
The Anti-Aging Potential of Neohesperidin and Its Synergistic Effects with Other Citrus Flavonoids in Extending Chronological Lifespan of Saccharomyces Cerevisiae BY4742


Abstract
:1. Introduction
2. Results
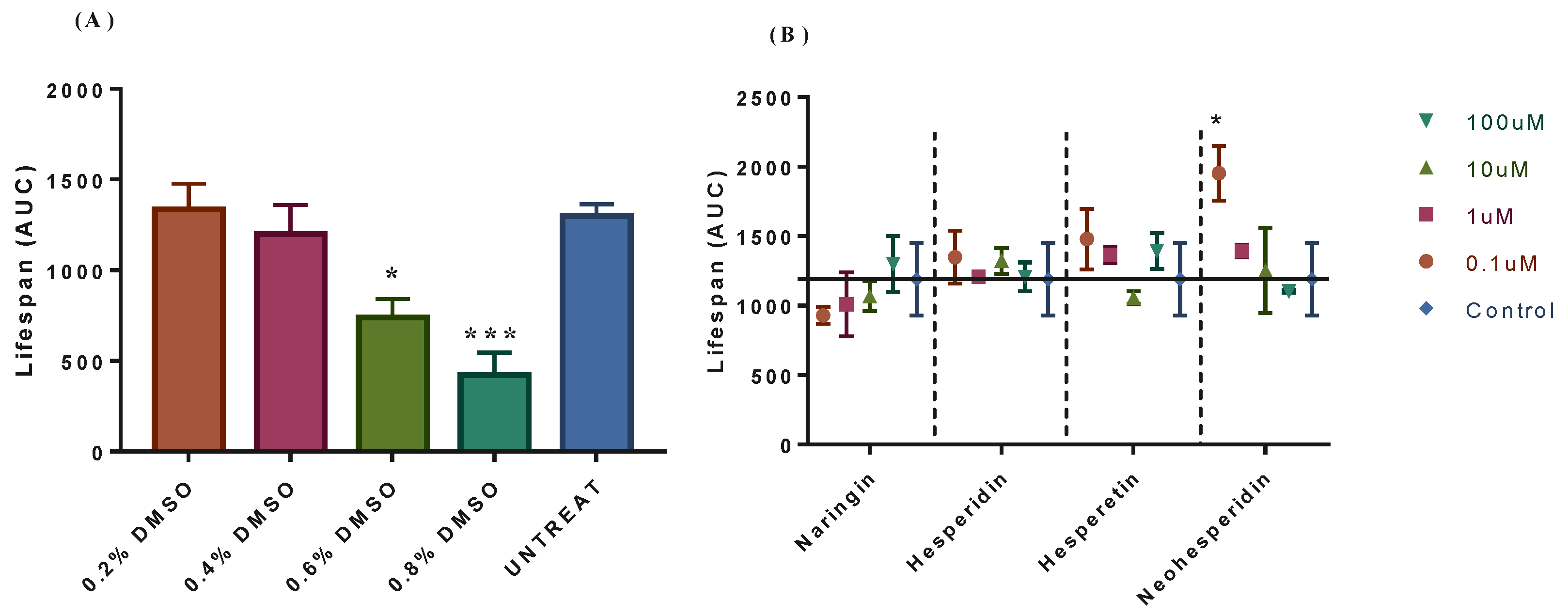
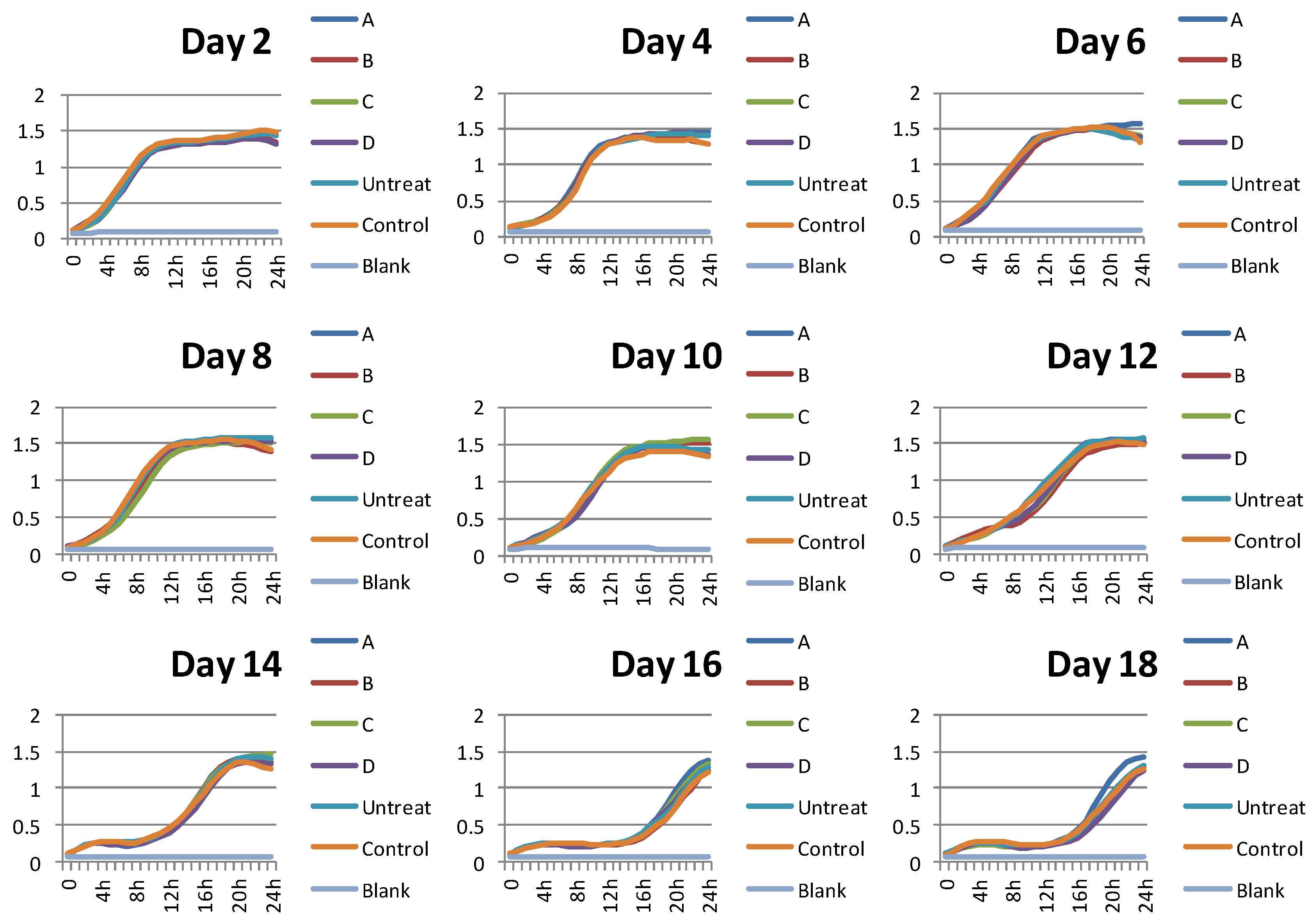
2.1. Neohesperidin Extended Yeast Lifespan in a Concentration-Dependent Manner
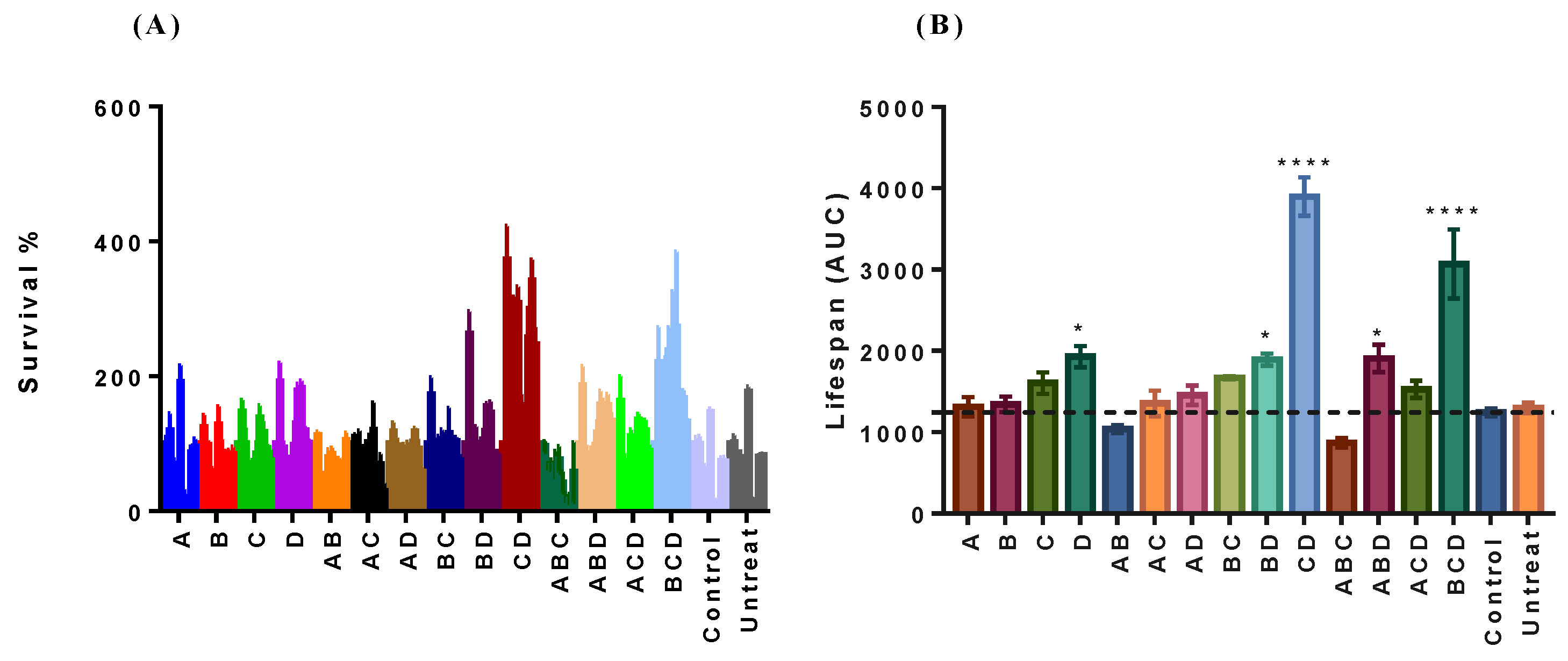
2.2. Neohesperidin Positively Interacted with Hesperetin for Extending the CLS of Yeast BY4742
2.3. Neohesperidin Significantly Reduced Intracellular Reactive Oxygen Species (ROS) Content
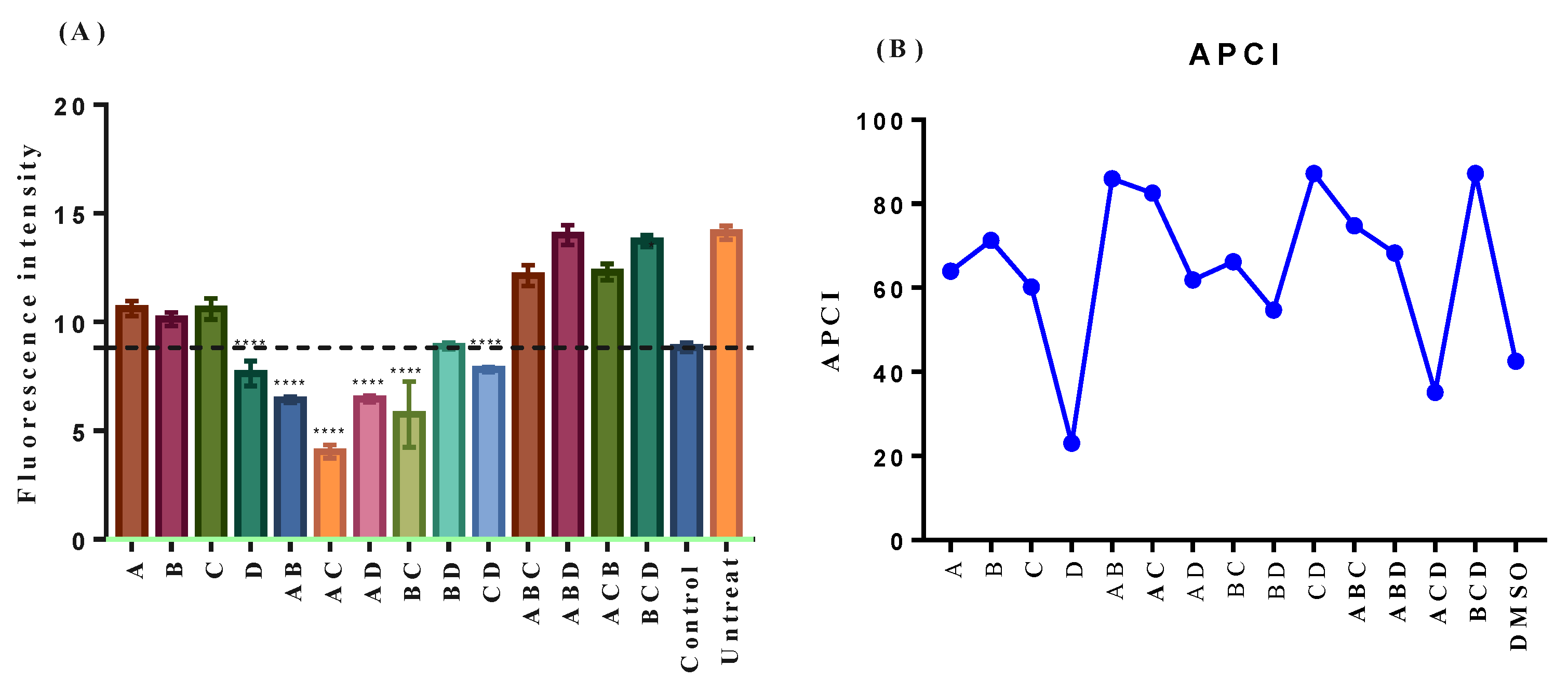
2.4. In Vitro Antioxidant Activity of Neohesperidin was Relatively Weak
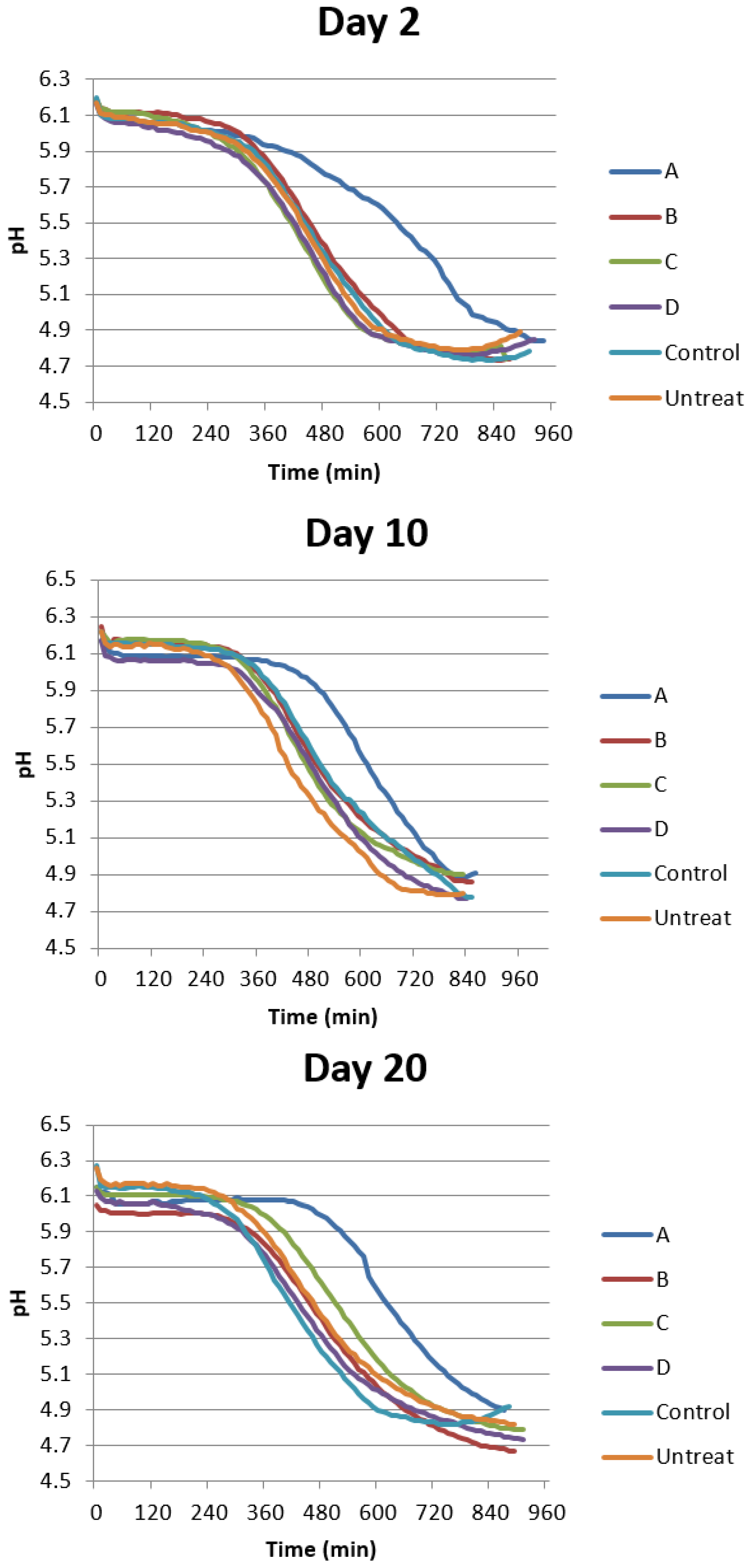
2.5. Neohesperidin Could Not Slow Down the Variation of Extracellular Acidification of Yeast BY4742
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Lifespan and Yeast Cell Growth Assay
4.3. Intracellular ROS Scavenging Ability Assays
4.4. Antioxidant Activity Assays
4.5. Extracellular pH Detection
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- Argyropoulou, A.; Aligiannis, N.; Trougakos, I.P.; Skaltsounis, A.L. Natural compounds with anti-ageing activity. Nat. Prod. Rep. 2013, 30, 1412–1437. [Google Scholar] [CrossRef]
- Ding, A.J.; Zheng, S.Q.; Huang, X.B.; Xing, T.K.; Wu, G.S.; Sun, H.Y.; Qi, S.H.; Luo, H.R. Current perspective in the discovery of anti-aging agents from natural products. Nat. Prod. Biol. 2017, 7, 1–70. [Google Scholar] [CrossRef]
- Pan, M.H.; Lai, C.S.; Tsai, M.L.; Wu, J.C.; Ho, C.T. Molecular mechanisms for anti-aging by natural dietary compounds. Mol. Nutr. Food Res. 2012, 56, 88–115. [Google Scholar] [CrossRef] [PubMed]
- Si, H.; Liu, D. Dietary antiaging phytochemicals and mechanisms associated with prolonged survival. J. Nutr. Biochem. 2014, 25, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Kaeberlein, M. Lessons on longevity from budding yeast. Nature 2010, 464, 513. [Google Scholar] [CrossRef] [PubMed]
- Prasain, J.K.; Carlson, S.H.; Wyss, J.M. Flavonoids and age-related disease, risk; benefits and critical windows. Maturitas 2010, 66, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Longo, V.D.; Shadel, G.S.; Kaeberlein, M.; Kennedy, B. Replicative and Chronological Aging in Saccharomyces cerevisiae. Cell Metab. 2012, 16, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Steinkraus, K.A.; Kaeberlein, M.; Kennedy, B.K. Replicative aging in yeast, the means to the end. Annu. Rev. Cell Dev. Biol. 2008, 24, 29. [Google Scholar] [CrossRef]
- Burtner, C.R.; Murakami, C.J.; Kennedy, B.K.; Kaeberlein, M. A molecular mechanism of chronological aging in yeast. Cell Cycle 2009, 8, 1256–1270. [Google Scholar] [CrossRef]
- Wagner, H. Synergy research, approaching a new generation of phytopharmaceuticals. Fitoterapia 2011, 82, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Benavente-Garcia, O.; Castillo, J. Update on uses and properties of citrus flavonoids, new findings in anticancer; cardiovascular; and anti-inflammatory activity. J. Agric. Food chem. 2008, 56, 6185–6205. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.J.; Ahn, W.S. Neuroprotective effects of chronic hesperetin administration in mice. Arch. Pharm. Res. 2008, 31, 1457. [Google Scholar] [CrossRef] [PubMed]
- Elavarasan, J.; Velusamy, P.; Ganesan, T.; Ramakrishnan, S.K.; Rajasekaran, D.; Periandavan, K. Hesperidin-mediated expression of Nrf2 and upregulation of antioxidant status in senescent rat heart. J. Pharm. Pharmacol. 2012, 64, 1472–1482. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.L.; Poon, C.Y.; Lin, C.Y.; Yan, T.; Kwong, D.W.; Yung, K.K.; Wong, M.S.; Bian, Z.X.; Li, H.W. Inhibition of β-amyloid aggregation by albiflorin; aloeemodin and neohesperidin and their neuroprotective effect on primary hippocampal cells against β-amyloid induced toxicity. Curr. Alzheimer Res. 2015, 12, 424–433. [Google Scholar] [CrossRef]
- Hwang, S.L.; Yen, G.C. Neuroprotective effects of the citrus flavanones against H2O2-induced cytotoxicity in PC12 cells. J. Agric. Food Chem. 2008, 56, 859–864. [Google Scholar] [CrossRef]
- Kandhare, A.D.; Raygude, K.S.; Ghosh, P.; Ghule, A.E.; Bodhankar, S.L. Neuroprotective effect of naringin by modulation of endogenous biomarkers in streptozotocin induced painful diabetic neuropathy. Fitoterapia 2012, 83, 650–659. [Google Scholar] [CrossRef]
- Kumar, A.; Dogra, S.; Prakash, A. Protective effect of naringin; a citrus flavonoid; against colchicine-induced cognitive dysfunction and oxidative damage in rats. J. Med. Food 2010, 13, 976–984. [Google Scholar] [CrossRef]
- Parhiz, H.; Roohbakhsh, A.; Soltani, F.; Rezaee, R.; Iranshahi, M. Antioxidant and anti-inflammatory properties of the citrus flavonoids hesperidin and hesperetin, an updated review of their molecular mechanisms and experimental models. Phytother. Res. 2015, 29, 323–331. [Google Scholar] [CrossRef]
- Sun, X.L.; Ishihara, S.; Matsuura, A.; Sakagami, Y.; Qi, J. Anti-aging effects of hesperidin on Saccharomyces cerevisiae via inhibition of reactive oxygen species and UTH1 gene expression. Biosci. Biotechnol. Biochem. 2012, 76, 640–645. [Google Scholar] [CrossRef]
- Xu, F.; Zang, J.; Chen, D.; Zhang, T.; Zhan, H.Y.; Lu, M.D.; Zhuge, H.X. Neohesperidin induces cellular apoptosis in human breast adenocarcinoma MDA-MB-231 cells via activating the Bcl-2/Bax-mediated signaling pathway. Nat. Prod. Commun. 2012, 7, 1475. [Google Scholar] [CrossRef] [PubMed]
- Murakami, C.; Delaney, J.R.; Chou, A.; Carr, D.; Schleit, J.; Sutphin, G.L.; An, E.H.; Castanza, A.S.; Fletcher, M.; Goswami, S. pH neutralization protects against reduction in replicative lifespan following chronological aging in yeast. Cell Cycle 2012, 11, 3087–3096. [Google Scholar] [CrossRef] [PubMed]
- Hazen, K.C. Influence of DMSO on antifungal activity during susceptibility testing in vitro. Diagn. Microbiol. Infect. Dis. 2013, 75, 60–63. [Google Scholar] [CrossRef]
- Murakami, C.J.; Burtner, C.R.; Kennedy, B.K.; Kaeberlein, M. A method for high-throughput quantitative analysis of yeast chronological life span. J. Gerontol. 2008, 63, 113. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Song, L.; Liu, S.Q.; Huang, D. A high throughput screening assay for determination of chronological lifespan of yeast. Exp. Gerontol. 2011, 46, 915–922. [Google Scholar] [CrossRef]
- Prabhakar, P.K.; Kumar, A.; Doble, M. Combination therapy: A new strategy to manage diabetes and its complications. Phytomedicine Int. J. Phytother. Phytopharm. 2014, 21, 123. [Google Scholar] [CrossRef]
- Liu, R.H. Potential synergy of phytochemicals in cancer prevention: Mechanism of action. J. Nutr. 2004, 134, 3479S–3485S. [Google Scholar] [CrossRef]
- Shen, J.Z.; Tlj, N.; Ho, W.S. Therapeutic potential of phytochemicals in combination with drugs for cardiovascular disorders. Curr. Pharm. Des. 2017, 23, 961–966. [Google Scholar] [CrossRef]
- Rayalam, S.; Della-Fera, M.A.; Baile, C.A. Synergism between resveratrol and other phytochemicals: Implications for obesity and osteoporosis. Mol. Nutr. Food Res. 2011, 55, 1177–1185. [Google Scholar] [CrossRef]
- Harman, D. Aging: A theory based on free radical and radiation chemistry. J. Gerontol. 1956, 11, 298. [Google Scholar] [CrossRef]
- Moradas-Ferreira, P.; Costa, V.; Piper, P.; Mager, W. The molecular defences against reactive oxygen species in yeast. Mol. Microbiol. 1996, 19, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.E.; Allen, P.; Brunton, N.; O’Grady, M.N.; Kerry, J.P. Phenolic composition and in vitro antioxidant capacity of four commercial phytochemical products: Olive leaf extract (Olea europaea L.), lutein, sesamol and ellagic acid. Food Chem. 2011, 126, 948–955. [Google Scholar] [CrossRef]
- Piper, P.W. Maximising the yeast chronological lifespan. Sub-Cell. Biochem. 2012, 57, 145. [Google Scholar]
- Burhans, W.C.; Weinberger, M. Acetic acid effects on aging in budding yeast: Are they relevant to aging in higher eukaryotes? Cell Cycle 2009, 8, 2300–2302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.L.; Shih, P.H.; Yen, G.C. Neuroprotective effects of citrus flavonoids. J. Agric. Food Chem. 2012, 60, 877. [Google Scholar] [CrossRef]
- Craker, L.E.; Cookson, C.; Osborne, D.J. Control of proton extrusion and cell elongation by ethylene and auxin in the water-plant Ranunculus sceleratus. Plant. Sci. Lett. 1978, 12, 379–385. [Google Scholar] [CrossRef]
- Areias, F.M.; Rego, A.C.; Oliveira, C.R.; Seabra, R.M. Antioxidant effect of flavonoids after ascorbate/Fe2+-induced oxidative stress in cultured retinal cells. Biochem. Pharmacol. 2001, 62, 111–118. [Google Scholar] [CrossRef]
- Qi, W.; Zhu, C.; Fan, X.; Yang, D.; Yang, M. Research progress on the correlation between antioxidants and life extension. Chin. J. Pharmacol. Toxicol. 2016, 3, 588–597. (In Chinese) [Google Scholar]
- Kumar, S.; Lefevre, S.D.; Veenhuis, M.; Klei, I.J.V.D. Extension of yeast chronological lifespan by methylamine. PLoS ONE 2012, 7, e48982. [Google Scholar] [CrossRef] [Green Version]
- Murakami, C.J.; Wall, V.; Basisty, N.; Kaeberlein, M. Composition and acidification of the culture medium influences chronological aging similarly in vineyard and laboratory yeast. PLoS ONE 2011, 6, e24530. [Google Scholar] [CrossRef] [Green Version]
- Morgunova, G.V.; Klebanov, A.A.; Marotta, F.; Khokhlov, A.N. Culture medium pH and stationary phase/chronological aging of different cells. Mosc. Univ. Biol. Sci. Bull. 2017, 72, 58–62. [Google Scholar] [CrossRef]
- Gilvarg, C.; Bloch, K. The utilization of acetic acid for amino acid synthesis in yeast. J. Biol. Chem. 1951, 193, 339. [Google Scholar] [PubMed]
- Yang, Y.; Zhao, X.J.; Pan, Y.; Zhou, Z. Identification of the chemical compositions of Ponkan peel by ultra performance liquid chromatography coupled with quadrupole time-of-flight mass spectrometry. Anal. Methods 2015, 8, 893–903. [Google Scholar] [CrossRef]
- Cypser, J.R.; Johnson, T.E. Multiple stressors in caenorhabditis elegans induce stress hormesis and extended longevity. J. Gerontol. Ser. A 2002, 57, B109–B114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludovico, P.; Burhans, W.C. Reactive oxygen species, ageing and the hormesis police. FEMS Yeast Res. 2014, 14, 33–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Y. Mitochondria, reactive oxygen species, and chronological aging: A message from yeast. Exp. Gerontol. 2011, 46, 847–852. [Google Scholar] [CrossRef]
- Mesquita, A.; Weinberger, M.; Silva, A.; Sampaiomarques, B.; Almeida, B.; Leão, C.; Costa, V.; Rodrigues, F.; Burhans, W.C.; Ludovico, P. Caloric restriction or catalase inactivation extends yeast chronological lifespan by inducing H2O2 and superoxide dismutase activity. Proc. Natl. Acad. Sci. USA 2010, 107, 15123–15128. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Schroeder, E.A.; Ocampo, A.; Barrientos, A.; Shadel, G.S. Regulation of yeast chronological life span by TORC1 via adaptive mitochondrial ROS signaling. Cell Metab. 2011, 13, 668–678. [Google Scholar] [CrossRef] [Green Version]
- Graziano, A.C.; Cardile, V.; Crascì, L.; Caggia, S.; Dugo, P.; Bonina, F.; Panico, A. Protective effects of an extract from Citrus bergamia against inflammatory injury in interferon-γ and histamine exposed human keratinocytes. Life Sci. 2012, 90, 968–974. [Google Scholar] [CrossRef]
- Nohara, K.; Mallampalli, V.; Nemkov, T.; Wirianto, M.; Yang, J.; Ye, Y.; Sun, Y.; Han, L.; Esser, K.A.; Mileykovskaya, E.; et al. Nobiletin fortifies mitochondrial respiration in skeletal muscle to promote healthy aging against metabolic challenge. Nat. Commun. 2019, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Song, L.; Liu, S.Q.; Huang, D. Tanshinones extend chronological lifespan in budding yeast Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2014, 98, 8617–8628. [Google Scholar] [CrossRef] [PubMed]
- Lutchman, V.; Medkour, Y.; Samson, E.; Arlia-Ciommo, A.; Dakik, P.; Cortes, B.; Rukundo, B. Discovery of plant extracts that greatly delay yeast chronological aging and have different effects on longevity-defining cellular processes. Oncotarget 2016, 7, 16542–16566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreca, D.; Bellocco, E.; Caristi, C.; Leuzzi, U.; Gattuso, G. Kumquat (Fortunella japonica Swingle) juice: Flavonoid distribution and antioxidant properties. Food Res. Int. 2011, 44, 2190–2197. [Google Scholar] [CrossRef]
- Hungder, J.; Chang, K.S.; Chang, T.C.; Chuanliang, H. Antioxidant potentials of buntan pumelo (Citrus grandis Osbeck) and its ethanolic and acetified fermentation products. Food Chem. 2010, 118, 554–558. [Google Scholar]
- Almeida, M.M.B.; de Sousa, P.H.M.; Arriaga, Â.M.C.; do Prado, G.M.; de Carvalho Magalhães, C.E.; Maia, G.A.; de Lemos, T.L.G. Bioactive compounds and antioxidant activity of fresh exotic fruits from northeastern Brazil. Food Res. Int. 2011, 44, 2155–2159. [Google Scholar] [CrossRef] [Green Version]
- Park, S.J.; Ahmad, F.; Philp, A.; Baar, K.; Williams, T.; Luo, H.; Ke, H.; Rehmann, H.; Taussig, R.; Brown, A.L. Resveratrol ameliorates aging-related metabolic phenotypes by inhibiting cAMP phosphodiesterases. Cell 2012, 148, 421–433. [Google Scholar] [CrossRef] [Green Version]
- Bottino, D.A.; Lopes, F.G.; Oliveira, F.J.D.; Mecenas, A.D.S.; Clapauch, R.; Bouskela, E. Relationship between biomarkers of inflammation, oxidative stress and endothelial/microcirculatory function in successful aging versus healthy youth: A transversal study. BMC Geriatr. 2015, 15, 41. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Samples are not available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Equation | R2 | The Linear Range |
|---|---|---|---|
| DPPH | y = −0.0029x + 0.6716 | 0.9997 | 0–200 µM |
| ABTS | y = −0.0006x + 0.644 | 0.9962 | 0–1000 µM |
| FRAP | y = 0.0023x − 0.003 | 0.9992 | 0–560 µM |
| Phytochemicals | DPPH | ABTS | FRAP | APCI | Rank |
|---|---|---|---|---|---|
| A | 11.33 ± 0.15 j | 550.18 ± 1.21 a | 350.74 ± 1.21 b | 63.96 | 9 |
| B | 33.31 ± 1.16 h | 533.57 ± 0.2 a | 302.64 ± 0.82 e | 71.32 | 6 |
| C | 41.39 ± 0.16 f | 447.33 ± 0.59 b | 160.01 ± 0.29 h | 60.17 | 10 |
| D | 28.74 ± 0.17 i | 119.33 ± 0.36 c | 7.54 ± 0.21 i | 23.02 | 15 |
| AB | 35.77 ± 0.06 g | 571.97 ± 7.13 a | 453.54 ± 0.86 a | 85.97 | 3 |
| AC | 45.8 ± 0.24 e | 548.94 ± 1.99 a | 350.96 ± 0.46 b | 82.51 | 4 |
| AD | 34.13 ± 0.10 h | 500.91 ± 1.03 b | 194.44 ± 0.62 g | 61.90 | 10 |
| BC | 53.74 ± 0.23 c | 432.93 ± 0.6 b | 162.59 ± 0.71 h | 66.19 | 8 |
| BD | 52.61 ± 0.05 c | 444.8 ± 27.13 b | 5.52 ± 0.22 i | 54.73 | 12 |
| CD | 58.52 ± 0.06 b | 546.17 ± 1.01 a | 323.97 ± 0.04 c | 87.23 | 1 |
| ABC | 58.61 ± 0.10 b | 477.95 ± 3.37 b | 208.58 ± 1.19 f | 74.82 | 5 |
| ABD | 47.58 ± 0.12 d | 489.66 ± 0.64 b | 191.63 ± 1.50 g | 68.30 | 7 |
| ACD | 61.75 ± 0.30 a | 25.05 ± 0.74 d | 4.03 ± 0.08 i | 35.09 | 14 |
| BCD | 61.23 ± 0.11 a | 538.31 ± 0.24 a | 310.05 ± 0.52 d | 87.21 | 2 |
| DMSO | 3.42 ± 0.18 k | 455.56 ± 0.54 b | 192.37 ± 0.26 g | 42.53 | 13 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, C.; Zhang, H.; Guan, X.; Zhou, Z. The Anti-Aging Potential of Neohesperidin and Its Synergistic Effects with Other Citrus Flavonoids in Extending Chronological Lifespan of Saccharomyces Cerevisiae BY4742. Molecules 2019, 24, 4093. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24224093
Guo C, Zhang H, Guan X, Zhou Z. The Anti-Aging Potential of Neohesperidin and Its Synergistic Effects with Other Citrus Flavonoids in Extending Chronological Lifespan of Saccharomyces Cerevisiae BY4742. Molecules. 2019; 24(22):4093. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24224093
Chicago/Turabian StyleGuo, Chunxia, Hua Zhang, Xin Guan, and Zhiqin Zhou. 2019. "The Anti-Aging Potential of Neohesperidin and Its Synergistic Effects with Other Citrus Flavonoids in Extending Chronological Lifespan of Saccharomyces Cerevisiae BY4742" Molecules 24, no. 22: 4093. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24224093