inhibited by cyanides, Tiron [
36], and various hydrophilic chelators [
35], but not affected by carbon monoxide [
35]. In the present study, the expression of Lathosterol oxidase in
T. mentagrophytes was reduced upon exposure to either berberine hydrochloride or clotrimazole, please see
Table 6 and
Table 7. This suggested that both medicines could inhibit the growth of
T. mentagrophytes by blocking the biosynthesis of Lathosterol oxidase.
Aldehydes are reduced to related alcohols by members of the aldo–keto reductase (AKR) superfamily, in an NADPH-dependent manner [
37]. Detoxification of acrolein occurs through involvement of AKR1B, AKR1B7, AKR1C, AKR7A1, and AKR7A2. AKR1A also participates in metabolic pathways requiring the reduction aldehyde portions in target compounds [
38]. Ascorbic acid (AsA) has been reported to be produced by the AKR1A and AKR1B enzymes [
39]. We found that the expression of Aflatoxin B1 aldehyde reductase member 2 was significantly increased upon
T. mentagrophytes exposure to either berberine hydrochloride or clotrimazole (
Table 6 and
Table 7); this might be the pressure response of
T. mentagrophytes to degrade berberine hydrochloride or clotrimazole for self-protection against the stress of medicine.
Most eukaryotic cell membranes comprise sterols, which play key roles in sustaining membrane integrity and fluidity. Azoles have been used as sterol biosynthesis inhibitors in systemic antifungal therapy in humans [
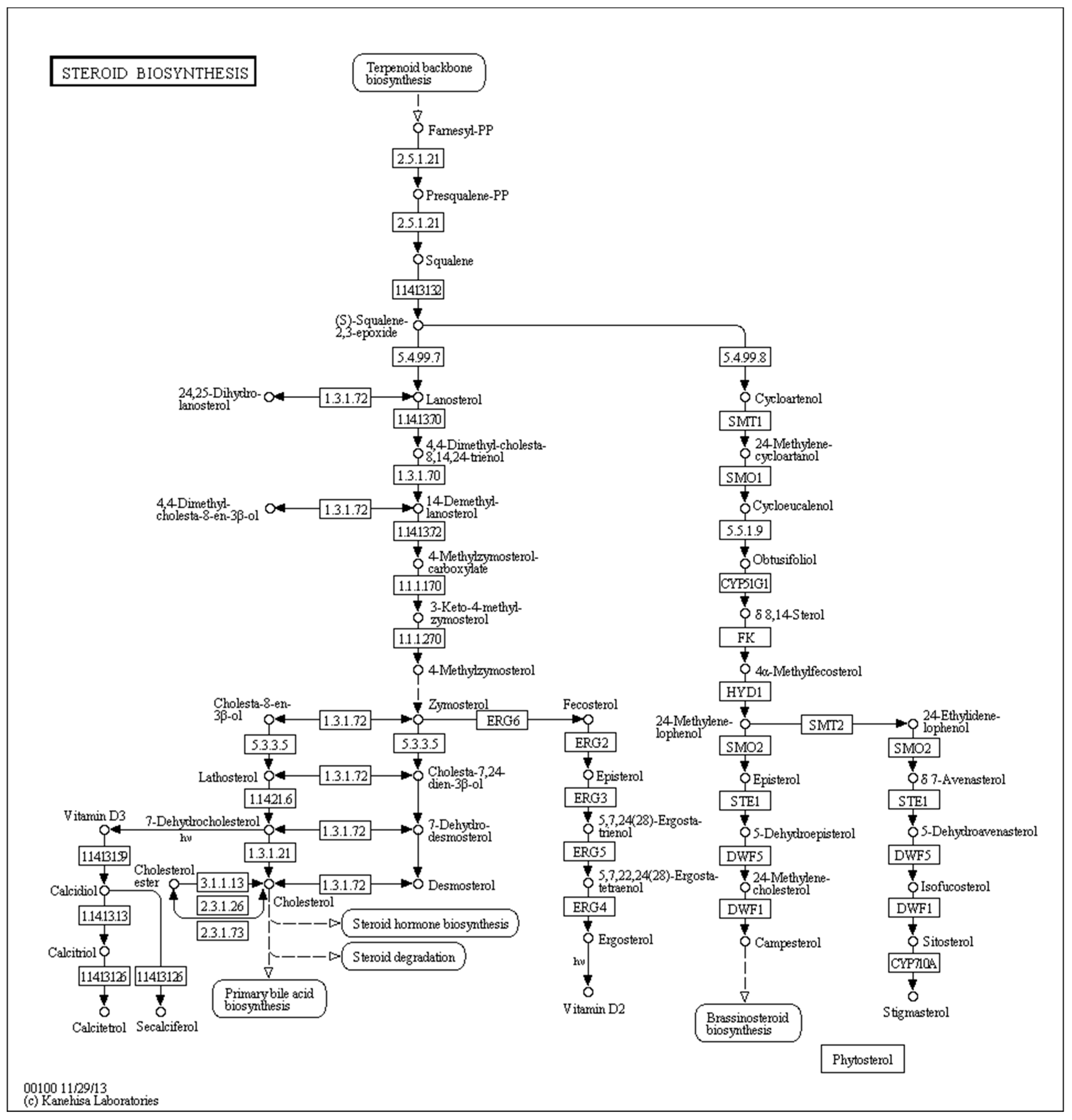
40]. In the present study, KEGG pathway analysis showed that steroid biosynthesis (map00100) (
p < 0.0024899) (
Figure 10) was very important in the activities clotrimazole against
T. mentagrophytes while berberine hydrochloride is affection the sterol synthesis. Some of the key genes in this pathway are sterol 14α-demethylase, methylsterol monooxygenase, and sterol 24-C-methyltransferase erg-4. We found that 14α-demethylase, methylsterol monooxygenase and sterol 24-C-methyltransferase erg-4 were down-regulated after treatment with either berberine hydrochloride or clotrimazole. These findings suggested that both medicines could inhibit steroid biosynthesis to prevent the growth of
T. mentagrophytes. Modification of the cyp51A gene is responsible for resistance of
A. fumigatus against azoles. Cyp51A counts as one of the two genes encoding sterol C14-demethylase isoforms in the above pathogen. Interestingly, we found that down-regulation of 14α-demethylase exposed to various medicines was slightly different, i.e., berberine hydrochloride (fold change −3.4956) and clotrimazole (fold change −2.1283) (
Table 5 and
Table 6) caused various degrees of alteration. This might be one of the anti-resistance mechanisms of berberine hydrochloride in
T. mentagrophytes.
4. Methods
4.1. Fungi Growth
Eumycetes from dermopathic rabbits were provided by the Institute of Internal Medicine, Shaoxing District (Shaoxing, China). The Chinese Academy of Medical Sciences confirmed the presence of the
T. mentagrophytes strain in eumycetes, which was grown in Tryptic Soy Broth (TSB) at 28 °C, TSB contained 17 g of Tryptone, 3 g of Phytone, 5 g of NaCl and 1 liter of distilled water, pH 7.1 [
41].
4.2. Growth Curve by Dry Weight Determination
Berberine hydrochloride (BC) (>98% purity, Lot No. 20130306) was manufactured by Shanghai Yuanye Biotechnology Co., Ltd. (Shanghai, China). Clotrimazole (CLO) (99% purity, Lot No. 23593-75-1) was provided by BaDaTong Medical Company (Taizhou, Zhejiang, China). Time and concentration effects of berberine and clotrimazole on
T. mentagrophytes were determined as proposed previously [
42]. Briefly, the fungi were cultured at 30,000 cells/mL, in presence of berberine hydrochloride (0.5, 1, or 2 mg/mL) or clotrimazole (5.0 μg/mL) at 37 °C for 0 h, 10 h, 22 h, 34 h, 46 h, 58 h and 60 h, respectively. Duplicate 1 mL aliquots of homogenized samples were removed from each conical flask and transferred to pre-weighed Eppendorf tubes. After centrifugation was performed 13,300×
g for 20 min, the sediments were dried in an oven at 60 °C. Differential weights were determined using an analytical balance. Each experiment was performed in duplicate. The growth of
T. mentagrophytes was observed.
4.3. Ultra-Structural Analysis by Electron Microscopy
The fungi were cultured at 30,000 cells/mL, Berberine hydrochloride (1 mg/mL) and Clotrimazole (0.5 μg/mL) were added in fungi for 6 h and the fungi were analyzed as well as DMSO controls. Berberine hydrochloride and clotrimazole were dissolved in DMSO, then dissolved in distilled water, the concentration of DMSO in water was 5%.
For TEM, the samples were fixed with 2.5% glutaraldehyde in phosphate buffer (0.1 M, pH 7.0; >4 h) followed by post-fixation with 1% OsO4 in phosphate buffer (1–2 h). After washing, the specimens were dehydrated by graded ethanol and incubated in acetone. After resin embedding, the samples were sectioned on a LEICA EM UC7 ultratome, followed by staining with uranyl acetate and alkaline lead citrate (each 5 to 10 min) and analysis under a Hitachi Model H-7650 TEM (HITACHI, Tokyo, Japan).
For SEM, the specimens were fixed and post-fixed as described above for TEM. Then, they were transferred into ethanol:iso-amyl acetate (1:1 v/v) for 30 min, and iso-amyl acetate overnight. After dehydration in a Hitachi Model HCP-2 critical point dryer (HITACHI, Tokyo, Japan) with liquid CO2, the specimens were coated with gold-palladium on a Hitachi Model E-1010 ion sputter (4–5 min) before analysis on a Hitachi Model TM-1000 SEM.
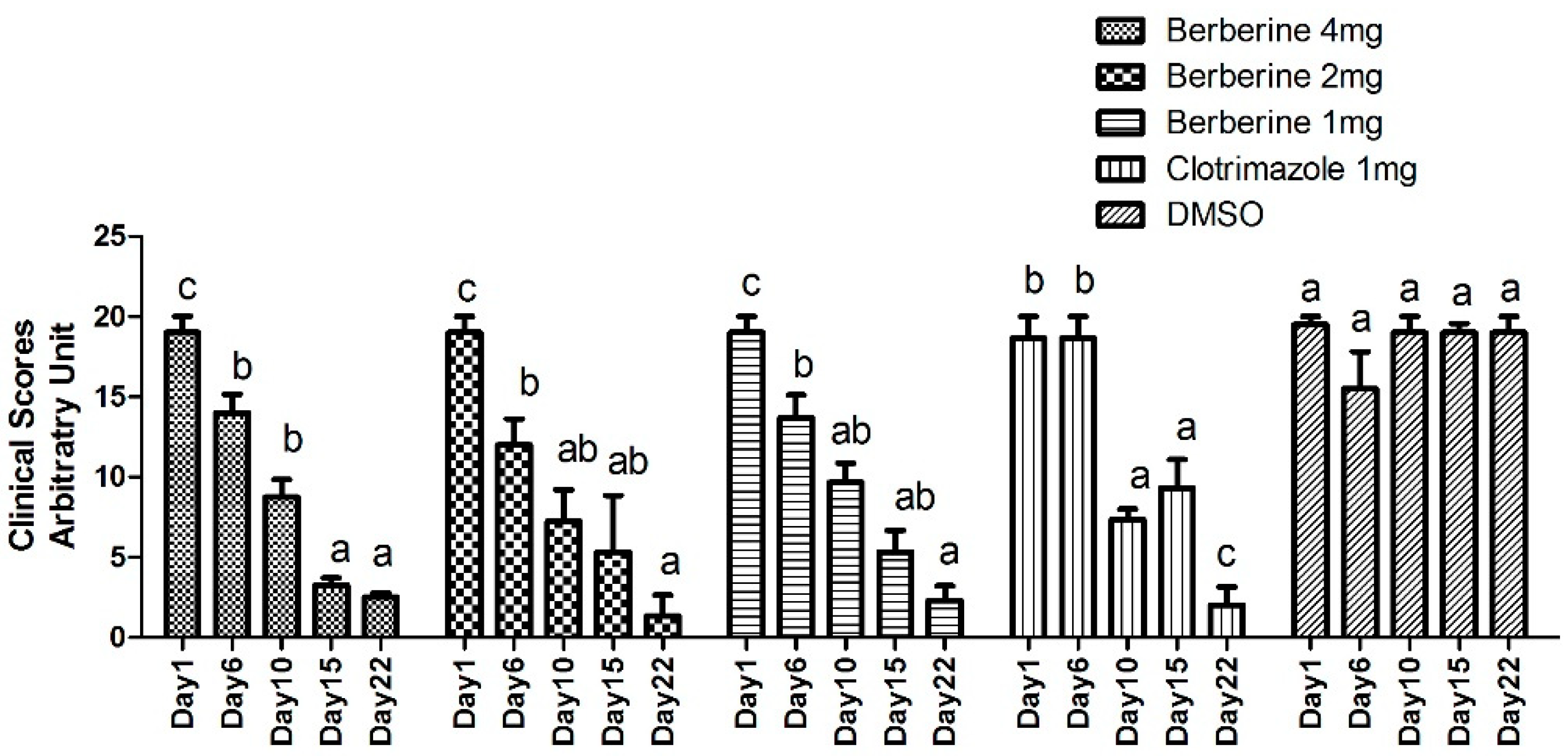
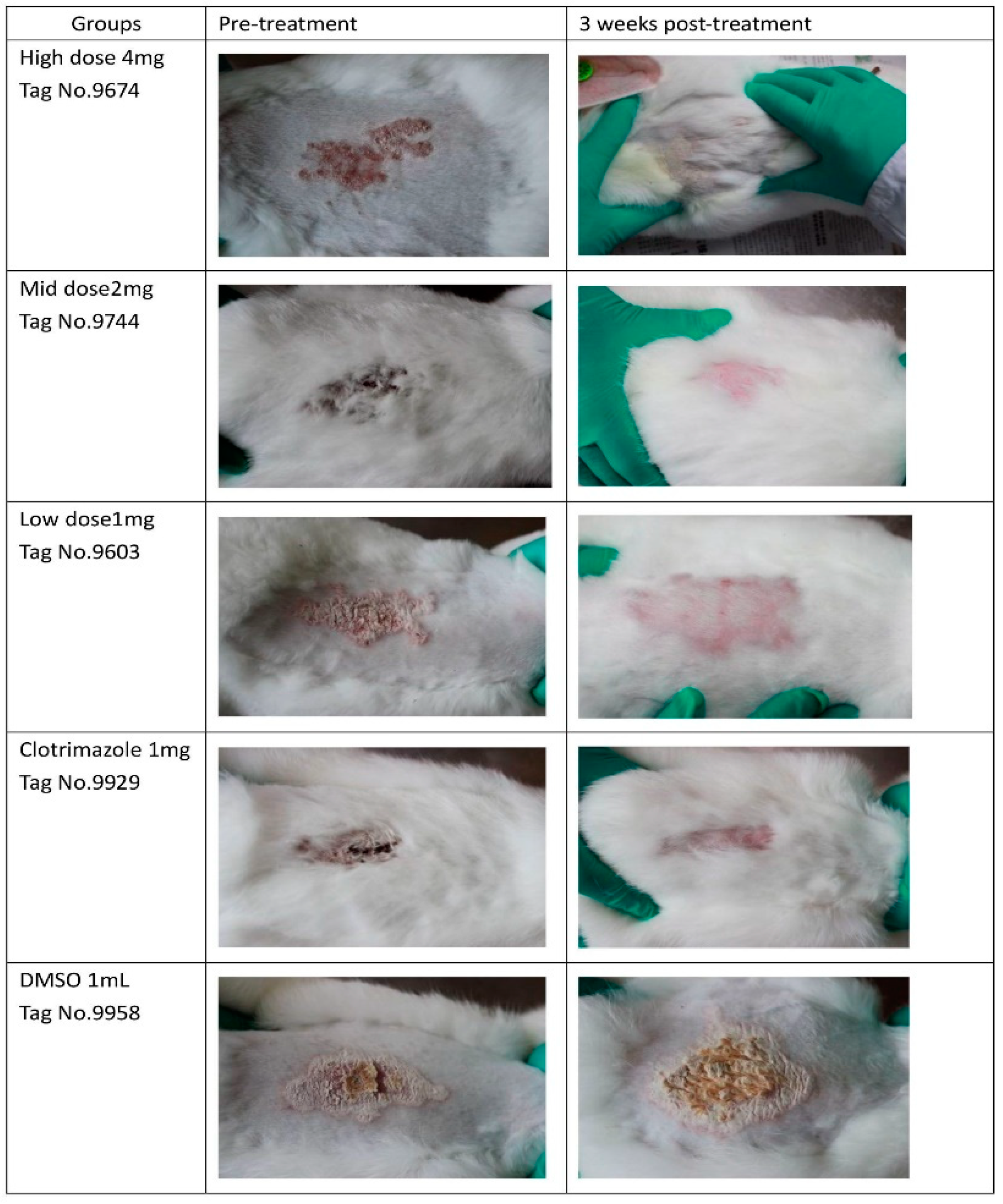
4.4. Experimental Animals and In Vivo Antifungal Assay
Twenty five New Zealand white male rabbits aged 45 days, weighing 800–950 g, were obtained from the experimental animal center at Zhejiang University, Hangzhou, China. The study was performed after approval from the Bioethics Committee of Zhejiang Academy of Agricultural Sciences (NO.320569), strictly following existing guidelines. The animals were assigned to five groups (n = 5). Three groups treated with berberine hydrochloride were classified as Group 1 (4 mg), Group 2 (2 mg), and Group 3 (1 mg). Two additional groups were designated as positive (PC; clotrimazole at 1 mg) and negative (NC; 1 mL DMSO) control groups, respectively.
Dermatophytosis was induced as described in a previous report [
43]. Briefly, a 1-mL suspension (1.0 × 10
6 cells) of
T. mentagrophytes was used for infection for 3 consecutive days (Day 1, 2, 3). Different doses of berberine (4 mg, 2 mg, 1 mg) were applied with solution form to groups 1, 2, and 3 from Day 4 to Day 6, for 3 consecutive days, they are applied by sprayed. Clotrimazole (1 mg) or DMSO (1 mL) was applied topically in parallel (
Table 8). The animals were assessed for symptoms for 22 days, according to a previously described methodology [
44]. In this evaluation, the infected area of skin from each rabbit was divided into four equal quadrants and each area was scored as follows: 0, normal; 1, slightly erythematous patches; 2, well-defined redness, swelling, with bristling hairs, bald patches, or scaly areas; 3, large areas of marked redness, scaling, exposed patches, or ulceration in places; 4, partial damage to the covering and loss of hair; and 5, extensive damage to the covering and complete loss of hair. The scores from various treatment groups were compared. The results were compared using a one-way ANOVA and Tukey’s HSD test. A
p value < 0.05 was considered statistically significant. Photos from each group were taken before treatment and 3 three weeks post-treatment.
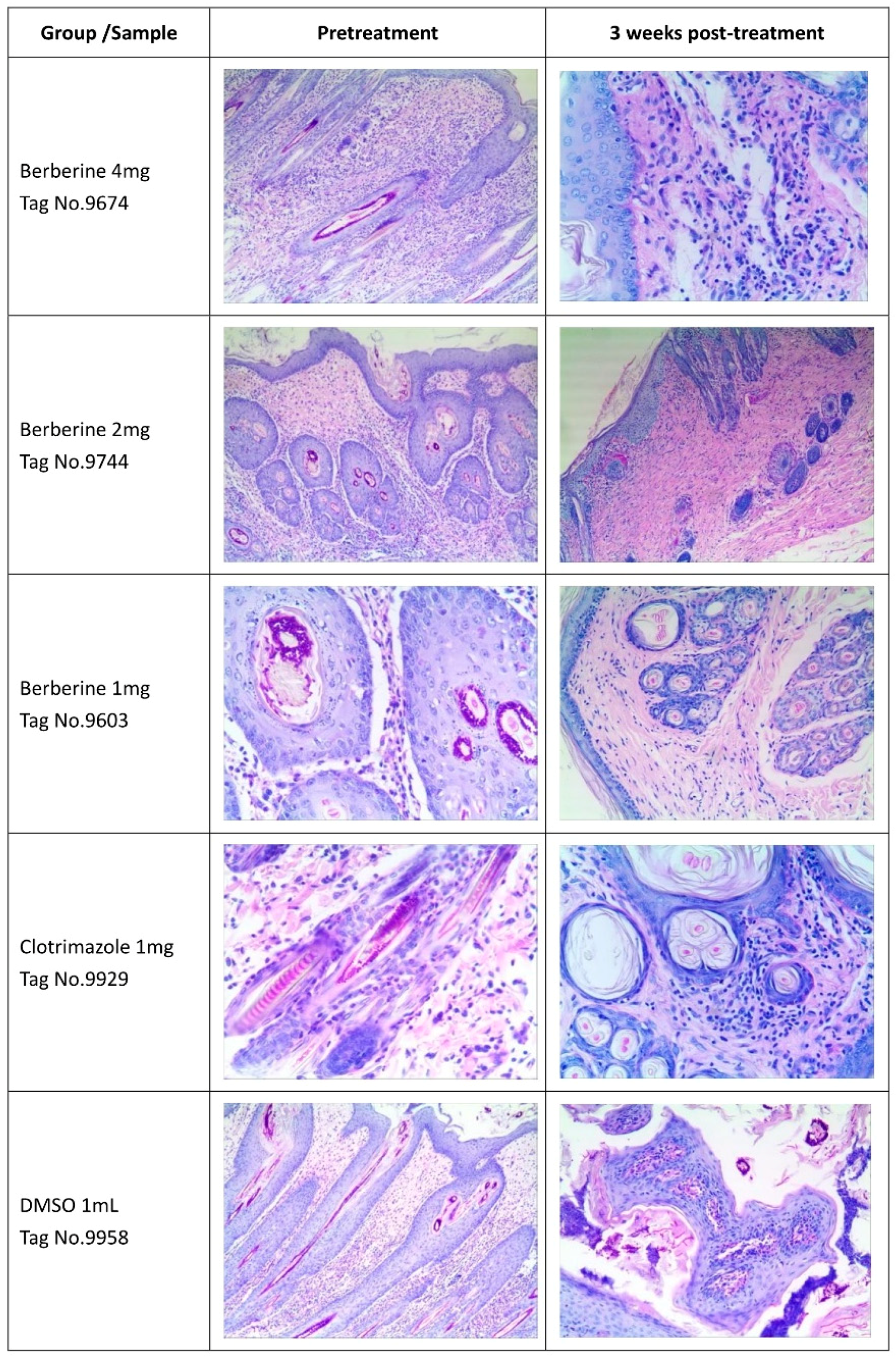
4.5. Skin Histology PAS Staining
Three skin tissue specimens were randomly obtained at the end of the study from each group, fixed and paraffin embedded, and assessed histologically and after PAS staining [
45].
4.6. Berberine Hydrochloride Treatment for Transcriptome Analysis
1.0 × 107 CFU in 1 mL normal saline were used in in vitro cultures. BC and clotrimazole treatment groups (n = 3) were submitted to 1/2 minimum inhibitory concentrations (MICs) of respective drugs (i.e., 1 mg/mL BC and 0.5 μg/mL Clotrimazole, in DMSO); control animals received DMSO. After 6 h of incubation (28 °C, 150 rpm) and filtration (gauze), the samples were frozen for further analysis.
4.7. RNA Purification, cDNA Library Preparation and Illumina Sequencing for Transcriptome Analysis
TRIzol Reagent (Invitrogen) was used for total RNA extraction, as directed by the manufacturer. Then, mRNA was obtained with oligo (dT) magnetic beads and fragmented using fragmentation buffer (70 °C, 4 min). SuperScript II was used to synthesize first strand cDNA; second strand cDNA was obtained after treatment with DNA polymerase I and RNaseH. Double-stranded cDNAs were incubated with T4 DNA polymerase, Klenow Enzyme (NEB), and T4 polynucleotide kinase (NEB). Then, single A base was added with Klenow exo-polymerase; ligation was performed with Adapter by DNA ligase (NEB). Ligation products were submitted to PCR and purified with QIAquick PCR Purification Kit (Qiagen, Dusseldorf, Germany) for the generation of the cDNA library, quantitatively assessed with Qubit (Invitrogen). A cluster of DNA fragments was amplified using bridge PCR on a flow cell chip. After multiple amplification rounds, the products were sequenced on an Illumina HiSeq 4000.
4.8. Assembly and Gene Identification
Reads were assembled with the Trinity software (
http://trinityrnaseq.sourceforge.net/) as previously described [
46]. Contig generation was performed by extension based on overlapping sequence pairs. Then, contigs were joined into transcripts. The longest were considered to be unigenes. The raw transcriptome data have been deposited in the Sequence Read Archive of the National Center for Biotechnology Information (NCBI) (accession No. GSE80604 and SRA: SRP073785).
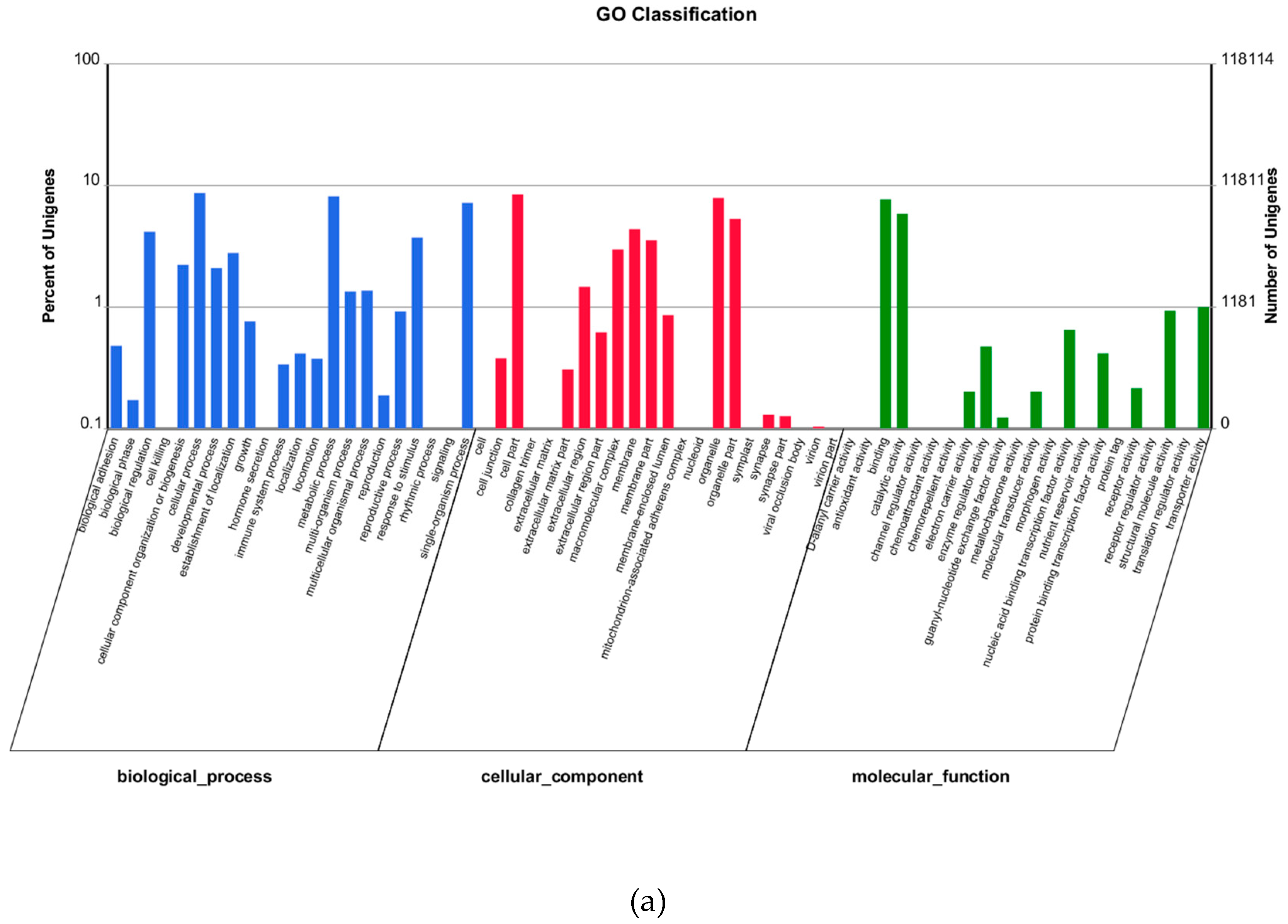
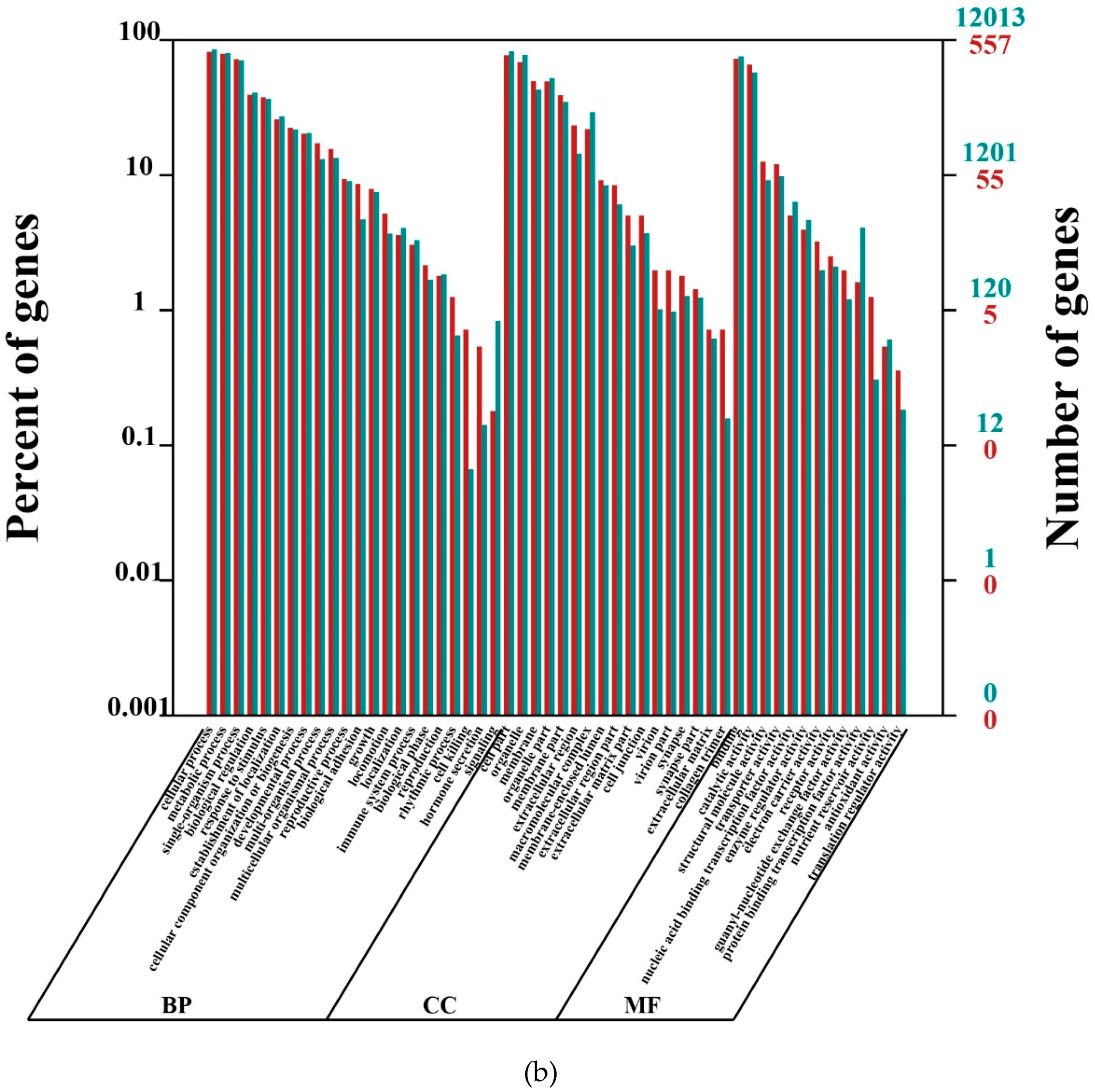
4.9. Functional Annotation for Unigenes
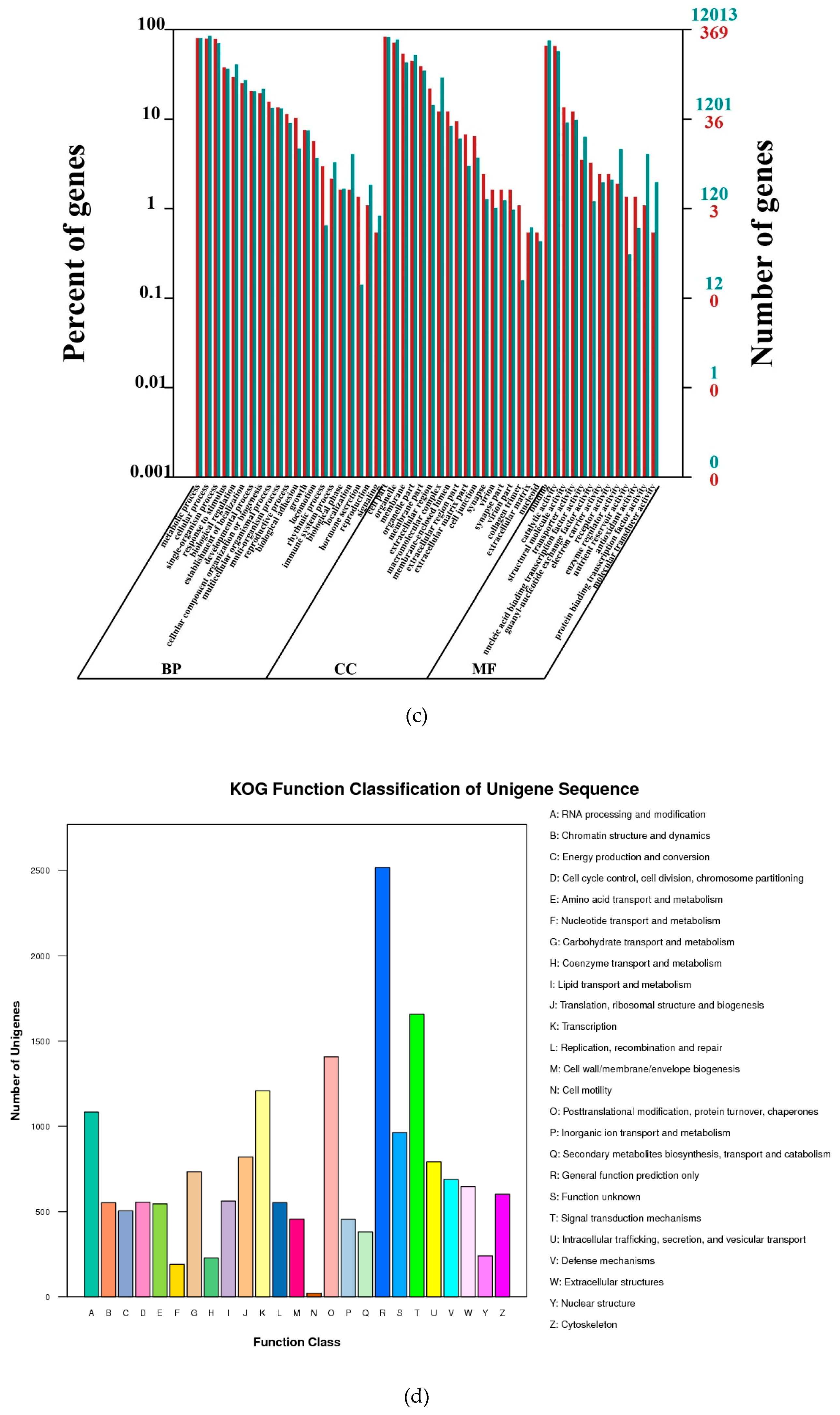
Nt and SwissProt were used for unigene alignment with BLAST (cut-off e-value = 10−5). Blast2GO was utilized for Gene ontology (GO) annotation. KOG and KEGG pathway annotations were also carried out with BLAST against Cluster of Orthologous Groups databases and Kyoto Encyclopedia of Genes and Genomes (cut-off e-value = 10−5).
4.10. Identification of Microsatellites
Unigenes ≥ 1 Kb were submitted to SSR assessment using the Microsatellite Identification tool (MISA;
http://pgrc.ipkgatersleben.de/misa/). Detection criteria were perfect repeat motifs of 1–6 bp, with minimum repeat numbers of 10, 6, 5, 5, 5 and 5 for mono-, di-, tri-, tetra-, penta- and hexa-nucleotide microsatellites, respectively.
4.11. Identification of Differentially Expressed Genes
Unigene abundance was assessed by mapping the reads from 9 specimens against the reference unigene set with the RSEM software (University of Wisconsin-Madison, Madison, WI, USA) on Bowtie2 [
47]. Relative transcript abundance rates were expressed as FPKM values (fragments per kilobase of exon per million mapped reads) [
48], derived as R~109C/NL(C is the number of mappable reads falling into the specific unigene; N is the total number of mappable reads; L is the unigene length. Differentially expressed genes were identified by the DEseq software as described previously, with false discovery rate (FDR) correction [
49]. Genes were considered to be differentially expressed with Qvalue < 0.05 and |log 2 ratio| > 1.
4.12. Real-time PCR
First strand cDNA was obtained from total RNA (1 µg) with a kit from Promega (Madison, WI, USA) as directed by the manufacturer. Quantitative RT-PCR was performed on an ABI StepOnePlus (Applied Biosystems, Thermo Fisher Scientific, Waltham, MA, USA) with SYBR Green Supermix (TaKaRa) as proposed by the manufacturer. Amplification was performed at 94 °C (10 min), followed by 40 cycles of 94 °C (15 s), 60 °C (31 s), and 72 °C (1 min). Primers are described in
Table 9. Cycle threshold (CT) values were obtained, and data analyzed by the 2
−ΔΔCt method [
50]. Normalization was carried out with 18S, as an endogenous housekeeping gene. Relative mRNA expression levels were provided as mean ± SD from triplicate experiments.
4.13. Statistic Analysis
Group comparison was performed by Duncan’s test. GraphPad Prism 5 (GraphPad Software, San Diego, CA, USA) was used for statistical analyses; statistical significance was set at p < 0.05. Pearson correlation were used to statistic analysis, determining the agreement between qRT-PCR analysis and Illumina RNA-seq analysis.
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}