
The Neuroprotective Effects of Histamine H3 Receptor Antagonist E177 on Pilocarpine-Induced Status Epilepticus in Rats

 , , ,
, , , 
Abstract
:1. Introduction
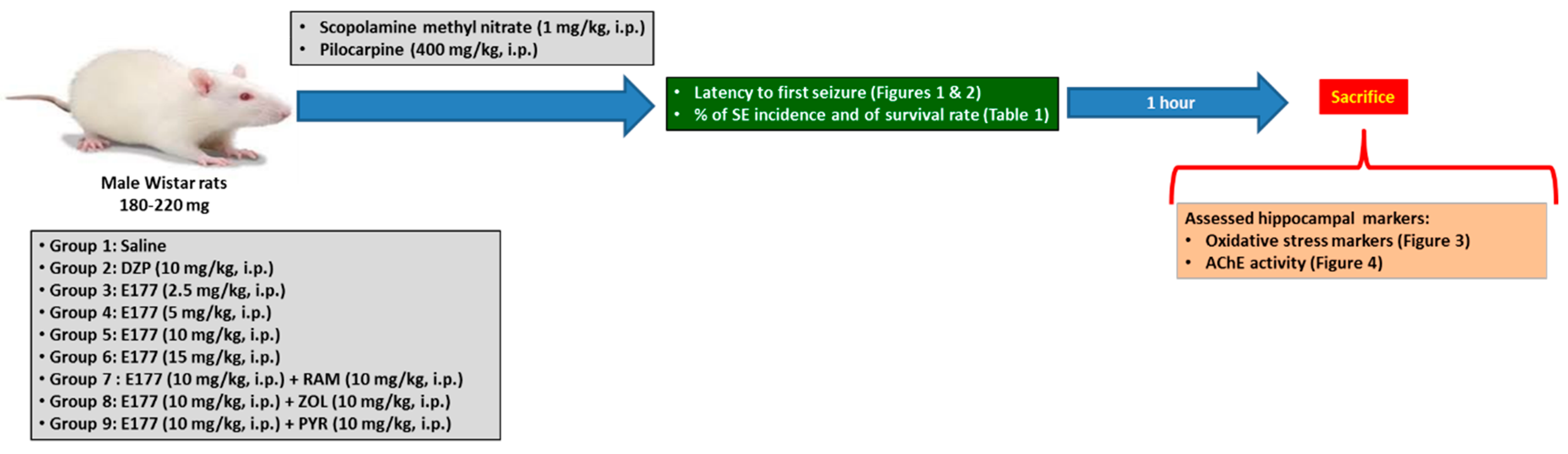
2. Results
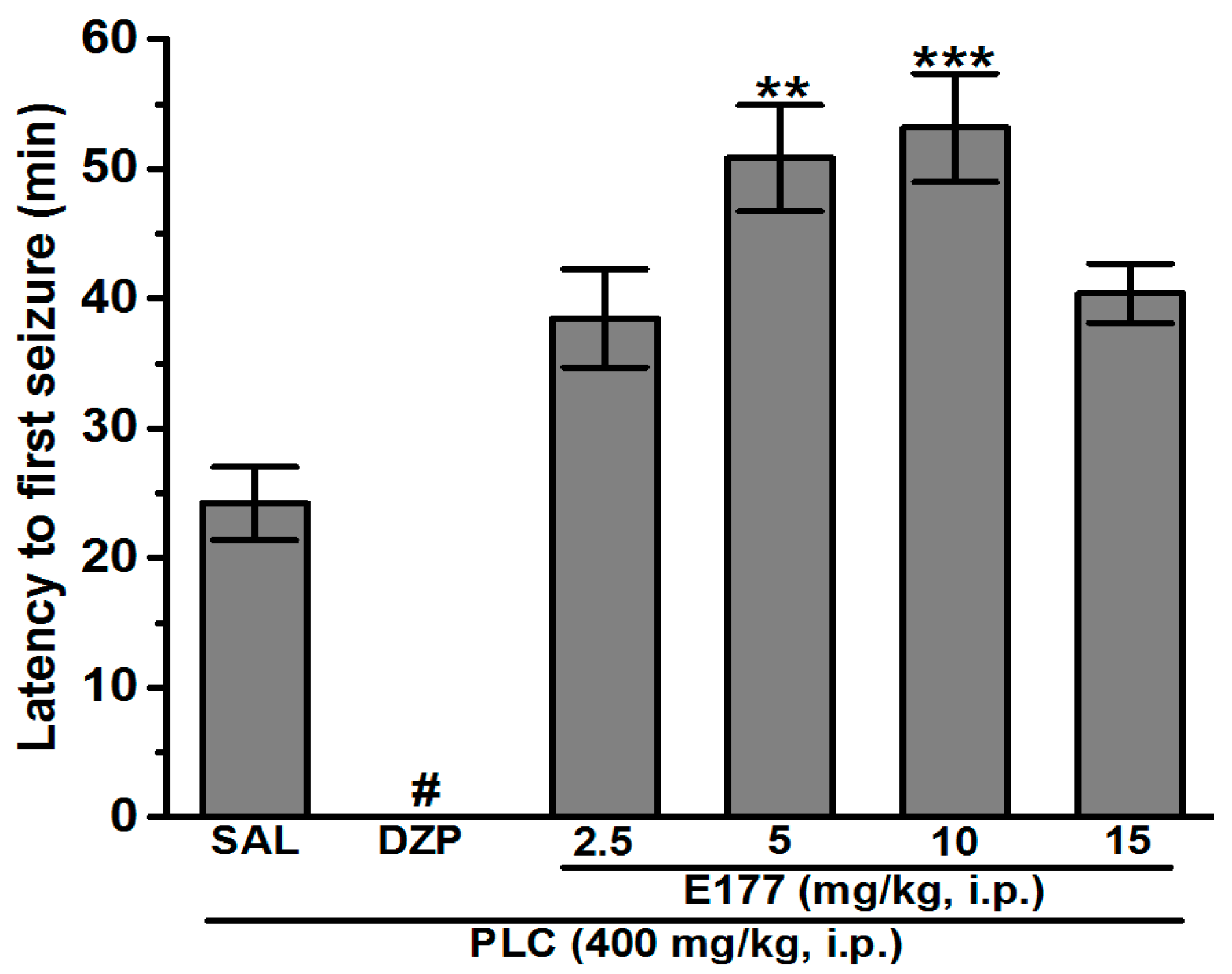
2.1. E177 Prolonged Latency Time to First Seizure
2.2. Effect of E177 on SE Incidence and Survival Rate
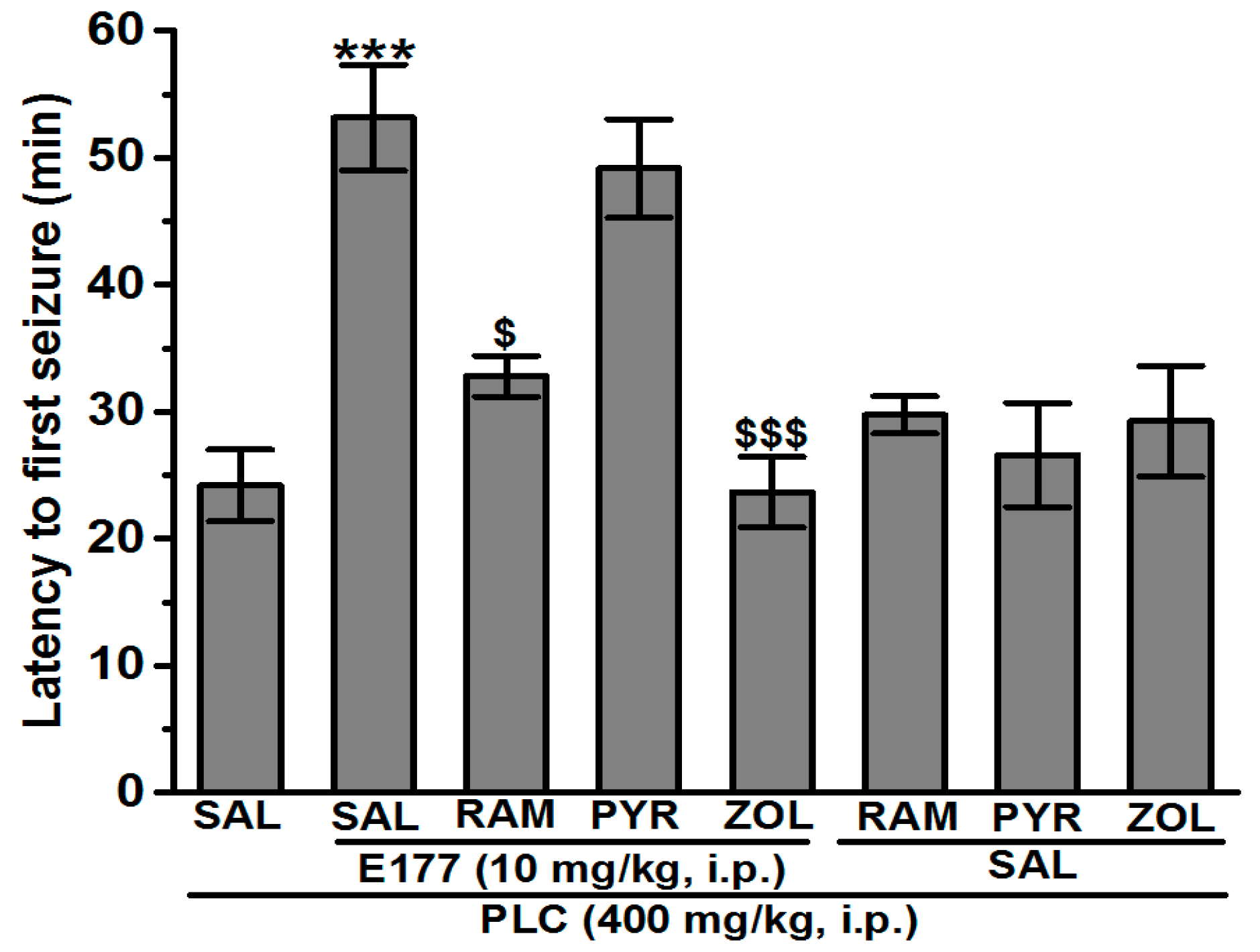
2.3. Effects of RAM, PYR, and ZOL on E177-Provided Protection against SE
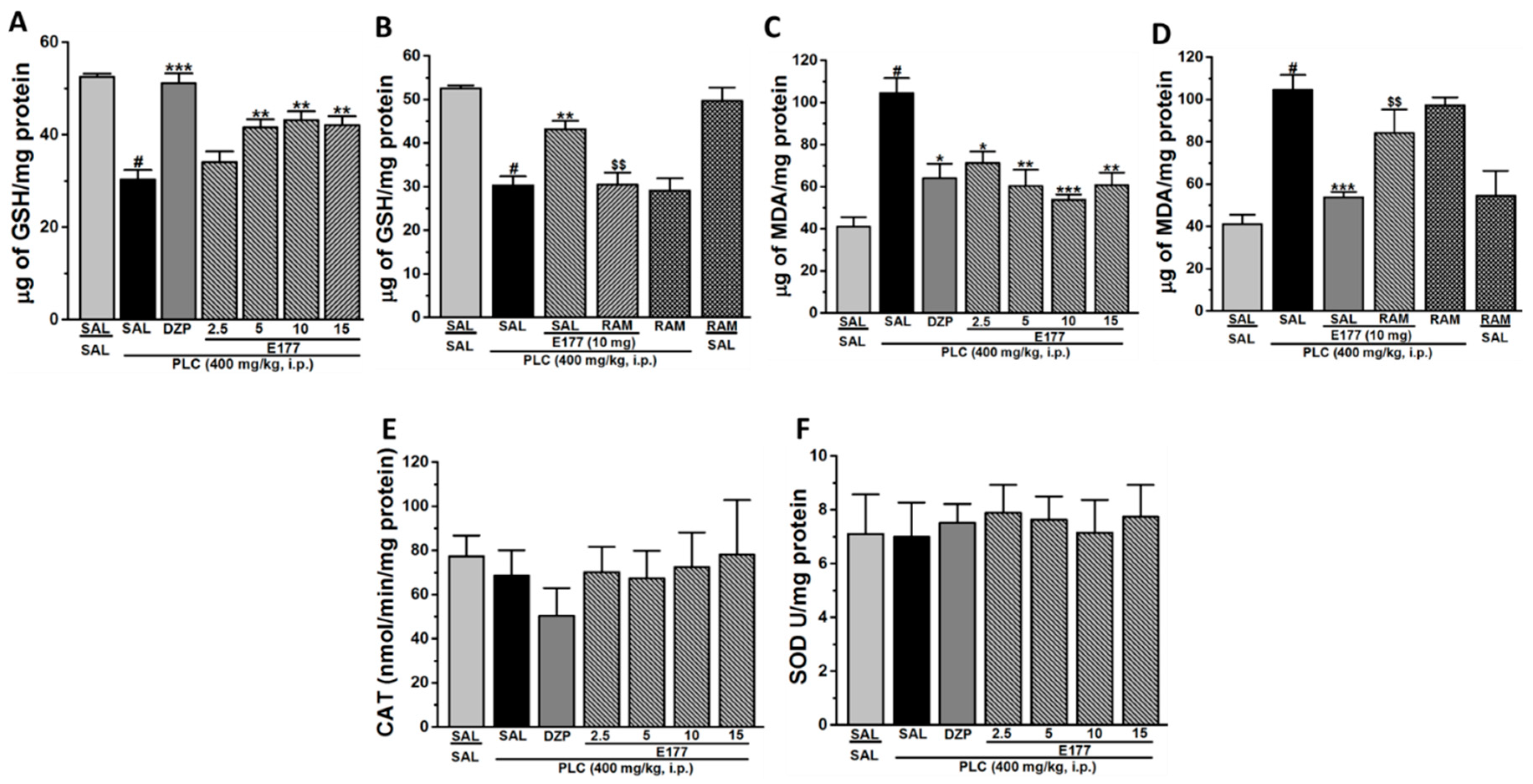
2.4. Effects of E177 on Oxidative Stress Markers
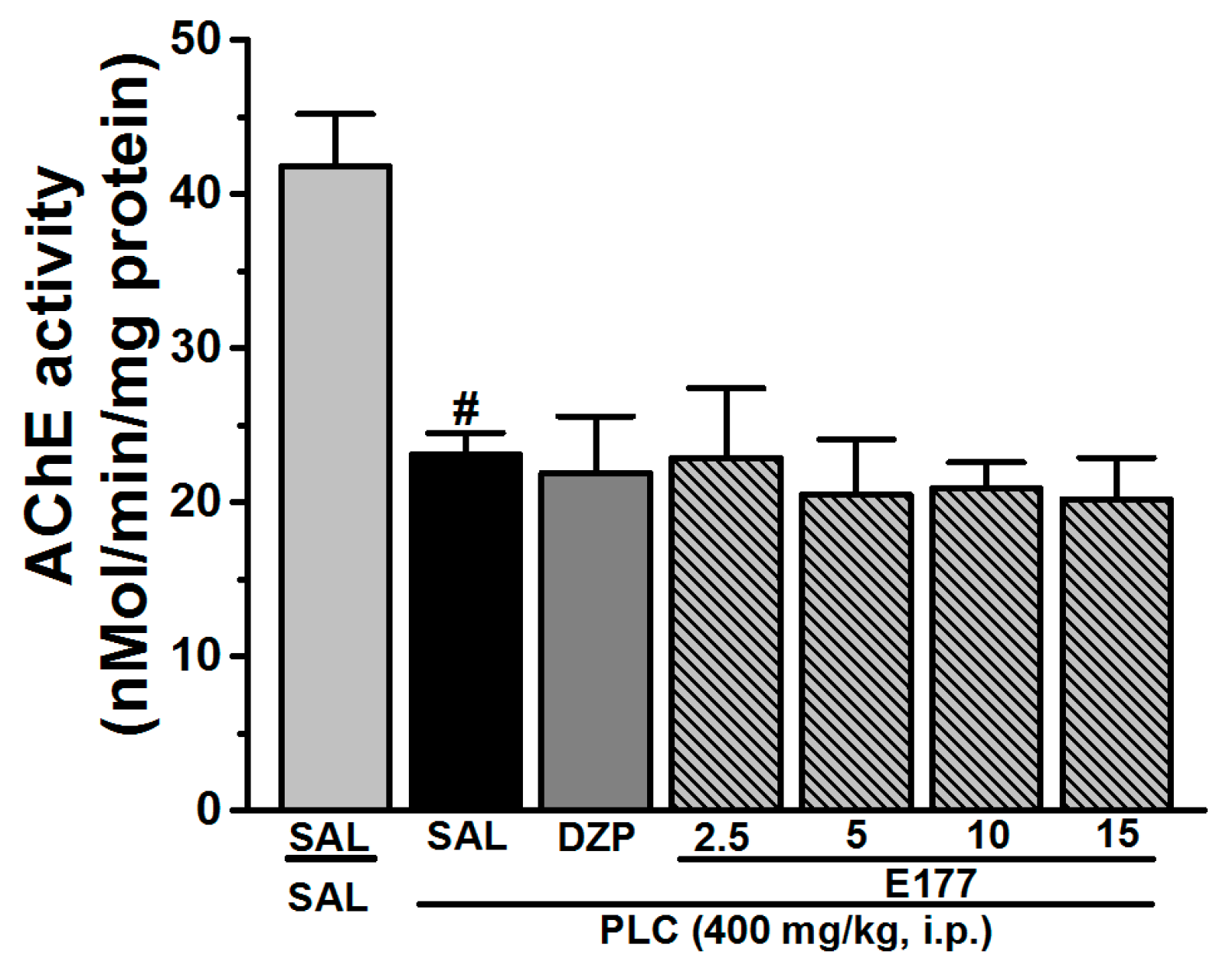
2.5. Effects of E177 on AChE Activity
3. Discussion
4. Material and Methods
4.1. Animals
4.2. Drugs
4.3. Experimental Procedure of PLC-Induced SE
4.4. Biochemical Assessments
4.4.1. Oxidative Stress
4.4.1.1. MDA
4.4.1.2. GSH
4.4.1.3. CAT
4.4.1.4. SOD
4.4.1.5. Acetylcholine Esterase (AChE) Activity
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Loscher, W.; Klitgaard, H.; Twyman, R.E.; Schmidt, D. New avenues for anti-epileptic drug discovery and development. Nat. Rev. Drug. Discov. 2013, 12, 757–776. [Google Scholar] [CrossRef] [PubMed]
- Kirsten, M.F.; Khara, M.S.; Samuel, W.; Scott, B.P.; Churl-Su, K.; Jonathan, D.; Tamara, P.; Diane, L.L.; Nathalie, J. Prevalence and incidence of epilepsy: A systematic review and meta-analysis of international studies. Neurology 2017, 89, 642. [Google Scholar]
- Treiman, D.M. Electroclinical features of status epilepticus. J. Clin. Neurophysiol. 1995, 12, 343–362. [Google Scholar] [CrossRef] [PubMed]
- Rossetti, A.O. Treatment options in the management of status epilepticus. Curr. Treat. Options. Neurol. 2010, 12, 100–112. [Google Scholar] [CrossRef]
- Theodore, W.H.; Porter, R.J.; Albert, P.; Kelley, K.; Bromfield, E.; Devinsky, O.; Sato, S. The secondarily generalized tonic-clonic seizure: A videotape analysis. Neurology 1994, 44, 1403–1407. [Google Scholar] [CrossRef]
- Agarwal, N.B.; Agarwal, N.K.; Mediratta, P.K.; Sharma, K.K. Effect of lamotrigine, oxcarbazepine and topiramate on cognitive functions and oxidative stress in PTZ-kindled mice. Seizure 2011, 20, 257–262. [Google Scholar] [CrossRef]
- Sutter, R.; Marsch, S.; Fuhr, P.; Kaplan, P.W.; Ruegg, S. Anesthetic drugs in status epilepticus: Risk or rescue? A 6-year cohort study. Neurology 2014, 82, 656–664. [Google Scholar] [CrossRef]
- Turski, W.A.; Cavalheiro, E.A.; Schwarz, M.; Czuczwar, S.J.; Kleinrok, Z.; Turski, L. Limbic seizures produced by pilocarpine in rats: Behavioural, electroencephalographic and neuropathological study. Behav. Brain. Res. 1983, 9, 315–335. [Google Scholar] [CrossRef]
- Ali, A.E.; Mahdy, H.M.; Elsherbiny, D.M.; Azab, S.S. Rifampicin ameliorates lithium-pilocarpine-induced seizures, consequent hippocampal damage and memory deficit in rats: Impact on oxidative, inflammatory and apoptotic machineries. Biochem. Pharmacol. 2018, 156, 431–443. [Google Scholar] [CrossRef]
- Tellez-Zenteno, J.F.; Hernandez-Ronquillo, L. A review of the epidemiology of temporal lobe epilepsy. Epilepsy. Res. Treat. 2012, 2012, 630853. [Google Scholar] [CrossRef]
- Dos Santos, P.S.; Costa, J.P.; Tome Ada, R.; Saldanha, G.B.; de Souza, G.F.; Feng, D.; de Freitas, R.M. Oxidative stress in rat striatum after pilocarpine-induced seizures is diminished by alpha-tocopherol. Eur. J. Pharmacol. 2011, 668, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Kamei, C. Involvement of central histamine in amygdaloid kindled seizures in rats. Behav. Brain. Res. 2001, 124, 243–250. [Google Scholar] [CrossRef]
- Kamei, C.; Ishizawa, K.; Kakinoki, H.; Fukunaga, M. Histaminergic mechanisms in amygdaloid-kindled seizures in rats. Epilepsy. Res. 1998, 30, 187–194. [Google Scholar] [CrossRef]
- Vohora, D.; Pal, S.N.; Pillai, K.K. Histamine and selective H3-receptor ligands: A possible role in the mechanism and management of epilepsy. Pharmacol. Biochem. Behav. 2001, 68, 735–741. [Google Scholar] [CrossRef]
- Ago, J.; Ishikawa, T.; Matsumoto, N.; Ashequr Rahman, M.; Kamei, C. Mechanism of imipramine-induced seizures in amygdala-kindled rats. Epilepsy. Res. 2006, 72, 1–9. [Google Scholar] [CrossRef]
- Scherkl, R.; Hashem, A.; Frey, H.H. Histamine in brain--its role in regulation of seizure susceptibility. Epilepsy. Res. 1991, 10, 111–118. [Google Scholar] [CrossRef]
- Miyata, I.; Saegusa, H.; Sakurai, M. Seizure-modifying potential of histamine H1 antagonists: A clinical observation. Pediatr. Int. 2011, 53, 706–708. [Google Scholar] [CrossRef]
- Takano, T.; Sakaue, Y.; Sokoda, T.; Sawai, C.; Akabori, S.; Maruo, Y.; Taga, T.; Ohno, M.; Takeuchi, Y. Seizure susceptibility due to antihistamines in febrile seizures. Pediatr. Neurol. 2010, 42, 277–279. [Google Scholar] [CrossRef]
- Zolaly, M.A. Histamine H1 antagonists and clinical characteristics of febrile seizures. Int. J. Gen. Med. 2012, 5, 277–281. [Google Scholar] [CrossRef]
- Gemkow, M.J.; Davenport, A.J.; Harich, S.; Ellenbroek, B.A.; Cesura, A.; Hallett, D. The histamine H3 receptor as a therapeutic drug target for CNS disorders. Drug. Discov. Today. 2009, 14, 509–515. [Google Scholar] [CrossRef]
- Bhowmik, M.; Khanam, R.; Vohora, D. Histamine H3 receptor antagonists in relation to epilepsy and neurodegeneration: A systemic consideration of recent progress and perspectives. Br. J. Pharmacol. 2012, 167, 1398–1414. [Google Scholar] [CrossRef] [PubMed]
- Sadek, B.; Saad, A.; Sadeq, A.; Jalal, F.; Stark, H. Histamine H3 receptor as a potential target for cognitive symptoms in neuropsychiatric diseases. Behav. Brain. Res. 2016, 312, 415–430. [Google Scholar] [CrossRef] [PubMed]
- Sadek, B.; Saad, A.; Latacz, G.; Kuder, K.; Olejarz, A.; Karcz, T.; Stark, H.; Kieć-Kononowicz, K. Non-imidazole-based histamine H3 receptor antagonists with anticonvulsant activity in different seizure models in male adult rats. Drug. Des. Devel. Ther. 2016, 10, 3879–3898. [Google Scholar] [CrossRef] [PubMed]
- Alachkar, A.; Lazewska, D.; Kiec-Kononowicz, K.; Sadek, B. The Histamine H3 Receptor Antagonist E159 Reverses Memory Deficits Induced by Dizocilpine in Passive Avoidance and Novel Object Recognition Paradigm in Rats. Front. Pharmacol. 2017, 8, 709. [Google Scholar] [CrossRef]
- Eissa, N.; Jayaprakash, P.; Azimullah, S.; Ojha, S.K.; Al-Houqani, M.; Jalal, F.Y.; Lazewska, D.; Kiec-Kononowicz, K.; Sadek, B. The histamine H3R antagonist DL77 attenuates autistic behaviors in a prenatal valproic acid-induced mouse model of autism. Sci. Rep. 2018, 8, 13077. [Google Scholar] [CrossRef]
- Harada, C.; Hirai, T.; Fujii, Y.; Harusawa, S.; Kurihara, T.; Kamei, C. Intracerebroventricular administration of histamine H3 receptor antagonists decreases seizures in rat models of epilepsia. Methods. Find. Exp. Clin Pharmacol. 2004, 26, 263–270. [Google Scholar]
- Witkin, J.M.; Nelson, D.L. Selective histamine H3 receptor antagonists for treatment of cognitive deficiencies and other disorders of the central nervous system. Pharmacol. Ther. 2004, 103, 1–20. [Google Scholar] [CrossRef]
- Alachkar, A.; Lazewska, D.; Latacz, G.; Frank, A.; Siwek, A.; Lubelska, A.; Honkisz-Orzechowska, E.; Handzlik, J.; Stark, H.; Kiec-Kononowicz, K.; et al. Studies on Anticonvulsant Effects of Novel Histamine H3R Antagonists in Electrically and Chemically Induced Seizures in Rats. Int. J. Mol. Sci. 2018, 19, 3386. [Google Scholar] [CrossRef]
- Arrang, J.M.; Garbarg, M.; Schwartz, J.C. Auto-inhibition of brain histamine release mediated by a novel class (H3) of histamine receptor. Nature 1983, 302, 832–837. [Google Scholar] [CrossRef]
- Sadek, B.; Saad, A.; Subramanian, D.; Shafiullah, M.; Łażewska, D.; Kieć-Kononowiczc, K. Anticonvulsant and procognitive properties of the non-imidazole histamine H3 receptor antagonist DL77 in male adult rats. Neuropharmacology 2016, 106, 46–55. [Google Scholar] [CrossRef]
- Panula, P.; Chazot, P.L.; Cowart, M.; Gutzmer, R.; Leurs, R.; Liu, W.L.; Stark, H.; Thurmond, R.L.; Haas, H.L. International Union of Basic and Clinical Pharmacology. XCVIII. Histamine Receptors. Pharmacol. Rev. 2015, 67, 601–655. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.C.; Frei, B. Toward a new recommended dietary allowance for vitamin C based on antioxidant and health effects in humans. Am. J. Clin. Nutr. 1999, 69, 1086–1107. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.A.; Hudes, E.S.; Tice, J.A. Relation of serum ascorbic acid to mortality among US adults. J. Am. Coll. Nutr. 2001, 20, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.F.; Freitas, R.L.; Xavier, S.M.; Saldanha, G.B.; Freitas, R.M. Neuroprotective actions of vitamin C related to decreased lipid peroxidation and increased catalase activity in adult rats after pilocarpine-induced seizures. Pharmacol. Biochem. Behav. 2008, 89, 1–5. [Google Scholar] [CrossRef]
- Xue, Y.; Xie, N.; Cao, L.; Zhao, X.; Jiang, H.; Chi, Z. Diazoxide preconditioning against seizure-induced oxidative injury is via the PI3K/Akt pathway in epileptic rat. Neurosci. Lett. 2011, 495, 130–134. [Google Scholar] [CrossRef]
- Lazewska, D.; Kaleta, M.; Hagenow, S.; Mogilski, S.; Latacz, G.; Karcz, T.; Lubelska, A.; Honkisz, E.; Handzlik, J.; Reiner, D.; et al. Novel naphthyloxy derivatives — Potent histamine H3 receptor ligands. Synthesis and pharmacological evaluation. Bioorg. Med. Chem. 2018, 26, 2573–2585. [Google Scholar] [CrossRef]
- Alachkar, A.; Khan, N.; Lazewska, D.; Kiec-Kononowicz, K.; Sadek, B. Histamine H3 receptor antagonist E177 attenuates amnesia induced by dizocilpine without modulation of anxiety-like behaviors in rats. Neuropsychiatr. Dis. Treat. 2019, 15, 531–542. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, H.; Onodera, K.; Iinuma, K.; Watanabe, T. Effect of thioperamide, a histamine H3 receptor antagonist, on electrically induced convulsions in mice. Eur J. Pharmacol 1993, 234, 129–133. [Google Scholar] [CrossRef]
- Alachkar, A.; Latacz, G.; Siwek, A.; Lubelska, A.; Honkisz, E.; Grybos, A.; Lazewska, D.; Handzlik, J.; Stark, H.; Kiec-Kononowicz, K.; et al. Anticonvulsant evaluation of novel non-imidazole histamine H3R antagonists in different convulsion models in rats. Pharmacol. Biochem. Behav. 2018, 170, 14–24. [Google Scholar] [CrossRef]
- Walker, M.C. Pathophysiology of status epilepticus. Neurosci. Lett. 2018, 667, 84–91. [Google Scholar] [CrossRef]
- Deng, X.; Wang, M.; Hu, S.; Feng, Y.; Shao, Y.; Xie, Y.; Wu, M.; Chen, Y.; Shi, X. The Neuroprotective Effect of Astaxanthin on Pilocarpine-Induced Status Epilepticus in Rats. Front. Cell. Neurosci. 2019, 13, 123. [Google Scholar] [CrossRef] [PubMed]
- Sadek, B.; Kuder, K.; Subramanian, D.; Shafiullah, M.; Stark, H.; Lazewska, D.; Adem, A.; Kiec-Kononowicz, K. Anticonvulsive effect of nonimidazole histamine H3 receptor antagonists. Behav. Pharmacol. 2014, 25, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Sadek, B.; Saad, A.; Schwed, J.S.; Weizel, L.; Walter, M.; Stark, H. Anticonvulsant effects of isomeric nonimidazole histamine H3 receptor antagonists. Drug. Des. Devel. Ther. 2016, 10, 3633–3651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadek, B.; Schwed, J.S.; Subramanian, D.; Weizel, L.; Walter, M.; Adem, A.; Stark, H. Non-imidazole histamine H3 receptor ligands incorporating antiepileptic moieties. Eur. J. Med. Chem. 2014, 77, 269–279. [Google Scholar] [CrossRef]
- Sadek, B.; Shehab, S.; Wiecek, M.; Subramanian, D.; Shafiullah, M.; Kiec-Kononowicz, K.; Adem, A. Anticonvulsant properties of histamine H3 receptor ligands belonging to N-substituted carbamates of imidazopropanol. Bioorg. Med. Chem. Lett. 2013, 23, 4886–4891. [Google Scholar] [CrossRef]
- Kasteleijn-Nolst Trenite, D.; Parain, D.; Genton, P.; Masnou, P.; Schwartz, J.C.; Hirsch, E. Efficacy of the histamine 3 receptor (H3R) antagonist pitolisant (formerly known as tiprolisant; BF2.649) in epilepsy: Dose-dependent effects in the human photosensitivity model. Epilepsy. Behav. 2013, 28, 66–70. [Google Scholar] [CrossRef]
- Vohora, D.; Pal, S.N.; Pillai, K.K. Thioperamide, a selective histamine H3 receptor antagonist, protects against PTZ-induced seizures in mice. Life. Sci. 2000, 66, l297–l301. [Google Scholar] [CrossRef]
- Dong, Y.; Wang, S.; Zhang, T.; Zhao, X.; Liu, X.; Cao, L.; Chi, Z. Ascorbic acid ameliorates seizures and brain damage in rats through inhibiting autophagy. Brain. Res. 2013, 1535, 115–123. [Google Scholar] [CrossRef]
- Minjarez, B.; Camarena, H.O.; Haramati, J.; Rodriguez-Yanez, Y.; Mena-Munguia, S.; Buritica, J.; Garcia-Leal, O. Behavioral changes in models of chemoconvulsant-induced epilepsy: A review. Neurosci. Biobehav. Rev. 2017, 83, 373–380. [Google Scholar] [CrossRef]
- Turski, L.; Ikonomidou, C.; Turski, W.A.; Bortolotto, Z.A.; Cavalheiro, E.A. Review: Cholinergic mechanisms and epileptogenesis. The seizures induced by pilocarpine: A novel experimental model of intractable epilepsy. Synapse 1989, 3, 154–171. [Google Scholar] [CrossRef]
- Song, S.; Fajol, A.; Chen, Y.; Ren, B.; Shi, S. Anticonvulsive effects of protodioscin against pilocarpine-induced epilepsy. Eur. J. Pharmacol. 2018, 833, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Benetti, F.; Izquierdo, I. Histamine infused into basolateral amygdala enhances memory consolidation of inhibitory avoidance. Int. J. Neuropsychopharmacol. 2013, 16, 1539–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benetti, F.; Baldi, E.; Bucherelli, C.; Blandina, P.; Passani, M.B. Histaminergic ligands injected into the nucleus basalis magnocellularis differentially affect fear conditioning consolidation. Int. J. Neuropsychopharmacol. 2013, 16, 575–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sadek, B.; Saad, A.; Subramanian, D.; Shafiullah, M.; Lazewska, D.; Kiec-Kononowiczc, K. Anticonvulsant and procognitive properties of the non-imidazole histamine H3 receptor antagonist DL77 in male adult rats. Neuropharmacology 2016, 106, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Azimullah, S.; Jayaprakash, P.; Jayaraj, R.L.; Reiner, D.; Ojha, S.K.; Beiram, R.; Stark, H.; Lazewska, D.; Kiec-Kononowicz, K.; et al. The dual-active histamine H3 receptor antagonist and acetylcholine esterase inhibitor E100 ameliorates stereotyped repetitive behavior and neuroinflammmation in sodium valproate induced autism in mice. Chem. Biol. Interact. 2019, 312, 108775. [Google Scholar] [CrossRef] [PubMed]
- Sudha, K.; Rao, A.V.; Rao, A. Oxidative stress and antioxidants in epilepsy. Clinica. Chimica. Acta. 2001, 303, 19–24. [Google Scholar] [CrossRef]
- Pearson-Smith, J.N.; Patel, M. Metabolic Dysfunction and Oxidative Stress in Epilepsy. Int. J. Mol. Sci. 2017, 18, 2365. [Google Scholar] [CrossRef] [Green Version]
- Puttachary, S.; Sharma, S.; Stark, S.; Thippeswamy, T. Seizure-induced oxidative stress in temporal lobe epilepsy. Biomed. Res. Int. 2015, 2015, 745613. [Google Scholar] [CrossRef]
- Packer, L. Oxidants, antioxidant nutrients and the athlete. J. Sports. Sci. 1997, 15, 353–363. [Google Scholar] [CrossRef]
- Bellissimo, M.I.; Amado, D.; Abdalla, D.S.; Ferreira, E.C.; Cavalheiro, E.A.; Naffah-Mazzacoratti, M.G. Superoxide dismutase, glutathione peroxidase activities and the hydroperoxide concentration are modified in the hippocampus of epileptic rats. Epilepsy. Res. 2001, 46, 121–128. [Google Scholar] [CrossRef]
- Dal-Pizzol, F.; Klamt, F.; Vianna, M.M.; Schroder, N.; Quevedo, J.; Benfato, M.S.; Moreira, J.C.; Walz, R. Lipid peroxidation in hippocampus early and late after status epilepticus induced by pilocarpine or kainic acid in Wistar rats. Neurosci. Lett. 2000, 291, 179–182. [Google Scholar] [CrossRef]
- Pazdernik, T.L.; Emerson, M.R.; Cross, R.; Nelson, S.R.; Samson, F.E. Soman-induced seizures: Limbic activity, oxidative stress and neuroprotective proteins. J. Appl. Toxicol. 2001, 21, S87–S94. [Google Scholar] [CrossRef] [PubMed]
- Freitas, R.M. The evaluation of effects of lipoic acid on the lipid peroxidation, nitrite formation and antioxidant enzymes in the hippocampus of rats after pilocarpine-induced seizures. Neurosci. Lett. 2009, 455, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Reyes, S.; Santillan-Cigales, J.J.; Jimenez-Osorio, A.S.; Pedraza-Chaverri, J.; Guevara-Guzman, R. Glycyrrhizin ameliorates oxidative stress and inflammation in hippocampus and olfactory bulb in lithium/pilocarpine-induced status epilepticus in rats. Epilepsy. Res. 2016, 126, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos Sales, I.M.; do Nascimento, K.G.; Feitosa, C.M.; Saldanha, G.B.; Feng, D.; de Freitas, R.M. Caffeic acid effects on oxidative stress in rat hippocampus after pilocarpine-induced seizures. Neurol. Sci. 2011, 32, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Freitas, R.M.; Vasconcelos, S.M.; Souza, F.C.; Viana, G.S.; Fonteles, M.M. Oxidative stress in the hippocampus after pilocarpine-induced status epilepticus in Wistar rats. Febs. J. 2005, 272, 1307–1312. [Google Scholar] [CrossRef]
- McDonough, J.H., Jr.; Shih, T.M. Neuropharmacological mechanisms of nerve agent-induced seizure and neuropathology. Neurosci. Biobehav. Rev. 1997, 21, 559–579. [Google Scholar] [CrossRef]
- Freitas, R.M.; Bezerra Felipe, C.F.; Nascimento, V.S.; Oliveira, A.A.; Viana, G.S.; Fonteles, M.M. Pilocarpine-induced seizures in adult rats: Monoamine content and muscarinic and dopaminergic receptor changes in the striatum. Comp. Biochem. Physiol. C. Toxicol. Pharmacol. 2003, 136, 103–108. [Google Scholar] [CrossRef]
- Giovagnoli, A.R.; Avanzini, G. Learning and memory impairment in patients with temporal lobe epilepsy: Relation to the presence, type, and location of brain lesion. Epilepsia 1999, 40, 904–911. [Google Scholar] [CrossRef]
- Giacobini, E. Cholinesterase inhibitor therapy stabilizes symptoms of Alzheimer disease. Alzheimer. Dis. Assoc. Disord. 2000, 14, S3–S10. [Google Scholar] [CrossRef]
- Costa, D.A.; de Oliveira, G.A.; Lima, T.C.; dos Santos, P.S.; de Sousa, D.P.; de Freitas, R.M. Anticonvulsant and antioxidant effects of cyano-carvone and its action on acetylcholinesterase activity in mice hippocampus. Cell Mol. Neurobiol. 2012, 32, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Ezz, H.S.; Khadrawy, Y.A.; Noor, N.A. The neuroprotective effect of curcumin and Nigella sativa oil against oxidative stress in the pilocarpine model of epilepsy: A comparison with valproate. Neurochem. Res. 2011, 36, 2195–2204. [Google Scholar] [PubMed]
- Freitas, R.M.; Sousa, F.C.; Viana, G.S.; Fonteles, M.M. Acetylcholinesterase activities in hippocampus, frontal cortex and striatum of Wistar rats after pilocarpine-induced status epilepticus. Neurosci. Lett. 2006, 399, 76–78. [Google Scholar] [CrossRef] [PubMed]
- Orsetti, M.; Ghi, P.; Di Carlo, G. Histamine H(3)-receptor antagonism improves memory retention and reverses the cognitive deficit induced by scopolamine in a two-trial place recognition task. Behav. Brain. Res. 2001, 124, 235–242. [Google Scholar] [CrossRef]
- Santos, I.M.; Tome Ada, R.; Saldanha, G.B.; Ferreira, P.M.; Militao, G.C.; Freitas, R.M. Oxidative stress in the hippocampus during experimental seizures can be ameliorated with the antioxidant ascorbic acid. Oxid. Med. Cell. Longev. 2009, 2, 214–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krause, M.; Stark, H.; Schunack, W. Azomethine prodrugs of (R)-alpha-methylhistamine, a highly potent and selective histamine H3-receptor agonist. Curr. Med. Chem. 2001, 8, 1329–1340. [Google Scholar] [CrossRef]
- Javed, H.; Azimullah, S.; Abul Khair, S.B.; Ojha, S.; Haque, M.E. Neuroprotective effect of nerolidol against neuroinflammation and oxidative stress induced by rotenone. BMC. Neurosci. 2016, 17, 58. [Google Scholar] [CrossRef] [Green Version]
- Javed, H.; Azimullah, S.; Meeran, M.F.N.; Ansari, S.A.; Ojha, S. Neuroprotective Effects of Thymol, a Dietary Monoterpene Against Dopaminergic Neurodegeneration in Rotenone-Induced Rat Model of Parkinson’s Disease. Int. J. Mol. Sci. 2019, 20, 1538. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Samples of the compounds reported in this contribution are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Percentage of SE Incidence (%) | Percentage of Survival (%) |
|---|---|---|
| SAL | 0.00 | 100.00 |
| PLC (400 mg/kg) | 100.00 # | 66.67 |
| DZP (10 mg/kg) | 0.00 ** | 100.00 |
| E177 (2.5 mg/kg) | 50.00 * | 100.00 |
| E177 (5 mg/kg) | 50.00 * | 100.00 |
| E177 (10 mg/kg) a | 33.33 ** | 100.00 |
| E177 (15 mg/kg) | 50.00* | 83.33 |
| E177 a + RAM b | 66.67 | 100.00 |
| E177 a + PYR c | 50.00 | 100.00 |
| E177 a + ZOL d | 100.00 $ | 83.33 |
| RAM + PLC | 83.33 | 100.00 |
| PYR + PLC | 83.33 | 83.33 |
| ZOL + PLC | 66.67 | 100.00 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alachkar, A.; Azimullah, S.; Ojha, S.K.; Beiram, R.; Łażewska, D.; Kieć-Kononowicz, K.; Sadek, B. The Neuroprotective Effects of Histamine H3 Receptor Antagonist E177 on Pilocarpine-Induced Status Epilepticus in Rats. Molecules 2019, 24, 4106. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24224106
Alachkar A, Azimullah S, Ojha SK, Beiram R, Łażewska D, Kieć-Kononowicz K, Sadek B. The Neuroprotective Effects of Histamine H3 Receptor Antagonist E177 on Pilocarpine-Induced Status Epilepticus in Rats. Molecules. 2019; 24(22):4106. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24224106
Chicago/Turabian StyleAlachkar, Alaa, Sheikh Azimullah, Shreesh K. Ojha, Rami Beiram, Dorota Łażewska, Katarzyna Kieć-Kononowicz, and Bassem Sadek. 2019. "The Neuroprotective Effects of Histamine H3 Receptor Antagonist E177 on Pilocarpine-Induced Status Epilepticus in Rats" Molecules 24, no. 22: 4106. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24224106