
Silymarin: Friend or Foe of UV Exposed Keratinocytes?

 ,
, 

Abstract
:
1. Introduction
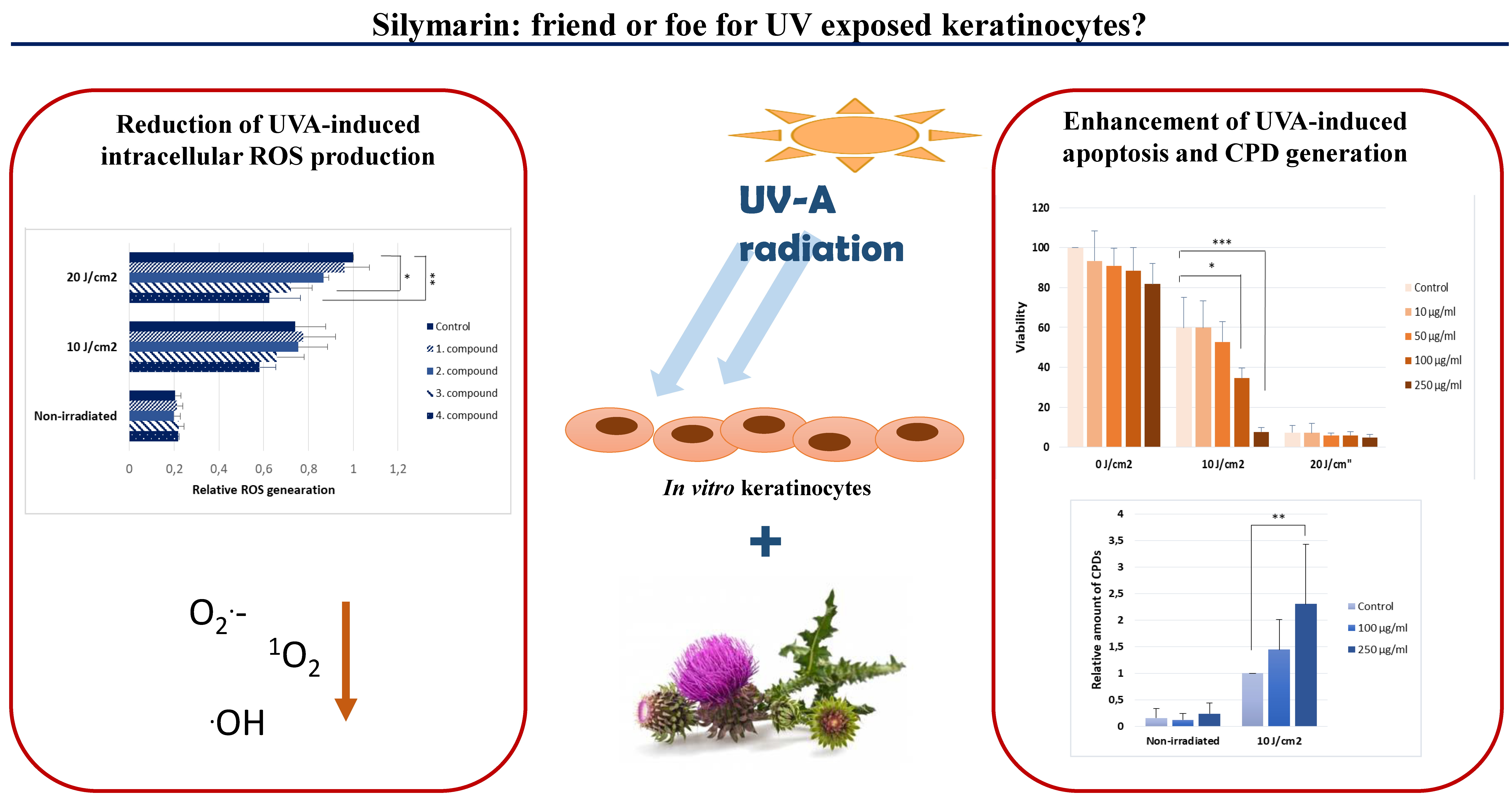
2. Results
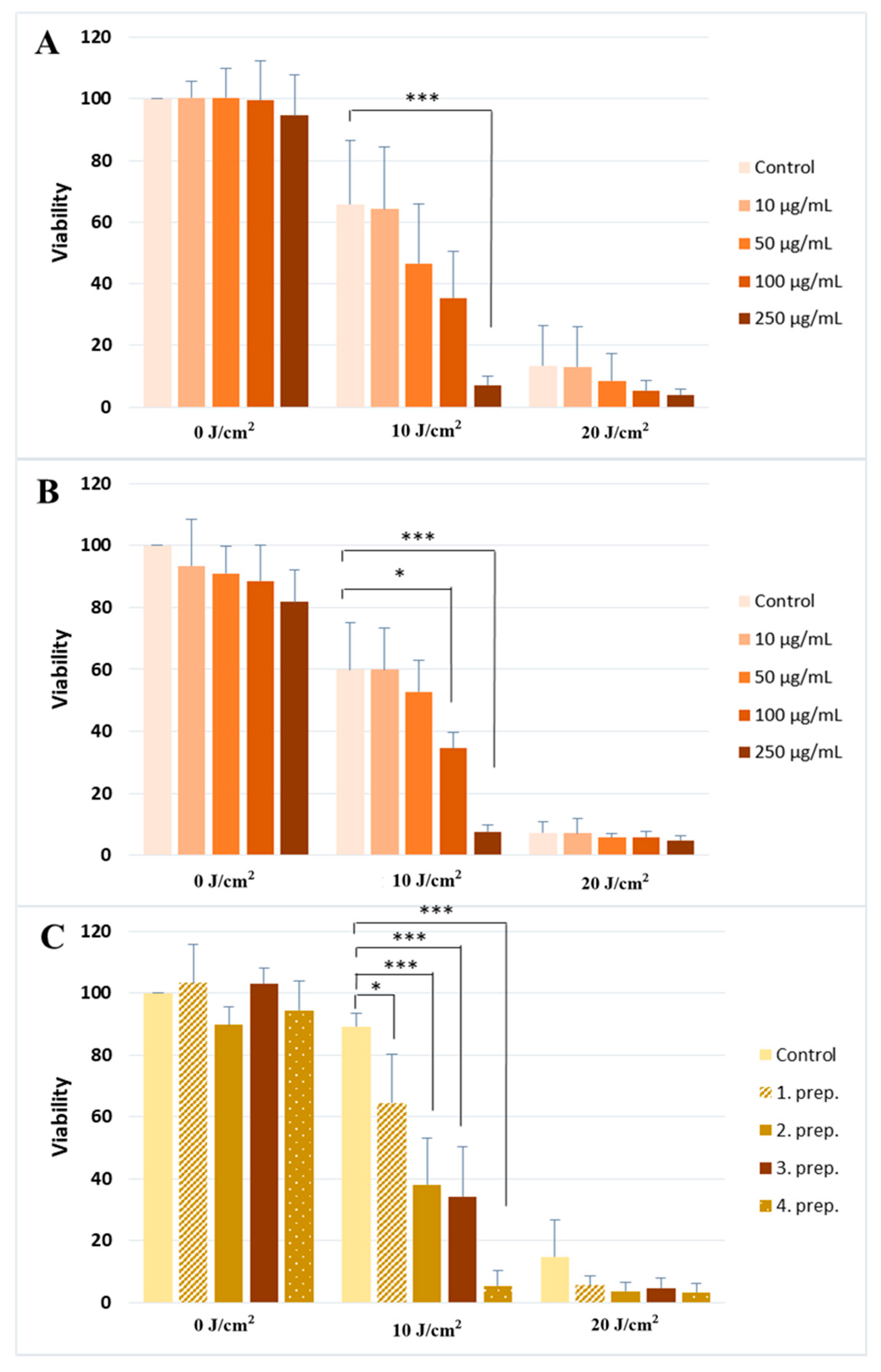
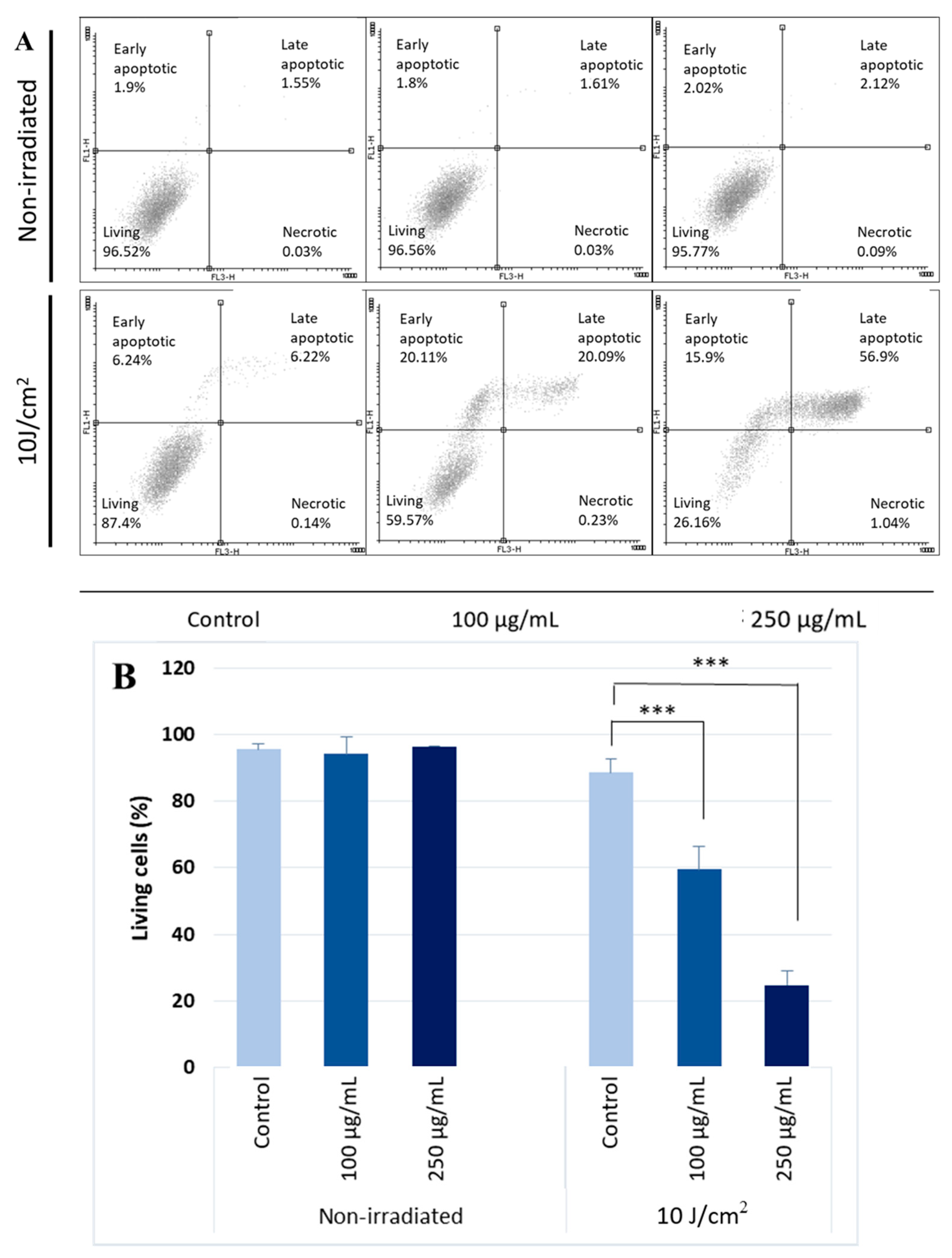
2.1. Silymarin Pre-Treatment Enhances the UVA-Induced Cytotoxicity
- (1)
- A silymarin extract (Sigma-Aldrich, St. Louis, MO, USA) dissolved in ethanol.
- (2)
- A commercially available silymarin compound (Silegon, Teva Pharmaceutical Industries Ltd., Petach Tikva, Israel).
- (3)
- Four different topical formulations of silymarin containing 250 µg/mL herbal extract dissolved in Transcutol HP (TC) and different sucrose-esters as penetration enhancers (see Table 1 at Materials for details). We have previously shown the efficacy of these enhancers on cell and skin penetration [36].
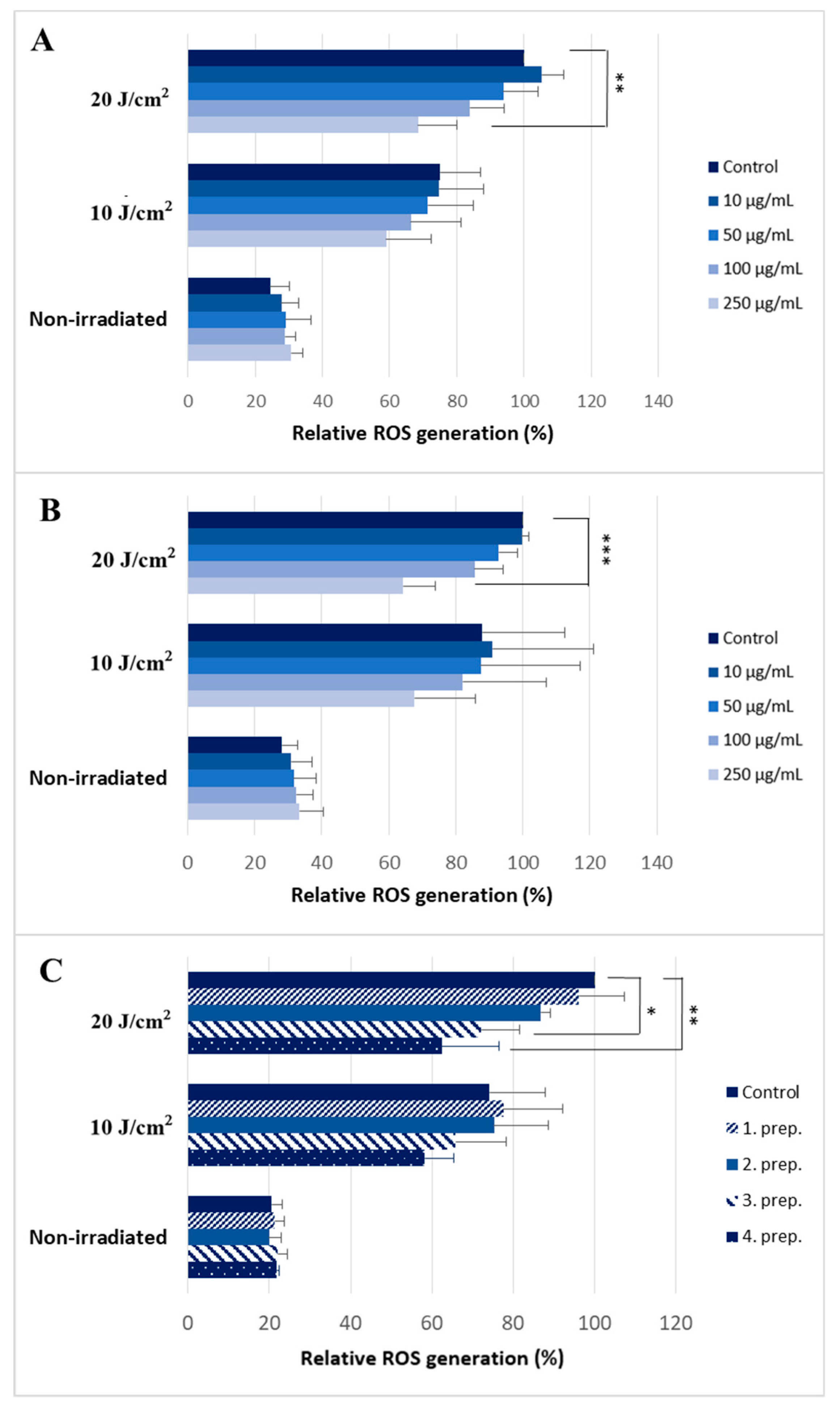
2.2. Silymarin Treatment Reduces Intracellular ROS Production after UVA Irradiation
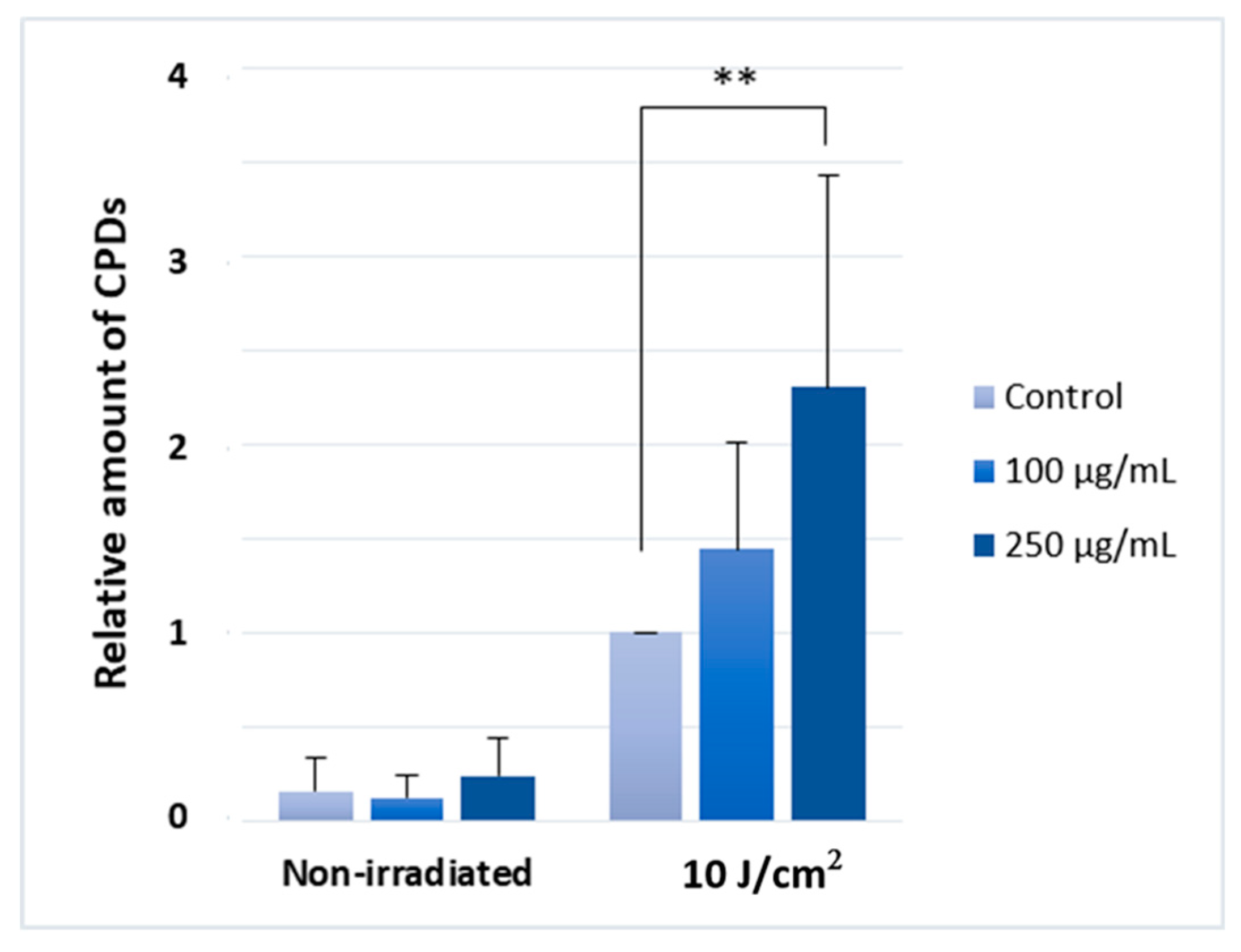
2.3. Silymarin Pre-Treatment Enhances the UVA-Induced CPD-Generation
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Preparation of Silymarin Formulations
4.2.1. Materials
4.2.2. Preparation of Compositions Containing Silymarin Powder
4.3. Silymarin Treatments
4.4. UVA Irradiation
4.5. MTT (3-[4–dimethylthiazol-2-yl]-2,5 diphenyl tetrazolium bromide) Assay
4.6. Annexin V/Propidium Iodide Staining (Apoptosis Assay)
4.7. ROS (Reactive Oxygen Species) Production Measurements
4.8. Enzyme-Linked Immunosorbent Assay (ELISA)
4.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Brem, R.; Guven, M.; Karran, P. Oxidatively-Generated damage to DNA and proteins mediated by photosensitized UVA. Free Radic. Biol. Med. 2017, 107, 101–109. [Google Scholar] [CrossRef]
- de Gruijl, F.R. Photocarcinogenesis: UVA vs. UVB. Methods Enzymol. 2000, 319, 359–366. [Google Scholar] [PubMed]
- Ridley, A.J.; Whiteside, J.R.; McMillan, T.J.; Allinson, S.L. Cellular and sub-cellular responses to UVA in relation to carcinogenesis. Int. J. Radiat. Biol. 2009, 85, 177–195. [Google Scholar] [CrossRef] [PubMed]
- Wondrak, G.T.; Jacobson, M.K.; Jacobson, E.L. Endogenous UVA-photosensitizers: Mediators of skin photodamage and novel targets for skin photoprotection. Photochem. Photobiol. Sci. 2006, 5, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Damiani, E.; Rosati, L.; Castagna, R.; Carloni, P.; Greci, L. Changes in ultraviolet absorbance and hence in protective efficacy against lipid peroxidation of organic sunscreens after UVA irradiation. J. Photochem. Photobiol. B Biol. 2006, 82, 204–213. [Google Scholar] [CrossRef]
- Sander, C.S.; Chang, H.; Salzmann, S.; Müller, C.S.; Ekanayake-Mudiyanselage, S.; Elsner, P.; Thiele, J.J. Photoaging is associated with protein oxidation in human skin in vivo. J. Investig. Dermatol. 2002, 118, 618–625. [Google Scholar] [CrossRef]
- Kozmin, S.; Slezak, G.; Reynaud-Angelin, A.; Elie, C.; de Rycke, Y.; Boiteux, S.; Sage, E. UVA radiation is highly mutagenic in cells that are unable to repair 7,8-dihydro-8-oxoguanine in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2005, 102, 13538–13543. [Google Scholar] [CrossRef] [PubMed]
- Javeri, A.; Lyons, J.G.; Huang, X.X.; Halliday, G.M. Downregulation of Cockayne syndrome B protein reduces human 8-oxoguanine DNA glycosylase-1 expression and repair of UV radiation-induced 8-oxo-7,8-dihydro-2’-deoxyguanine. Cancer Sci. 2011, 102, 1651–1658. [Google Scholar] [CrossRef]
- Besaratinia, A.; Synold, T.W.; Chen, H.H.; Chang, C.; Xi, B.; Riggs, A.D.; Pfeifer, G.P. DNA lesions induced by UV A1 and B radiation in human cells: Comparative analyses in the overall genome and in the p53 tumor suppressor gene. Proc. Natl. Acad. Sci. USA 2005, 102, 10058–10063. [Google Scholar] [CrossRef]
- Sage, E.; Girard, P.M.; Francesconi, S. Unravelling UVA-induced mutagenesis. Photochem. Photobiol. Sci. 2012, 11, 74–80. [Google Scholar] [CrossRef]
- Douki, T.; Reynaud-Angelin, A.; Cadet, J.; Sage, E. Bipyrimidine photoproducts rather than oxidative lesions are the main type of DNA damage involved in the genotoxic effect of solar UVA radiation. Biochemistry 2003, 42, 9221–9226. [Google Scholar] [CrossRef]
- Courdavault, S.; Baudouin, C.; Charveron, M.; Favier, A.; Cadet, J.; Douki, T. Larger yield of cyclobutane dimers than 8-oxo-7,8-dihydroguanine in the DNA of UVA-irradiated human skin cells. Mutat. Res. 2004, 556, 135–142. [Google Scholar] [CrossRef]
- Courdavault, S.; Baudouin, C.; Charveron, M.; Canguilhem, B.; Favier, A.; Cadet, J.; Douki, T. Repair of the three main types of bipyrimidine DNA photoproducts in human keratinocytes exposed to UVB and UVA radiations. DNA Repair 2005, 4, 836–844. [Google Scholar] [CrossRef] [PubMed]
- Lo, H.L.; Nakajima, S.; Ma, L.; Walter, B.; Yasui, A.; Ethell, D.W.; Owen, L.B. Differential biologic effects of CPD and 6-4PP UV-induced DNA damage on the induction of apoptosis and cell-cycle arrest. BMC Cancer 2005, 5, 135. [Google Scholar] [CrossRef]
- D’Errico, M.; Teson, M.; Calcagnile, A.; Proietti De Santis, L.; Nikaido, O.; Botta, E.; Zambruno, G.; Stefanini, M.; Dogliotti, E. Apoptosis and efficient repair of DNA damage protect human keratinocytes against UVB. Cell Death Differ. 2003, 10, 754–756. [Google Scholar] [CrossRef] [PubMed]
- Drouin, R.; Therrien, J.P. UVB-induced cyclobutane pyrimidine dimer frequency correlates with skin cancer mutational hotspots in p53. Photochem. Photobiol. 1997, 66, 719–726. [Google Scholar] [CrossRef] [PubMed]
- You, Y.H.; Lee, D.H.; Yoon, J.H.; Nakajima, S.; Yasui, A.; Pfeifer, G.P. Cyclobutane pyrimidine dimers are responsible for the vast majority of mutations induced by UVB irradiation in mammalian cells. J. Biol. Chem. 2001, 276, 44688–44694. [Google Scholar] [CrossRef]
- Szabados-Furjesi, P.; Pajtas, D.; Barta, A.; Csepanyi, E.; Kiss-Szikszai, A.; Tosaki, A.; Bak, I. Synthesis, in Vitro Biological Evaluation, and Oxidative Transformation of New Flavonol Derivatives: The Possible Role of the Phenyl-N,N-Dimethylamino Group. Molecules 2018, 23, 3161. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Liu, Z.; Yang, H.; Li, C.; Chen, H.; Liu, Y.; Zhao, M.; Zhu, Y. Luteolin decreases the UVA-induced autophagy of human skin fibroblasts by scavenging ROS. Mol. Med. Rep. 2016, 14, 1986–1992. [Google Scholar] [CrossRef] [PubMed]
- Mejía-Giraldo, J.C.; Winkler, R.; Gallardo, C.; Sánchez-Zapata, A.M.; Puertas-Mejía, M.A. Photoprotective Potential of Baccharis antioquensis (Asteraceae) as Natural Sunscreen. Photochem. Photobiol. 2016, 92, 742–752. [Google Scholar] [CrossRef]
- Sim, G.S.; Lee, B.C.; Cho, H.S.; Lee, J.W.; Kim, J.H.; Lee, D.H.; Kim, J.H.; Pyo, H.B.; Moon, D.C.; Oh, K.W.; et al. Structure activity relationship of antioxidative property of flavonoids and inhibitory effect on matrix metalloproteinase activity in UVA-irradiated human dermal fibroblast. Arch. Pharm. Res. 2007, 30, 290–298. [Google Scholar] [CrossRef]
- Tobi, S.E.; Gilbert, M.; Paul, N.; McMillan, T.J. The green tea polyphenol, epigallocatechin-3-gallate, protects against the oxidative cellular and genotoxic damage of UVA radiation. Int. J. Cancer 2002, 102, 439–444. [Google Scholar] [CrossRef]
- Saliou, C.; Kitazawa, M.; McLaughlin, L.; Yang, J.P.; Lodge, J.K.; Tetsuka, T.; Iwasaki, K.; Cillard, J.; Okamoto, T.; Packer, L. Antioxidants modulate acute solar ultraviolet radiation-induced NF-kappa-B activation in a human keratinocyte cell line. Free Radic. Biol. Med. 1999, 26, 174–183. [Google Scholar] [CrossRef]
- Deep, G.; Agarwal, R. Chemopreventive efficacy of silymarin in skin and prostate cancer. Integr. Cancer Ther. 2007, 6, 130–145. [Google Scholar] [CrossRef]
- Gupta, O.P.; Sing, S.; Bani, S.; Sharma, N.; Malhotra, S.; Gupta, B.D.; Banerjee, S.K.; Handa, S.S. Anti-inflammatory and anti-arthritic activities of silymarin acting through inhibition of 5-lipoxygenase. Phytomedicine 2000, 7, 21–24. [Google Scholar] [CrossRef]
- Vaid, M.; Prasad, R.; Singh, T.; Elmets, C.A.; Xu, H.; Katiyar, S.K. Silymarin inhibits ultraviolet radiation-induced immune suppression through DNA repair-dependent activation of dendritic cells and stimulation of effector T cells. Biochem. Pharmacol. 2013, 85, 1066–1076. [Google Scholar] [CrossRef]
- Svobodová, A.; Zdarilová, A.; Walterová, D.; Vostálová, J. Flavonolignans from Silybum marianum moderate UVA-induced oxidative damage to HaCaT keratinocytes. J. Dermatol. Sci. 2007, 48, 213–224. [Google Scholar] [CrossRef]
- Katiyar, S.K.; Korman, N.J.; Mukhtar, H.; Agarwal, R. Protective effects of silymarin against photocarcinogenesis in a mouse skin model. J. Natl. Cancer Inst. 1997, 89, 556–566. [Google Scholar] [CrossRef]
- Vostálová, J.; Tinková, E.; Biedermann, D.; Kosina, P.; Ulrichová, J.; Rajnochová Svobodová, A. Skin Protective Activity of Silymarin and its Flavonolignans. Molecules 2019, 24, 1022. [Google Scholar] [CrossRef]
- Marchiori, M.C.L.; Rigon, C.; Camponogara, C.; Oliveira, S.M.; Cruz, L. Hydrogel containing silibinin-loaded pomegranate oil based nanocapsules exhibits anti-inflammatory effects on skin damage UVB radiation-induced in mice. J. Photochem. Photobiol. B 2017, 170, 25–32. [Google Scholar] [CrossRef]
- Rigby, C.M.; Roy, S.; Deep, G.; Guillermo-Lagae, R.; Jain, A.K.; Dhar, D.; Orlicky, D.J.; Agarwal, C.; Agarwal, R. Role of p53 in silibinin-mediated inhibition of ultraviolet B radiation-induced DNA damage, inflammation and skin carcinogenesis. Carcinogenesis 2017, 38, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Narayanapillai, S.; Agarwal, C.; Tilley, C.; Agarwal, R. Silibinin is a potent sensitizer of UVA radiation-induced oxidative stress and apoptosis in human keratinocyte HaCaT cells. Photochem. Photobiol. 2012, 88, 1135–1140. [Google Scholar] [CrossRef] [PubMed]
- Rajnochová Svobodová, A.; Zálešák, B.; Biedermann, D.; Ulrichová, J.; Vostálová, J. Phototoxic potential of silymarin and its bioactive components. J. Photochem. Photobiol. B 2016, 156, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Katiyar, S.K.; Roy, A.M.; Baliga, M.S. Silymarin induces apoptosis primarily through a p53-dependent pathway involving Bcl-2/Bax, cytochrome c release, and caspase activation. Mol. Cancer Ther. 2005, 4, 207–216. [Google Scholar] [PubMed]
- Baptista, M.S.; Cadet, J.; Di Mascio, P.; Ghogare, A.A.; Greer, A.; Hamblin, M.R.; Lorente, C.; Nunez, S.C.; Ribeiro, M.S.; Thomas, A.H.; et al. Type I and Type II Photosensitized Oxidation Reactions: Guidelines and Mechanistic Pathways. Photochem. Photobiol. 2017, 93, 912–919. [Google Scholar] [CrossRef] [PubMed]
- Fehér, P.; Ujhelyi, Z.; Váradi, J.; Fenyvesi, F.; Róka, E.; Juhász, B.; Varga, B.; Bombicz, M.; Priksz, D.; Bácskay, I.; et al. Efficacy of Pre- and Post-Treatment by Topical Formulations Containing Dissolved and Suspended Silybum marianum against UVB-Induced Oxidative Stress in Guinea Pig and on HaCaT Keratinocytes. Molecules 2016, 21, 1269. [Google Scholar] [CrossRef]
- Kunisada, M.; Masaki, T.; Ono, R.; Morinaga, H.; Nakano, E.; Yogianti, F.; Okunishi, K.; Sugiyama, H.; Nishigori, C. Hydrochlorothiazide enhances UVA-induced DNA damage. Photochem. Photobiol. 2013, 89, 649–654. [Google Scholar] [CrossRef]
- Robinson, K.S.; Traynor, N.J.; Moseley, H.; Ferguson, J.; Woods, J.A. Cyclobutane pyrimidine dimers are photosensitised by carprofen plus UVA in human HaCaT cells. Toxicol. In Vitro 2010, 24, 1126–1132. [Google Scholar] [CrossRef]
- Kvasnicka, F.; Bíba, B.; Sevcík, R.; Voldrich, M.; Krátká, J. Analysis of the active components of silymarin. J. Chromatogr. A 2003, 990, 239–245. [Google Scholar] [CrossRef]
- Dhanalakshmi, S.; Mallikarjuna, G.U.; Singh, R.P.; Agarwal, R. Dual efficacy of silibinin in protecting or enhancing ultraviolet B radiation-caused apoptosis in HaCaT human immortalized keratinocytes. Carcinogenesis 2004, 25, 99–106. [Google Scholar] [CrossRef]
- Mudambi, S.; Pera, P.; Washington, D.; Remenyik, E.; Fidrus, E.; Shafirstein, G.; Bellnier, D.; Paragh, G. Photodynamic therapy does not induce cyclobutane pyrimidine dimers in the presence of melanin. Photodiagnosis Photodyn. Ther. 2018, 22, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Boukamp, P.; Petrussevska, R.T.; Breitkreutz, D.; Hornung, J.; Markham, A.; Fusenig, N.E. Normal keratinization in a spontaneously immortalized aneuploid human keratinocyte cell line. J. Cell Biol. 1988, 106, 761–771. [Google Scholar] [CrossRef]
- Kahol, A.P.; Singh, K.L.; Tandon, S.; Kumar, S. Process for Isolation of Hepatoprotective Agent Silymarin from the Seeds of the Plant Silybum marianum. U.S. Patent US6309678B1, 30 October 2001. [Google Scholar]
- Kuki, Á.; Nagy, L.; Deák, G.; Nagy, M.; Zsuga, M.; Kéki, S. Identification of Silymarin Constituents: An Improved HPLC-MS Method. Chromatographia 2012, 75, 175–180. [Google Scholar] [CrossRef]
- Boros, G.; Miko, E.; Muramatsu, H.; Weissman, D.; Emri, E.; Rózsa, D.; Nagy, G.; Juhász, A.; Juhász, I.; van der Horst, G.; et al. Transfection of pseudouridine-modified mRNA encoding CPD-photolyase leads to repair of DNA damage in human keratinocytes: A new approach with future therapeutic potential. J. Photochem. Photobiol. B 2013, 129, 93–99. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the silymarin compounds are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Preparations | ||||
|---|---|---|---|---|
| Ingredients (g) | 1 | 2 | 3 | 4 |
| Silymarin powder | 0.25 | 0.25 | 0.25 | 0.25 |
| Transcutol | ---- | ----- | 0.71 | 0.71 |
| Sucrose ester SP50 | 0.15 | ---- | 0.15 | --- |
| Sucrose ester SP70 | --- | 0.15 | --- | 0.15 |
| Propylene Glycol | 0.25 | 0.25 | 0.25 | 0.25 |
| Cell culture media | ad 100 | ad 100 | ad 100 | ad 100 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fidrus, E.; Ujhelyi, Z.; Fehér, P.; Hegedűs, C.; Janka, E.A.; Paragh, G.; Vasas, G.; Bácskay, I.; Remenyik, É. Silymarin: Friend or Foe of UV Exposed Keratinocytes? Molecules 2019, 24, 1652. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24091652
Fidrus E, Ujhelyi Z, Fehér P, Hegedűs C, Janka EA, Paragh G, Vasas G, Bácskay I, Remenyik É. Silymarin: Friend or Foe of UV Exposed Keratinocytes? Molecules. 2019; 24(9):1652. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24091652
Chicago/Turabian StyleFidrus, Eszter, Zoltán Ujhelyi, Pálma Fehér, Csaba Hegedűs, Eszter Anna Janka, György Paragh, Gábos Vasas, Ildikó Bácskay, and Éva Remenyik. 2019. "Silymarin: Friend or Foe of UV Exposed Keratinocytes?" Molecules 24, no. 9: 1652. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24091652