
Inhibitory Effects of Myrtucommuacetalone 1 (MCA-1) from Myrtus Communis on Inflammatory Response in Mouse Macrophages

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Inhibition of NO Production by Myrtucommuacetalone 1 (MCA-1) in THP1 Monocyte and U138MG Cells
2.2. Effect of MCA-1 on LPS-Induced NFκB and p38 Kinase Activation
2.3. Effect of MCA-1 on iNOS Expression, NFκB Phosphorylation, and the iNOS Protein
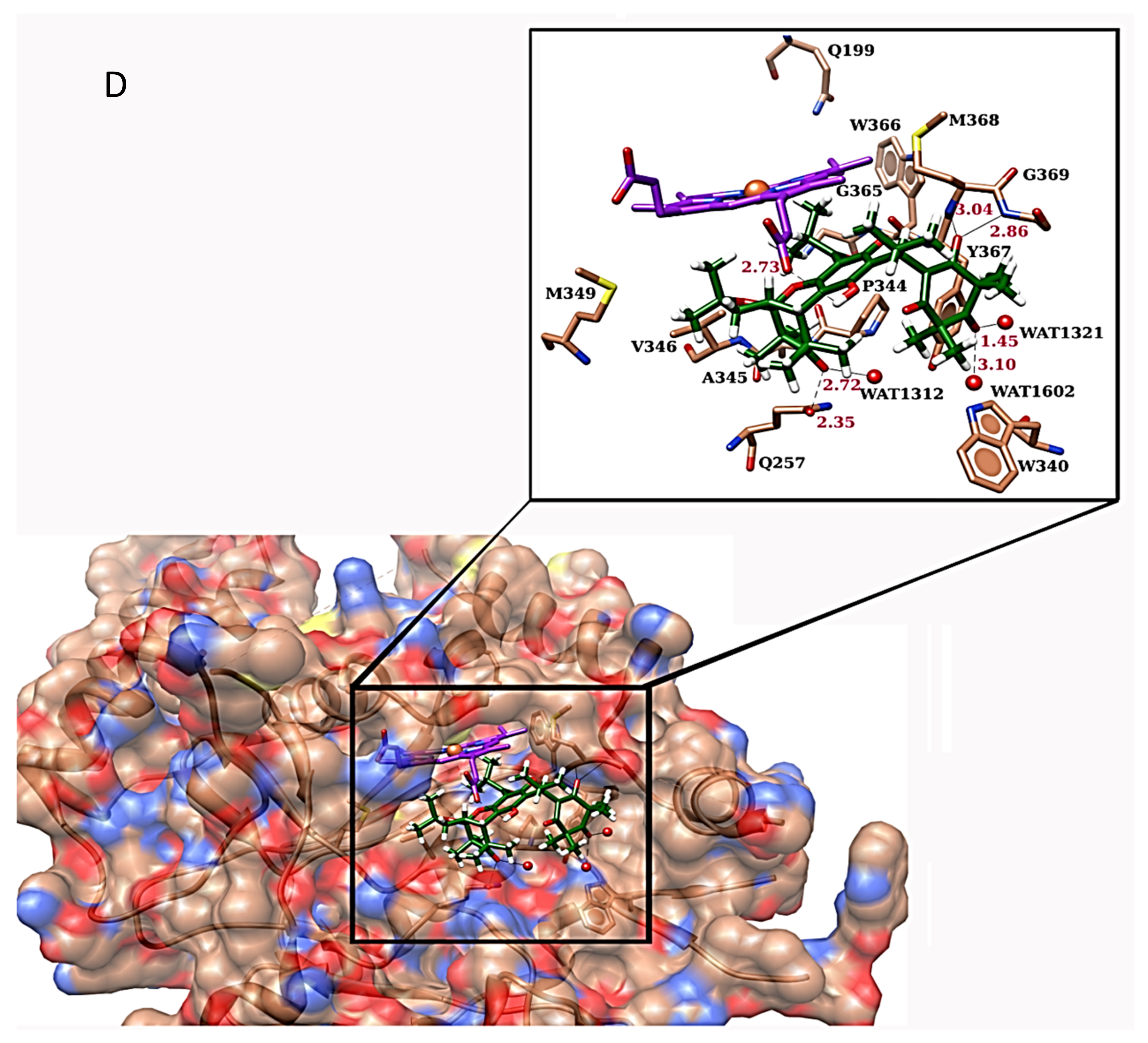
2.4. Molecular Docking of iNOS Ligand with MCA-1
2.5. Effect of MCA-1 on Cell Viability and Anticancer Activity
2.6. Effect of Compound on ROS Generation and H2O2 Production
3. Discussion
4. Materials and Methods
4.1. Plant Material, Extraction and Isolation
4.2. Animals, Cells, Chemicals, and Reagents
4.3. Cell Culture
4.4. Determination of Nitrite Oxide Concentration
4.5. Determination of NFκB and p38 Kinase Translocation
4.6. Determination of iNOS Expression by RT-PCR
4.7. Detection of iNOS and p65 Proteins
4.8. Determination of Superoxide Production by NBT Assay
4.9. H2O2 Determination
4.10. Determination of Cytotoxicity
4.11. Molecular Docking Studies for iNOS Inhibition
4.11.1. FRED 3.0.0
4.11.2. Redocking Experiment
4.12. Physicochemical and Pharmacokinetic Predictions of MCA-1
4.13. Data Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Semeraro, N.; Ammollo, C.T.; Semeraro, F.; Colucci, M. Sepsis-associated dissemenated intravascular coagulation and thromboembolic disease. J. Hematol. Infect. Dis. 2010, 2, 2010–2024. [Google Scholar]
- Choudhary, M.I.; Khan, N.; Ahmad, M.; Yousuf, S.; Fun, H.K.; Soomro, S.; Asif, M.; Mesaik, M.A.; Shaheen, F. New inhibitors of ROS generation and T-cell proliferation from Myrtus communis. Org. Lett. 2013, 15, 1862–1865. [Google Scholar] [CrossRef] [PubMed]
- Aktan, F. iNOS-mediated nitric oxide production and its regulation. Life Sci. 2004, 75, 639–653. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, J.; Galley, H.F.; Webster, N.R. Oxidative stress and gene expression in sepsis. Br. J. Anaesth. 2003, 90, 221–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karin, M. NF-κB as a critical link between inflammation and cancer. Cold Spring Harb Perspect Biol. 2009, 1, 5. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Kasai, K.; Gross, S.S. NO suppresses while peroxynitrite sustains NF-κB: A paradigm to rationalize cytoprotective and cytotoxic actions attributed to NO. Cardiovasc. Res. 2004, 63, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Song, D.; Ye, X.; Liu, S.F. A pivotal role of endothelial-specific NF-kappaB signaling in the pathogenesis of septic shock and septic vascular dysfunction. J. Immunol. 2009, 183, 4031–4038. [Google Scholar] [CrossRef] [Green Version]
- Leoni, L.A.; Fukushima, A.R.; Rocha, L.Y.; Maifrino, L.B.; Rodrigues, B. Physical activity on endothelial and erectile dysfunction: A literature review. Aging Male. 2014, 4, 1–6. [Google Scholar] [CrossRef]
- Griscavage, J.M.; Fukuto, J.M.; Komori, Y.; Ignarro, L.J. Nitric oxide inhibits neuronal nitric oxide synthase by interacting with the heme prosthetic group: Role of tetrahydrobiopterin in modulating the inhibitory action of nitric oxide. J. Biol. Chem. 1994, 269, 21644–21649. [Google Scholar]
- Watts, C. Identifying the neighborhoods of LPS signaling. Nat. Immunol. 2008, 9, 343–345. [Google Scholar] [CrossRef]
- Chen, Y.C.; Shen, S.C.; Lee, W.R.; Hou, W.C.; Yang, L.L.; Lee, T.J. Inhibition of nitric oxide synthase inhibitors and lipopolysaccharide induced inducible NOS and cyclooxygenase-2 gene expressions by rutin, quercetin, and quercetin pentaacetate in RAW 264.7 macrophages. J. Cell Biochem. 2001, 82, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.C.; Jo, J.; Park, J.; Kang, H.K.; Park, Y. NF-κB Signaling Pathways in Osteoarthritic Cartilage Destruction. Cells 2019, 8, 734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelletier, J.P.; Jovanovic, D.V.; Lascau-Coman, V.; Fernandes, J.C.; Manning, P.T.; Connor, J.R.; Currie, M.G.; Martel-Pelletier, J. Selective inhibition of inducible nitric oxide synthase reduces progression of experimental osteoarthritis in vivo: Possible link with the reduction in chondrocyte apoptosis and caspase 3 level. Arthritis Rheum. 2000, 43, 1290–1299. [Google Scholar] [CrossRef]
- Fujioka, S.; Niu, J.; Schmidt, C.; Sclabas, G.M.; Peng, B.; Uwagawa, T.; Li, Z.; Evans, D.B.; Abbruzzese, J.L.; Chiao, P.J. NF-kappaB and AP-1 connection: Mechanism of NF-kappaB-dependent regulation of AP-1 activity. Mol. Cell. Biol. 2004, 24, 7806–7819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilgun-Sherki, Y.; Melamed, E.; Offen, D. The role of oxidative stress in the pathogenesis of multiple sclerosis: The need for effective antioxidant therapy. J. Neurol. 2004, 251, 261–268. [Google Scholar]
- Van de Schans, S.A.; Janssen-Heijnen, M.L.; Biesma, B.; Smeenk, F.W.; van de Poll-Franse, L.V.; Seynaeve, C.; Coebergh, J.W. COPD in cancer patients: Higher prevalence in the elderly, a different treatment strategy in case of primary tumours above the diaphragm, and a worse overall survival in the elderly patient. Eur. J. Cancer. 2007, 43, 2194–2202. [Google Scholar] [CrossRef]
- Hamalainen, M.; Nieminen, R.; Vuorela, P.; Heinonen, M.; Moilanen, E. Anti-inflammatory effects of flavonoids: Genistein, Kaempferol, Quercetin, and Daidzein inhibit STAT-1 and NF-κB activations, whereas Flavone, Isorhamnetin, Naringenin, and Pelargonidin inhibit only NF-κB activation along with their inhibitory effect on iNOS Expression and NO production in activated macrophages. Mediators Inflamm. 2007, 4, 56–73. [Google Scholar]
- Khasnavis, S.; Jana, A.; Roy, A.; Mazumder, M.; Bhushan, B.; Wood, T.; Ghosh, S.; Watson, R.; Pahan, K. Suppression of nuclear factor-kappa B activation and inflammation in microglia by a physically- modified saline. J. Biol. Chem. 2012, 287, 29529–29542. [Google Scholar] [CrossRef] [Green Version]
- Naeem, N.; Haneef, K.; Kabir, N.; Iqbal, H.; Jamall, S.; Salim, A. DNA methylation inhibitors, 5-azacytidine and zebularine potentiate the trans-differentiation of rat bone marrow mesenchymal stem cells into cardiomyocytes. Cardiovasc Ther. 2013, 31, 201–209. [Google Scholar] [CrossRef]
- Tsai, S.-H.; Lin-Shiau, S.-Y.; Lin, J.-K. Suppression of nitric oxide synthase and the down-regulation of the activation of NFκB in macrophages by resveratrol. Br. J. Pharmacol. 1999, 126, 673–680. [Google Scholar] [CrossRef] [Green Version]
- Leiro, J.; Alvarez, E.; Arranz, J.A.; Laguna, R.; Uriarte, E.; Orallo, F. Effects of cis-resveratrol on inflammatory murine macrophages: Antioxidant activity and down-regulation of inflammatory genes. J. Leukoc. Biol. 2004, 75, 1156–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drexhage, H.A.; van der Gaag, R.D.; Namavar, F. Nitroblue tetrazolium-dye reduction by rat peritoneal macrophages during the uptake of Diplococcus pneumoniae, type VI. Antonie van Leeuwenhoek 1978, 44, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Serrander, L.; Cartier, L.; Bedard, K.; Banfi, B.; Lardy, B.; Plastre, O.; Sienkiewicz, A.; Fórró, L.; Schlegel, W.; Krause, K.H. NOX4 activity is determined by mRNA levels and reveals a unique pattern of ROS generation. Biochem. J. 2007, 406, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Vachala, S.D.; Keloth, K.S.; Peralam, Y.P. Synthesis and evaluation of fused pyrimidine derivatives as anti- inflammatory, antiproliferative and antimicrobial agents. Med. Chem. Res. 2012, 21, 2998–3005. [Google Scholar] [CrossRef]
- Kuete, V.; Wabo, H.K.; Eyong, K.O.; Feussi, M.T.; Wiench, B.; Krusche, B.; Tane, P.; Folefoc, G.N.; Efferth, T. Anticancer Activities of Six Selected Natural Compounds of Some Cameroonian Medicinal Plants. PLoS ONE 2011, 6, 21762. [Google Scholar] [CrossRef]
- McGann, M.R.; Almond, H.R.; Nicholls, A.; Grant, J.A.; Brown, F.K. Gaussian. Biopolymers 2003, 68, 76–90. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, P.C.D.; Skillman, A.G.; Warren, G.L.; Ellingson, B.A.; Stahl, M.T. Conformer generation with OMEGA: Algorithm and validation using high quality structures from the Protein Databank and Cambridge Structural Database. J. Chem. Inf. Model. 2010, 50, 572–584. [Google Scholar] [CrossRef] [PubMed]
- Davey, D.D.; Adler, M.; Arnaiz, D.; Eagen, K.; Erickson, S.; Guilford, W.; Kenrick, M.; Morrissey, M.M.; Ohlmeyer, M.; Pan, G.; et al. Design, synthesis, and activity of 2- imidazol-1-ylpyrimidine derived inducible nitric oxide synthase dimerization inhibitors. J. Med. Chem. 2007, 50, 1146–1157. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the compound bank of Dr Panjwani center for molecular medicine and drug research, ICCBS, UoK. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physicochemical Properties | Formula: C38H52O9; molecular weight = 652.81 g/mol; number of heavy atoms = 47, volume = 617.87; number of aromatic heavy atoms = 6; fraction Csp3 = 0.68; number of rotatable bonds = 5; number of H-bond acceptors = 9; number of H-bond donors = 3; molar refractivity = 180.26; TPSA = 147.43 Ų; lipophilicity [Log Po/w (iLOGP)] = 3.22; water solubility (Log S) = −8.05; Caco2 (human colorectal carcinoma cell permeability) = 22.8225 nm/sec |
| Pharmacokinetic Properties | GI absorption = moderate; BBB permeant = no; P-gp substrate = yes; CYP1A2 inhibitor = no; CYP2C19 inhibitor = no; CYP2C9 inhibitor = no; CYP2D6 inhibitor = no; CYP3A4 inhibitor = yes; Log Kp (skin permeation) = −5.33 cm/s |
| Drug Likeness | Lipinski = yes, 1 violation: MW > 500; Ghose = no, 4 violations: MW > 480, WLOGP > 5.6, MR > 130, number of atoms > 70; Veber = no, 1 violation: TPSA > 140; Egan = no, 2 violations: WLOGP > 5.88, TPSA > 131.6; Muegge = no, 2 violations: MW > 600, XLOGP3 > 5; bioavailability score = 0.56 |
| Medicinal Chemistry | PAINS = 0 alert; Brenk = 1 alert: beta_keto_anhydride; leadlikeness = no, 2 violations: MW > 350, XLOGP3 > 3.5; synthetic accessibility = 7.31 |
| Toxicity | AMES test = nonmutagenic; carcinogenicity = none; predicted LD50 = 995 mg/kg; hepatotoxicity = none |
| Cancer Cell line Prediction | Hepatoblastoma (HepG2), prostate carcinoma (DU-145), adult T-acute lymphoblastic leukemia (MT4), and melanoma (M19-MEL) |
| Nontumor Cell Line Prediction | None |
| Compound | ROS (IC50) µg/mL | Oxidative Burst% Inhibition at 25 µg/mL | |
|---|---|---|---|
| O2− | H2O2 | ||
| MCA-1 | >100 | 42.5 | 53 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soomro, S.; Mesaik, M.A.; Shaheen, F.; Khan, N.; Halim, S.A.; Ul-Haq, Z.; Ali Siddiqui, R.; Choudhary, M.I. Inhibitory Effects of Myrtucommuacetalone 1 (MCA-1) from Myrtus Communis on Inflammatory Response in Mouse Macrophages. Molecules 2020, 25, 13. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25010013
Soomro S, Mesaik MA, Shaheen F, Khan N, Halim SA, Ul-Haq Z, Ali Siddiqui R, Choudhary MI. Inhibitory Effects of Myrtucommuacetalone 1 (MCA-1) from Myrtus Communis on Inflammatory Response in Mouse Macrophages. Molecules. 2020; 25(1):13. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25010013
Chicago/Turabian StyleSoomro, Samreen, M. Ahmed Mesaik, Farzana Shaheen, Noureen Khan, Sobia Ahsan Halim, Zaheer Ul-Haq, Rafat Ali Siddiqui, and Muhammad Iqbal Choudhary. 2020. "Inhibitory Effects of Myrtucommuacetalone 1 (MCA-1) from Myrtus Communis on Inflammatory Response in Mouse Macrophages" Molecules 25, no. 1: 13. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25010013