
Cytotoxicity of Triterpene Seco-Acids from Betula pubescens Buds
by
 ,
,
Łukasz Szoka
1, 
Valery Isidorov
2,*,
Jolanta Nazaruk
3,
Marcin Stocki
2 and
Leszek Siergiejczyk
4 1
Department of Medicinal Chemistry, Medical University of Bialystok, 15-222 Białystok, Poland
2
Forest Faculty, Białystok University of Technology, 17-200 Hajnówka, Poland
3
Department of Pharmacognosy, Medical University of Bialystok, 15-222 Białystok, Poland
4
Institute of Chemistry, University of Białystok, 15-245 Białystok, Poland
*
Author to whom correspondence should be addressed.
Molecules 2019, 24(22), 4060; https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24224060
Submission received: 9 October 2019
/
Revised: 1 November 2019
/
Accepted: 6 November 2019
/
Published: 9 November 2019
(This article belongs to the Collection Molecular Medicine)
Abstract
:The present study investigated the magnitude and mechanism of the cytotoxic effect on selected cancer cell lines of 3,4-seco-urs-4(23),20(30)-dien-3-oic acid (1), 3,4-seco-olean-4(24)-en-19-oxo-3-oic acid (2), and 3,4-seco-urs-4(23),20(30)-dien-19-ol-3-oic acid (3) isolated from downy birch (Betula pubescens) buds by carbon dioxide supercritical fluid extraction and gradient column chromatography. Cell viability in six human cancer lines exposed to these compounds was determined by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Apoptosis was quantified by annexin V/propidium iodide staining of gastric cancer AGS and colorectal cancer DLD-1 cells. To evaluate the mechanism of apoptosis, the expression of apoptosis-related proteins was analyzed by Western blot. Compound 1 exhibited non-specific toxicity, while compounds 2 and 3 were specifically toxic to colon and stomach cancer cells. The toxicity of compounds 2 and 3 against these two cell lines was greater than for compound 1. Cleavage of caspase-8, -9, and -3 was found in AGS and DLD-1 cells treated with all three seco-acids, indicating the induction of apoptosis via extrinsic and intrinsic pathways. Therefore, triterpene seco-acids (1–3) decreased cell viability by apoptosis induction. AGS and DLD-1 cells were more susceptible to seco-acids with an oxidized C19 than normal fibroblasts. Hence, it made them a new group of triterpenes with potential anticancer activity.
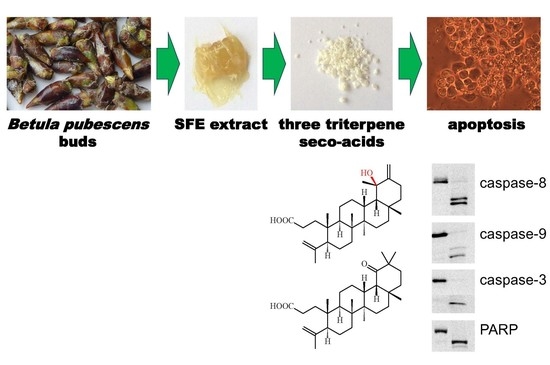
1. Introduction
Birch trees are deciduous hardwood species from the Betulaceae family and are widespread in the Northern Hemisphere. Since ancient times, different parts of these plants have been used in traditional medicine [1,2]. In particular, ethanol tinctures of winter buds have been used to treat toothache, colds, and rheumatoid conditions. A number of laboratory and clinical studies have confirmed the therapeutic properties of birch bud extracts, including their diuretic action as well as antiseptic and antioxidant properties [3,4,5,6]. In Russia, a birch bud preparation, Gemmae Betulae, is a standardized medicine used mainly to treat urinary tract diseases [7].
In recent decades, a number of studies have been conducted on the anti-cancer activity of birch buds [8,9,10,11]. In particular, it was found that various extracts from the buds of two species of white birch exhibited distinct time- and concentration-dependent cytotoxicity with respect to many human cancer cell lines [10,11]. At the same time, the highest cytotoxic activity was demonstrated by extracts obtained by carbon dioxide supercritical fluid extraction (SFE). These results suggest that birch buds are a promising source of compounds with cytotoxic activity against various forms of cancer. Notably, birch buds have a high content of triterpene compounds [12,13]. Many members of this group of secondary metabolites have well-documented anti-cancer activity [14,15,16]. However, to date, research has been limited to the action of whole extracts obtained from birch buds, and no attempt has been made to study the anti-cancer effect of individual constituent compounds.
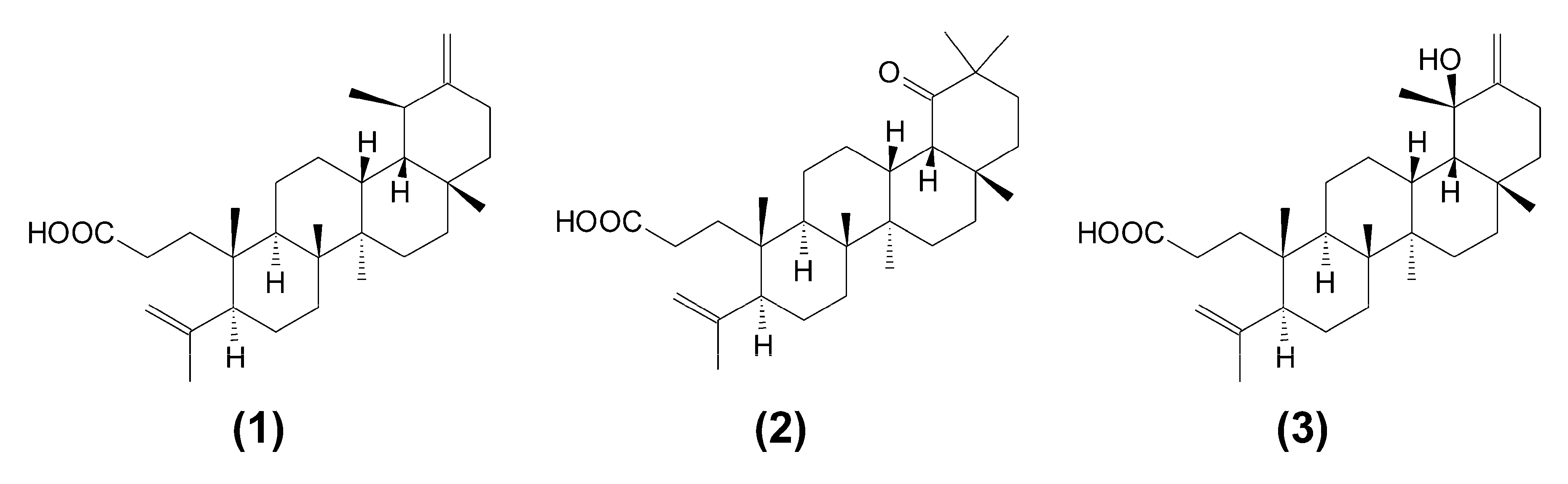
This work reports on the cytotoxic effect of triterpene seco-acids (1–3), which were isolated for the first time from buds of downy birch, Betula pubescens, as seen in Figure 1.
2. Results
2.1. Triterpene Seco-Acids Decreased Viability of Cancer Cells
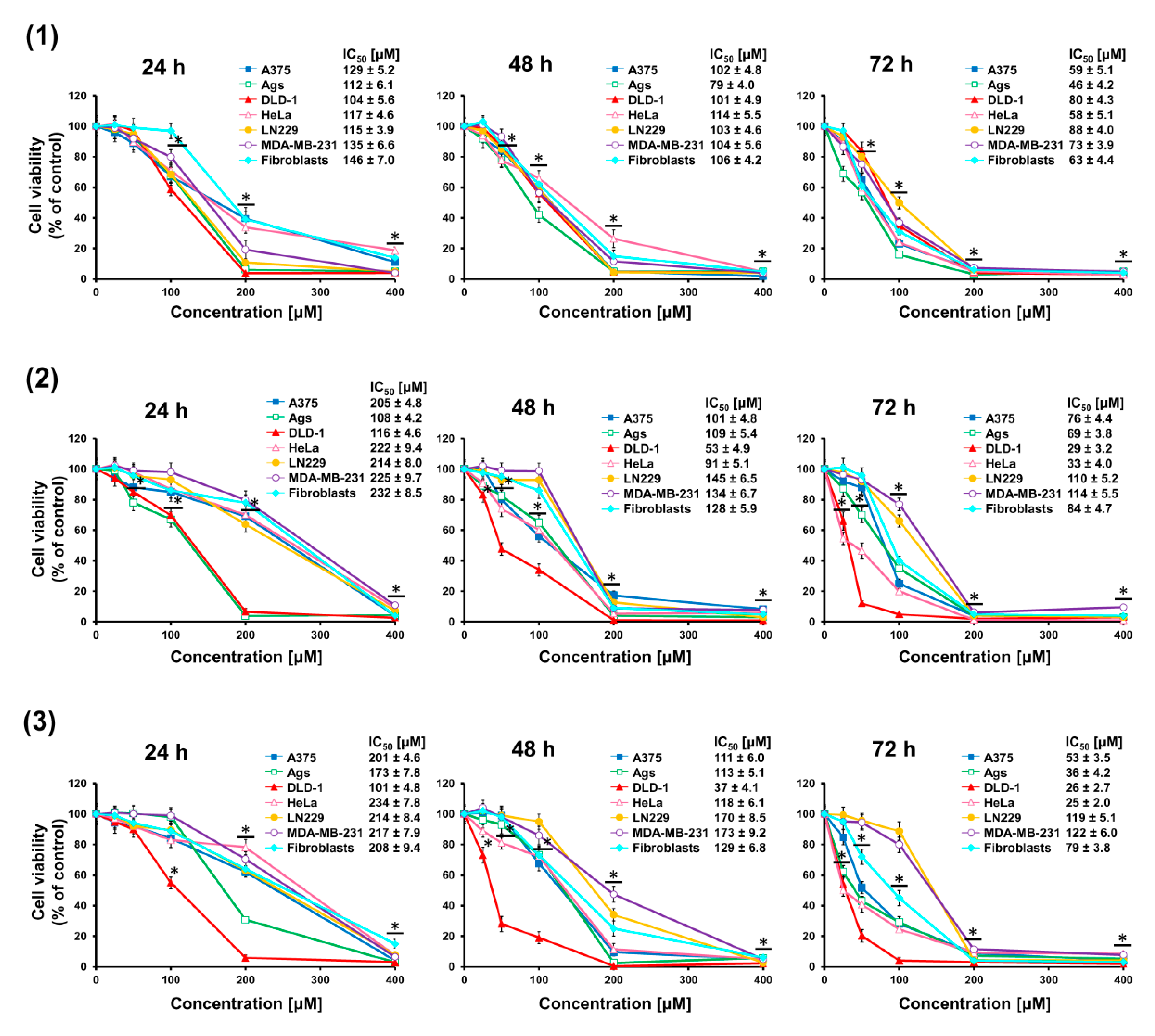
Cancer cells A375, AGS, DLD-1, HeLa, LN229, MDA-MB-231, and normal skin fibroblasts were treated with increasing doses (25–400 μM) of seco-acids (compounds 1–3) for 24, 48, and 72 h, and changes in cell viability were quantified using the MTT assay, as depicted in Figure 2. A dose- and time-dependent decrease in cell viability was observed. Based on the dose–response curves, the IC50 values were calculated. It was found that the decrease in viability of cells treated with seco-acid (1) was similar. By contrast, bifunctional compounds keto-seco-acid (2) and hydroxy-seco-acid (3) exerted higher activity against DLD-1 cells. In addition, DLD-1 cells were more sensitive to the action of these seco-acids than normal fibroblasts. The most relevant IC50 values are presented in Table 1.
2.2. Triterpene Seco-Acids Increased Apoptosis in AGS and DLD-1 Cells
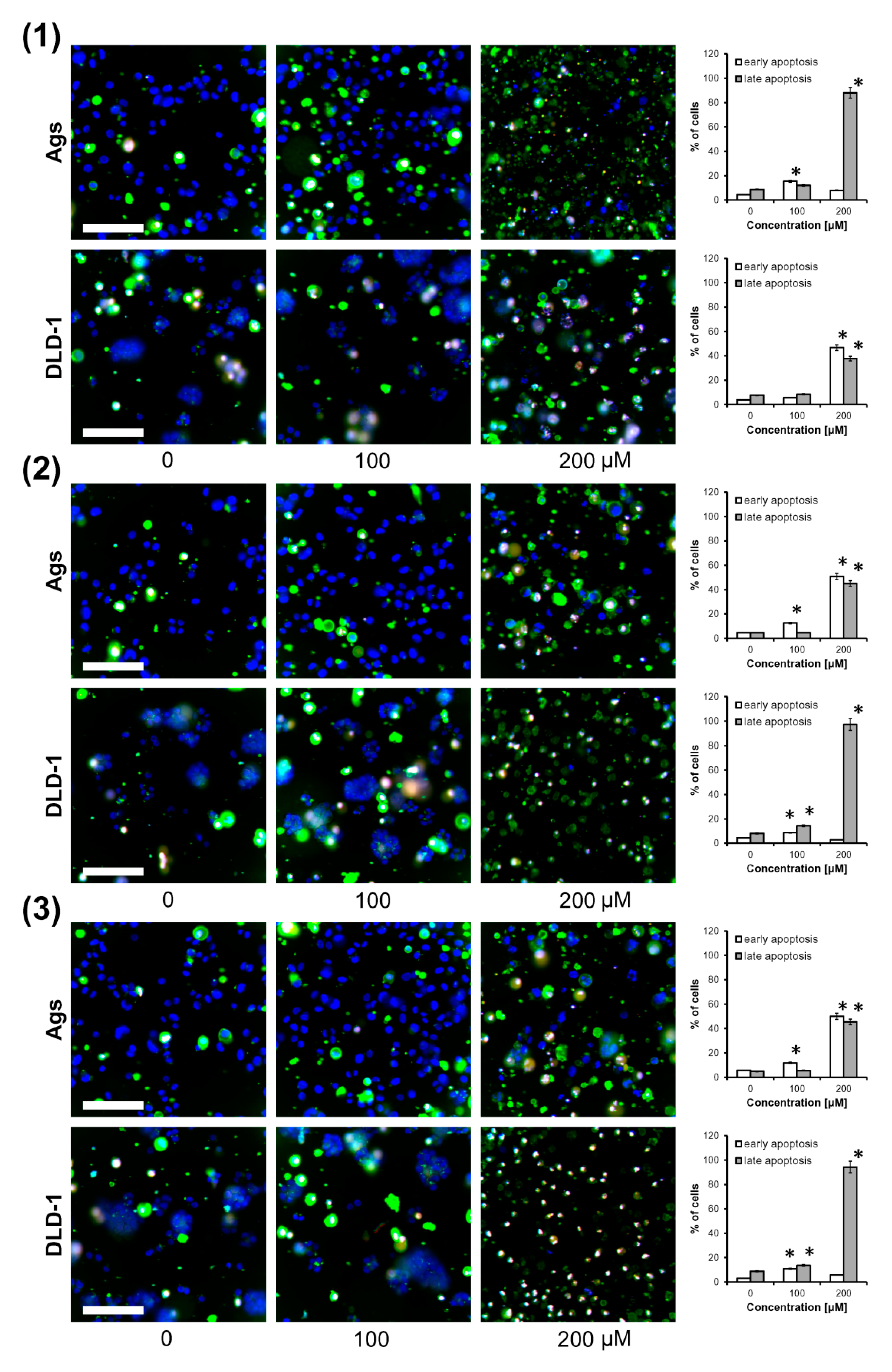
Since birch bud extracts were used in Russian medicine as a diuretic agent taken by mouth [2,3], gastric cancer cells (AGS) and colorectal cancer cells (DLD-1) were chosen for the examination of the apoptotic activity of triterpene seco-acids isolated from them. Apoptosis was quantified by triple staining of cells with annexin V-FITC conjugate, propidium iodide (PI), and Hoechst 33342, and cells were visualized by fluorescent microscopy, as depicted in Figure 3. Apoptotic cells were characterized by the exposure of phosphatidylserine on the outer leaflet of the cellular membrane. Thus, annexin V, a natural ligand of phosphatidylserine, was used for their detection. Membrane integrity in early apoptotic cells made them impermeable to PI; however, DNA of late apoptotic or dead cells could bind it. In contrast to PI, Hoechst 33342 had the ability to bind the DNA of living and dead cells. We found that after 24 h of treatment, triterpene seco-acids increased apoptotic cell death in a dose-dependent manner. Compound 1 at 200 μM induced a late apoptotic stage in almost all AGS cells, while the number of early and late apoptotic DLD-1 cells was similar. In contrast, treatment of AGS cells with compounds 2 and 3 at 200 μM resulted in a similar increase in the number of early and late apoptotic cells, and put almost all DLD-1 cells into a late apoptotic stage.
2.3. Triterpene Seco-Acids Activated Extrinsic and Intrinsic Pathway of Apoptosis in AGS and DLD-1 Cells
To evaluate the mechanism of apoptosis induction by triterpene seco-acids in AGS and DLD-1 cells, the expression of apoptosis-related proteins was analyzed by Western blot, as shown in Figure 4. The final stage of apoptosis was carried out by caspases—a group of cysteine-aspartyl-specific proteases. Activation of caspases depended on intrachain cleavage of inactive zymogens to form active cleaved caspases. There were two major apoptosis-triggering pathways: extrinsic and intrinsic.
The extrinsic pathway was activated by ligands of transmembrane death receptors, resulting in activation of caspase-8. Increase in caspase-8 processing was observed by all triterpene seco-acids in both cell lines used in the study, with the data suggesting induction of the extrinsic apoptosis pathway.
Multiple internal stimuli are known to trigger the intrinsic pathway of apoptosis by increasing the permeability of outer mitochondrial membranes. This resulted in translocation of cytochrome c into the cytoplasm and caspase-9 activation. Among intrinsic pathway-inducing proteins, expression levels of BID and p53 were assayed. BID was previously shown to be cleaved by caspase-8 to form truncated BID (tBID), which translocated into mitochondrial membranes, resulting in cytochrome c release. Our data showed a decrease in BID expression in both lines, except for seco-acid (1)-treated AGS cells. By contrast, there was no increase of p53 level in wild-type p53-containing AGS cells or in DLD-1 cells carrying an inactivating mutation in the TP53 gene [17]. Thus, BID but not p53 could mediate induction of the intrinsic pathway. Finally, we found an increase in caspase-9 cleavage in both cell lines treated with all three triterpene seco-acids, indicating intrinsic pathway induction.
Both apoptosis pathways were previously shown to converge in the induction of executioner caspases, such as caspase-3 and caspase-7, in turn catalyzing the proteolytic degradation of many cellular components, leading to cell death. Treatment with the three triterpene seco-acids induced an increase in active cleaved caspase-3 levels in AGS and DLD-1 cells with a concomitant decrease in expression of caspase-3 pro-form in DLD-1 cells. Although cleaved caspase-7 was not detected in DLD-1 cells after 24 h of treatment, the observed decrease in its pro-form level indicates caspase-7 activation. In contrast, treatment of AGS cells with seco-acids resulted in not only increased levels of cleaved caspase-3 but also increased expression of executioner caspase pro-forms. Cleavage of poly(ADP-ribose) polymerase (PARP), the substrate for caspase-3, was concomitant with caspase-3 processing in both cell lines, indicating enzymatic activation of caspase-3. Taken together, these data suggest the activation of executioner caspases through the extrinsic and intrinsic pathways of apoptosis. The highest levels of cleaved caspase-8, -9, and -3 and cleaved PARP were observed in AGS cells treated with compound 1, and in DLD-1 cells treated with compounds 2 and 3. Thus, cells recognized as late apoptotic on annexin V and PI staining had higher levels of apoptosis markers.
3. Discussion
The genus Betula is a rich source of triterpenes that exert cytotoxic activity and are thus considered as potential anticancer agents, with betulinic acid being a well-known example [18]. In turn, less-active compounds such as betulin, which is the major constituent of the outer bark of birch trees, are used for obtaining semi-synthetic derivatives with promising activity [19,20]. Triterpene biosynthesis pathways vary depending on the plant organ [21]; hence, searching for triterpenes is not limited to birch bark. Indeed, the presence of both ursane and oleanane triterpenes was reported in floral spikes of Betula platyphylla [16]. Triterpenes belonging to the oleanane and lupane groups, besides ursane derivatives, are also of interest due to their possible use in cancer therapy [15].
Birch buds seem to be another interesting source of triterpenes. In a recent paper [11], the cytotoxicity of various birch bud extracts against a wide range of cancer cells was demonstrated. In the current study, we isolated triterpene seco-acids (compounds 1–3) from B. pubescens buds and determined their cytotoxicity to cancer cells and normal fibroblasts.
One of the isolated compounds, seco-acid 1 was previously reported to show weak cellular toxicity in multiple cell lines [16]. Our data supported these findings; however, we also demonstrated a time dependency in all tested cell lines. Moreover, the addition of a hydroxyl group to C19 of seco-acid 1 caused an increase in cytotoxicity in some of the cell lines—particularly in colorectal and gastric cancer cells. Therefore, the incorporation of polar groups into ursane type seco-acids, as in compound 3, might have a beneficial impact on the possible application range. Additionally, it seemed that this feature was not unique to ursane-type seco-acids, because oleanane type derivative 2, characterized by a keto group at C19, retained high cytotoxicity in colorectal cancer culture. This was consistent with an earlier report that an increase in the polarity of triterpenes causes an increase in anticancer activity [22].
Cancer cells are characterized by apoptosis resistance, and apoptosis induction is a common mechanism of anticancer drugs [23]. Apoptosis is a highly regulated process characterized by membrane blebbing, chromatin condensation, and cell shrinkage [24]. One of the most important events defining cell fate is the loss of phospholipid asymmetry with the externalization of phosphatidylserine, which functions as a chemical signal for phagocytosis [24]. Oleanane and ursane triterpenes have been shown to promote apoptosis in many cancer cells [15,16]. In the current study, both colorectal and gastric cancer cells treated with triterpene seco-acids showed externalization of phosphatidylserine, indicating apoptosis. Moreover, to different extents, cells lost membrane integrity—a phenomenon common to the late stage of apoptosis or necrosis.
The majority of apoptosis events are driven by caspases. These members of the cysteine proteases exist in the cytoplasm as zymogens [24]. Executioner caspases (e.g., caspase-3 and -7) catalyze the degradation of enzymes and structural proteins. The activation of executioner caspases is regulated by initiator caspases (e.g., caspase-8 and -9). Oleanane and ursane triterpenoids have been reported to activate the intrinsic pathway of apoptosis [15]. In addition, the three triterpene seco-acids examined in the current study acted as activators of caspase-9, which was a mediator of the intrinsic pathway [25]. Consequently, activation of caspase-3, -7 and degradation of PARP occurred in DLD-1 and AGS cells. Notably, the level of p53 was unchanged in both cell lines; however, BID was down-regulated. tBID serves as a switch for activation of the intrinsic pathway by caspase-8, which in turn acts as the main intermediate of the extrinsic pathway [26,27]. It was reported that triterpenes can promote apoptosis via the extrinsic pathway [28]. Indeed, activation of caspase-8 by all three triterpene seco-acids was observed in the current study. Accordingly, apoptosis induced by the triterpene seco-acids was the result of the activation of both extrinsic and intrinsic pathways.
In conclusion, this study showed that triterpene seco-acids decreased cell viability by apoptosis induction in gastric and colorectal cancer cells. Furthermore, these cancer cells were more susceptible to the toxicity of seco-acids when they had an oxidized C19, compared to normal fibroblasts. As such, they might be considered for further investigations as a novel group of drug candidates for the treatment of colorectal cancer.
4. Materials and Methods
4.1. Reagents and Chemicals
Stationary phases and eluents used in column chromatography and extraction were: silica gel (mesh 0.063–0.2 mm, Merck, Darmstadt, Germany); polyamide (Roth, Karlsrue, Germany); n-hexane, diethyl ether, benzene, and chloroform (Chempur, Pekary Śląskie, Poland); methanol, ethyl acetate, and acetone (POCH, Gliwice, Poland).
Antibody against β-actin (A2066, dilution 1:2000), horseradish peroxidase-conjugated secondary antibodies, anti-rabbit (A9169, dilution 1:5000) and anti-mouse (A9044, dilution 1:5000), and 3-(4,5-dimethylthiazole-2-yl)-2,5-diphenyltetrazolium bromide (MTT) were obtained from Sigma-Aldrich (Poznań, Poland). Dulbecco’s modified Eagle’s medium (DMEM), penicillin, streptomycin, fetal bovine serum (FBS), phosphate-buffered saline, trypsin, and Hoechst 33342 were purchased from Thermo Fisher Scientific (Warszawa, Poland). Antibodies against caspase-3 (9662, dilution 1:1000), cleaved caspase-3 (9664, dilution 1:1000), caspase-7 (12827, dilution 1:1000), caspase-8 (9746, dilution 1:1000), caspase-9 (9508, dilution 1:1000), Bcl-2 protein family member, BID (2002, dilution 1:1000), poly(ADP-ribose) polymerase (PARP; 9542, dilution 1:1000) and cleaved PARP (5625, dilution 1:1000) were obtained from Cell Signaling Technology (Danvers, MA, USA). Antibodies against p53 (sc-126, dilution 1:1000) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
4.2. Plant Material
Buds of downy birch (B. pubescens Ehrh.) were gathered in August 2015 from trees growing in a non-protected area of the Biebrza National Park in north-eastern Poland (53° 32’ N, 22° 43’ E). A voucher specimen (No. BO-17035) was deposited with the herbarium of the Department of Pharmacognosy, Medical University of Bialystok, Poland. The birch species was identified using the approach described in [13].
4.3. Bud Extraction and Chemical Analysis
Downy birch buds (600 g) were ground and extracted by carbon dioxide SFE on a Waters SFE-1000F-2-FMC50 (Milford, MA, USA) system at 40 °C and a pressure of 300 bar. The dry residue from SFE was further extracted with n-hexane in a Soxhlet apparatus. The SFE extract (90 g) was subjected to Sephadex LH-20 gel filtration with chloroform:methanol (1:1) elution. The hexane extract (210 g) was separated on silica gel columns, eluted with hexane−chloroform, and then exposed to a chloroform−ethyl acetate step gradient (the analytical procedure is described in detail in [29]). Fractions containing triterpene compounds were subjected to further separation on polyamide columns with benzene−methanol as eluents. After multistep chromatographic separations and re-crystallization from ethanol, pure 3,4-seco-urs-4(23),20(30)-dien-3-oic acid (compound 1), 3,4-seco-olean-4(24)-en-19-oxo-3-oic acid (compound 2), and 3,4-seco-urs-4(23),20(30)-dien-19-ol-3-oic acid (compound 3) were isolated (Figure 1). The composition of the collected fractions, as well as the purity of the isolated seco-acids (1–3) were determined by a gas chromatography–mass spectrometry (GC-MS) method (for details, see Supplementary Information). Mass spectra of isolated seco-acids, as their trimethylsilyl (TMS) derivatives, are also presented as Supplementary Information (Figures EMS-1–EMS-3). Table S1 contains the 13C nuclear magnetic resonance (NMR) chloroform solvent (CDCl3) spectra of the obtained compounds registered on a Bruker Advance II 400 spectrometer at 100 MHz. Results of NMR identification agreed with previous reports [12,16]. This was the first isolation of these triterpene seco-acids from B. pubescens buds.
4.4. Cell Culture and Treatment
Breast cancer MDA-MB-231 cells, colorectal cancer DLD-1 cells, gastric cancer AGS cells, glioblastoma LN229 cells, cervix cancer HeLa cells, and human skin fibroblasts CCD-25Sk were obtained from the American Type Culture Collection. Melanoma A375 cells were purchased from Sigma-Aldrich (Poznań, Poland). Cells were cultured in DMEM supplemented with 10% FBS, 100 units/mL penicillin and 100 μg/mL streptomycin in a humidified 5% CO2 atmosphere at 37 °C. Triterpene seco-acids were dissolved in dimethyl sulfoxide (DMSO) and added to the culture medium at each designated time. The final DMSO concentration in the medium for the control and compound-treated cells was 0.1%.
4.5. Cell Viability Assay
Viability of cells was determined by MTT assay [30]. Cells were cultured in 96-well plates at 1 × 104 cells per well for 24 h, and then treated with triterpene seco-acids. After 24, 48, or 72 h, MTT solution was added to each well and cells were incubated at 37 °C for 4 h. The medium was then removed and formazan crystals were dissolved in 100 μL of DMSO and 12.5 μL of Sorensen’s glycine buffer on a plate shaker. Optical density was measured in an Asys UVM340 microplate reader (Biochrom Ltd., Cambridge, UK) at 570 nm.
4.6. Apoptosis Assay
Cells were seeded at a density of 1 × 105 cells per well in 6-well plates. After 24 h, the medium was changed and triterpene seco-acids were added for the next 24 h. Floating and adherent cells were collected and assayed with a Dead Cell Apoptosis Kit with annexin V-fluorescein isothiocyanate (FITC) and propidium iodide (PI) for flow cytometry (Thermo Fisher Scientific, Warszawa, Poland), according to the manufacturer’s protocol, as described in [19]. Briefly, cells were dispersed in 100 μL annexin-binding buffer containing 5 μL annexin V-FITC conjugate solution, 1 μg/mL PI, and 1 μg/mL Hoechst 33342, and incubated for 15 min at room temperature. Then, 400 μL annexin-binding buffer was added, and the cells were transferred to 96-well plates and visualized by fluorescent microscopy on a BD Pathway 855 system (Becton Dickinson, San Jose, CA, USA). Early apoptotic cells showed green and blue fluorescence, while late apoptotic cells showed green, red, and blue fluorescence.
4.7. Western Blot
Protein concentrations in the samples were determined by the method of Lowry et al. [31]. Proteins were resolved by sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE), transferred to nitrocellulose membranes, and probed with primary antibodies overnight at 4 °C. Afterwards, secondary antibodies were added for 1 h. Signals were visualized using Enhanced Chemiluminescence (ECL) reagent purchased from GE Healthcare (Waukesha, WI, USA) and recorded with a BioSpectrum Imaging System (Ultra-Violet Products, Ltd., Murray, UT, USA).
4.8. Statistical Analysis
The results were submitted to statistical analysis using one-way analysis of variance (ANOVA) followed by Tukey’s test, accepting p < 0.05 as a significant difference in comparison with controls. Half-maximal inhibitory concentration (IC50) values were calculated by nonlinear regression analysis using GraphPad Prism version 7.04 (GraphPad Software, San Diego, CA, USA).
Supplementary Materials
The following are available online. Text S1: Gas chromatographic–mass spectrometric analysis of separated fractions and isolated compounds; Text S2: Spectroscopic and gas-chromatographic characteristics of triterpene seco-acids (1–3) isolated from Betula pubescens buds; Table S1: Data on 13C NMR (100 MHz, CDCl3) spectra of triterpene seco-acids (1–3).
Author Contributions
Conceptualization, V.I. and Ł.S.; Development of methodology, J.N. and Ł.S.; Sample preparation: M.S.; Acquisition of data: Ł.S. (cytotoxicity assays), V.I. (GC-MS), L.S. (NMR); Analysis and interpretation of data: Ł.S. and V.I.; Writing—original draft preparation: V.I. and Ł.S.; Writing—review and editing: V.I.
Funding
This research was funded by the National Science Centre (Poland), grant number 2016/23/B/NZ7/03360 as well as by Białystok University of Technology Program S/ZWL/1/2017.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Culpeper, N. Culpeper’s Complete Herbal: A Book of Natural Remedies of Ancient Ills; (The Wordsworth Collection Reference Library); NTC/Contemporary Publishing Company (Reprint of 1653 Work): London, UK, 1995. [Google Scholar]
- Shikov, A.N.; Pozharitskaya, O.N.; Makarov, V.G.; Wagner, H.; Verpoorte, R.; Heinrich, M. Medicinal plants of the Russian Pharmacopoeia; their history and applications. J. Ethnopharmacol. 2014, 154, 481–536. [Google Scholar] [CrossRef] [PubMed]
- Raal, A. Birch (Betula spp.). In Recent Progress in Medical Plants; Awaad, A.S., Singh., V.K., Govil, J.N., Eds.; Drug Plants II. Studium Press LLC: Houston, TX, USA, 2010; Volume 28, pp. 123–142. [Google Scholar]
- Mashentseva, A.A.; Dehaen, W.; Seitembetov, T.S.; Seitembetova, A.J. Comparison of the antioxidant activity of the different Betula pendula Roth. extracts from northern Kazakhstan. J. Phytol. 2011, 3, 18–25. [Google Scholar]
- Duric, K.; Kovac-Besovic, E.; Niksic, H.; Sofic, E. Antibacterial activity of methanolic extracts, decoction and isolated triterpene products from different parts birch, Betula pendula Roth. J. Plant. Stud. 2013, 2, 61–70. [Google Scholar] [CrossRef]
- Tolmacheva, A.A.; Rogozhin, E.A.; Deryabin, D.G. Antibacterial and quorum sensing regulatory activities of some traditional Eastern-European medicinal plants. Acta Pharm. 2014, 64, 173–186. [Google Scholar] [CrossRef] [Green Version]
- State Pharmacopoeia of USSR, 11-th ed.; Medizina: Moscow, Russia, 1990; Volume 2. (In Russian)
- Goun, E.A.; Petrichenko, V.M.; Solodnikov, S.U.; Suhinina, T.V.; Kline, M.A.; Cunningham, G.; Nguyen, C.; Miles, H. Anticancer and antithrombin activity of Russian plants. J. Ethnopharmacol. 2002, 81, 337–342. [Google Scholar] [CrossRef]
- Spiridonov, N.A.; Konovalov, D.A.; Arkhipov, V.V. Cytotoxicity of some Russian ethnomedicinal plants and plant compounds. Phytother. Res. 2005, 19, 428–432. [Google Scholar] [CrossRef]
- Isidorov, V.A.; Bakier, S.; Moskwa, J.; Borawska, M.H. Anticancer activity of some plant extracts on U87MG human gliobastoma cell. In Book of Abstract of 6th Rus.-Korean Conf. Current Issues of Biologically Active Compound Chemistry and Biotechnology; Novosibirsk Institute of Organic Chemistry: Novosibirsk, Russua, 2015; p. 189. [Google Scholar]
- Isidorov, V.; Szoka, Ł.; Nazaruk, J. Cytotoxicity of white birch bud extracts: Perspectives for therapy of tumors. PLoS ONE 2018, 13, e0201949. [Google Scholar] [CrossRef]
- Vedernikov, D.N.; Roshchin, V.I. Extractive compounds of Betulaceae family buds (Betula pendula Roth.): V. Composition of triterpene seco-acids. Russ. J. Bioorg. Chem. 2012, 38, 762–768. [Google Scholar] [CrossRef]
- Isidorov, V.; Szczepaniak, L.; Wróblewska, A.; Pirożnikow, E.; Vetchinnikova, L. Gas chromatographic-mass spectrometric examination of chemical composition of two Eurasian birch (Betula L.) bud exudates and its taxonomical implication. Biochem. System. Ecol. 2014, 52, 41–48. [Google Scholar] [CrossRef]
- Prokofʹeva, N.G.; Chaikina, E.L.; Pokhilo, N.D.; Anisimov, M.M. Hemolytic and cytotoxic activity of dammarane-type triterpenoids. Chem. Nat. Comp. 2007, 43, 72–75. [Google Scholar] [CrossRef]
- Laszczyk, M.N. Pentacyclic triterpenes of the lupane, oleanane and ursane group as tools in cancer therapy. Planta Med. 2009, 75, 1549–1560. [Google Scholar] [CrossRef] [PubMed]
- Kashiwada, Y.; Sekiya, M.; Yamazaki, K.; Ikeshiro, Y.; Fujioka, T.; Yamagishi, T.; Kitagawa, S.; Takaishi, Y. Triterpenoids from the floral spikes of Betula platyphylla var. japonica and their reversing activity against multidrug-resistant cancer cells. J. Nat. Prod. 2007, 70, 623–627. [Google Scholar] [CrossRef] [PubMed]
- Bouaoun, L.; Sonkin, D.; Ardin, M.; Hollstein, M.; Byrnes, G.; Zavadil, J.; Olivier, M. TP53 Variations in Human Cancers: New Lessons from the IARC TP53 Database and Genomics Data. Hum. Mutat. 2016, 37, 865–876. [Google Scholar] [CrossRef] [PubMed]
- Ali-Seyed, M.; Jantan, I.; Vijayaraghavan, K.; Bukhari, S.N. Betulinic acid: Recent advances in chemical modifications, effective delivery, and molecular mechanisms of a promising anticancer therapy. Chem. Biol. Drug Des. 2016, 87, 517–536. [Google Scholar] [CrossRef]
- Szoka, L.; Karna, E.; Hlebowicz-Sarat, K.; Karaszewski, J.; Boryczka, S.; Palka, J.A. Acetylenic derivative of betulin induces apoptosis in endometrial adenocarcinoma cell line. Biomed. Pharmacother. 2017, 95, 429–436. [Google Scholar] [CrossRef]
- Yang, S.J.; Liu, M.C.; Xiang, H.M.; Zhao, Q.; Xue, W.; Yang, S. Synthesis and in vitro antitumor evaluation of betulin acid ester derivatives as novel apoptosis inducers. Eur. J. Med. Chem. 2015, 102, 249–255. [Google Scholar] [CrossRef]
- Thimmappa, R.; Geisler, K.; Louveau, T.; O’Maille, P.; Osbourn, A. Triterpene biosynthesis in plants. Annu. Rev. Plant. Biol. 2014, 65, 225–257. [Google Scholar] [CrossRef]
- Yang, H.; Kim, H.W.; Kim, Y.C.; Sung, S.H. Cytotoxic activities of naturally occurring oleanane-, ursane-, and lupane-type triterpenes on HepG2 and AGS cells. Pharmacogn. Mag. 2017, 13, 118–122. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, R.W.; Ruefl, A.A.; Lowe, S. Apoptosis: A link between cancer genetics and chemotherapy. Cell. 2002, 108, 153–164. [Google Scholar] [CrossRef]
- Fadeel, B.; Orrenius, S. Apoptosis: A basic biological phenomenon with wide-ranging implications in human disease. J. Intern. Med. 2005, 258, 479–517. [Google Scholar] [CrossRef]
- Kroemer, G.; Reed, J.C. Mitochondrial control of cell death. Nat. Med. 2000, 6, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Nagata, S. Apoptosis by death factor. Cell 1997, 88, 355–365. [Google Scholar] [CrossRef]
- Schug, Z.T.; Gonzalvez, F.; Houtkooper, R.H.; Vaz, F.M.; Gottlieb, E. BID is cleaved by caspase-8 within a native complex on the mitochondrial membrane. Cell Death Differ. 2011, 18, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Mishra, T.; Arya, R.K.; Meena, S.; Joshi, P.; Pal, M.; Meena, B.; Upreti, D.K.; Rana, T.S.; Datta, D. Isolation, characterization and anticancer potential of cytotoxic triterpenes from Betula utilis bark. PLoS ONE 2016, 11, e0159430. [Google Scholar] [CrossRef] [PubMed]
- Isidorov, V.; Stocki, M.; Nazaruk, J.; Bakier, S. Metabolomic GC-MS study of Betula pubescens bud extracts. Trees 2019. submitted. [Google Scholar]
- Plumb, J.A.; Milroy, R.; Kaye, S.B. Effects of the pH dependence of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide-formazan absorption on chemosensitivity determined by a novel tetrazolium-based assay. Cancer Res. 1989, 49, 4435–4440. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
Sample Availability: Samples of the compounds 1,3 are available from the authors. |
Figure 1.
Chemical structure of triterpene seco-acids: 3,4-seco-urs-4(23),20(30)-dien-3-oic acid (1), 3,4-seco-olean-4(24)-en-19-oxo-3-oic acid (2), and 3,4-seco-urs-4(23),20(30)-dien-19-ol-3-oic acid (3).
Figure 1.
Chemical structure of triterpene seco-acids: 3,4-seco-urs-4(23),20(30)-dien-3-oic acid (1), 3,4-seco-olean-4(24)-en-19-oxo-3-oic acid (2), and 3,4-seco-urs-4(23),20(30)-dien-19-ol-3-oic acid (3).

Figure 2.
Triterpene seco-acids decrease cell viability. Effect of 3,4-seco-urs-4(23),20(30)-dien-3-oic acid (1), 3,4-seco-olean-4(24)-en-19-oxo-3-oic acid (2), and 3,4-seco-urs-4(23),20(30)-dien-19-ol-3-oic acid (3) on the viability of cancer cells A375, AGS, DLD-1, HeLa, LN229, MD-MB-231 and normal dermal fibroblasts after 24, 48, and 72 h treatment, as assessed by MTT assay. Data are presented as mean ± standard error of the mean (SEM) from three independent experiments. * p < 0.05 compared to control group. The half-maximal inhibitory concentration (IC50) values of seco-acids for the cell lines are shown.
Figure 2.
Triterpene seco-acids decrease cell viability. Effect of 3,4-seco-urs-4(23),20(30)-dien-3-oic acid (1), 3,4-seco-olean-4(24)-en-19-oxo-3-oic acid (2), and 3,4-seco-urs-4(23),20(30)-dien-19-ol-3-oic acid (3) on the viability of cancer cells A375, AGS, DLD-1, HeLa, LN229, MD-MB-231 and normal dermal fibroblasts after 24, 48, and 72 h treatment, as assessed by MTT assay. Data are presented as mean ± standard error of the mean (SEM) from three independent experiments. * p < 0.05 compared to control group. The half-maximal inhibitory concentration (IC50) values of seco-acids for the cell lines are shown.

Figure 3.
Triterpene seco-acids promote apoptosis in AGS and DLD-1 cells. AGS and DLD-1 cells treated with 3,4-seco-urs-4(23),20(30)-dien-3-oic acid (1), 3,4-seco-olean-4(24)-en-19-oxo-3-oic acid (2), and 3,4-seco-urs-4(23),20(30)-dien-19-ol-3-oic acid (3) for 24 h were triple stained with annexin V-FITC conjugate (green fluorescence), propidium iodide (red fluorescence), and Hoechst 33342 (blue fluorescence), and were visualized by fluorescence microscopy. Representative merged photographs are shown. Scale bar = 100 μm. Data are presented as mean ± standard deviation (SD) from three assays. * p < 0.05 compared to control group.
Figure 3.
Triterpene seco-acids promote apoptosis in AGS and DLD-1 cells. AGS and DLD-1 cells treated with 3,4-seco-urs-4(23),20(30)-dien-3-oic acid (1), 3,4-seco-olean-4(24)-en-19-oxo-3-oic acid (2), and 3,4-seco-urs-4(23),20(30)-dien-19-ol-3-oic acid (3) for 24 h were triple stained with annexin V-FITC conjugate (green fluorescence), propidium iodide (red fluorescence), and Hoechst 33342 (blue fluorescence), and were visualized by fluorescence microscopy. Representative merged photographs are shown. Scale bar = 100 μm. Data are presented as mean ± standard deviation (SD) from three assays. * p < 0.05 compared to control group.

Figure 4.
Triterpene seco-acids trigger extrinsic and intrinsic pathways of apoptosis in AGS and DLD-1 cells. AGS and DLD-1 cells were treated with 3,4-seco-urs-4(23),20(30)-dien-3-oic acid (1), 3,4-seco-olean-4(24)-en-19-oxo-3-oic acid (2), and 3,4-seco-urs-4(23),20(30)-dien-19-ol-3-oic acid (3) for 24 h, and the expression levels of apoptosis-related proteins were determined by Western blotting. Antibodies against caspase-8, BID, p53, caspase-9, caspase-3, cleaved caspase-3, caspase-7, PARP and cleaved PARP were used. The expression of β-actin served to normalize protein loading.
Figure 4.
Triterpene seco-acids trigger extrinsic and intrinsic pathways of apoptosis in AGS and DLD-1 cells. AGS and DLD-1 cells were treated with 3,4-seco-urs-4(23),20(30)-dien-3-oic acid (1), 3,4-seco-olean-4(24)-en-19-oxo-3-oic acid (2), and 3,4-seco-urs-4(23),20(30)-dien-19-ol-3-oic acid (3) for 24 h, and the expression levels of apoptosis-related proteins were determined by Western blotting. Antibodies against caspase-8, BID, p53, caspase-9, caspase-3, cleaved caspase-3, caspase-7, PARP and cleaved PARP were used. The expression of β-actin served to normalize protein loading.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
IC50 values (μM) of compounds 2 and 3 for fibroblasts and selected cancer cell lines after 48 and 72 h of treatment.
Table 1.
IC50 values (μM) of compounds 2 and 3 for fibroblasts and selected cancer cell lines after 48 and 72 h of treatment.
| Compound 2 48 h | Compound 2 72 h | Compound 3 48 h | Compound 3 72 h | |
|---|---|---|---|---|
| Fibroblasts | 128 ± 5.9 | 84 ± 4.7 | 129 ± 6.8 | 79 ± 3.8 |
| A.375 | 101 ± 4.8 | 76 ± 4.4 | 111 ± 6.0 | 53 ± 3.5 |
| Ags | 109 ± 5.4 | 69 ± 3.8 | 113 ± 5.1 | 36 ± 4.2 |
| DLD-1 | 53 ± 4.9 | 29 ± 3.2 | 37 ± 4.1 | 26 ± 2.7 |
| HeLa | 91 ± 5.1 | 33 ± 4.0 | 118 ± 6.1 | 25 ± 2.0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Szoka, Ł.; Isidorov, V.; Nazaruk, J.; Stocki, M.; Siergiejczyk, L. Cytotoxicity of Triterpene Seco-Acids from Betula pubescens Buds. Molecules 2019, 24, 4060. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24224060
AMA Style
Szoka Ł, Isidorov V, Nazaruk J, Stocki M, Siergiejczyk L. Cytotoxicity of Triterpene Seco-Acids from Betula pubescens Buds. Molecules. 2019; 24(22):4060. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24224060
Chicago/Turabian StyleSzoka, Łukasz, Valery Isidorov, Jolanta Nazaruk, Marcin Stocki, and Leszek Siergiejczyk. 2019. "Cytotoxicity of Triterpene Seco-Acids from Betula pubescens Buds" Molecules 24, no. 22: 4060. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24224060