
Specificity Assessment of CRISPR Genome Editing of Oncogenic EGFR Point Mutation with Single-Base Differences

 , and
, and
Abstract
:1. Introduction
2. Results
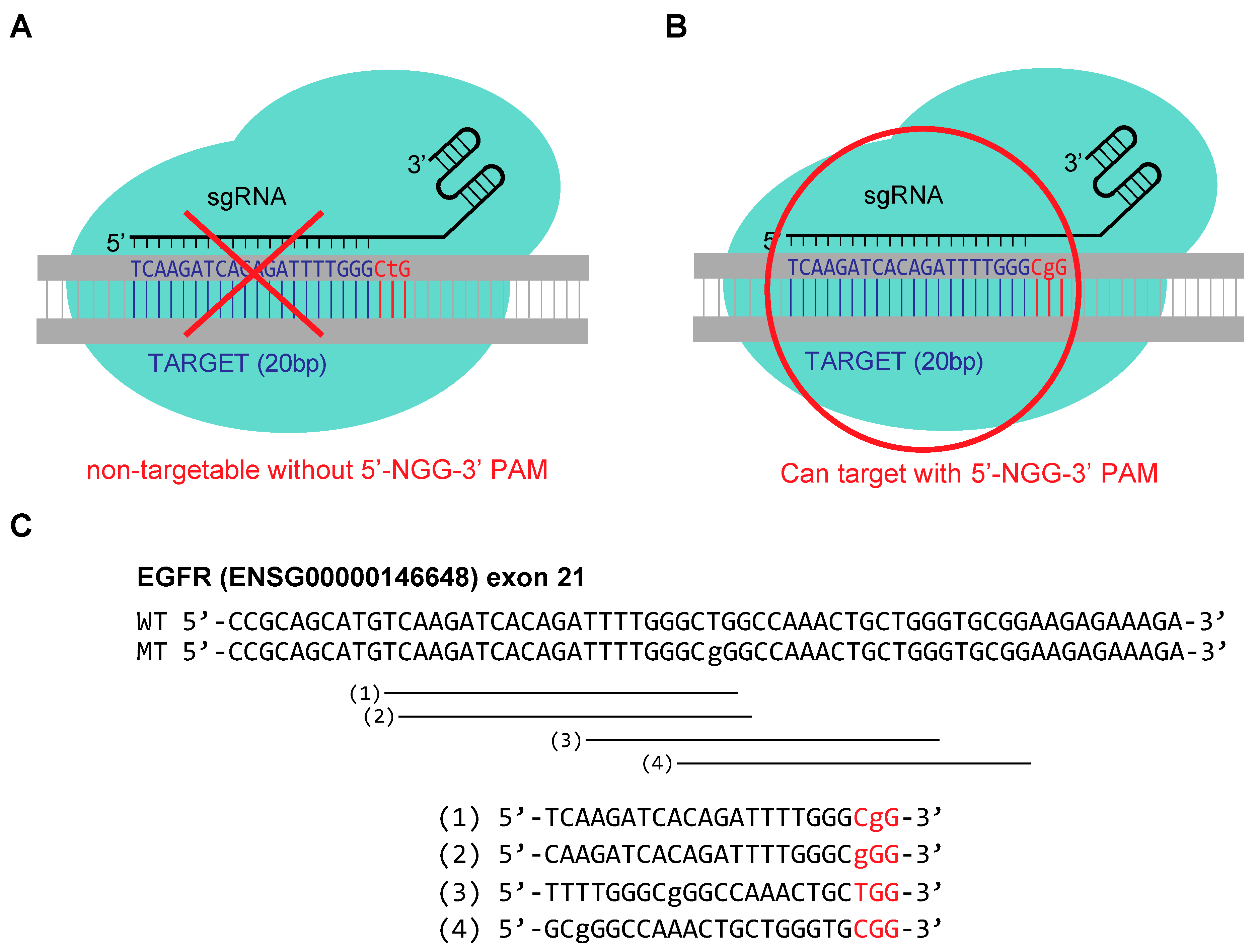
2.1. Tested Guide RNA Designs for Targeting Oncogenic Point Mutation in EGFR (2573T > G)
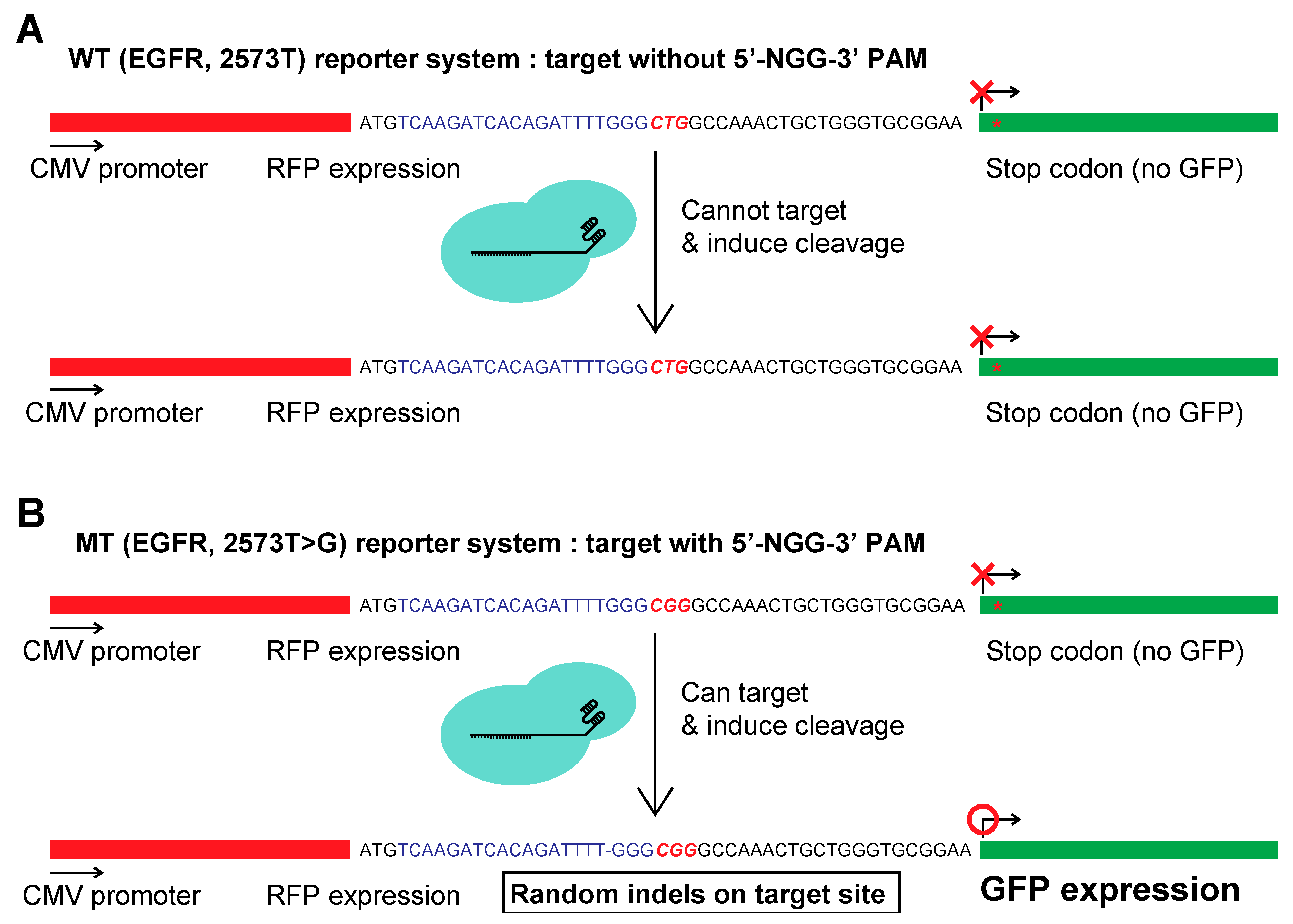
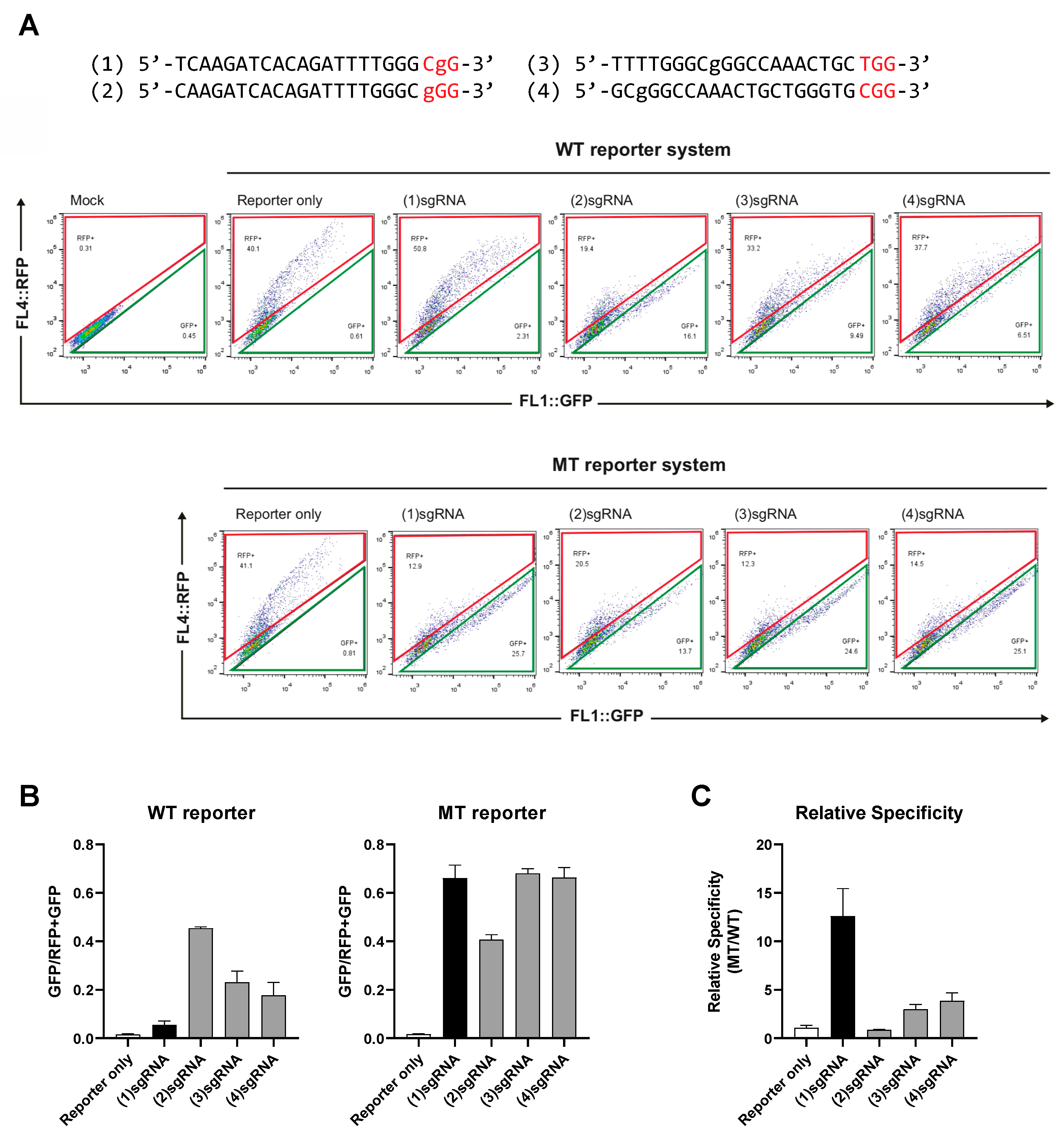
2.2. Constructing a Dual Fluorescence Reporter System for the Quantification of Genome Editing Efficiency
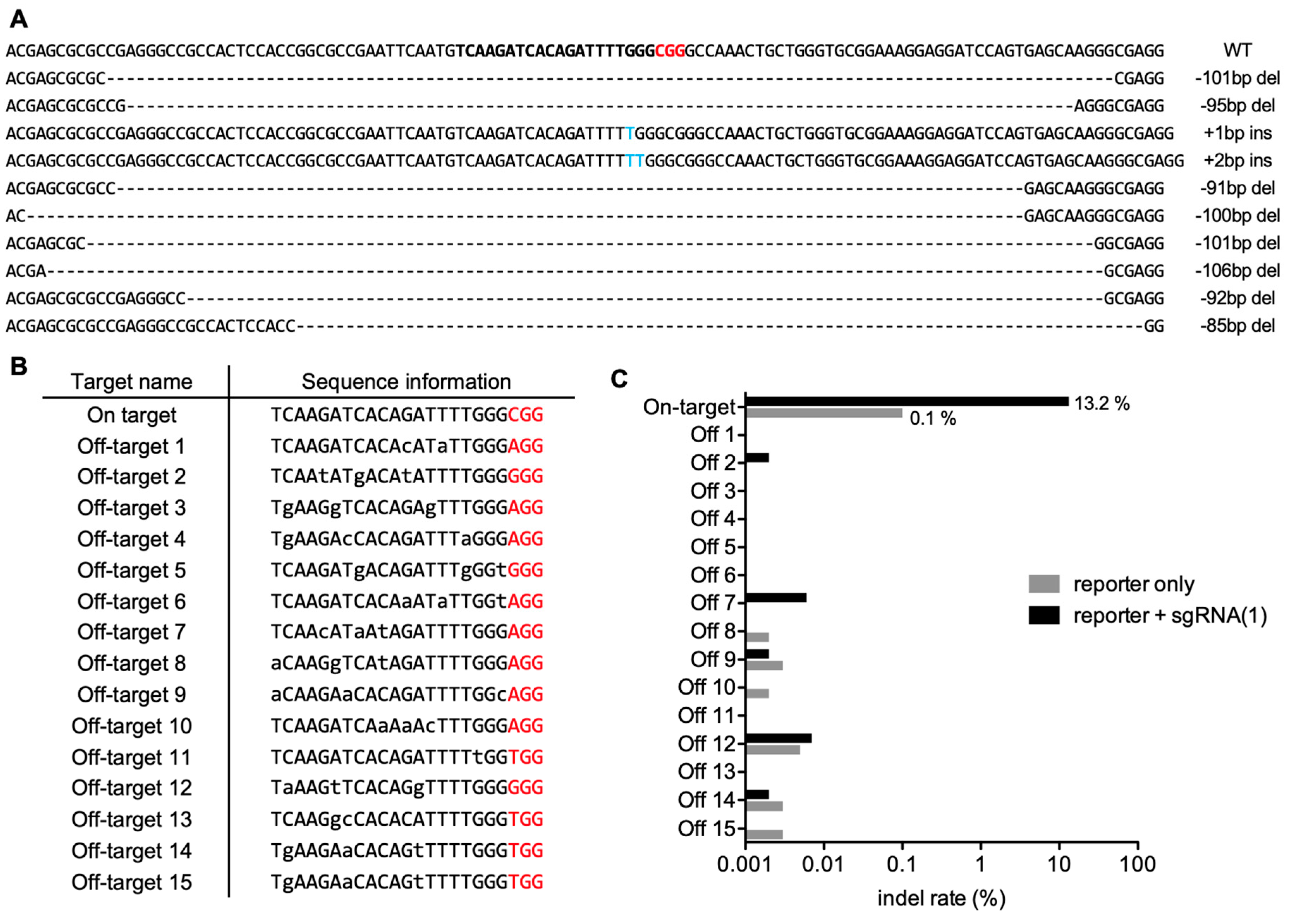
2.3. Analyses of the Indel Patterns and Genome-Wide Off-Targets of the PAM-Based Guide RNA
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Transfection
4.2. Targeted Deep Sequencing
4.3. T7E1 Assay
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Doudna, J.A.; Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA–Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Mali, P.; Yang, L.; Esvelt, K.M.; Aach, J.; Guell, M.; DiCarlo, J.E.; Norville, J.E.; Church, G.M. RNA-guided human genome engineering via Cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jinek, M.; Jiang, F.; Taylor, D.W.; Sternberg, S.H.; Kaya, E.; Ma, E.; Anders, C.; Hauer, M.; Zhou, K.; Lin, S.; et al. Structures of Cas9 Endonucleases Reveal RNA-Mediated Conformational Activation. Science 2014, 343, 1247997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimasu, H.; Ran, F.A.; Hsu, P.D.; Konermann, S.; Shehata, S.I.; Dohmae, N.; Ishitani, R.; Zhang, F.; Nureki, O. Crystal structure of Cas9 in complex with guide RNA and target DNA. Cell 2014, 156, 935–949. [Google Scholar] [CrossRef] [Green Version]
- Bae, S.; Park, J.; Kim, J.-S. Cas-OFFinder: A fast and versatile algorithm that searches for potential off-target sites of Cas9 RNA-guided endonucleases. Bioinformatics 2014, 30, 1473–1475. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Park, Y.H.; Jin, Y.B.; Kim, S.U.; Hur, J.K. CRISPR Diagnosis and Therapeutics with Single Base Pair Precision. Trends Mol. Med. 2019. [Google Scholar] [CrossRef]
- Anders, C.; Niewoehner, O.; Duerst, A.; Jinek, M. Structural basis of PAM-dependent target DNA recognition by the Cas9 endonuclease. Nature 2014, 513, 569–573. [Google Scholar] [CrossRef]
- Fonfara, I.; Le-Rhun, A.; Chylinski, K.; Makarova, K.S.; Lecrivain, A.L.; Bzdrenga, J.; Koonin, E.V.; Charpentier, E. Phylogeny of Cas9 determines functional exchangeability of dual-RNA and Cas9 among orthologous type II CRISPR-Cas systems. Nucleic Acids Res. 2014, 42, 2577–2590. [Google Scholar] [CrossRef]
- Li, Y.; Mendiratta, S.; Ehrhardt, K.; Kashyap, N.; White, M.A.; Bleris, L. Exploiting the CRISPR/Cas9 PAM Constraint for Single-Nucleotide Resolution Interventions. PLOS ONE 2016, 11, e0144970. [Google Scholar] [CrossRef]
- Kim, W.; Lee, S.; Kim, H.S.; Song, M.; Cha, Y.H.; Kim, Y.H.; Shin, J.; Lee, E.S.; Joo, Y.; Song, J.J.; et al. Targeting mutant KRAS with CRISPR-Cas9 controls tumor growth. Genome Res. 2018, 28, 374–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbst, R.S. Review of epidermal growth factor receptor biology. Int J. Radiat. Oncol. Biol. Phys. 2004, 59, 21–26. [Google Scholar] [CrossRef] [PubMed]
- da Cunha Santos, G.; Shepherd, F.A.; Tsao, M.S. EGFR mutations and lung cancer. Annu Rev. Pathol. 2011, 6, 49–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, B.; Zhang, S.; Ma, S. Management of non-small cell lung cancer with EGFR mutation: THE role of radiotherapy in the era of tyrosine kinase inhibitor therapy-opportunities and challenges. J. Thorac. Dis. 2017, 9, 3385–3393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Liu, X.; Liu, R.; Shi, T.; Li, X.; Zhong, D.; Wang, Y.; Chen, G.; Chen, J. Concurrent epidermal growth factor receptor T790M secondary mutation and epithelial-mesenchymal transition in a lung adenocarcinoma patient with EGFR-TKI drug resistance. Thorac. Cancer 2017, 8, 693–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, H.; Xue, W.; Anderson, D.G. CRISPR–Cas: A tool for cancer research and therapeutics. Nat. Rev. Clin. Oncol. 2019, 16, 281–295. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Lage, M.; Puig-Serra, P.; Menendez, P.; Torres-Ruiz, R.; Rodriguez-Perales, S. CRISPR/Cas9 for Cancer Therapy: Hopes and Challenges. Biomedicines 2018, 6, 105. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Gu, T.; Patel, S.; Bode, A.M.; Lee, M.H.; Dong, Z. CRISPR/Cas9-An evolving biological tool kit for cancer biology and oncology. NPJ Precis. Oncol. 2019, 3, 8. [Google Scholar] [CrossRef]
- Koo, T.; Yoon, A.R.; Cho, H.Y.; Bae, S.; Yun, C.O.; Kim, J.S. Selective disruption of an oncogenic mutant allele by CRISPR/Cas9 induces efficient tumor regression. Nucleic Acids Res. 2017, 45, 7897–7908. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.W.; Kim, S.; Kim, J.M.; Kim, J.S. Targeted genome engineering in human cells with the Cas9 RNA-guided endonuclease. Nat. Biotechnol. 2013, 31, 230–232. [Google Scholar] [CrossRef]
- Gu, W.; Crawford, E.D.; O-Donovan, B.D.; Wilson, M.R.; Chow, E.D.; Retallack, H.; DeRisi, J.L. Depletion of Abundant Sequences by Hybridization (DASH): Using Cas9 to remove unwanted high-abundance species in sequencing libraries and molecular counting applications. Genome Biol. 2016, 17, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Yu, J.; Hwang, G.H.; Kim, S.; Kim, H.S.; Ye, S.; Kim, K.; Park, J.; Park, D.Y.; Cho, Y.K.; et al. CUT-PCR: CRISPR-mediated, ultrasensitive detection of target DNA using PCR. Oncogene 2017, 36, 6823–6829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pattanayak, V.; Lin, S.; Guilinger, J.P.; Ma, E.; Doudna, J.A.; Liu, D.R. High-throughput profiling of off-target DNA cleavage reveals RNA-programmed Cas9 nuclease specificity. Nat. Biotechnol. 2013, 31, 839–843. [Google Scholar] [CrossRef] [PubMed]
- Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; van der Oost, J.; Regev, A.; et al. Cpf1 Is a Single RNA-Guided Endonuclease of a Class 2 CRISPR-Cas System. Cell 2015, 163, 759–771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Samples of the reagents are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene (Primer Direction) | Primer Sequence (5′ to 3′) | Off-Target Sequence |
|---|---|---|
| EGFR_Off_1_F1 | TAAATTGAGTGCAGAGCCTTG | TCAAGATCACAcATaTTGGGAGG |
| EGFR_Off_1_R1 | GCAATGGTACCCTTTTCTTC | |
| EGFR_Off_1_F2 | AAACCCTGTGTTTGGAATTCA | |
| EGFR_Off_1_R2 | TTTGTTGTCCTTATATGTCATTGTG | |
| EGFR_Off_2_F1 | TCAGAGCCCAGTTTATCATAG | TCAAtATgACAtATTTTGGGGGG |
| EGFR_Off_2_R1 | GTAAATTAGCTGGTCTCAGG | |
| EGFR_Off_2_F2 | GGCATCAAGTAAATTACTCAAAACA | |
| EGFR_Off_2_R2 | CATGTAGCCAAAACAAAACAAACA | |
| EGFR_Off_3_F1 | ATTCAACAACGATCTGTTGTG | TgAAGgTCACAGAgTTTGGGAGG |
| EGFR_Off_3_R1 | AAAGCAAAACCCCAAAGCGA | |
| EGFR_Off_3_F2 | TCTAGTGGAGACAGATGTTAAAC | |
| EGFR_Off_3_R2 | GGGCTTTCCTCTCTTCTGT | |
| EGFR_Off_4_F1 | TGCATTTTGACTCTTCTCGA | TgAAGAcCACAGATTTaGGGAGG |
| EGFR_Off_4_R1 | AGAAGAAAGCTTCTGATGCC | |
| EGFR_Off_4_F2 | ATATCCAATAACAACAATACTAGC | |
| EGFR_Off_4_R2 | CCATCATTGAATTGAGGTCAC | |
| EGFR_Off_5_F1 | GGAAATCAAATTTGGGGTTG | TCAAGATgACAGATTTgGGtGGG |
| EGFR_Off_5_R1 | CTGCTGTTCCCCAACTTATT | |
| EGFR_Off_5_F2 | TCTGAGACTGGGTCATTCAT | |
| EGFR_Off_5_R2 | GCTTTTTACAACTATCTTACTAATAAC | |
| EGFR_Off_6_F1 | CCTTGTTGATTTACATTGATGTG | TCAAGATCACAaATaTTGGtAGG |
| EGFR_Off_6_R1 | CATTCCATAACATTCTCAGGT | |
| EGFR_Off_6_F2 | CCAATGTAAATCAACAAGGGT | |
| EGFR_Off_6_R2 | AATCATGAACATATTTATGCTTTCC | |
| EGFR_Off_7_F1 | CAAACTCACAATTGTGAGGG | TCAAcATaAtAGATTTTGGGAGG |
| EGFR_Off_7_R1 | AGCAGAGCATGGAGCTCATA | |
| EGFR_Off_7_F2 | ATGGACCGGGTGGCTTAAA | |
| EGFR_Off_7_R2 | AATGAAGCATGGTGACAGAC | |
| EGFR_Off_8_F1 | ACCAGGATGGTCTTGATCTC | aCAAGgTCAtAGATTTTGGGAGG |
| EGFR_Off_8_R1 | AAGTGTTTAAAACAGTGCCA | |
| EGFR_Off_8_F2 | CTGCCCAGGCTTCATCTTAA | |
| EGFR_Off_8_R2 | CCAGGCTGAAATGATCAAAG | |
| EGFR_Off_9_F1 | ATTTCAAAGGGTGGGGCTTT | aCAAGAaCACAGATTTTGGcAGG |
| EGFR_Off_9_R1 | GAAGTCTCAGATCAAGGTCC | |
| EGFR_Off_9_F2 | TTGGGACTTGTCATCCTTTT | |
| EGFR_Off_9_R2 | GGAGAAGAGCATGAGTGCTA | |
| EGFR_Off_10_F1 | TAGACTAGTCACCAGAATTCC | TCAAGATCAaAaAcTTTGGGAGG |
| EGFR_Off_10_R1 | TCACCATGCAGTTGTACATA | |
| EGFR_Off_10_F2 | CAAGGAAGACAAAAGAGACA | |
| EGFR_Off_10_R2 | TCCCCAGTCTGTTCTCTCTT | |
| EGFR_Off_11_F1 | GTTATGTGAGGTGTTTGTGT | TCAAGATCACAGATTTTtGGTGG |
| EGFR_Off_11_R1 | AGAACACACCATGTTAGAGG | |
| EGFR_Off_11_F2 | TGATGATAATTTCATGTGTGTTACC | |
| EGFR_Off_11_R2 | ACTCTCTTTGGTGAGAAGGA | |
| EGFR_Off_12_F1 | TACAGTTGTGTGGCTTTCGA | TaAAGtTCACAGgTTTTGGGGGG |
| EGFR_Off_12_R1 | GCCGTCTCAATACTTGTGAA | |
| EGFR_Off_12_F2 | AACAACTATGGTATGGGCCA | |
| EGFR_Off_12_R2 | GGTAAAACCCCATCTACTAAAAA | |
| EGFR_Off_13_F1 | GCCTCTTATCTGAACGAGAA | TCAAGgcCACAcATTTTGGGTGG |
| EGFR_Off_13_R1 | ATTAGTTGCAGTTCAAAGCC | |
| EGFR_Off_13_F2 | AAATCTACTTGGAGCAATGC | |
| EGFR_Off_13_R2 | TTTTTGGTAGGAGCCTGCAG | |
| EGFR_Off_14_F1 | AGTGTTTCGATAGATGGAGG | TgAAGAaCACAGtTTTTGGGTGG |
| EGFR_Off_14_R1 | CTACCCAAAACCTTTGTCCC | |
| EGFR_Off_14_F2 | AGAAGAGCACGAGTGGTAAA | |
| EGFR_Off_14_R2 | CACCCTTTTTCTTCCTCCAT | |
| EGFR_Off_15_F1 | GGTGGGGAAAAAAGTTTTTGG | TgAAGAaCACAGtTTTTGGGTGG |
| EGFR_Off_15_R1 | CTACACCTTCTTTTCCCGAC | |
| EGFR_Off_15_F2 | GAGTGACGAGGAGGAGGAAA | |
| EGFR_Off_15_R2 | TCAACACCCTTTTCCCCAT | |
| EGFR_on-target_F | TACTTGAAGCTGTCCTTCCC | TCAAGATCACAGATTTTGGG |
| EGFR_on-target_R | CCGTCGTCCTTGAAGAAGAT |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bae, T.; Kim, H.; Kim, J.H.; Kim, Y.J.; Lee, S.H.; Ham, B.-J.; Hur, J.K. Specificity Assessment of CRISPR Genome Editing of Oncogenic EGFR Point Mutation with Single-Base Differences. Molecules 2020, 25, 52. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25010052
Bae T, Kim H, Kim JH, Kim YJ, Lee SH, Ham B-J, Hur JK. Specificity Assessment of CRISPR Genome Editing of Oncogenic EGFR Point Mutation with Single-Base Differences. Molecules. 2020; 25(1):52. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25010052
Chicago/Turabian StyleBae, Taegeun, Hanseop Kim, Jeong Hee Kim, Yong Jun Kim, Seung Hwan Lee, Byung-Joo Ham, and Junho K. Hur. 2020. "Specificity Assessment of CRISPR Genome Editing of Oncogenic EGFR Point Mutation with Single-Base Differences" Molecules 25, no. 1: 52. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25010052