Silibinin Downregulates the NF-κB Pathway and NLRP1/NLRP3 Inflammasomes in Monocytes from Pregnant Women with Preeclampsia

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Clinical Characteristics
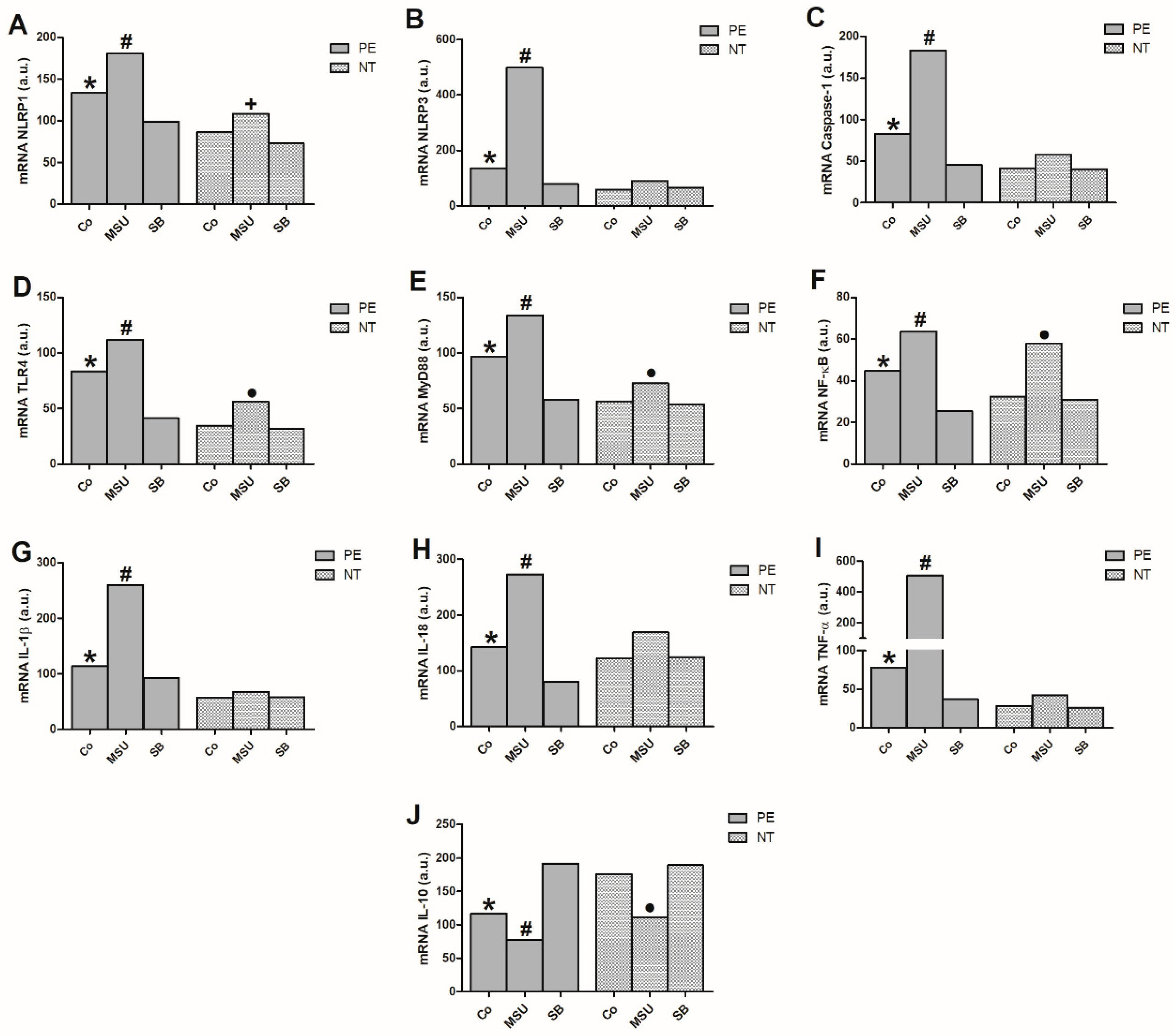
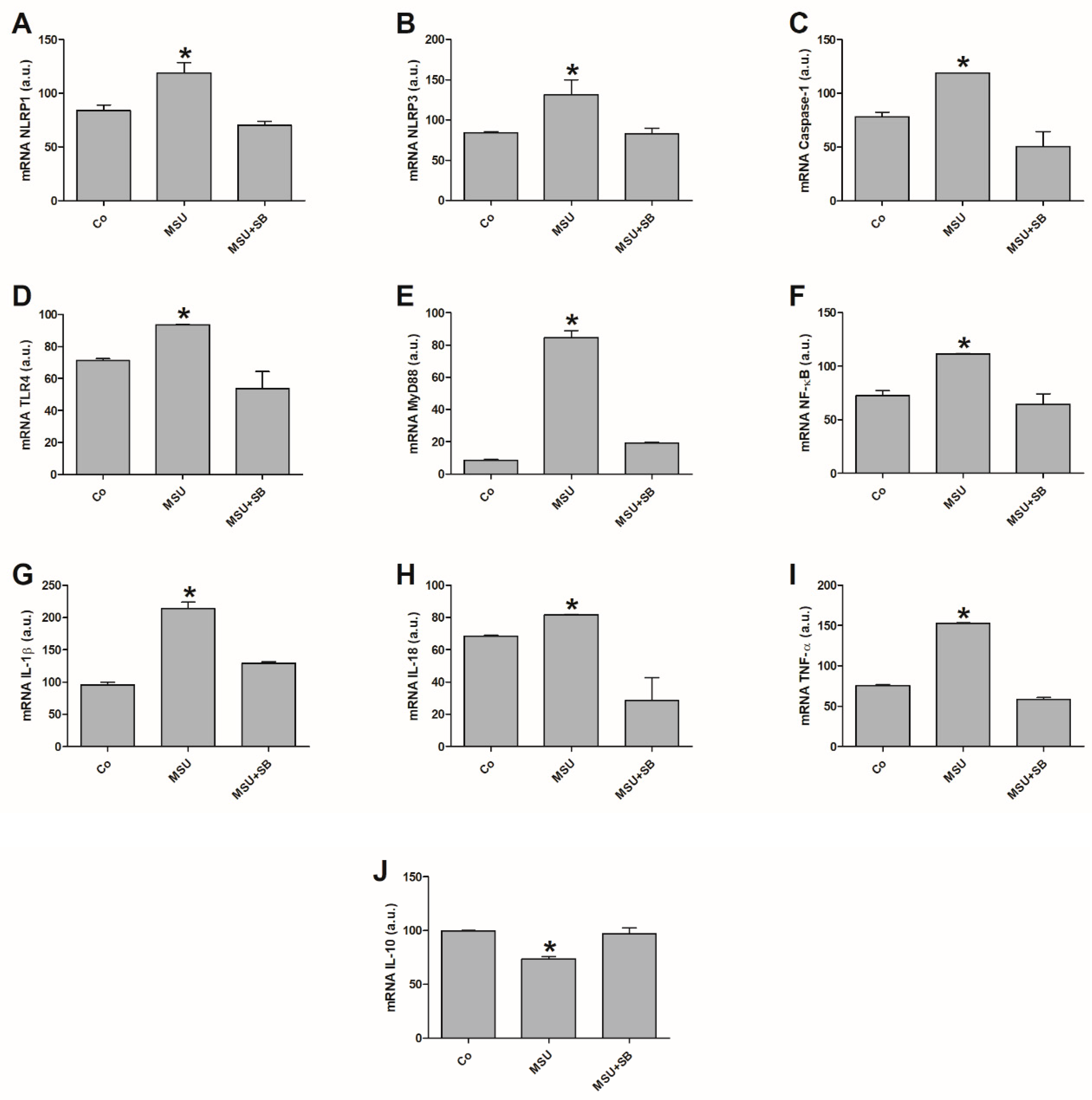
2.2. Gene Expression in Monocytes Cultured with MSU or SB
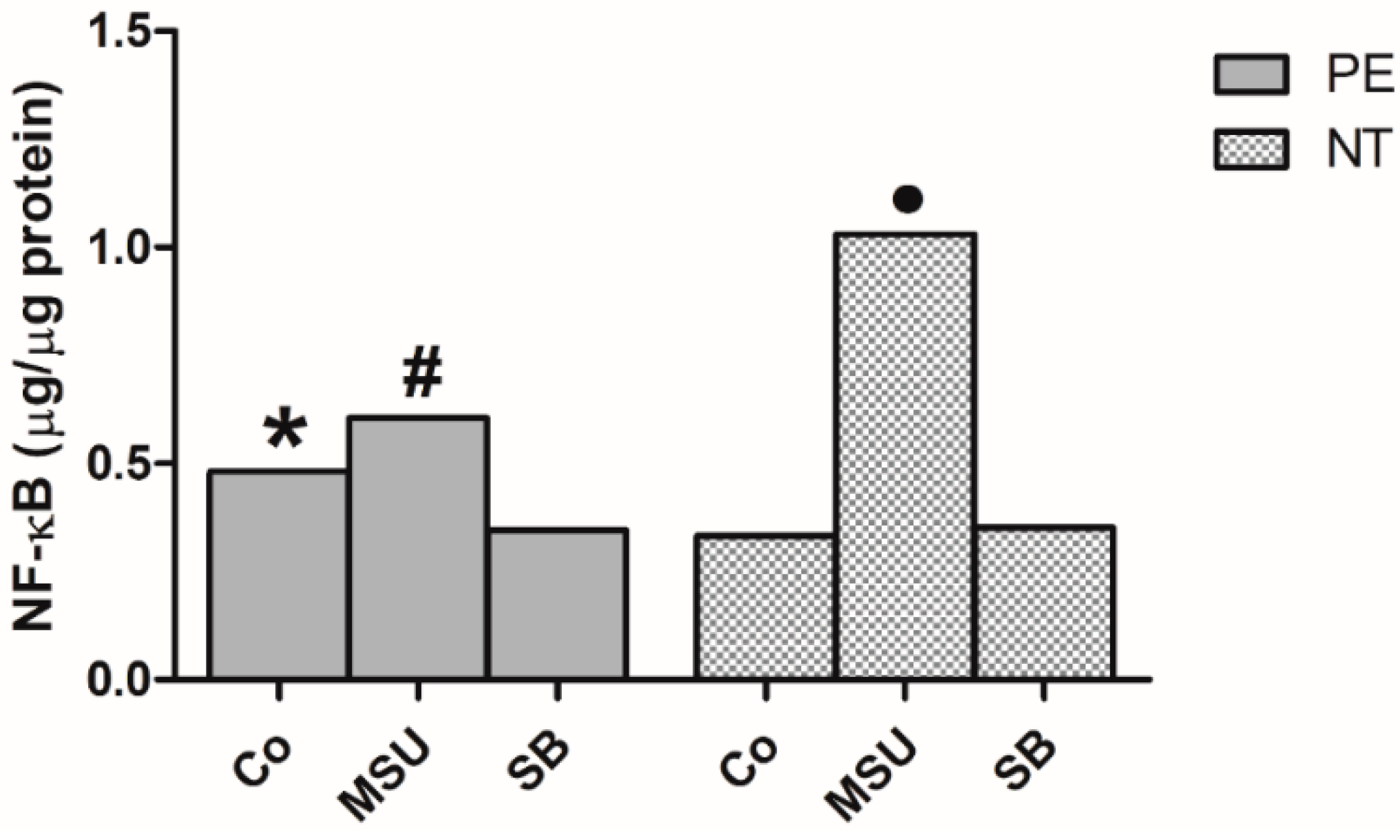
2.3. Determination of NF-κB in Monocyte Nuclear Extracts
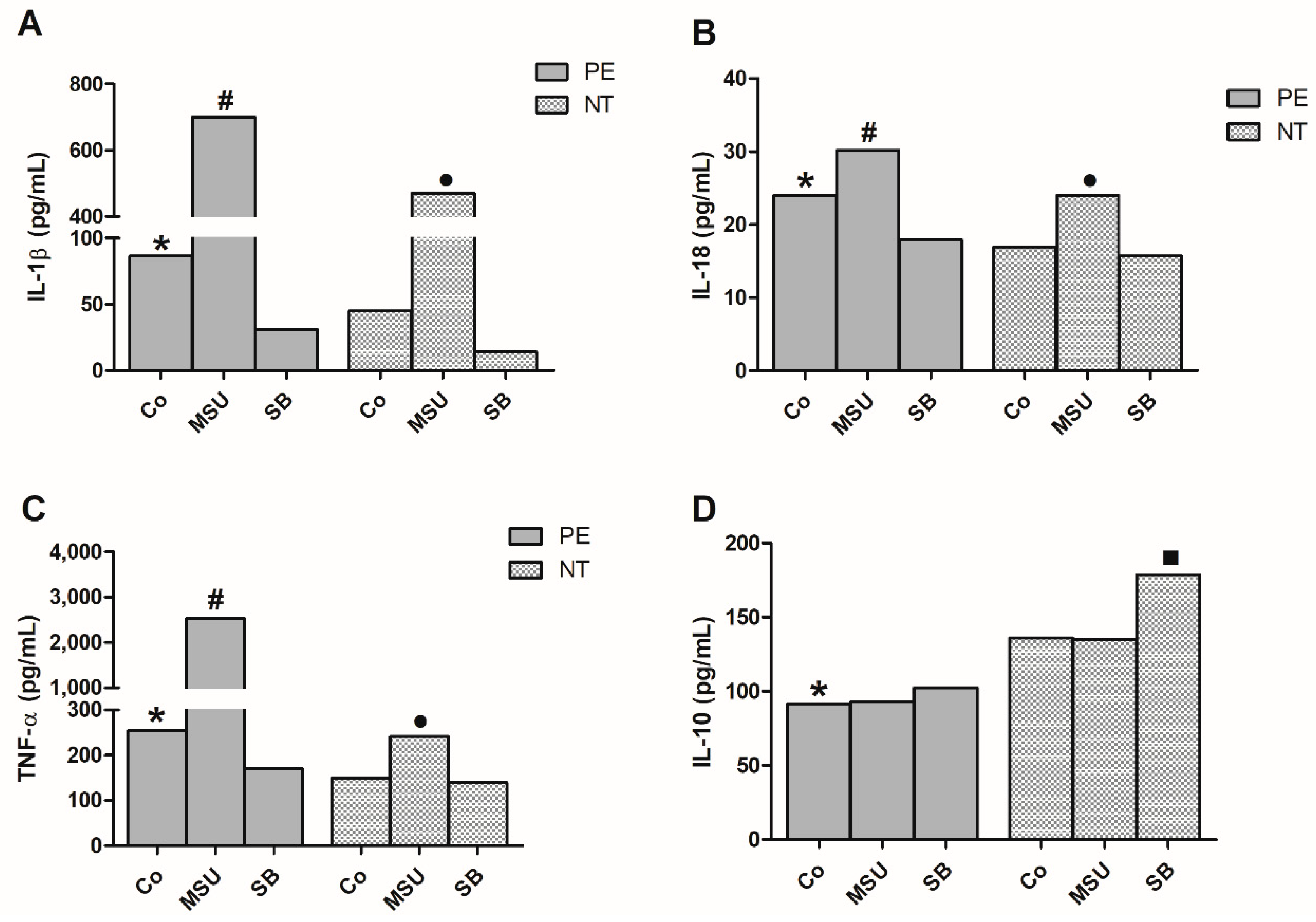
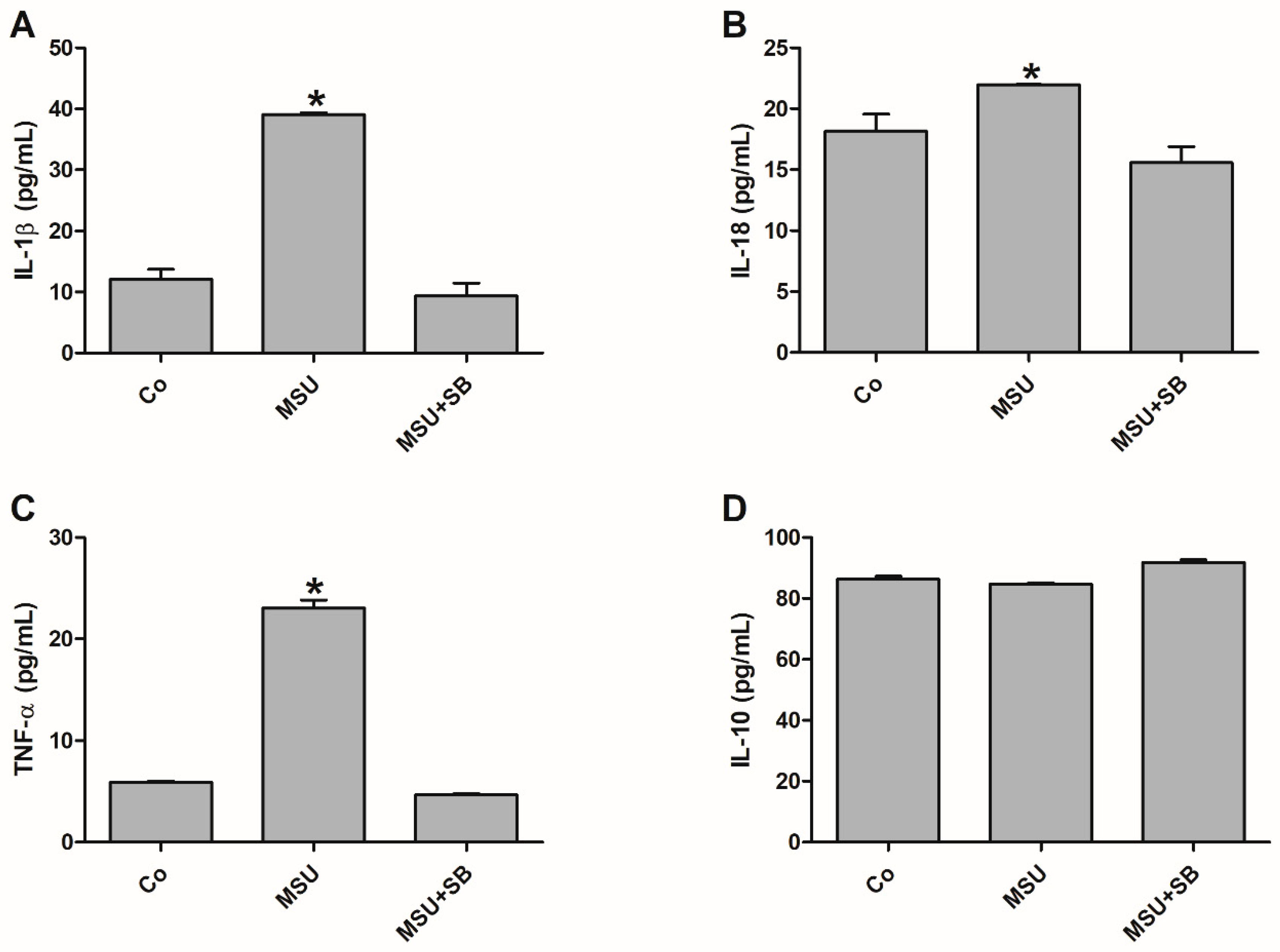
2.4. Cytokine Production by Monocytes from Pregnant Women
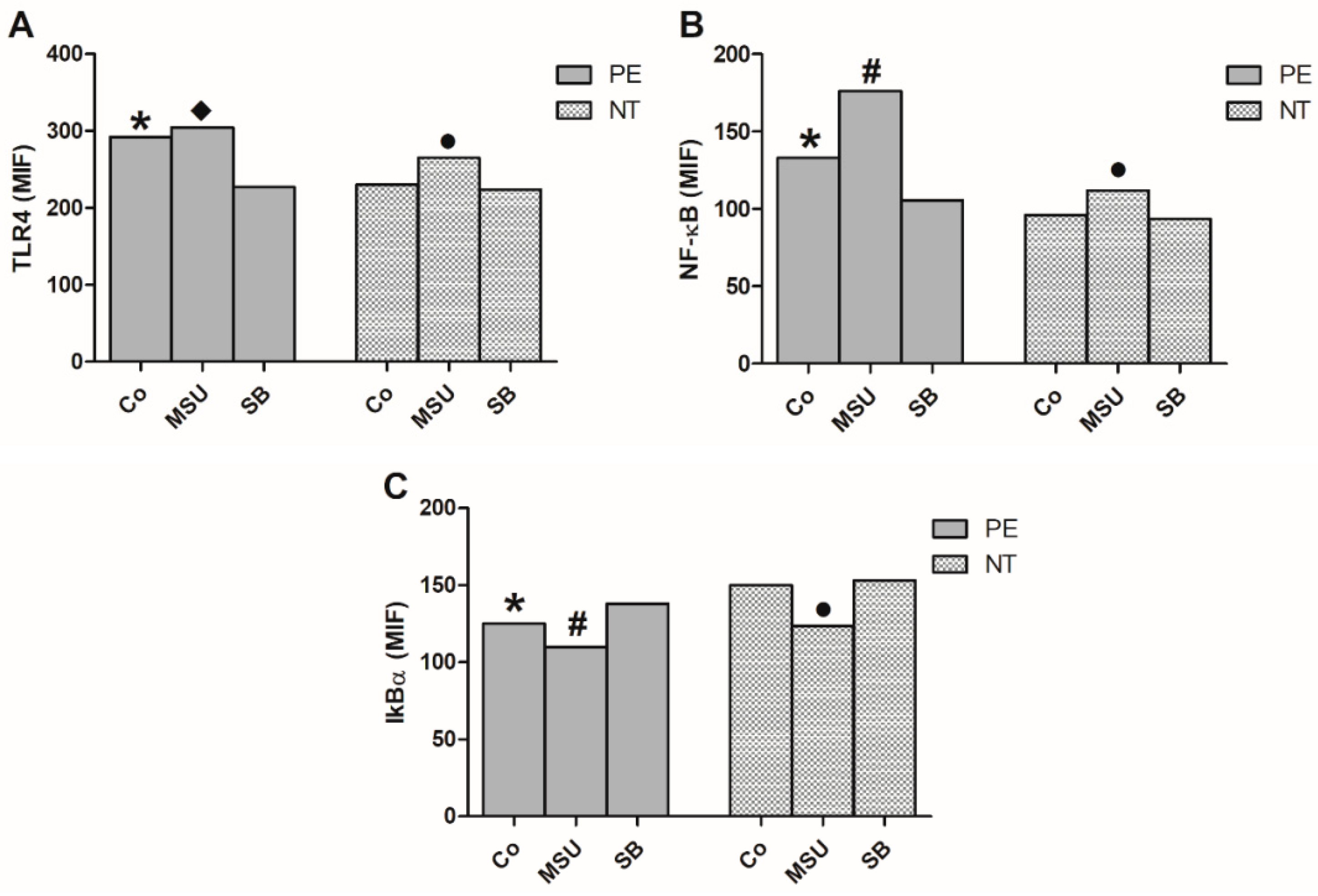
2.5. Analysis of the Expression of NF-κB Pathway in Monocytes
2.6. Gene Expression in THP-1 Cells Cultured with MSU or SB
2.7. Cytokine Production by THP-1 cells Cultured with MSU and/or SB
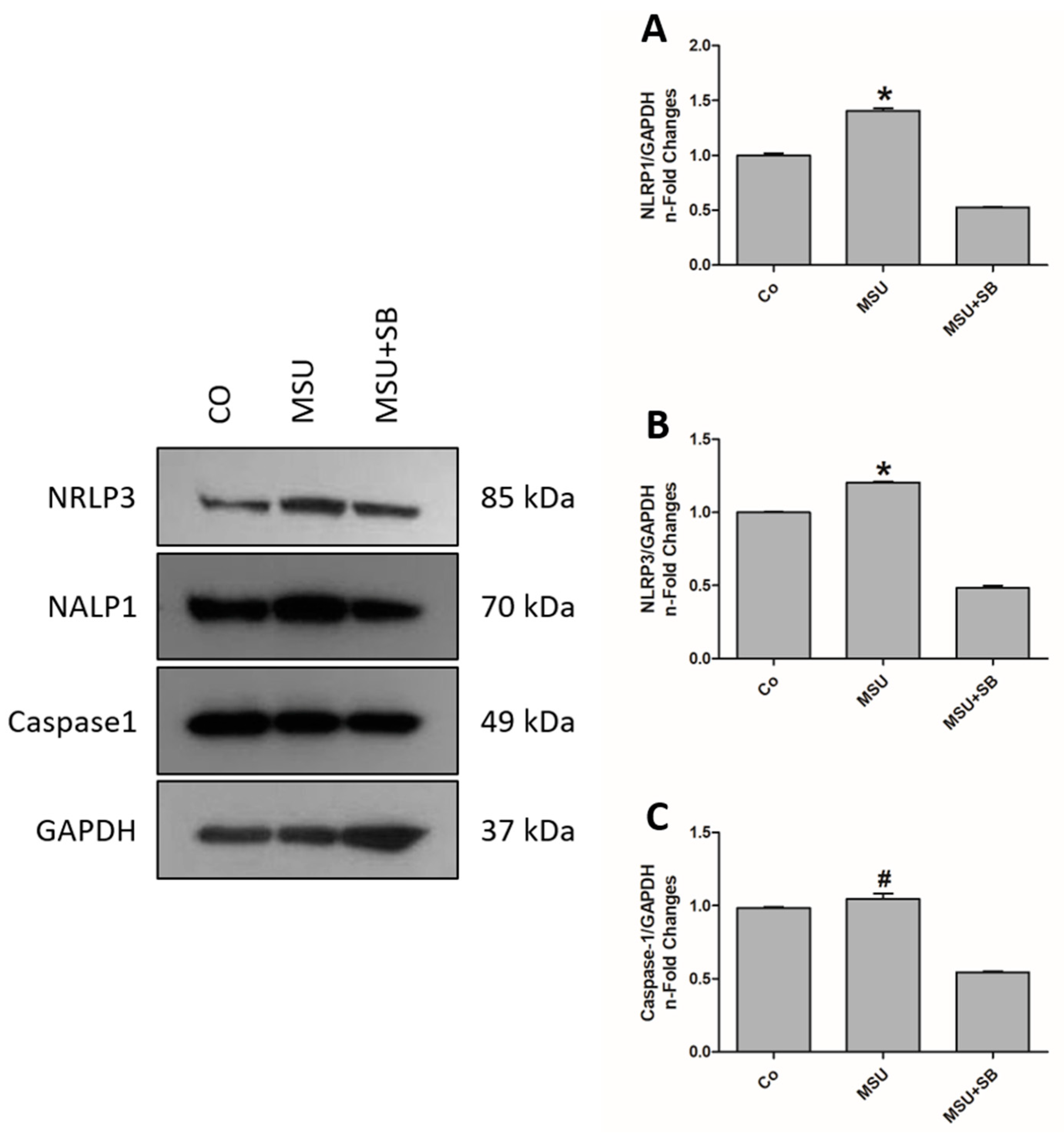
2.8. Inflammasomes Protein Componentes in THP-1 Cells Cultured with MSU and/or SB
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Blood Sampling
4.3. Monocyte Cultures
4.4. THP-1 Cell culture
4.5. Nuclear Extraction of Monocytes
4.6. Determination of p65NF-κB Activity
4.7. Cytokine Determinations
4.8. Expression of Transcripts Related to Inflammation
4.9. Expression of Intracytoplasmic Factors and TLR4 Receptor in Monocytes
4.10. Evaluation of Proteins Related to Inflammasomes in THP-1 Cells
4.11. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Dixit, N.; Baboota, S.; Kohli, K.; Ahmad, S.; Ali, J. Sylimarin: A review of pharmacological aspects and bioavailabity enhacement approaches. Indian J. Pharmacol. 2007, 39, 172–179. [Google Scholar] [CrossRef]
- Abenavoli, L.; Izzo, A.A.; Milić, N.; Cicala, C.; Santini, A.; Capasso, R. Milk thistle (Silybum marianum): A concise overview on its chemistry, pharmacological, and nutraceutical uses in liver diseases. Phytother. Res. 2018, 32, 2202–2213. [Google Scholar] [CrossRef] [PubMed]
- Bannwart, C.F.; Nakaira-Takahagi, E.; Golim, M.A.; Medeiros, L.T.L.; Romão, M.; Weel, I.C.; Peraçoli, M.T. Downregulation of nuclear factor kappa B (NF-κB) pathway by silibinin in human monocytes challenged with Paracoccidioides brasiliensis. Life Sci. 2010, 86, 880–886. [Google Scholar] [CrossRef] [PubMed]
- Bannwart, C.F.; Peraçoli, J.C.; Nakaira-Takahagi, E.; Peraçoli, M.T. Inhibitory effect of silibinin on tumour necrosis factor-alpha and hydrogen peroxide production by human monocytes. Nat. Prod. Res. 2010, 24, 1747–1757. [Google Scholar] [CrossRef]
- Manna, S.K.; Mukhopadhyay, A.; Van, N.T.; Aggarwal, B.B. Silymarin suppress TNF-induced activation of NF-kB, c-Jun N-terminal kinase, and apoptosis. J. Immunol. 1999, 163, 6800–6809. [Google Scholar]
- Esmaeil, N.; Anaraki, S.B.; Gharagozloo, M.; Moayedi, B. Silymarin impacts on immune system as an immunomodulator: One key for many locks. Int. Immunopharmacol. 2017, 50, 194–201. [Google Scholar] [CrossRef]
- Sibai, B.; Dekker, G.; Kupferminc, M. Pre-eclampsia. Lancet 2005, 365, 785–799. [Google Scholar] [CrossRef]
- Ghulmiyyah, L.; Sibai, B. Maternal mortality from preeclampsia/eclampsia. Semin. Perinatol. 2012, 36, 56–59. [Google Scholar] [CrossRef]
- American College of Obstetricians and Gynecologists; Task Force on Hypertension in Pregnancy. Hypertension in pregnancy: Report of the American College of Obstetricians and Gynecologists’ Task Force on Hypertension in Pregnancy. Obstet. Gynecol. 2013, 122, 1122–1131. [Google Scholar]
- Tranquilli, A.L.; Dekker, G.; Magee, L.; Roberts, J.; Sibai, B.M.; Steyn, W.; Zeeman, G.G.; Brown, M.A. The classification, diagnosis and management of the hypertensive disorders of pregnancy: A revised statement from the ISSHP. Pregnancy Hypertens. 2014, 4, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Mol, B.W.; Roberts, C.T.; Thangaratinam, S.; Magee, L.A.; Groot, C.G.; Hofmeyr, G.J. Pre-eclampsia. Lancet 2016, 387, 999–1011. [Google Scholar] [CrossRef]
- Luppi, P.; Deloia, J.A. Monocytes of preeclamptic women spontaneously synthesize pro-inflammatory cytokines. Clin. Immunol. 2006, 118, 268–275. [Google Scholar] [CrossRef]
- Peraçoli, J.C.; Rudge, M.V.C.; Peraçoli, M.T. Tumor necrosis fator-alpha in gestation and pueperium of women with gestational hypertension and pre-eclampsia. Am. J. Reprod Immunol. 2007, 57, 177–185. [Google Scholar] [CrossRef]
- Cristofalo, R.; Bannwart-Castro, C.F.; Magalhães, C.G.; Borges, V.T.; Peraçoli, J.C.; Witkin, S.S.; Peraçoli, M.T. Silibinin attenuates oxidative metabolism and cytokine production by monocytes from preeclamptic women. Free Radic. Res. 2013, 47, 268–275. [Google Scholar] [CrossRef]
- Peraçoli, M.T.S.; Bannwart, C.F.; Cristofalo, R.; Borges, V.T.M.; Costa, R.A.A.; Witkin, S.S.; Peraçoli, J.C. Increased reactive oxygen species and tumor necrosis factor-alpha production by monocytes are associated with elevated levels of uric acid in pre-eclamptic women. Am. J. Reprod Immunol. 2011, 66, 460–467. [Google Scholar] [CrossRef]
- Livingston, J.R.; Payne, B.; Brown, M.; Roberts, J.M.; Côté, A.M.; Magee, L.A.; von Dadelszen, P.; PIERS Study Group. Uric acid as a predictor of adverse maternal and perinatal outcomes in women hospitalized with preeclampsia. J. Obstet. Gynaecol. Can. 2014, 36, 870–877. [Google Scholar] [CrossRef]
- Zhao, J.; Zheng, D.Y.; Yang, J.M.; Wang, M.; Zhang, X.T.; Sun, L.; Yun, X.G. Maternal serum uric acid concentration is associated with the expression of tumour necrosis factor-α and intercellular adhesion molecule-1 in patients with preeclampsia. J. Hum. Hypertens. 2016, 30, 456–462. [Google Scholar] [CrossRef]
- Martinon, F.; Petrilli, V.; Mayor, A.; Tardivel, A.; Tshopp, J. Gout-associated uric acid crystals activate the NALP3 inflammasome. Nature 2006, 440, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Kingsbury, S.R.; Conaghan, P.G.; McDermott, M. The role of the NLRP3 inflammasome in gout. J. Inflamm. Res. 2011, 4, 39–49. [Google Scholar] [Green Version]
- Stutz, A.; Kolbe, C.C.; Stahl, R.; Horvath, G.L.; Franklin, B.S.; van Ray, O.; Brinkschulte, R.; Geyer, M.; Meissner, F.; Latz, E. NLRP3 inflammasome assembly is regulated by phosphorylation of the pyrin domain. J. Exp. Med. 2017, 214, 1725–1736. [Google Scholar] [CrossRef]
- Schroder, K.; Tschopp, J. The inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef]
- Jo, E.K.; Kim, J.K.; Shin, D.M.; Sasakawa, C. Molecular mechanisms regulating NLRP3 inflammasome activaction. Cell Mol. Immunol. 2016, 13, 148–159. [Google Scholar] [CrossRef]
- Giorgi, V.S.; Peraçoli, M.T.; Peraçoli, J.C.; Witkin, S.S.; Bannwart-Castro, C.F. Silibinin modulates the NF-κB pathway and pro-inflammatory cytokine production by mononuclear cells from preeclamptic women. J. Reprod. Immunol. 2012, 95, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Matias, M.L.; Romão, M.; Weel, I.C.; Ribeiro, V.R.; Nunes, P.R.; Borges, V.T.; Araújo, J.P., Jr.; Peraçoli, J.C.; de Oliveira, L.; Peraçoli, M.T. Endogenous and uric acid-induced activation of NLRP3 inflammasome in pregnant women with preeclampsia. PLoS ONE 2015, 10, e0129095. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.O.; Peraçoli, M.T.; Weel, I.C.; Bannwart, C.F.; Romão, M.; Nakaira-Takahagi, E.; Medeiros, L.T.; Silva, M.G.; Peraçoli, J.C. Hepatoprotective and anti-inflammatory effects of silibinin on experimental preeclampsia induced by L-NAME in rats. Life Sci. 2012, 91, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Borzychowski, A.M.; Sargent, I.L.; Redman, C.W. Inflammation and pre-eclampsia. Semin. Fetal Neonatal Med. 2006, 11, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Romão-Veiga, M.; Matias, M.L.; Ribeiro, V.R.; Nunes, P.R.; Borges, V.T.M.; Peraçoli, J.C.; Peraçoli, M.T.S. Induction of systemic inflammation by hyaluronan and hsp70 in women with pre-eclampsia. Cytokine 2018, 105, 23–31. [Google Scholar] [CrossRef]
- Chen, W.; Qian, L.; Wu, F.; Li, M.; Wang, H. Significance of toll-like receptor 4 signaling in peripheral blood monocytes of pre-eclamptic patients. Hypertens Pregnancy 2015, 34, 486–494. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, L.T.; Peraçoli, J.C.; Bannwart-Castro, C.F.; Romão, M.; Weel, I.C.; Golim, M.A.; de Oliveira, L.G.; Kurokawa, C.S.; Borges, V.T.M.; Peraçoli, M.T. Monocytes from pregnant women with pre-eclampsia are polarized to a M1 phenotype. Am. J. Reprod. Immunol. 2014, 72, 5–13. [Google Scholar] [CrossRef]
- Khan, R.N.; Hay, D.P. A clear and present danger: Inflammasomes DAMPing down disorders of pregnancy. Hum. Reprod. Update 2015, 21, 388–405. [Google Scholar] [CrossRef]
- Peraçoli, J.C.; Bannwart-Castro, C.F.; Romão, M.; Weel, I.C.; Ribeiro, V.R.; Borges, V.T.; Rudge, M.V.; Witkin, S.S.; Peraçoli, M.T. High levels of heat shock protein 70 are associated with pro-inflammatory cytokines and may differentiate early- from late-onset preeclampsia. J. Reprod. Immunol. 2013, 100, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Romão, M.; Weel, I.C.; Lifshitz, S.J.; Peraçoli, M.T.; Witkin, S. Elevated hyaluronan and extracellular matrix metalloproteinase inducer levels in women with preeclampsia. Arch. Gynecol. Obstet. 2014, 289, 575–579. [Google Scholar] [CrossRef]
- Naruse, K.; Sado, T.; Noguchi, T.; Tsunemi, T.; Yoshida, S.; Akasaka, J.; Koike, N.; Oi, H.; Kobayashi, H. Peripheral RAGE (receptor for advance glycation endproducts)-ligands in normal pregnancy and preeclampsia: Novel markers of inflammatory response. J. Reprod. Immunol. 2012, 93, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Afonina, I.S.; Zhong, Z.; Karin, M.; Beyaert, R. Limiting inflammation-the negative regulation of NF-κB and the NLRP3 inflammasome. Nat. Immunol. 2017, 18, 861–869. [Google Scholar] [CrossRef]
- Striz, I.; Brabcova, E.; Kolesar, L.; Liu, X.D.; Brabcova, I.; Sekerkova, A.; Poole, J.A.; Jaresova, M.; Slavcev, A.; Rennard, S.I. Epithelial cells modulate genes associated with NF kappa B activation in co-cultured human macrophages. Immunobiology 2011, 216, 1110–1116. [Google Scholar] [CrossRef] [Green Version]
- Vallabhapurapu, S.; Karin, M. Regulation and function of NF-kappaB transcription factors in the immune system. Annu. Rev. Immunol. 2009, 27, 693–733. [Google Scholar] [CrossRef]
- Moore, K.W.; de Waal Malefyt, R.; Coffman, R.L.; O’Garra, A. Interleukin-10 and the interleukin-10 receptor. Annu. Rev. Immunol. 2001, 19, 683–765. [Google Scholar] [CrossRef] [PubMed]
- Cubro, H.; Kashyap, S.; Nath, M.C.; Ackerman, A.W.; Garovic, V.D. The role of interleukin-10 in the pathophysiology of preeclampsia. Curr. Hypertens. Rep. 2018, 20, 36. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.R.; Seo, H.S.; Ku, J.M.; Kim, G.J.; Jeon, C.Y.; Park, J.H.; Jang, B.H.; Park, S.J.; Shin, Y.C.; Ko, S.G. Silibinin inhibits the production of pro-inflammatory cytokines through inhibition of NF-κB signaling pathway in HMC-1 human mast cells. Inflamm. Res. 2013, 62, 941–950. [Google Scholar] [CrossRef] [Green Version]
- Saurai, P. Silymarin as a natural antioxidant: An overview of the current evidence and perspectives. Antioxidants 2015, 4, 204–247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Wang, B.; Cao, S.; Wang, Y.; Wu, D. Silybin attenuates LPS-induced lung injury in mice by inhibiting NF-κB signaling and NLRP3 activation. Int. J. Mol. Med. 2017, 39, 1111–1118. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Xu, D.; She, L.; Wang, Z.; Yang, N.; Sun, R.; Zhang, Y.; Yan, C.; Wei, Q.; Aa, J.; et al. Silybin inhibits NLRP3 inflammasome assembly through the NAD+/SIRT2 pathway in mice with nonalcoholic fatty liver disease. FASEB J. 2018, 32, 757–767. [Google Scholar] [CrossRef]
- Li, C.Y.; Lam, K.W.; Yam, L.T. Esterases in human leukocytes. J. Histochem. Cytochem. 1973, 21, 1–12. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 93, 265–275. [Google Scholar]
Sample Availability: Samples of the compounds are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Pregnant Women with Preeclampsia (n = 20) | NT Pregnant Women (n = 20) |
|---|---|---|
| Age (years) | 26 (17–41) | 27 (18–40) |
| Gestational age (weeks) | 34 (23–39) | 35 (23–40) |
| Systolic Blood Pressure (mmHg) | 160 * (140–200) | 110 (90–112) |
| Diastolic Blood Pressure (mmHg) | 110 * (90–120) | 69 (63–70) |
| Proteinuria (mg/24 h) | 7250 * (300–18800) | <300 |
| Uric acid (mg/dL) | 6.2 * (3.9–10.1) | 3.2 (2.3–4.7) |
| Gene | Forward Primer (5′–3′) | Reverse Primer (5′–3′) | GenBank |
|---|---|---|---|
| NLRP1 | (1728)TCCGGCTCCCATTAGACAGA(1747) | (1810)AGACCCATCCTGGCTCATCT(1791) | NM_033004.3 |
| NLRP3 | (2826)GAGGAAAAGGAAGGCCGACA(2845) | (2917)TGGCTGTTCACCAATCCATGA(2897) | NM_004895.4 |
| CASP1 | (1065)AGACATCCCACAATGGGCTC(1084) | (1172)TGAAAATCGAACCTTGCGGAAA(1151) | NM_033292.3 |
| TLR4 | (2274)TGCTTCTTGCTGGCTGCATA(2293) | (2359)CCAGTCCTCATCCTGGCTTG(2340) | NM_138554.4 |
| MYD88 | (263)GTCTCCTCCACATCCTCCCT(282) | (344)TCCGCACGTTCAAGAACAGA(325) | NM_001172567.1 |
| NFKB1 | (1072) TGCAGCAGACCAAGGAGATG(1091) | (1211) TGCATTGGGGGCTTTACTGT (1192) | NM_003998.3 |
| IL1B | (544)GAGCAACAAGTGGTGTTCTCC(564) | (653)AACACGCAGGACAGGTACAG(634) | NM_000576.2 |
| IL18 | (438)ACTGTAGAGATAATGCACCCCG(459) | (517)AGTTACAGCCATACCTCTAGGC(496) | NM_001562.3 |
| TNF | (325)GCTGCACTTTGGAGTGATCG(344) | (462)GGGTTTGCTACAACATGGGC(443) | NM_000594.3 |
| IL10 | (361)AAGACCCAGACATCAAGGCG(380) | (445)ATTCGATGACAGCGCCGTAG(426) | NM_000572.2 |
| GAPDH | (684)CGTGGAAGGACTCATGACCA(703) | (801)GGCAGGGATGATGTTCTGGA(782) | NM_002046.4 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matias, M.L.; Gomes, V.J.; Romao-Veiga, M.; Ribeiro, V.R.; Nunes, P.R.; Romagnoli, G.G.; Peracoli, J.C.; Peracoli, M.T.S. Silibinin Downregulates the NF-κB Pathway and NLRP1/NLRP3 Inflammasomes in Monocytes from Pregnant Women with Preeclampsia. Molecules 2019, 24, 1548. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24081548
Matias ML, Gomes VJ, Romao-Veiga M, Ribeiro VR, Nunes PR, Romagnoli GG, Peracoli JC, Peracoli MTS. Silibinin Downregulates the NF-κB Pathway and NLRP1/NLRP3 Inflammasomes in Monocytes from Pregnant Women with Preeclampsia. Molecules. 2019; 24(8):1548. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24081548
Chicago/Turabian StyleMatias, Mariana Leticia, Virginia Juliani Gomes, Mariana Romao-Veiga, Vanessa Rocha Ribeiro, Priscila Rezeck Nunes, Graziela Gorete Romagnoli, Jose Carlos Peracoli, and Maria Terezinha Serrao Peracoli. 2019. "Silibinin Downregulates the NF-κB Pathway and NLRP1/NLRP3 Inflammasomes in Monocytes from Pregnant Women with Preeclampsia" Molecules 24, no. 8: 1548. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24081548