
An Improved Method for the Synthesis of Butein Using SOCl2/EtOH as Catalyst and Deciphering Its Inhibition Mechanism on Xanthine Oxidase


Abstract
:1. Introduction
2. Results and Discussion
2.1. Synthesis of Butein
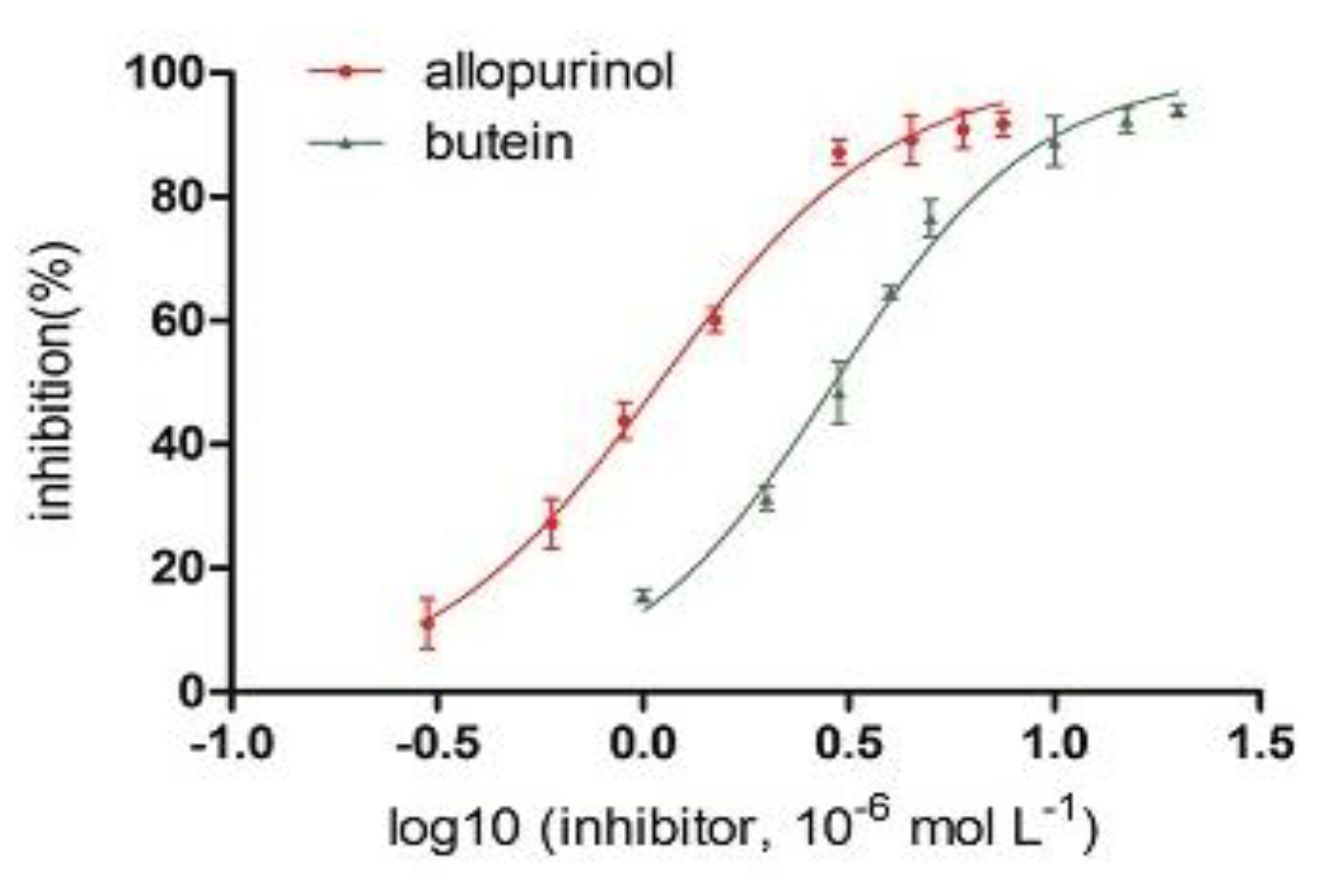
2.2. Inhibition of Butein on XO Activity
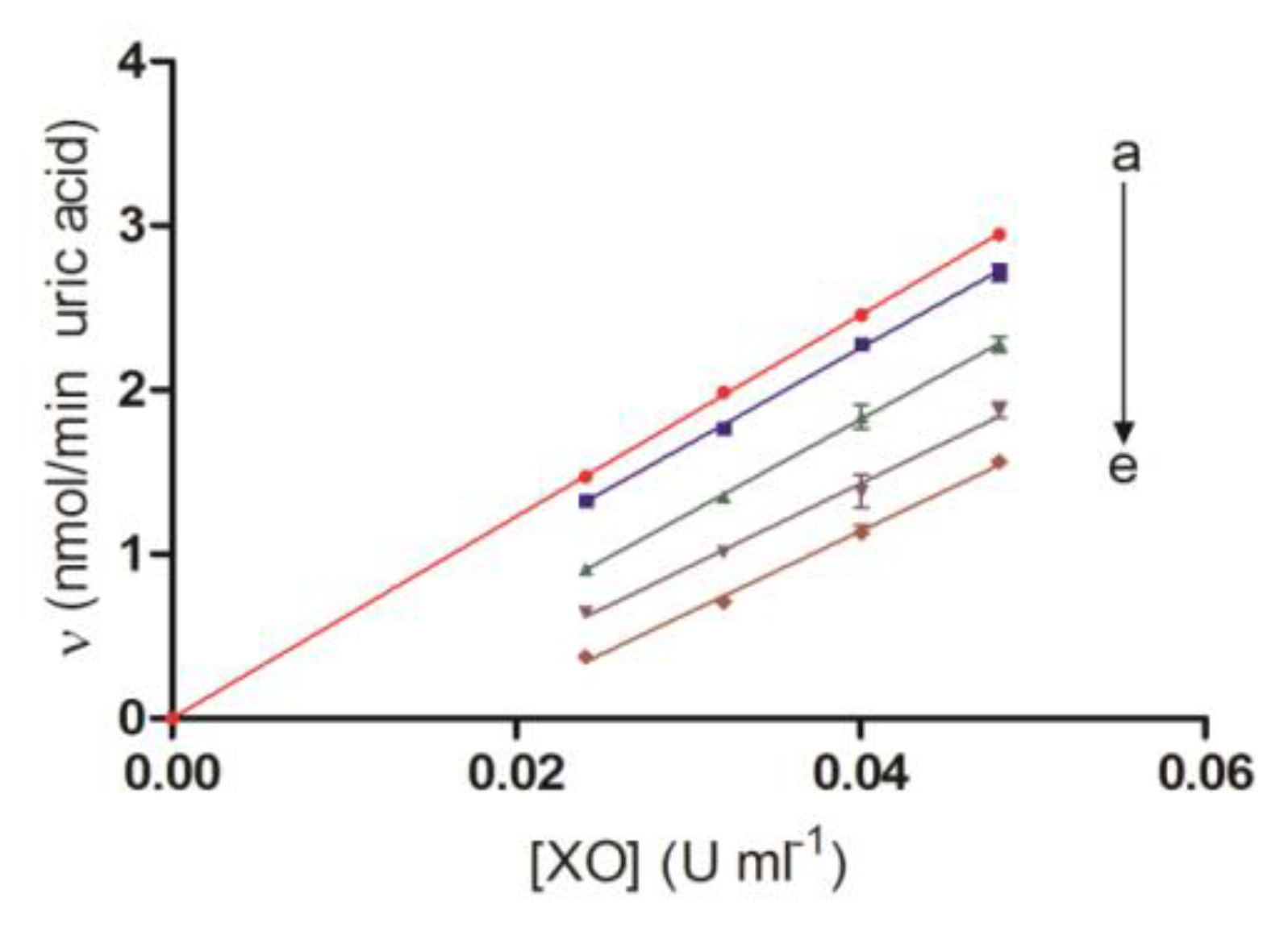
2.3. Irreversibility
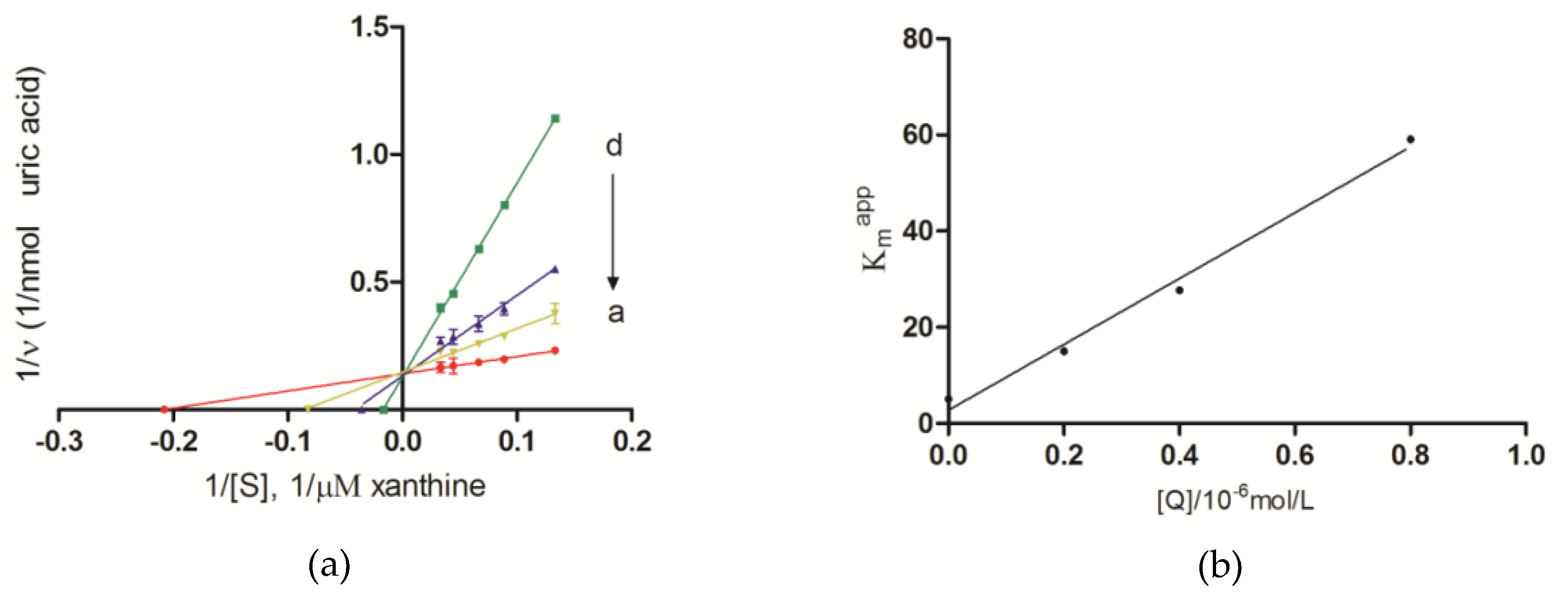
2.4. Inhibition Kinetic Analysis
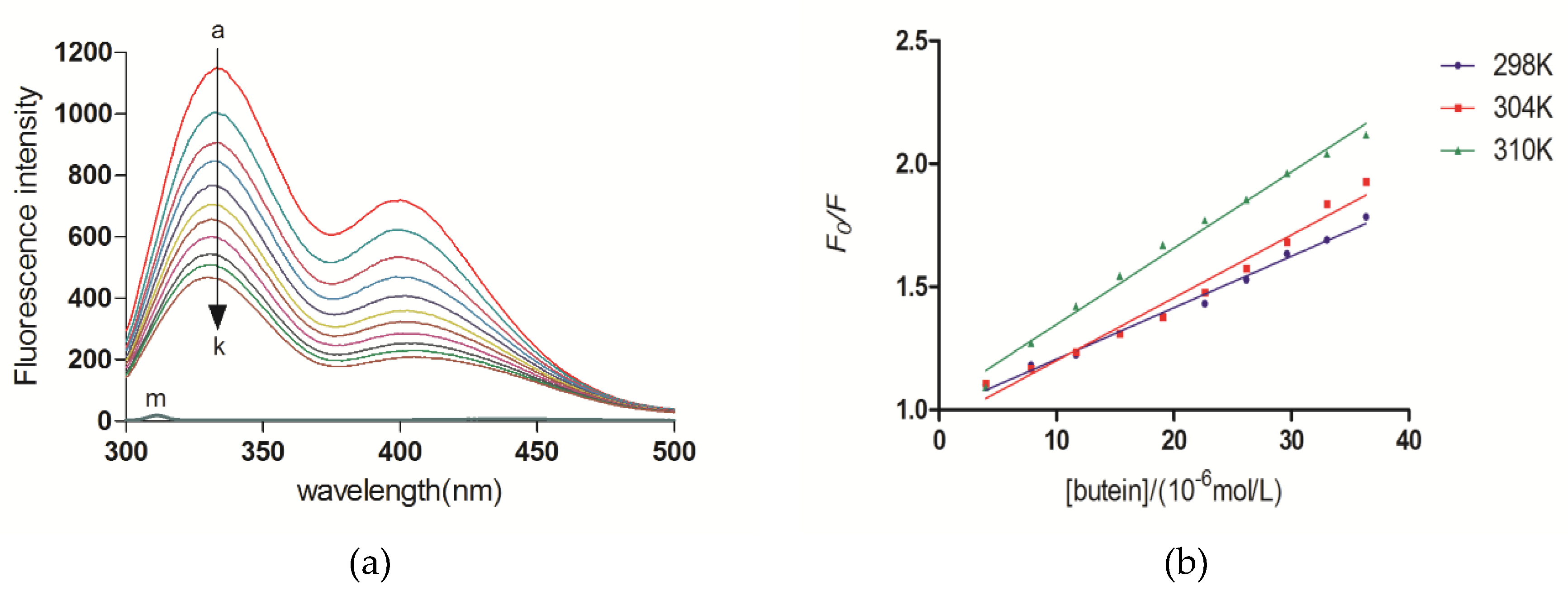
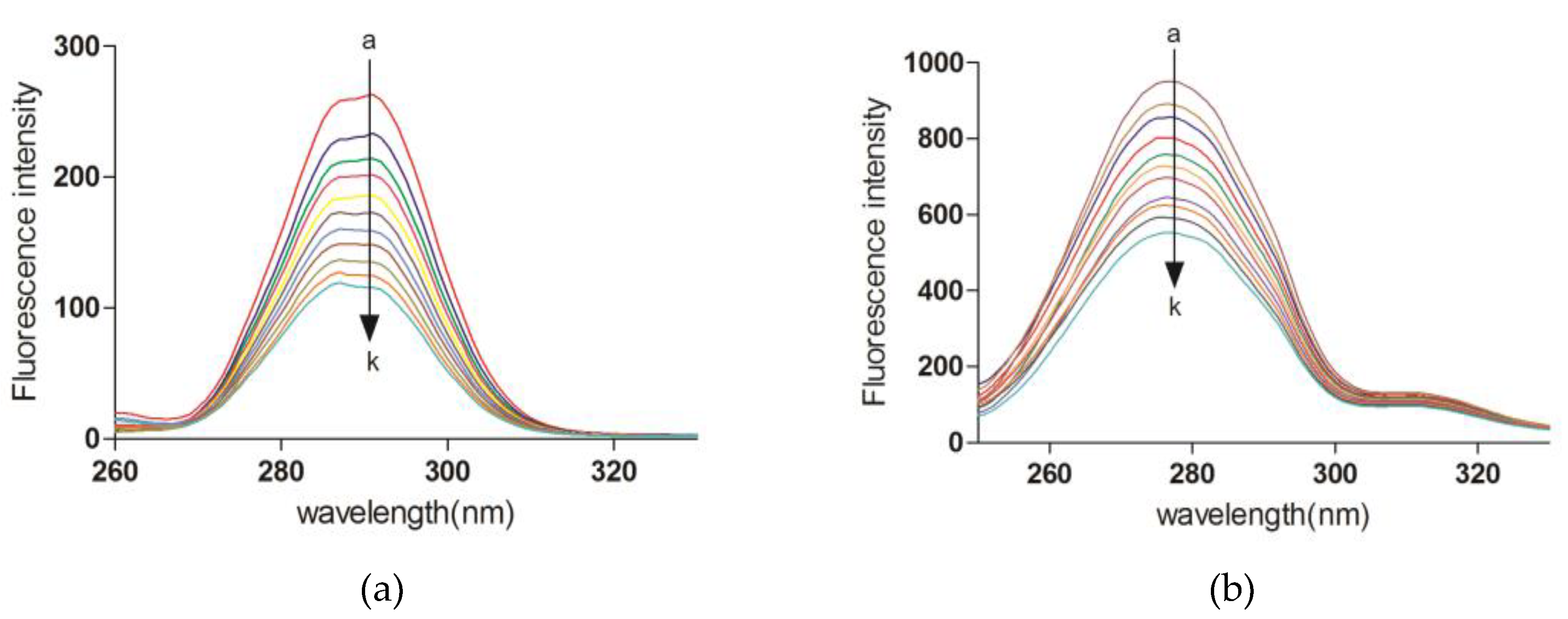
2.5. Fluorescence Quenching of XO upon Butein
2.6. Thermodynamic Analysis
2.7. Synchronous Fluorescence
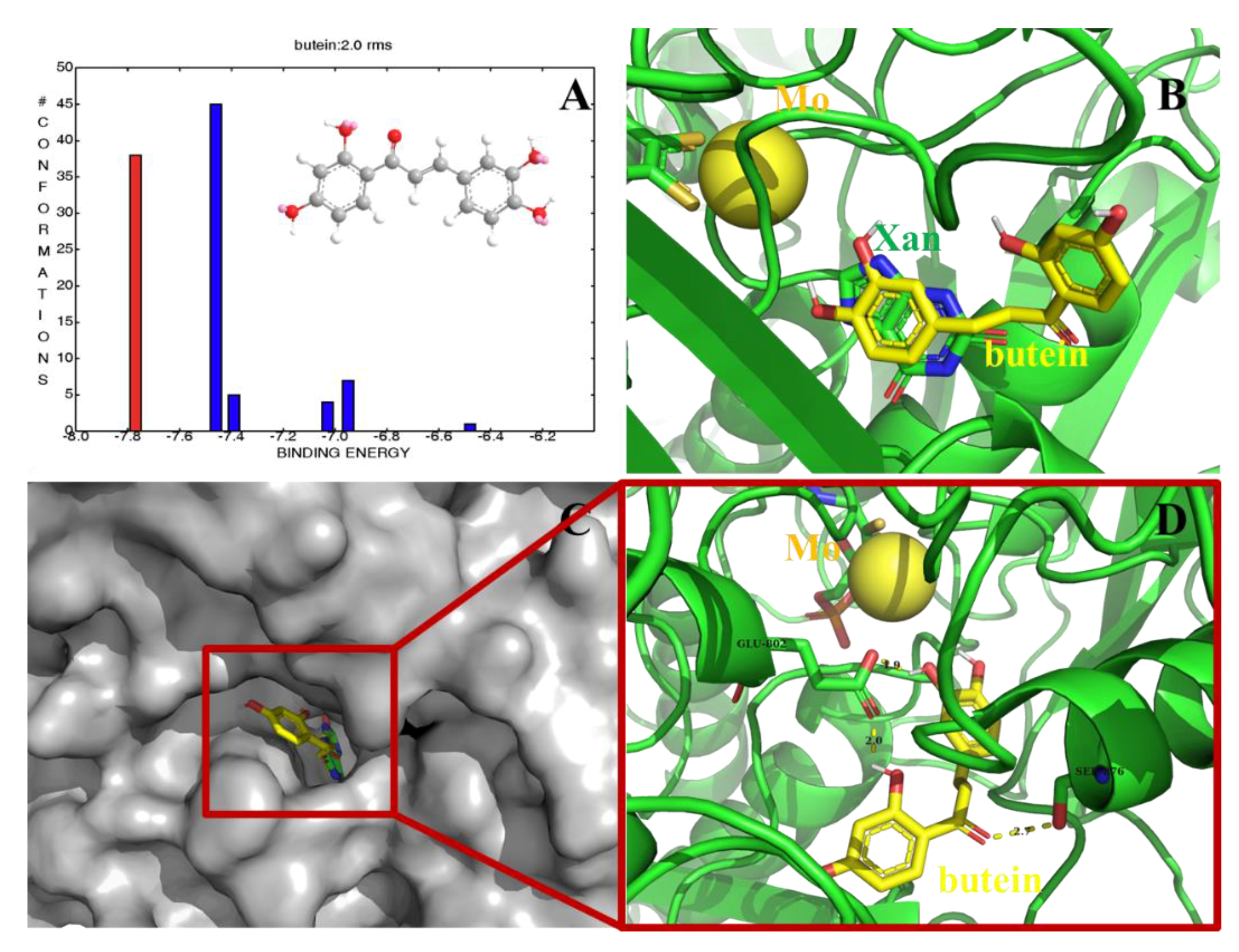
2.8. Computational Docking of the Butein–XO Complex
3. Experimental Section
3.1. Materials
3.2. General Procedure for the Synthesis of Butein
3.3. Enzyme Activity Assay
3.4. Kinetic Analysis for Inhibitory
3.5. Fluorescence Titration
3.6. Molecular Docking
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Maruhashi, T.; Hisatome, I.; Kihara, Y.; Higashi, Y. Hyperuricemia and endothelial function: From molecular background to clinical perspectives. Atherosclerosis 2018, 278, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Ragab, G.; Elshahaly, M.; Bardin, T. Gout: An old disease in new perspective—A review. J. Adv. Res. 2017, 8, 495–511. [Google Scholar] [CrossRef]
- Punzi, L.; Scanu, A.; Ramonda, R.; Oliviero, F. Gout as autoinflammatory disease: New mechanisms for more appropriated treatment targets. Autoimmun. Rev. 2012, 12, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Wang, H.; Xia, W.; Chang, X.; Wang, M.; An, L. Prevalence and correlates of hyperuricemia in the middle-aged and older adults in China. Sci. Rep. 2018, 8, 4314–4322. [Google Scholar] [CrossRef] [PubMed]
- Punzi, L.; So, A. Serum uric acid and gout: From the past to molecular biology. Curr. Med. Res. Opin. 2013, 29, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Zhu, H.; Liu, J.; Fan, H.; Sun, L.; Lu, W.; Liu, X.; Li, L. 3,5,2′,4′-tetrahydroxychalcone, a new non-purine xanthine oxidase inhibitor. Chem. Biol. Interact. 2011, 189, 161–166. [Google Scholar] [CrossRef]
- Enroth, C.; Eger, B.T.; Okamoto, K.; Nishino, T.; Pai, E.F. Crystal structures of bovine milk xanthine dehydrogenase and xanthine oxidase: Structure-based mechanism of conversion. Proc. Natl. Acad. Sci. USA. 2000, 97, 10723–10728. [Google Scholar] [CrossRef] [Green Version]
- Terkeltaub, R.; Bushinsky, D.A.; Becker, M.A. Recent developments in our understanding of the renal basis of hyperuricemia and the development of novel antihyperuricemic therapeutics. Arthritis Res. Ther. 2006, 8, S4. [Google Scholar] [CrossRef] [PubMed]
- Burns, C.M.; Wortmann, R.L. Gout therapeutics: New drugs for an old disease. Lancet 2011, 377, 165–177. [Google Scholar] [CrossRef]
- Diaz-Torné, C.; Perez-Herrero, N.; Perez-Ruiz, F. New medications in development for the treatment of hyperuricemia of gout. Curr. Opin. Rheumatol. 2015, 27, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.S.; Dai, Y.S.; Pwu, R.F.; Chen, Y.H.; Chang, N.C. Risk estimates for drugs suspected of being associated with Stevens-Johnson syndrome and toxic epidermal necrolysis: A case-control study. Int. Med. J. 2005, 35, 188–190. [Google Scholar] [CrossRef] [PubMed]
- Reinders, M.K.; Jansen, T.L. Management of hyperuricemia in gout: Focus on febuxostat. Clin. Interv. Aging 2010, 5, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Jordan, A.; Gresser, U. Side effects and interactions of the xanthine oxidase inhibitor febuxostat. Pharmaceuticals 2018, 11, 51. [Google Scholar] [CrossRef]
- Lin, C.W.; Huang, W.I.; Chao, P.H.; Chen, W.W.; Hsiao, F.Y. Risk of cutaneous adverse reactions associated with allopurinol or febuxostat in real-world patients: A nationwide study. Int. J. Clin. Pract. 2019, e13316. [Google Scholar] [CrossRef]
- Cheng, Z.J.; Kuo, S.C.; Chan, S.C.; Ko, F.N.; Teng, C.M. Antioxidant properties of butein isolated from Dalbergia odorifera. Biochim. Biophys. Acta 1998, 1392, 291–299. [Google Scholar] [CrossRef]
- Hofmann, E.; Webster, J.; Do, T.; Kline, R.; Snider, L.; Hauser, Q.; Higginbottom, G.; Campbell, A.; Ma, L.; Paula, S. Hydroxylated chalcones with dual properties: Xanthine oxidase inhibitors and radical scavengers. Bioorg. Med. Chem. 2016, 24, 578–587. [Google Scholar] [CrossRef] [PubMed]
- Padmavathi, G.; Roy, N.K.; Bordoloi, D.; Arfuso, F.; Mishra, S.; Sethi, G.; Bishayee, A.; Kunnumakkara, A.B. Butein in health and disease: A comprehensive review. Phytomedicine 2017, 25, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Yang, Z.; Li, C.; Zhu, Z.; Shen, X.; Hu, L. Design, synthesis and SAR study of hydroxychalcone inhibitors of human β-secretase (BACE1). J. Enzyme Inhib. Med. Chem. 2011, 26, 643–648. [Google Scholar] [CrossRef] [Green Version]
- Sultan, A.; Raza, A.R.; Abbas, M.; Khan, K.M.; Tahir, M.N.; Saari, N. Evaluation of silica-H2SO4 as an efficient heterogeneous catalyst for the synthesis of chalcones. Molecules 2013, 18, 10081–10094. [Google Scholar] [CrossRef]
- Hu, Z.; Liu, J.; Dong, Z.; Guo, L.; Wang, D.; Zeng, P. Synthesis of chalcones catalyzed by SOCl2/EtOH. J. Chem. Res. 2004, 2, 158–159. [Google Scholar] [CrossRef]
- Petrov, O.; Ivanova, Y.; Gerova, M. SOCl2/EtOH: Catalytic system for synthesis of chalcones. Catal. Commun. 2008, 9, 315–316. [Google Scholar] [CrossRef]
- Bui, T.H.; Nguyen, N.T.; Dang, P.H. Design and synthesis of chalcone derivatives as potential non-purine xanthine oxidase inhibitors. SpringerPlus 2016, 5, 1789–1797. [Google Scholar] [CrossRef]
- Ponce, A.; Blanco, S.; Molina, A.; García-Domenech, R.; Gálvez, J. Study of the action of flavonoids on xanthine-oxidase by molecular topology. J. Chem. Inf. Comput. Sci. 2000, 40, 1039–1045. [Google Scholar] [CrossRef]
- Costantino, L.; Rastelli, G.; Albasini, A. A rational approach to the design of flavones as xanthine oxidase inhibitors. Eur. J. Med. Chem. 1996, 31, 693–699. [Google Scholar] [CrossRef]
- Mathew, B.; Suresh, J.; Mathew, G.; Rasheed, S.; Vilapurathu, J.; Jayaraj, P. Flavonoids: An outstanding structural core for the inhibition of xanthine oxidase enzyme. Curr. Enzym. Inhib. 2015, 11, 108–115. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, G.; Pan, J.; Gong, D. Novel insights into the inhibitory mechanism of kaempferol on xanthine oxidase. J. Agric. Food. Chem. 2015, 63, 526–534. [Google Scholar] [CrossRef]
- Lin, S.; Zhang, G.; Liao, Y.; Pan, J. Inhibition of chrysin on xanthine oxidase activity and its inhibition mechanism. Int. J. Biol. Macromol. 2015, 81, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Zou, B.; Zeng, H.; Zhang, L.; Chen, M.; Fu, G. Inhibitory effect of verbascoside on xanthine oxidase activity. Int. J. Biol. Macromol. 2016, 93, 609–614. [Google Scholar] [CrossRef]
- Huo, L.N.; Wang, W.; Zhang, C.Y.; Shi, H.B.; Liu, Y.; Liu, X.H.; Guo, B.H.; Zhao, D.M.; Gao, H. Bioassay-guided isolation and identification of xanthine oxidase inhibitory constituents from the leaves of, Perilla frutescens. Molecules 2015, 20, 17848–17859. [Google Scholar] [CrossRef]
- Yan, J.; Zhang, G.; Hu, Y.; Ma, Y. Effect of luteolin on xanthine oxidase: Inhibition kinetics and interaction mechanism merging with docking simulation. Food Chem. 2013, 141, 3766–3773. [Google Scholar] [CrossRef]
- Zhang, G.; Wang, L.; Pan, J. Probing the binding of the flavonoid diosmetin to human serum albumin by multispectroscopic techniques. J. Agric. Food Chem. 2012, 60, 2721–2729. [Google Scholar] [CrossRef]
- Peng, W.; Ding, F.; Jiang, Y.T.; Sun, Y.; Peng, Y.K. Evaluation of the biointeraction of colorant flavazin with human serum albumin: Insights from multiple spectroscopic studies, in silico docking and molecular dynamics simulation. Food Funct. 2014, 5, 1203–1217. [Google Scholar] [CrossRef]
- Zhang, G.; Ma, Y.; Wang, L.; Zhang, Y.; Zhou, J. Multispectroscopic studies on the interaction of maltol, a food additive, with bovine serum albumin. Food Chem. 2012, 133, 264–270. [Google Scholar] [CrossRef]
- Bi, S.; Ding, L.; Tian, Y.; Song, D.; Zhou, X.; Liu, X.; Zhang, H. Investigation of the interaction between flavonoids and human serum albumin. J. Mol. Struct. 2004, 703, 37–45. [Google Scholar] [CrossRef]
- Shuai, L.; Chen, Z.; Fei, P.; Wang, Q.; Yang, T. Spectroscopic analysis on the interaction of ferulic acid and tetramethylpyrazine with trypsin. Luminescence 2014, 29, 79–86. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, W.; Qiu, L. Synthesis of butyl-isobutyl-phthalate and its interaction with α-glucosidase in vitro. J. Biochem. 2011, 149, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Ross, P.D.; Subramanian, S. Thermodynamics of protein association reactions: Forces contributing to stability. Biochemistry 1981, 20, 3096–3102. [Google Scholar] [CrossRef]
- Teng, Y.; Liu, R.; Yan, S.; Pan, X.; Zhang, P.; Wang, M. Spectroscopic investigation on the toxicological interactions of 4-aminoantipyrine with bovine hemoglobin. J. Fluoresc. 2010, 20, 381–387. [Google Scholar] [CrossRef]
- Pauff, J.M.; Cao, H.; Hille, R. Substrate orientation and catalysis at the molybdenum site in xanthine oxidase: Crystal structures in complex with xanthine and lumazine. J. Boil. Chem. 2009, 284, 8760–8767. [Google Scholar] [CrossRef]
- Si, Y.X.; Yin, S.J.; Park, D.; Chung, H.Y.; Yan, L.; Lü, Z.R.; Zhou, H.M.; Yang, J.M.; Qian, G.Y.; Park, Y.D. Tyrosinase inhibition by isophthalic acid: Kinetics and computational simulation. Int. J. Biol. Macromol. 2011, 48, 700–704. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Ma, Y. Mechanistic and conformational studies on the interaction of food dye amaranth with human serum albumin by multispectroscopic methods. Food Chem. 2013, 136, 442–449. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; Mooney, C.; Shields, D.C.; Fitzqerald, R.J. Inhibition of dipeptidyl peptidase IV and xanthine oxidase by amino acids and dipeptides. Food Chem. 2013, 141, 644–653. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Sample of butein is available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
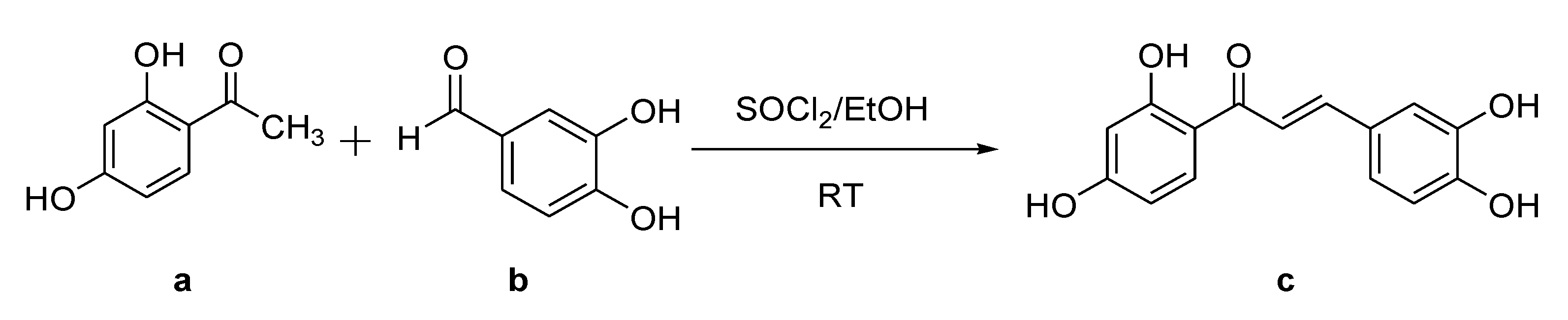{kind=link}
| Entry | SOCl2 (mol L−1) | Molar Ratio (a/b) | Time (h) | Yield (%) |
|---|---|---|---|---|
| 1 | 0.25 | 1:1 | 2 | 2 |
| 2 | 0.5 | 1:1 | 2 | 74 |
| 3 | 1 | 1:1 | 2 | 85 |
| 4 | 1.5 | 1:1 | 2 | 37 |
| 5 | 2 | 1:1 | 2 | 9 |
| 6 | 1 | 1:0.6 | 2 | 50 |
| 7 | 1 | 1:0.8 | 2 | 60 |
| 8 | 1 | 1:1.2 | 2 | 75 |
| 9 | 1 | 1:1.4 | 2 | 76 |
| 10 | 1 | 1:1 | 0.5 | 84 |
| 11 | 1 | 1:1 | 1 | 88 |
| 12 | 1 | 1:1 | 1.5 | 87 |
| T (K) | Kq (× 1012 L mol−1 S−1) | Ra | Ka (× 104 L mol−1) | Rb | ∆H (kJ mol−1) | n | ∆G (kJ mol−1) | ∆S (J mol−1 K−1) |
|---|---|---|---|---|---|---|---|---|
| 298 | 2.09 | 0.9953 | 2.96 | 0.9921 | 103.45 | 0.91 | −24.09 | 427.94 |
| 304 | 2.55 | 0.9878 | 3.86 | 0.9931 | 0.75 | −26.66 | ||
| 310 | 3.10 | 0.9944 | 10.51 | 0.9991 | 1.00 | −29.23 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, Y.-X.; Sun, S.-W.; Liu, Y.; Li, Y.; Liu, X.-H.; Wang, W.; Zhang, S.; Wang, W. An Improved Method for the Synthesis of Butein Using SOCl2/EtOH as Catalyst and Deciphering Its Inhibition Mechanism on Xanthine Oxidase. Molecules 2019, 24, 1948. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24101948
Hou Y-X, Sun S-W, Liu Y, Li Y, Liu X-H, Wang W, Zhang S, Wang W. An Improved Method for the Synthesis of Butein Using SOCl2/EtOH as Catalyst and Deciphering Its Inhibition Mechanism on Xanthine Oxidase. Molecules. 2019; 24(10):1948. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24101948
Chicago/Turabian StyleHou, Yu-Xue, Shi-Wei Sun, Yang Liu, Yan Li, Xiao-Hong Liu, Wei Wang, Shuang Zhang, and Wei Wang. 2019. "An Improved Method for the Synthesis of Butein Using SOCl2/EtOH as Catalyst and Deciphering Its Inhibition Mechanism on Xanthine Oxidase" Molecules 24, no. 10: 1948. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24101948