
Iridoids: Research Advances in Their Phytochemistry, Biological Activities, and Pharmacokinetics


Abstract
:1. Introduction
2. Phytochemistry
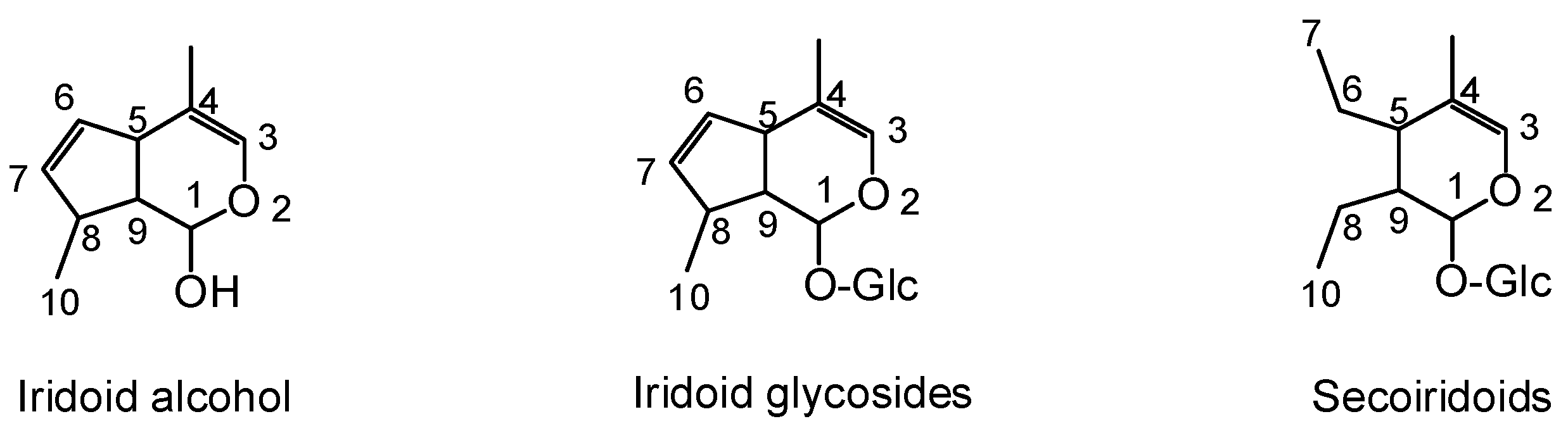
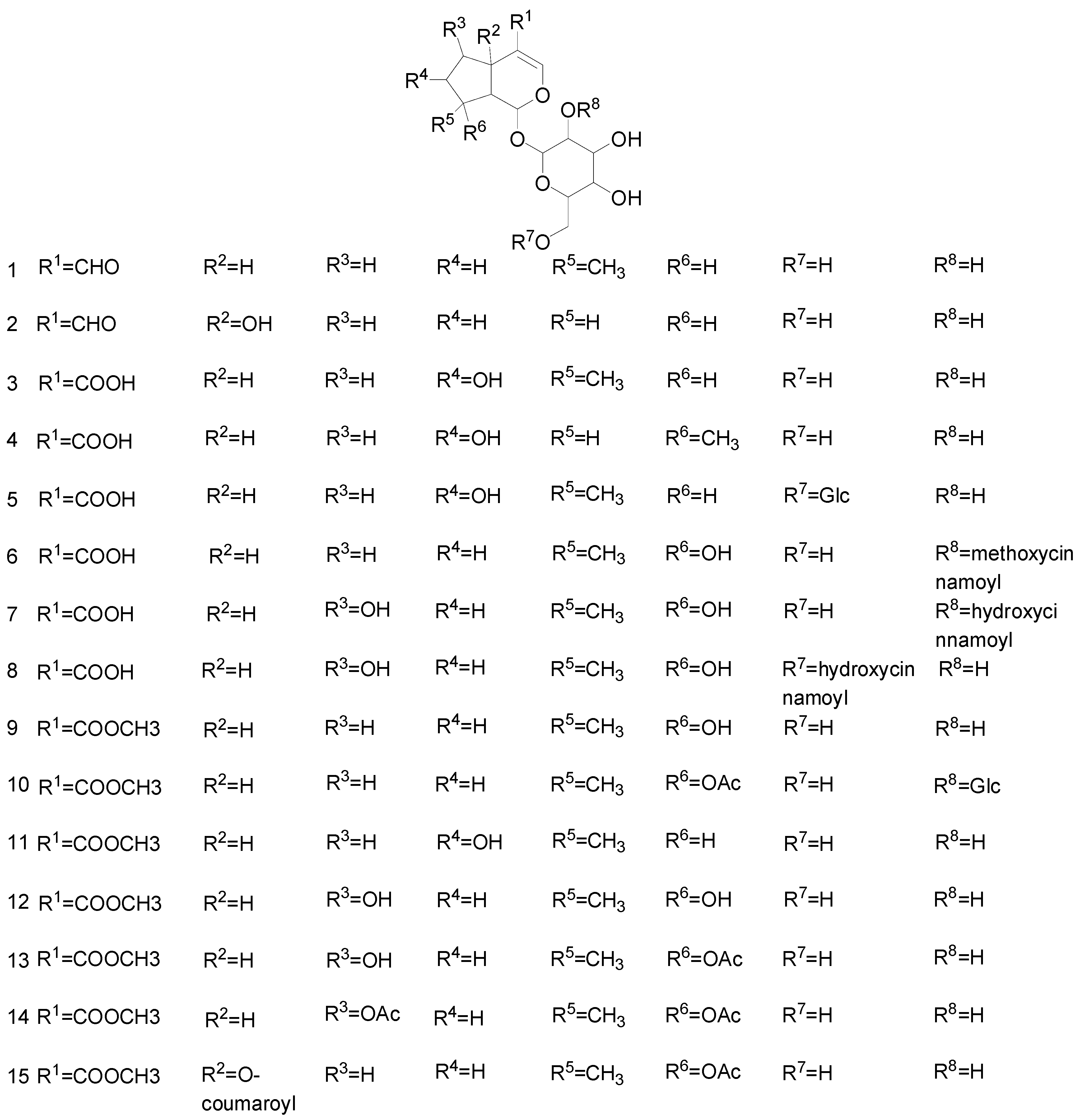
2.1. Iridoid Glycosides
2.2. Secoiridoids Glycosides
2.3. Bis-Iridoids
2.4. Non-Glycosidic Iridoids
3. Biological Activities
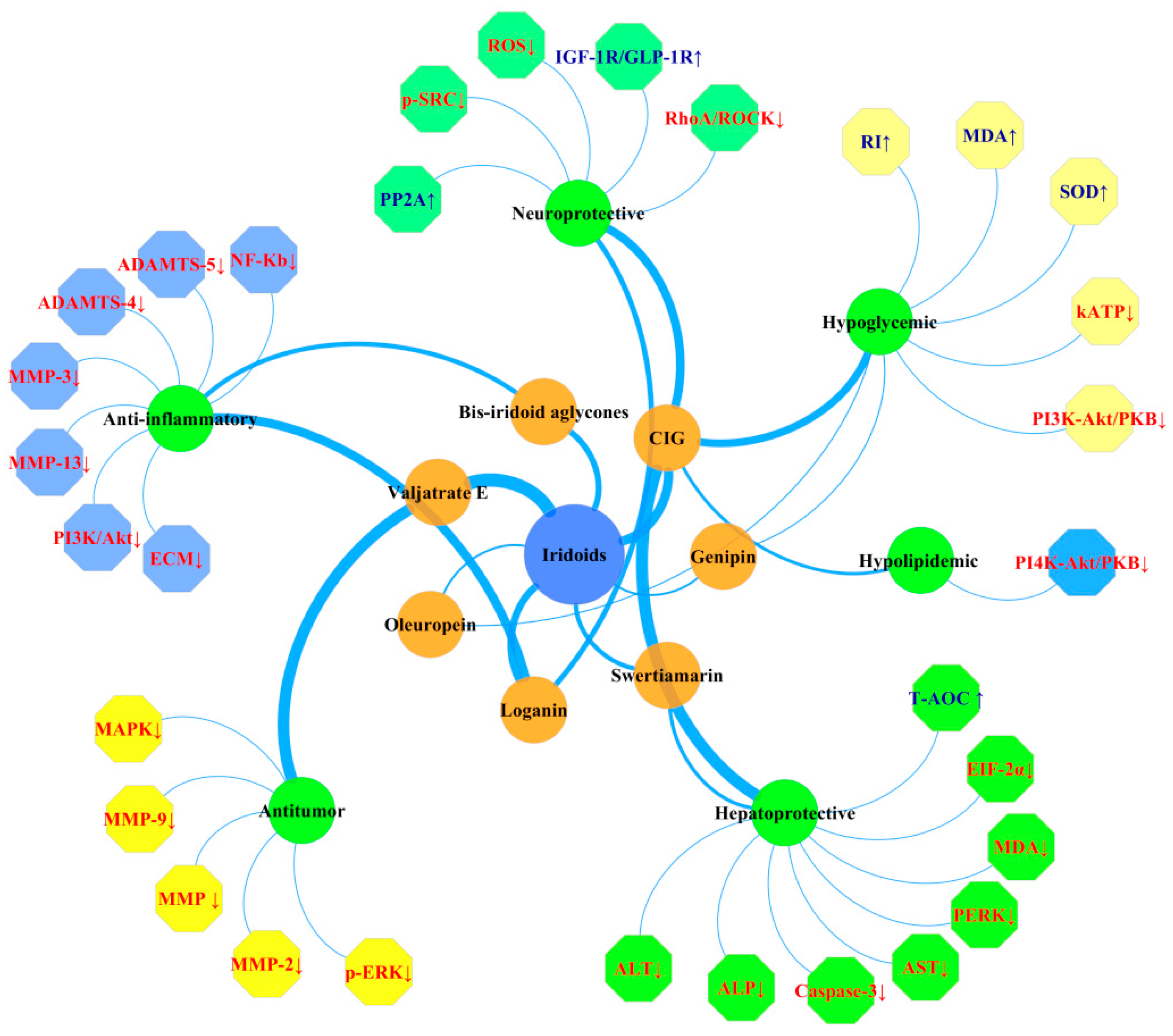
3.1. Neuroprotective Effects
3.2. Hepatoprotective Effects
3.3. Anti-Inflammatory Activities
3.4. Hypoglycemic and Hypolipidemic Activities
3.5. Antitumor Activities
3.6. Others
4. Pharmacokinetic of Iridoids
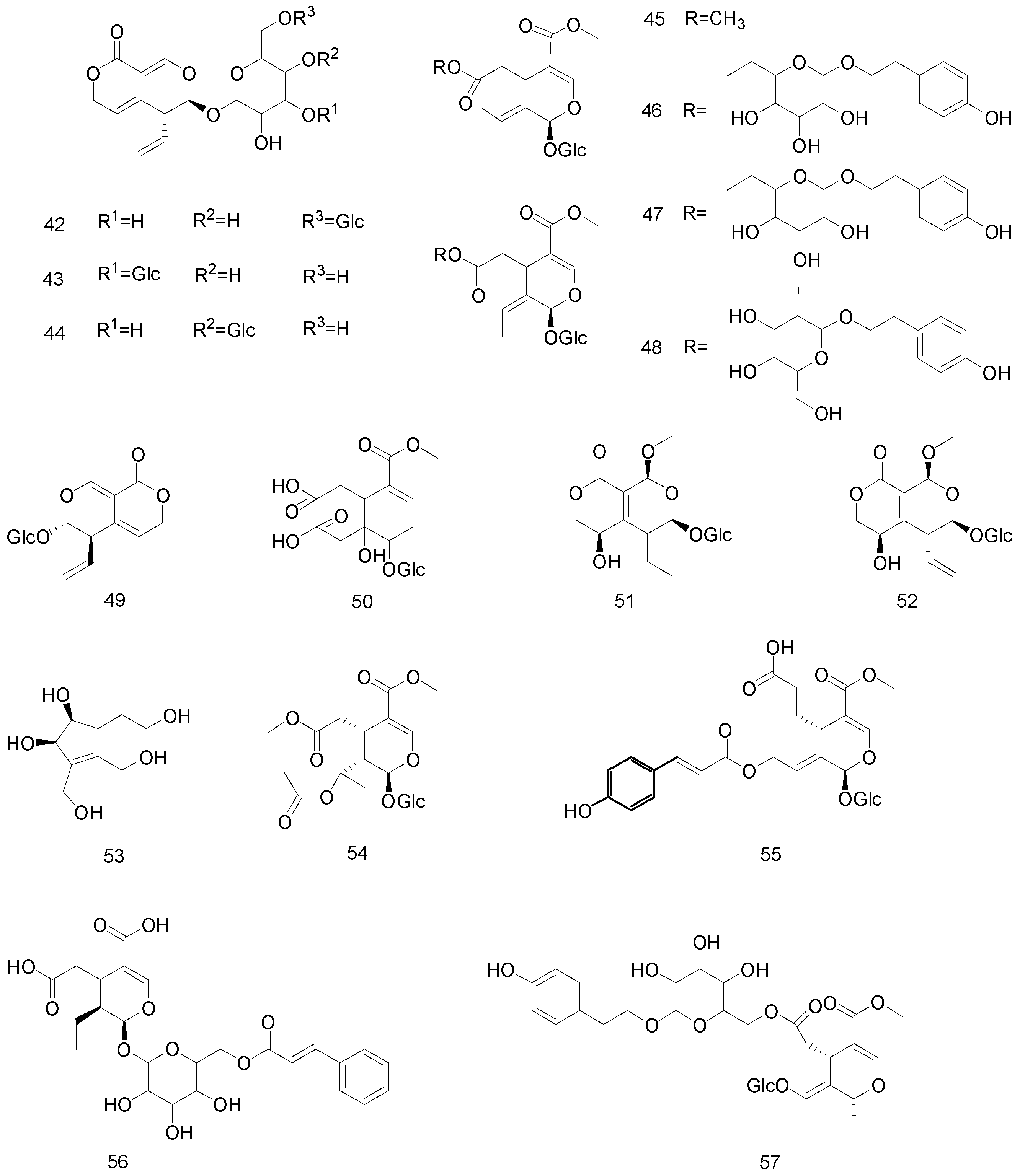
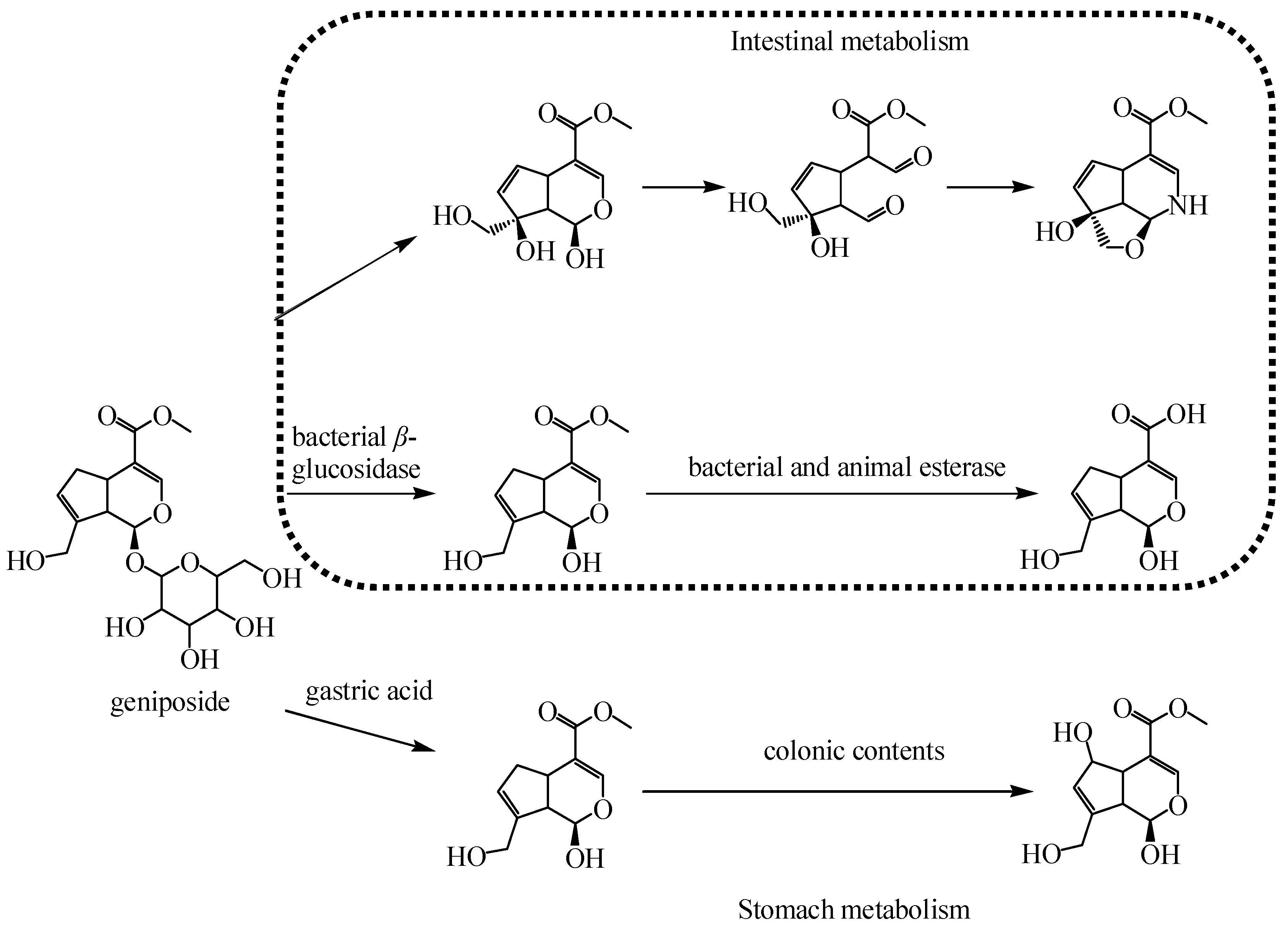
4.1. Iridoid Glycosides
4.2. Secoiridoids
4.3. Others
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Li, C.M.; Luo, Y.W.; Tian, B.Y. Research Progress on Mass Spectral Fragmentation of Iridoids. J. Hebei Norm. Univ. (Nat Sci. Edit). 2015, 39, 522–526. [Google Scholar]
- Pei, Y.H.; Lou, H.X. Natural Medicinal Chemistry; People’s Health Publishing House: Beijing, China, 2016; pp. 192–195. [Google Scholar]
- Bianco, A. Recent developments in iridoids chemistry. Pure Appl. Chem. 1994, 66, 2335–2338. [Google Scholar] [CrossRef]
- Zheng, L.S.; Liu, X.Q. Research progress of iridoids. Res. Dev. Nat. Prod. 2009, 21, 702–711. [Google Scholar]
- Ren, Z.J.; Zhang, L.M.; He, K.Z. Extraction Technology and Pharmacological Research Progress of Main Components of Gardenia jasminoides Ellis. Res. Dev. Nat. Prod. 2005, 17, 831–836. [Google Scholar]
- Picerno, P.; Autore, G.; Marzocco, S.; Meloni, M.; Sanogo, R.; Aquino, R.P. Anti-inflammatory activity of verminoside from kigelia a fricana and evaluation of cutaneous irritation in cell cultures and reconstituted human epidermis. J. Nat. Prod. 2005, 68, 1610–1614. [Google Scholar] [CrossRef] [PubMed]
- Akihisa, T.; Matsumoto, K.; Tokuda, H.; Yasukawa, K.; Seino, K.I.; Nakamoto, K.; Kimura, Y. Anti-inflammatory and potential cancer chemopreventive constituents of the fruits of Morinda citrifolia (Noni). J. Nat. Prod. 2007, 70, 754–757. [Google Scholar] [CrossRef]
- Dong, J.E.; Zhang, J. Advances in the Research of Iridoids Occurring in Plants. J. Northwest For. Univ. 2004, 19, 131–135. [Google Scholar]
- El-Naggar, L.J.; Beal, J.L. Iridoids. A review. J. Nat. Prod. 1980, 43, 649–707. [Google Scholar] [CrossRef]
- Zheng, H.R.; Zhu, Y.; Li, W.; Ding, L. Research progress on pharmacokinetics of natural iridoids. Drug Eval. Res. 2018, 41, 217–223. [Google Scholar]
- Wang, F.F.; Zhang, Y.M.; Zheng, X.W.; Dai, Z.; Liu, B.; Ma, S.C. Research Progress of the Structure and Biological Activities of Iridoids Compounds. China Acad. J. Electron. Publ. House 2019, 33, 91–98. [Google Scholar]
- Tundis, R.; Loizzo, M.; Menichini, F.; Statti, G.A.; Menichini, F. Biological and Pharmacological Activities of Iridoids: Recent Developments. Mini Rev. Med. Chem. 2008, 8, 399–420. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; He, X.; Tao, R.; Cao, X.Y. Phytochemistry and Pharmacology of the Genus Pedicularis Used in Traditional Chinese Medicine. Am. J. Chin. Med. 2014, 42, 1071–1098. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.J.; Jin, H.Z.; Shen, Y.H.; Qin, J.J.; Wang, Y.; Huang, Y.; Zeng, Q.; Zhang, W.D. Chemical constituents of plants from the genus incarvillea. Chem. Biodivers. 2009, 6, 818–826. [Google Scholar] [CrossRef] [PubMed]
- Zajdel, S.M.; Graikou, K.; GOwniak, K.; Chinou, L. Chemical analysis of Penstemon campanulatus (Cav.) Willd.—Antimicrobial activities. Fitoterapia 2012, 83, 373–376. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.Q.; Yang, Z.L.; Xu, L.; Li, P.; Hu, Y.Z. Akebia, saponin D, a saponin component from Dipsacus asper Wall, protects PC 12 cells against amyloid-β induced cytotoxicity. Cell Biol. Int. 2009, 33, 1102–1110. [Google Scholar] [CrossRef]
- Daniil, O.; Nina, K.; Nadezhda, C.; Nadezhda, C.; Larisa, T. Iridoids and Flavonoids of Four Siberian Gentians: Chemical Profile and Gastric Stimulatory Effect. Molecules 2015, 20, 19172–19188. [Google Scholar]
- Behbahani, M. Evaluation of anti-HIV-1 activity of a new iridoid glycoside isolated from Avicenna marina, in vitro. Int. Immunopharmacol. 2014, 23, 262–266. [Google Scholar] [CrossRef]
- Zhang, H.; Feng, N.; Xu, Y.T.; Li, T.X.; Gao, X.M.; Zhu, Y.; Song, Y.S.; Wang, Y.N.; Wu, H.H. Chemical constituents from the flowers of wild Gardenia jasminoides Ellis. Chem. Biodivers. 2017, 14, e1600437. [Google Scholar] [CrossRef]
- Ata, A.; Kalhari, K.S.; Samarasekera, R. Chemical constituents of Barleria prionitis and their enzyme inhibitory and free radical scavenging activities. Phytochem. Lett. 2009, 2, 37–40. [Google Scholar] [CrossRef]
- Yu, Y.; Xie, Z.L.; Gao, H.; Ma, W.W.; Dai, Y.; Wang, Y.; Zhong, Y.; Yao, X.S. Bioactive Iridoid Glucosides from the Fruit of Gardenia jasminoides. J. Nat. Prod. 2009, 72, 1459–1464. [Google Scholar] [CrossRef]
- Ata, A.; Mbong, N.; Iverson, C.D.; Samarasekera, R. Minor Chemical Constituents of Vitex pinnata. Nat. Prod. Commun. 2009, 4. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Huang, X.; Du, X.; Sun, W.J.; Zhang, Y.M. Study of chemical composition and antimicrobial activity of leaves and roots of Scrophularia ningpoensis. Nat. Prod. Res. 2009, 23, 775–780. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.; Qi, J.; Yu, B.Y. Iridoid and phenylpropanoid glycosides from Scrophularia ningpoensis Hemsl. and their α-Glucosidase inhibitory activities. Fitoterapia 2014, 93, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Bai, M.; Li, S.F.; Liu, S.F.; Wang, X.B.; Huang, X.X.; Song, S.J. Iridoid glycoside and lignans from a wild vegetable (Patrinia villosa, Juss.) with antioxidant activity. J. Food Biochem. 2018, 42, e12521. [Google Scholar] [CrossRef]
- Ali, E.G.; Shaza, A.M.; Ghada, F.; Hanan, A.; Adnan, A.R.; Omer, B.; Maged, A.K.; Nurhayat, T.; James, B. Chemical Composition of Buddleja polystachya Aerial Parts and its Bioactivity against Aedes aegypti. Nat. Prod. Lett. 2018, 32, 2775–2782. [Google Scholar]
- Vestena, A.; Piton, Y.; Bordignon, S.A.; Garcia, S.; Arbo, M.D.; Zuanazzi, J.A.; Poser, G. Hepatoprotective activity of Verbena litoralis, Verbena montevidensis and their main iridoid, brasoside. J. Ethnopharmacol. 2019, 239, 111906. [Google Scholar] [CrossRef]
- Anh, N.T.H.; Tam, K.T.; Tuan, N.V.; Thien, D.D.; Qua, T.D.; Tam, N.T.; Bao, N.C.; Do, T.T.; Nga, N.T.; Thuy, T.T.; et al. Chemical constituents of Oldenlandia pinifolia and their antiproliferative activities. Nat. Prod. Lett. 2019, 33, 796–802. [Google Scholar]
- Adom, M.B.; Taher, M.; Mutalabisin, M.F.; Amri, M.S.; Kudos, M.B.A.; Sulaiman, M.W.A.W.; Sengupta, P.; Susanti, D. Chemical constituents and medical benefits of Plantago major. Biomed. Pharmacother. Biomed. Pharmacother. 2017, 96, 348–360. [Google Scholar] [CrossRef]
- Berger, A.; Fasshuber, H.; Schinnerl, J.; Robien, W.; Brecker, L.; Valant-Vetschera, K. Iridoids as chemical markers of false ipecac (Ronabea emetica), a previously confused medicinal plant. J. Ethnopharmacol. 2011, 138, 756–761. [Google Scholar] [CrossRef]
- Saracoglu, I.; Harput, U.S. In Vitro Cytotoxic Activity and Structure Activity Relationships of Iridoid Glucosides Derived from Veronica species. Phytother. Res. 2012, 26, 148–152. [Google Scholar] [CrossRef]
- Shancai, T.; Qiuxia, L.; Yueyue, S.; Yiran, S.; Fang, C.; Lin, T. Iridoid Glycosides Fraction Isolated from Veronica ciliata Fisch. Protects against Acetaminophen-Induced Liver Injury in Mice. Evid.-Based Complement. Altern. Med. 2017, 2017, 6106572. [Google Scholar]
- Khan, M.F.; Dixit, P.; Jaiswal, N.; Tamrakar, A.K.; Srivastava, A.K.; Maurya, R. Chemical constituents of Kigelia pinnata twigs and their GLUT4 translocation modulatory effect in skeletal muscle cells. Fitoterapia 2012, 83, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Lv, T.; Xu, M.; Wang, D.; Zhu, H.T.; Yang, C.R.; Zhang, T.T.; Zhang, Y.J. The chemical constituents from the roots of Gentiana crassicaulisand their inhibitory effects on inflammatory mediators NO and TNF-α. Nat. Prod. Bioprospect. 2012, 2, 217–221. [Google Scholar] [CrossRef] [Green Version]
- Gao, B.B.; She, G.M.; She, D.M. Chemical Constituents and Biological Activities of Plants from the Genus Ligustrum. Chem. Biodivers. 2013, 10, 96–128. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, J.; Zhao, Y.L.; Huang, H.Y.; Wang, Y.Z. Comprehensive Quality Assessment Based Specific Chemical Profiles for Geographic and Tissue Variation in Gentiana rigescens Using HPLC and FTIR Method Combined with Principal Component Analysis. Front. Chem. 2017, 5, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khobrakova, V.B.; Budaeva, E.R.; Olennikov, D.N.; Zilfikarov, I.N. Immunomodulating Activity of Extract of Gentiana algida Pall. Pharm. Chem. J. 2017, 51, 384–387. [Google Scholar] [CrossRef]
- Zhang, L.; Zou, D.Z.; Bai, S.; Li, Z.H.; Zhang, C.H. Chemical constituents from Gentianella turkestanorum (Gentianaceae). Biochem. Syst. Ecol. 2016, 65, 89–92. [Google Scholar] [CrossRef]
- Gao, L.; Li, C.; Wang, Z.; Liu, X.Q.; You, Y.; Wei, H.; Guo, T. Ligustri Lucidi Fructus as a traditional Chinese medicine: A review of its phytochemistry and pharmacology. Nat. Prod. Res. 2015, 29, 493–510. [Google Scholar] [CrossRef]
- Qiu, Z.C.; Zhao, X.X.; Wu, Q.C.; Fu, J.R.; Dai, Y.; Wong, Z.S.; Yao, X.S. New secoiridoids from the fruits of Ligustrum lucidum. J. Asian Nat. Prod. Res. 2018, 20, 431–438. [Google Scholar] [CrossRef]
- Chen, Y.; Yu, H.; Guo, F.; Wu, Y.C.; Li, Y.M. Antinociceptive and anti-inflammatory activities of a standardizedextract of bis-iridoids from Pterocephalus hookeri. J. Ethnopharmacol. 2018, 216, 233–238. [Google Scholar] [CrossRef]
- de Ávila, J.M.; Pereira, A.O.; Zachow, L.L.; Gehm, A.Z.; Santos, M.Z.; Mostardeiro, M.A.; Back, D.; Morel, A.F.; Dalcol, I.I. Chemical constituents from Valeriana polystachya Smith and evaluation of their effects on the acetylcholinesterase and prolyl oligopeptidase activities. Fitoterapia 2018, 131, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.S.S.; Veitch, N.C.; Simmonds, M.S.J. Leaf Chemistry and Foliage Avoidance by the Thrips Frankliniella occidentalis and Heliothrips haemorrhoidalis in Glasshouse Collections. J. Chem. Ecol. 2011, 37, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Mueller, L.G.; de Andrade Salles, L.; Sakamoto, S. Effect of Storage Time and Conditions on the Diene Valepotriates Content of the Extract of Valeriana glechomifolia Obtained by Supercritical Carbon Dioxide. Phytochem. Anal. 2012, 23, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Ralambonirina Rasoarivelo, T.S.; Grougnet, R.; Michel, S.; Guillou, C.R.; Deguin, B. Chemical study of Anthospermum emirnense (Rubiaceae). Biochem. Syst. Ecol. 2017, 70, 186–191. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Han, H.; Yang, B.Y.; Xia, Y.K.; Kuang, H.X. Two New Iridoid Glycosides from the Root Barks of Sambucus williamsii Hance. Molecules 2011, 16, 3869–3874. [Google Scholar] [CrossRef] [PubMed]
- Dinda, B. Pharmacology of Iridoids; Pharmacology and Applications of Naturally Occurring Iridoids; Springer: Cham, Switzerland, 2019; pp. 1–15. [Google Scholar] [CrossRef]
- Qi, J.; Li, N.; Zhou, J.; Yu, B.Y.; Qiu, S.X. Isolation and Anti-Inflammatory Activity Evaluation of Triterpenoids and a Monoterpenoid Glycoside from Harpagophytum procumbens. Planta Med. 2010, 76, 1892–1896. [Google Scholar] [CrossRef] [Green Version]
- Alaniya, M.D.; Sutiashvili, M.G.; Shalashvili, K.G.; Skhirtladze, A.V.; Mshvildadze, V.D.; Pichette, A. Iridoid Glycosides from Roots of Verbascum laxum. Chem. Nat. Compd. 2014, 50, 842–845. [Google Scholar] [CrossRef]
- Xu, R.X. The mechanism and effect of nerve growth factor in nerve injury repair. Chin. J. Neurotrauma Surg. (Electron. Ed.) 2016, 2, 1–4. [Google Scholar]
- Dong, T.J.; Cui, Y.L.; Tian, J.S.; Yao, K.D. Advances in studies on natural iridoids. Chin. Tradit. Herb. Drugs. 2011, 42, 185–194. [Google Scholar]
- Habtemariam, S. Iridoids and Other Monoterpenes in the Alzheimer’s Brain: Recent Development and Future Prospects. Molecules. 2018, 23, 117. [Google Scholar] [CrossRef] [Green Version]
- Tseng, Y.T.; Lin, W.J.; Chang, W.H.; Lo, Y.C. The novel protective effects of loganin against 1-methyl-4-phenylpyridinium-induced neurotoxicity: Enhancement of neurotrophic signaling, activation of IGF-1R/GLP-1R, and inhibition of RhoA/ROCK pathway. Phytother. Res. 2019, 33, 690–701. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.C.; Kuai, X.X.; Zhang, L.; Li, Y.L.; Li, L.; Zhang, L. Effect of cornel iridoid glycoside on PP2Ac phosphorylation in okadaic acid-induced neurotoxicity cells. Acta Pharm. Sin. 2018, 53, 1036–1041. [Google Scholar]
- Ji, L.; Wang, X.; Li, J.; Zhong, X.; Zhang, B.; Juan, J.; Shang, X. New Iridoid Derivatives from the Fruits of Cornus officinalis and Their Neuroprotective Activities. Molecules 2019, 24, 625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chester, K.; Paliwal, S.; Khan, W.; Ahmad, S. UPLC-ESI-MS/MS and HPTLC Method for Quantitative Estimation of Cytotoxic Glycosides and Aglycone in Bioactivity Guided Fractions of Solanum nigrum L. Front. Pharm. 2017, 8, 434. [Google Scholar] [CrossRef]
- Tian, C.; Zhang, T.; Wang, L.; Shan, Q.; Jiang, L. The hepatoprotective effect and chemical constituents of total iridoids and xanthones extracted from Swertia mussotii Franch. J. Ethnopharmacol. 2014, 154, 259–266. [Google Scholar] [CrossRef]
- Li, D.X.; Jin, Y.R.; Jin, H.Y.; Li, C.H. Protective effects and the mechanisms of Boschniakia rossica iridoid glycoside on liver fibrosis in rats. J. Med. Sci. Yanbian Univ. 2016, 39, 175–178. [Google Scholar]
- Ma, Y.X.; Wu, M.H.; Jiang, Z.Q.; Zhao, F.M.; Li, L.; Li, M.H.; Lu, T.L.; Hang, A.W. Protective effect of cornel iridoid glycoside on hepatocytes injured by D-galactosamine/tumor necrosis factor-α. Chin. Pharmacol. Bull. 2018, 34, 118–122. [Google Scholar]
- Wang, L.H.; Ji, H.; Zhang, B.T.; Xiao, P.Z.; Zhang, B. Research Progress on Hepatoprotective Effect of Chinese Herbal Medicine. Addit. World. 2012, 11, 41–45. [Google Scholar]
- Liu, Y.; Yao, H.; Huang, H. Research progress on hepatoprotective effect of Uygur medicines. Northwest Pharm. J. 2018, 33, 279–284. [Google Scholar]
- Zhu, W.J. Preventive and Therapeutic Effects of Iridiod Glycosides from Paederia Scandens on Uric Acid Nephropathy in Rats and Its Mechanisms. Master’s Thesis, Anhui Medical University, Hefei, China, May 2013. [Google Scholar]
- Guo, J.; Zhang, X.S.; Liu, J.P. Regulation of iridoid glucosides from Corni officinalis Fructus on AGEs-induced inflammatory response in renal mesangial cells. Chin. Tradit. Pat. Med. 2013, 35, 2067–2072. [Google Scholar]
- Liang, l.; Bi, Q.; Dong, J.C.; Yang, X.X.; Yu, J. Progress on the development of natural medicine with hepatoprotective effects. Biot. Resour. 2018, 40, 148–158. [Google Scholar]
- Zhao, D.D.; Li, X.K.; Yu, N.; Mu, Q.Q.; Zhang, Y.; Fang, X.; Mo, F.F.; Gao, S.H. Effects of Iridoid Glycoside Extract from Corni Fructus Extract on Mitochondrial Oxygen Consumption and related Genes Expression in 3T3-L1 Mature Adipocytes. Chin. J. Exp. Tradit. Med. Formulae 2016, 22, 122–126. [Google Scholar]
- Mao, G.H.; Zhang, L.Q.; Qian, F.; Chen, K.X.; Xu, J.W.; Li, Y.M. Research progress on anti-inflammatory activity of three kinds of iridoid glycosides in Chinese materia medica. Chin. Tradit. Herb. Drugs. 2019, 50, 225–233. [Google Scholar]
- Luo, S.H.; Fu, S.C. The progress of study on anti -inflammatory mechanism of Chinese medicine. Pharm. Care Res. 2003, 3, 47–49. [Google Scholar]
- Huan, B.; Lei, X.Y. Progress in the research on the anti-inflammatory effect of traditional Chinese medicine and its mechanism. Chin. J. Clin. Ration. Med. Use 2015, 8, 177–179. [Google Scholar]
- Miao, Y.L.; Zhang, W.X.; Wang, Y.E.; Li, Y.; Yue, Y.H.; Ni, Y. Anti-inflammatory Mechanism of Antipyretic and Detoxifying Traditional Chinese Medicine. Chin. J. Exp. Tradit. Med. Formulae 2018, 24, 228–235. [Google Scholar]
- Li, J.; Yu, X.; Ma, Y.X.; Li, X.R. Advances in the research on anti-inflammatory mechanism of traditional Chinese medicine and its active components. Acta Chin. Med. Pharmacol. 2010, 38, 134–137. [Google Scholar]
- Suh, W.S.; Kwon, O.K.; Lee, T.H.; Subedi, L.; Kim, S.Y.; Lee, K.R. Secoiridoid Glycosides from the Twigs of Ligustrum obtusifolium Possess Anti-inflammatory and Neuroprotective Effects. Chem. Pharm. Bull. 2018, 66, 78–83. [Google Scholar] [CrossRef] [Green Version]
- Lei, X.X.; Su, Y.Y.; Li, M.Y.; Gui, H. Research progress of loganin and morroniside from cornel iridoid glycosides. Shanghai J. Tradit. Chin. Med. 2018, 52, 104–108. [Google Scholar]
- Yang, Y.; Gu, Y.T.; Zhao, H.; Zhang, S.L. Loganin Attenuates Osteoarthritis in Rats by Inhibiting IL-1β-Induced Catabolism and Apoptosis in Chondrocytes Via Regulation of Phosphatidylinositol 3-Kinases (PI3K)/Akt. Med. Sci. Monit. 2019, 25, 4159–4168. [Google Scholar] [CrossRef]
- Wang, F.; Jia, Q.W.; Yuan, Z.H.; Lv, L.Y.; Li, M.; Jiang, Z.B.; Liang, D.L.; Zhang, D.Z. An anti-inflammatory C-stiryl iridoid from Camptosorus sibiricus Rupr. Fitoterapia 2019, 134, 378–381. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Weimin, L.I. Retrospective Study of the Efficacy of Acarbose Combined with Metformin in the Treatment of Type 2 Diabetes Complicated with Hyperlipidemia. China Pharm. 2017, 20, 284–286. [Google Scholar]
- Jiefang, K.; Chen, G.; Rodolfo, T. Hypoglycemic, hypolipidemic and antioxidant effects of iridoid glycosides extracted from Corni fructus: Possible involvement of the PI3K–Akt/PKB signaling pathway. RSC Adv. 2018, 8, 30539–30549. [Google Scholar]
- Parton, L.E.; Ye, C.P.; Coppari, R. Glucose sensing by POMC neurons regulates glucose homeostasis and is impaired in obesity. Nature 2007, 449, 228–232. [Google Scholar] [CrossRef]
- Al-Azzawie, H.F.; Alhamdani, M.S.S. Hypoglycemic and antioxidant effect of oleuropein in alloxan-diabetic rabbits. Life Sci. 2006, 78, 1371–1377. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, D.C. Research progress on mechanism of hypoglycemic Chinese medicine. Chin. J. Clin. Ration. Drug Use 2017, 10, 159–161. [Google Scholar]
- Zhang, M.; Wang, H.H.; Zang, L.Q. Research progress on ingredients and medmechanisms of hypoglycemic traditional Chinese medicines. J. Guangdong Pharm. Univ. 2011, 3, 320–323. [Google Scholar]
- Chen, M.L.; Hou, X.T.; Hao, E.W.; Du, Z.C.; Xie, J.L.; Qin, J.F.; Deng, J.G. Research progress on the pharmacological action and mechanism of hypoglycemic action of traditional Chinese medicine. J. Chin. Med. Mater. 2018, 41, 2016–2020. [Google Scholar]
- Gong, X.; Ji, M.Y.; Xu, J.P.; Zhang, C.H.; Li, M.H. Hypoglycemic effects of bioactive ingredients from medicine food homology and medicinal health food species used in China. Crit. Rev. Food Sci. Nutr. 2019, 59, 1–24. [Google Scholar] [CrossRef]
- Yao, D.D.; Shu, L.; Yang, L.; Jia, X.B. Advance in studies on anti-diabetic mechanism of Gardeniae Fructus and its active ingredient geniposide. China J. Chin. Mater. Med. 2014, 8, 1368–1373. [Google Scholar]
- Li, X.H.; Yang, C.L.; Shen, H. Gentiopicroside exerts convincing antitumor effects in human ovarian carcinoma cells (SKOV3) by inducing cell cycle arrest, mitochondrial mediated apoptosis and inhibition of cell migration. J. BUON 2019, 24, 280–284. [Google Scholar] [PubMed]
- Sun, Y.; Lan, M.; Chen, X.; Dai, Y.; Zhao, X.; Wang, L.; Zhang, T.T.; Li, Y.B.; Zhu, J.L.; Zhang, X.M.; et al. Anti-invasion and anti-metastasis effects of Valjatrate E via reduction of matrix metalloproteinases expression and suppression of MAPK/ERK signaling pathway. Biomed. Pharmacother. 2018, 104, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Huang, H.; Lv, J.; Jiang, N.; Li, Y.; Liu, X.; Zhao, H. Iridoid compounds from the aerial parts of Swertia mussotii Franch. with cytotoxic activity. Nat. Prod. Res. 2019, 1–6. [Google Scholar] [CrossRef]
- Li, F.; Li, W.; Li, X. Geniposide attenuates inflammatory response by suppressing P2Y14 receptor and downstream ERK1/2 signaling pathway in oxygen and glucose deprivation-induced brain microvascular endothelial cells. J. Ethnopharmacol. 2016, 185, 77–86. [Google Scholar] [CrossRef]
- Shoda, J.; Miura, T.; Utsunomiya, H. Genipin enhances Mrp2 (Abcc2)-mediated bile formation and organic anion transport in rat liver. Hepatology 2004, 39, 167–178. [Google Scholar] [CrossRef]
- Ding, S.T.; Liu, H.T.; Li, W.M. Protective effects of geniposide on human umbilical vein endothelial cell injury induced by H2O2 in vitro. Chin. Pharmacol. Bull. 2009, 25, 725–729. [Google Scholar]
- Suzuki, Y.; Kondo, K.; Ikeda, Y. Antithrombotic Effect of Geniposide and Genipin in the Mouse Thrombosis Model. Planta Med. 2001, 67, 807–810. [Google Scholar] [CrossRef] [Green Version]
- Villasenor, I.M. Bioactivities of iridoids. Anti-Inflamm. Anti-Allergy Agents Med. Chem. 2007, 6, 307–314. [Google Scholar] [CrossRef]
- Lian, M.; Sun, Y.; Lin., Y. p-JAK2 plays a key role in catalpol-induced protection against rat intestinal ischemia/reperfusion injury. RSC Adv. 2017, 7, 54369–54378. [Google Scholar] [CrossRef] [Green Version]
- Jun-Sheng, T.; Bi-Yun, S.; Huan, X. 1H-NMR-Based Metabonomic Studies on the Anti-Depressant Effect of Genipin in the Chronic Unpredictable Mild Stress Rat Model. PLoS ONE 2013, 8, e75721. [Google Scholar]
- Gutierrez, R.M.P.; Solis, R.V.; Baez, E.G. Effect on capillary permeability in rabbits of iridoids from Buddleia scordioides. Phytother. Res. 2006, 20, 542–545. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.R.; Lee, M.Y.; Ahn, K. Suppressive effect of verproside isolated from Pseudolysimachion longifolium on airway inflammation in a mouse model of allergic asthma. Int. Immunopharmacol. 2006, 6, 978–986. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Chen, K.; Feng, M.Y.; Shao, W.; Wu, J.; Chen, K.; Liang, T.M.; Liu, C. Genipin alleviates high-fat diet-induced hyperlipidemia and hepatic lipid accumulation in mice via miR-142a-5p/SREBP-1c axis. FEBS J. 2018, 285, 501–517. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.T.; Hu, W.J.; Han, G.H.; Lu, W.Q.; Jia, D.X.; Hu, M.; Wang, D. Involvement of bone morphogenetic protein–related pathways in the effect of aucubin on the promotion of osteoblast differentiation in MG63 cells. Chem.-Biol. Interact. 2018, 283, 51–58. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, D.U.; Jeong, C.S. Gardenia jasminoides Ellis ethanol extract and its constituents reduce the risks of gastritis and reverse gastric lesions in rats. Food Chem. Toxicol. 2009, 47, 1127–1131. [Google Scholar] [CrossRef]
- Ma, W.W.; Tao, Y.; Wang, Y.Y.; Peng, I.F. Effects of Gardenia jasminoides extracts on cognition and innate immune response in an adult Drosophila model of Alzheimer’s disease. Chin. J. Nat. Med. 2017, 15, 899–904. [Google Scholar] [CrossRef]
- Koo, H.J.; Lee, S.; Shin, K.H.; Kim, B.C.; Lim, C.J.; Park, E.H. Geniposide, an anti-angiogenic compound from the fruits of Gardenia jasminoides. Planta Med. 2004, 70, 467–469. [Google Scholar]
- Wang, Y.; Dai, L.; Wu, H.; Zhang, Z.R.; Wang, W.Y.; Fu, J.; Deng, R.; Li, F.; Dai, X.J.; Zhang, X. Novel anti-inflammatory target of geniposide: Inhibiting Itgbeta1/Ras-Erk1/2 signal pathway via the miRNA-124a in rheumatoid arthritis syno-vial fibroblasts. Int. Immunopharmacol. 2018, 65, 284–294. [Google Scholar] [CrossRef]
- Liu, Z.X.; Liu, C.T.; Liu, Q.B.; Ren, J.; Li, L.Z.; Huang, X.X.; Wang, Z.Z.; Song, S.J. Iridoid glycosides from the flower buds of Lonicera japonica and their nitric oxide production and α-glucosidase inhibitory activities. J. Funct. Foods. 2015, 18, 512–519. [Google Scholar] [CrossRef]
- Toriizuka, K.; Kamiki, H.; Ohmura, N.Y.; Fujii, M.; Hori, Y.; Fukumura, M.; Hirai, Y.; Isoda, S.; Nemoto, Y.; Ida, Y. Anxiolytic effect of Gardeniae Fructus-extract containing active ingredient from Kamishoyosan (KSS), a Japanese traditional Kampo medicine. Life Sci. 2005, 77, 3010–3020. [Google Scholar] [CrossRef]
- Chen, J.L.; Blanc, P.; Stoddart, C.A.; Bogan, M.; Rozhon, E.J.; Parkinson, N.; Ye, Z.J.; Cooper, R.; Balick, M.; Nanakorn, W.; et al. New Iridoids from the Medicinal Plant Barleria, prionitis with Potent Activity against Respiratory Syncytial Virus. J. Nat. Prod. 1998, 61, 1295–1297. [Google Scholar] [CrossRef]
- Chen, L.; Yan, Y.; Zhang, L. Aucubin promotes angiogenesis via estrogen receptor beta in a mouse model of hindlimb ischemia. J. Steroid Biochem. Mol. Biol. 2017, 172, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Peng, J. Central Inhibition of Arachnoid Ether Terpenoids. Master’s Thesis, Southwest Jiaotong University, Xi’an, China, May 2009. [Google Scholar]
- Shi, R.R.; Wang, J.; Yan, X.L.; Hu, J.H.; Gao, Z.P.; Jia, M.X.; Yu, X.; Wang, J. Study on the mechanism of arachnoid iridoids and terpenes on rats with irritable bowel syndrome. J. Beijing Univ. Tradit. Chin. Med. 2014, 37, 304–308. [Google Scholar]
- Modaressi, M.; Delazar, A.; Nazemiyeh, H.; Fathi-Azad, F.; Smith, E.; Rahman, M.; Gibbons, S.; Nahar, L.; Sarker, S.D. Antibacterial Iridoid Glucosides from Eremostachys laciniata. Phytother. Res. PTR 2009, 23, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Cimanga, K.; Kambu, K.; Tona, L.; Hermans, N.; Apers, S.; Totté, J.; Pieters, L.; Vlietinck, A.J. Antiamoebic Activity of Iridoids from Morinda morindoides Leaves. Planta Med. 2007, 72, 751–753. [Google Scholar] [CrossRef]
- Pungitore, C.R.; Ayub, M.J.; García, M. Iridoids as Allelochemicals and DNA Polymerase Inhibitors. J. Nat. Prod. 2004, 67, 357–361. [Google Scholar] [CrossRef]
- Akihisa, T.; Seino, K.I.; Kaneko, E.; Watanabe, K.; Tochizawa, S.; Fukatsu, M.; Banno, N.; Metori, K.; Kimura, Y. Melanogenesis inhibitory activities of iridoid-, hemiterpene-, and fatty acid-glycosides from the fruits of Morinda citrifolia (Noni). J. Oleo Sci. 2010, 59, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Yan, H.; Zhang, L.; Shan, M.; Chen, P.; Ding, A.; Li, S.F. A Review on the Phytochemistry, Pharmacology, and Pharmacokinetics of Amentoflavone, a Naturally-Occurring Biflavonoid. Molecules 2017, 22, 299. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Zhang, Y.; Guo, L.; Zhang, Q.; Zhu, H. Study on the Absorption Mechanism of Geniposide in the Chinese Formula Huang-Lian-Jie-Du-Tang in Rats. AAPS PharmSciTech. 2017, 18, 1382–1392. [Google Scholar] [CrossRef]
- Qu, K.; Zhao, L.; Luo, X.; Zhang, C.; Hou, P.; Bi, K.; Chen, X. An LC-MS method for simultaneous determination of five iridoids from Zhi-zi-chi Decoction in rat brain microdialysates and tissue homogenates: Towards an in depth study for its antidepressive activity. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2014, 965, 206–215. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, L.; Zheng, Z.; Pi, Z.; Liu, Z.; Song, F. Online microdialysis-ultra performance liquid chromatography-mass spectrometry method for comparative pharmacokinetic investigation on iridoids from Gardenia jasminoides Ellis in rats with different progressions of type 2 diabetic complications. J. Pharm. Biomed. Anal. 2017, 140, 146–154. [Google Scholar] [CrossRef]
- Han, H.; Yang, L.; Xu, Y.; Ding, Y.; Bligh, S.W.; Zhang, T.; Wang, Z. Identification of metabolites of geniposide in rat urine using ultra-performance liquid chromatography combined with electrospray ionization quadrupole time-of-flight tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2011, 25, 3339–3350. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.J.; Zhao, L.M. Serum Pharmacochemistry and Phamarcokinetics Study of Naoxuening Injection. Tradit. Chin. Drug Res. Clin. Pharmacol. 2008, 5, 52–55. [Google Scholar]
- Ramakrishna, R.; Bhateria, M.; Singh, R.; Puttrevu, S.K.; Bhatta, R.S. Plasma pharmacokinetics, bioavailability and tissue distribution of agnuside following peroral and intravenous administration in mice using liquid chromatography tandem mass spectrometry. J. Pharm. Biomed. Anal. 2016, 125, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.L.; Li, H.L.; He, J.C.; Feng, E.F.; Shi, P.P.; Liu, Y.Q.; Liu, C.X. Comparative pharmacokinetics of swertiamarin in rats after oral administration of swertiamarin alone, Qing Ye Dan tablets and co-administration of swertiamarin and oleanolic acid. J. Ethnopharmacol. 2013, 149, 49–54. [Google Scholar] [CrossRef]
- Wang, Z.G.; Wang, X.J.; Sun, H.; Chen, L.; Ma, C.M. Determination of novel nitrogen-containing metabolite after oral administration of swertiamarin to rats. J. Asian Nat. Prod. Res. 2012, 14, 176–181. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, S.; Sun, Y.; Wang, H.; Chen, G.; Wang, X.; Hattori, M.; Zhang, H. New analytical method for the study of the metabolism of gentiopicroside in rats after oral administration by LC-TOF-MS following picolinoyl derivatization. J. Sep. Sci. 2014, 37, 237–243. [Google Scholar] [CrossRef]
- Wu, X.; Tang, S.; Jin, Y.; Zhang, Y.; Hattori, M.; Wang, X.; Zhang, H.; Wang, Z. New analytical method for the study of metabolism of swertiamarin in rats after oral administration by UPLC-TOF-MS following DNPH derivatization. Biomed. Chromatogr. 2015, 29, 1184–1189. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, Z.; Li, S.; Ye, X.; Li, X.; He, K. Synergy effects of herb extracts: Pharmacokinetics and pharmacodynamic basis. Fitoterapia 2014, 92, 133–147. [Google Scholar] [CrossRef]
- Badal, S.; Williams, S.A.; Huang, G.; Francis, S.; Vedantam, P.; Dunbar, O.; Jacobs, H.; Tzeng, T.J.; Gangemi, J.; Delgoda, R. Cytochrome P450 1 enzyme inhibition and anticancer potential of chromene amides from Amyris plumieri. Fitoterapia 2011, 82, 230–236. [Google Scholar] [CrossRef]
- Wu, Y.; Ai, Y.; Wang, F.; Ma, W.; Bian, Q.; Lee, D.Y.; Dai, R. Simultaneous determination of four secoiridoid and iridoid glycosides in rat plasma by ultra performance liquid chromatography-tandem mass spectrometry and its application to a comparative pharmacokinetic study. Biomed. Chromatogr. 2016, 30, 97–104. [Google Scholar] [CrossRef]
- Yang, T.M.; Liu, Y.X.; Fu, H.Y.; Lan, W.; Su, H.B.; Tang, H.B.; Yin, Q.B.; Li, H.D.; Wang, L.P.; Wu, H.L. Pharmacokinetic Analysis of Four Bioactive Iridoid and Secoiridoid Glycoside Components of Radix Gentianae Macrophyllae and Their Synergistic Excretion by HPLC-DAD Combined with Second-Order Calibration. Nat. Prod. Bioprospect. 2017, 7, 445–459. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Xue, B.; Ma, B.; Zhang, Q.; Liu, M.; Liu, L.; Yao, D.; Qi, H.; Wang, Y.; Ying, H.; et al. A pre-clinical pharmacokinetic study in rats of three naturally occurring iridoid glycosides, Picroside-I, II and III, using a validated simultaneous HPLC-MS/MS assay. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2015, 993–994, 47–59. [Google Scholar] [CrossRef]
- Zahiruddin, S.; Khan, W.; Nehra, R.; Alam, M.J.; Mallick, M.N.; Parveen, R.; Ahmad, S. Pharmacokinetics and comparative metabolic profiling of iridoid enriched fraction of Picrorhiza kurroa—An Ayurvedic Herb. J. Ethnopharmacol. 2017, 197, 157–164. [Google Scholar] [CrossRef]
- Hou, Y.C.; Tsai, S.Y.; Lai, P.Y.; Chao, P.D.L. Metabolism and pharmacokinetics of genipin and geniposide in rats. Food Chem. Toxicol. 2008, 46, 2764–2769. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
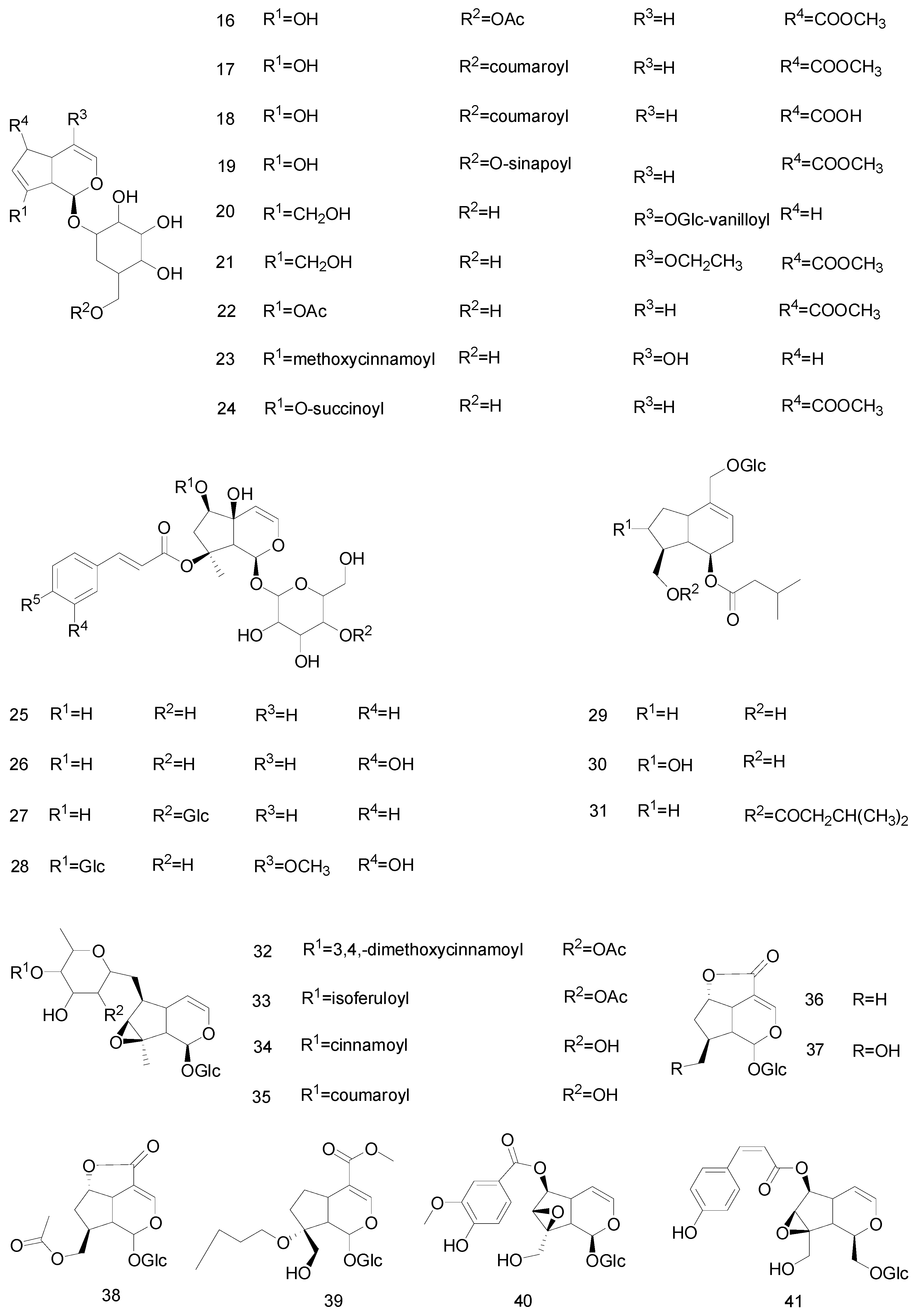{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
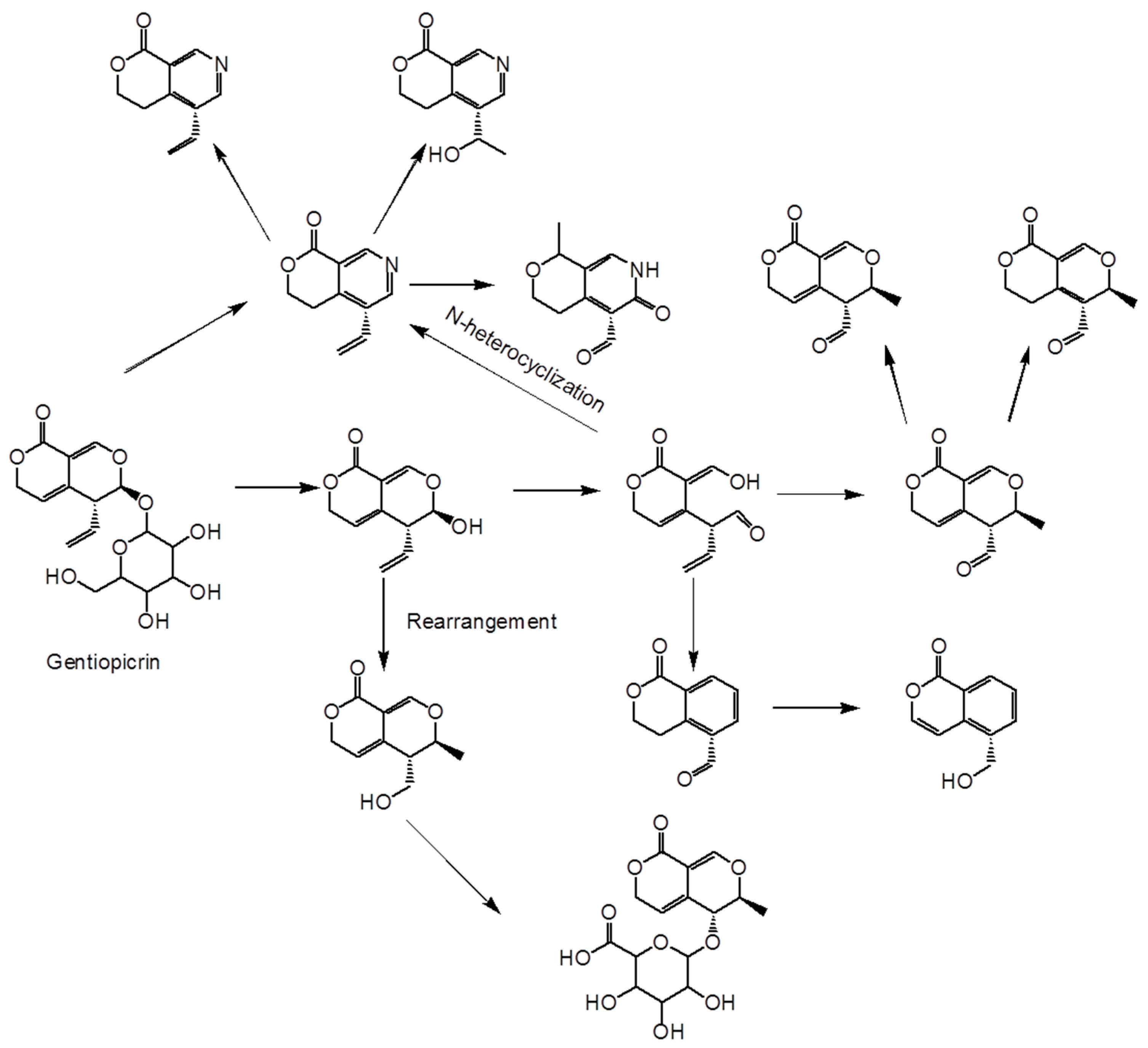{kind=link}
| No. | Classes | Compounds | Sources | Ref. |
|---|---|---|---|---|
| 1 | Iridoid glycosides | Euphroside | Scrophulariaceae | [13] |
| 2 | Plantarenaloside | Plantaginaceae, Bignoniaceae | [14,15] | |
| 3 | Geniposidic acid | Scrophulariaceae, Labiatae | [13] | |
| 4 | Loganic acid | Acanthaceae | [16] | |
| 5 | Loganic acid-6′-O-β-d-glucoside | Gentianaceae | [17] | |
| 6 | 2′-O-(4-methoxycinnamoyl) mussaenosidic acid | Acanthaceae | [18] | |
| 7 | 2′-O-trans-coumaroylshanzhiside | Rubiaceae | [19] | |
| 8 | 6′-O-trans-coumaroyl-shanzhiside | Rubiaceae | [19] | |
| 9 | Mussaenoside | Scrophulariaceae | [13] | |
| 10 | Lupulinoside | Acanthaceae | [20] | |
| 11 | 8-epideoxyloganic acid | Bignoniaceae | [14] | |
| 12 | Shanzhiside methyl ester | Acanthaceae | [20] | |
| 13 | Barlerin | Acanthaceae | [20] | |
| 14 | Acetylbarlerin | Acanthaceae | [20] | |
| 15 | 6-O-trans-p-coumaroyl-8-O-acetylshanzhi side methyl ester | Acanthaceae | [20] | |
| 16 | 6′-O-acetylgeniposide | Rubiaceae | [21] | |
| 17 | 6′-O-trans-p-coumaroylgeniposide | Rubiaceae | [21] | |
| 18 | 6′-O-trans-p-coumaroyl-geniposidic acid | Rubiaceae | [21] | |
| 19 | 6′-O-trans-sinapoylgeniposide | Rubiaceae | [21] | |
| 20 | Pinnatoside | Verbenaceae | [22] | |
| 21 | 6β-ethoxygeniposide | Rubiaceae | [19] | |
| 22 | 10-O-acetylgeniposide | Rubiaceae | [21] | |
| 23 | Eurostoside | Scrophulariaceae | [23] | |
| 24 | 10-O-succinoylgeniposide | Rubiaceae | [21] | |
| 25 | Ninpogenin | Scrophulariaceae | [24] | |
| 26 | 8-O-(coumaroyl)harpagide | Scrophulariaceae | [24] | |
| 27 | 6-O-α-d-galactopyranmosylharpagoside | Scrophulariaceae | [24] | |
| 28 | 8-O-feruloylharpagide | Scrophulariaceae | [24] | |
| 29 | Dihydropenstemide | Plantaginaceae | [15] | |
| 30 | Patrinalloside A | Valerianaceae | [25] | |
| 31 | 10-isovaleroyl-dihydropenstemide | Loganiaceae | [15] | |
| 32 | Buddlejoside A9 | Loganiaceae | [26] | |
| 33 | 6-O-α-L-(″-acetyl-4″-O-trans-isoferuloyl) rhamnopyranosyl catalpol | Loganiaceae | [26] | |
| 34 | 6-O-α-L-(4″-O-trans-cinnamoyl) rhamnopyranosyl catalpol | Loganiaceae | [26] | |
| 35 | 6-O-α-L-(4″-O-trans-p-coumaroyl) rham-nopyranosyl catalpol | Loganiaceae | [26] | |
| 36 | Brasoside | Verbenaceae | [27] | |
| 37 | Deacetyl asperuloside | Rubiaceae | [28] | |
| 38 | Asperuloside | Rubiaceae, Plantaginaceae | [28,29,30] | |
| 39 | 8α-butylgardenoside B | Rubiaceae | [19] | |
| 40 | Amphicoside | Scrophulariaceae | [31,32] | |
| 41 | Specioside | Bignoniaceae | [33] | |
| 42 | Secoiridoids | 6′-O-β-d-glucopyranosyl gentiopicroside | Gentianaceae | [34] |
| 43 | 3′-O-β-d-glucopyranosyl gentiopicroside | Gentianaceae | [34] | |
| 44 | 4′-O-β-d-glucopyranosyl gentiopicroside | Gentianaceae | [34] | |
| 45 | Oleoside dimethyl ester | Oleaceae | [35] | |
| 46 | (8E)-nuezhenide | Oleaceae | [35] | |
| 47 | (8Z)-nuezhenide | Oleaceae | [35] | |
| 48 | (8Z)-nuezhenide A | Oleaceae | [35] | |
| 49 | Gentiopicroside | Gentianaceae | [17,34,36,37,38] | |
| 50 | Oleuropein | Oleaceae | [35,39] | |
| 51 | Qinjiaosides B | Gentianaceae | [34] | |
| 52 | Qinjiaoside A | Gentianaceae | [34] | |
| 53 | 7-hydroxy eucommiol | Bignoniaceae | [33] | |
| 54 | 7-methoxydiderroside | Acanthaceae | [20] | |
| 55 | Isojaslanceoside B | Oleaceae | [40] | |
| 56 | 6′-O-trans-cinnamoyl-secologanoside | Oleaceae | [40] | |
| 57 | Nuezhenelenoliciside | Oleaceae | [40] | |
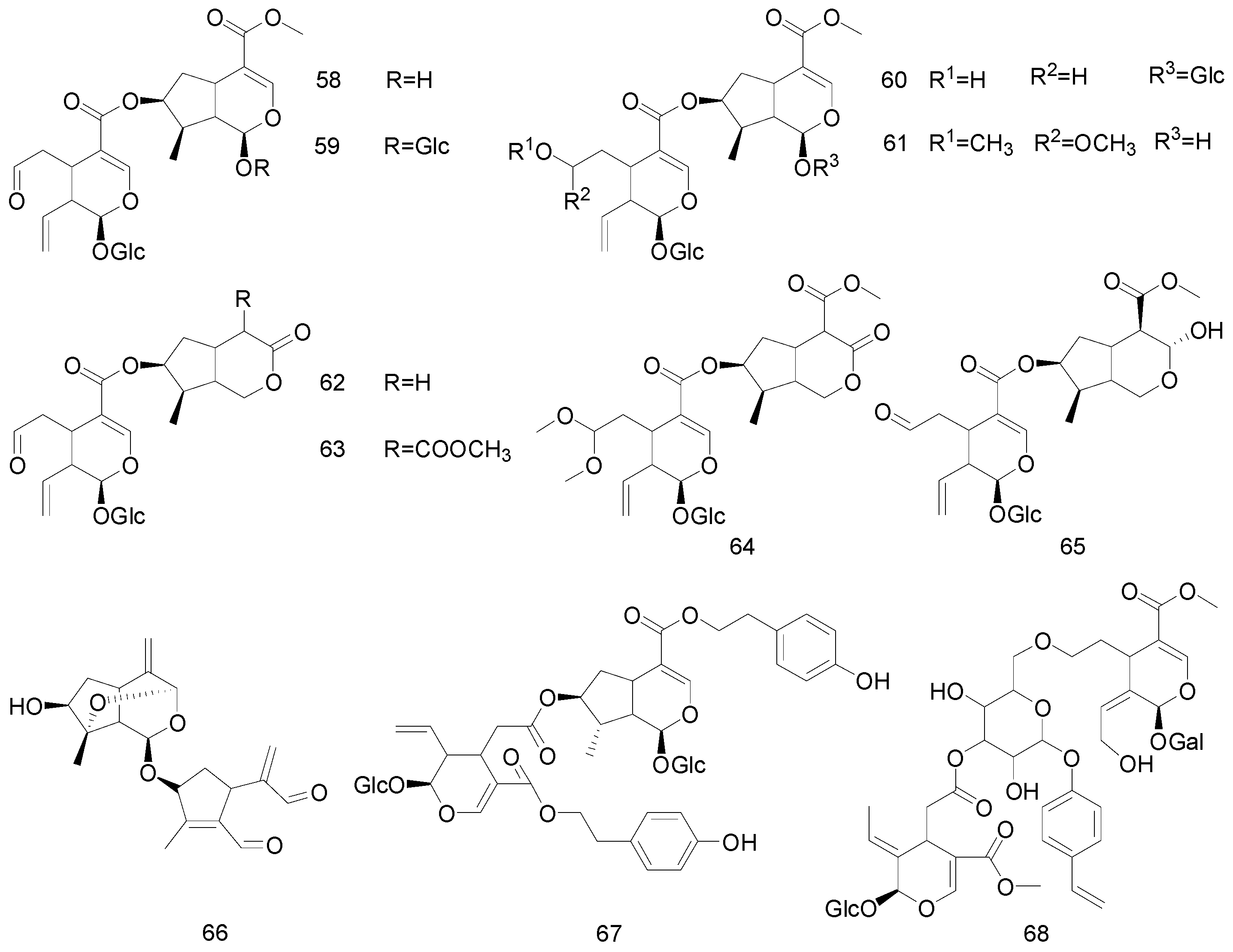
| 58 | Bis-iridoids | Sylvestroside III | Dipsacaceae | [41] |
| 59 | Cantleyosid | Dipsacaceae | [41] | |
| 60 | Sylvestroside I | Dipsacaceae | [41] | |
| 61 | Sylvestroside IV Dimethyl acetal | Dipsacaceae | [41] | |
| 62 | Laciniatoside II | Dipsacaceae | [41] | |
| 63 | Sylvestroside IV | Dipsacaceae | [41] | |
| 64 | sylvestroside III dimethyl acetal | Dipsacaceae | [41] | |
| 65 | Laciniatoside I | Dipsacaceae | [41] | |
| 66 | Polystachyn A | Valerianaceae | [42] | |
| 67 | Sclerochitonoside C | Acanthaceae | [43] | |
| 68 | Oleoside dimethyl ester | Oleaceae | [35] | |
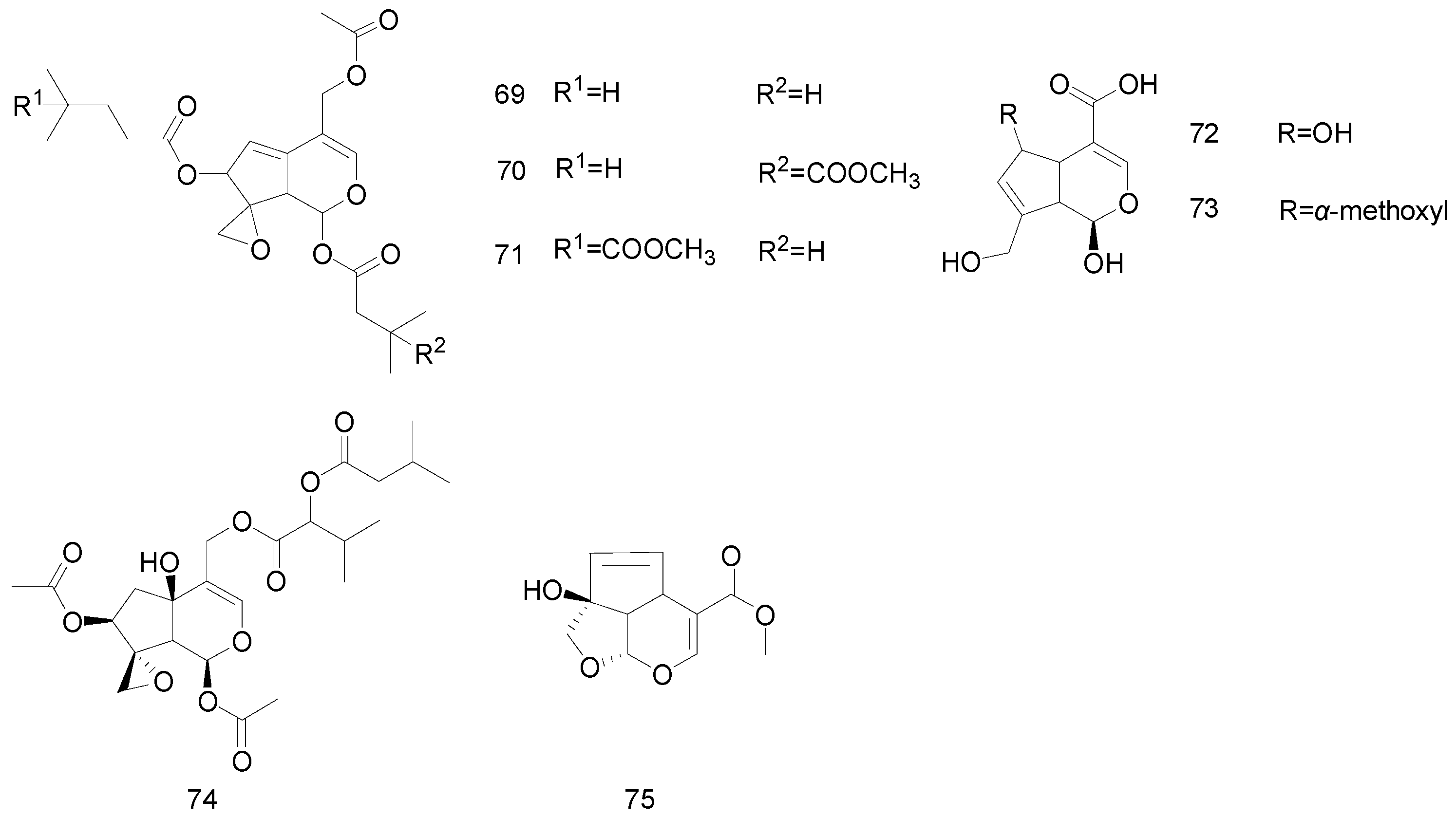
| 69 | Non-glycosidic iridoids | Valtrate | Valerianaceae | [44] |
| 70 | 1-β-acevaltrate | Valerianaceae | [44] | |
| 71 | Acevaltrate | Valerianaceae | [44] | |
| 72 | 6α-hydroxygenipi | Rubiaceae | [19] | |
| 73 | 6α-methoxygenipin | Rubiaceae | [19] | |
| 74 | IIHD-acevaltrate | Valerianaceae | [42] | |
| 75 | garjasmine | Rubiaceae | [19] |
| Function | Compounds | Dose | Model | Efficacy Evaluation | Ref. |
|---|---|---|---|---|---|
| Cardiocerebrovascular activity | Geniposide | 33.2 μg/mL | Brain microvascular endothelial cell (BMEC) with oxygen–glucose deprivation (OGD) | Declining the productions of IL-8, IL-1β and monocyte chemotactic protein 1 (MCP-1) | [87] |
| Digestive activity | Gentiopicroside | 20 mg/kg | Male rats (Sprague Dawley; six weeks old) | In all experimental groups an increase of gastric juice volume, total and free HCl concentration as well as pepsin concentration was observed. | [32] |
| Cholagogic effects | Genipin | 1 μmol/100 g·min | Male sprague dawley rats and hyperbilirubinemic rats | Enhanced the ability of hepatocytes to secrete independent bile salts | [88] |
| Antioxidant | Geniposide | 12.5, 25, 50 μg/mL | Human umbilical vein endothelial cell | Increased the activities of SOD, GSH-Px, NOS and NO production | [89] |
| Antithrombotic activity | Geniposide and genipin | 20, 40 mg/kg | Male ICR mice | Significantly prolonging the time required for thrombotic occlusion | [90] |
| Anti-senescence | Catalpol | 0.01, 0.1, 1 mg/mL | Human fibroblast Hs68 cell line irradiated by ultraviolet B (UVB) | Inhibited the formation of matrix metalloproteinase-1 (MMP-1) | [91] |
| Against intestinal ischemia/reperfusion (I/R) injury | Catalpol | 25, 50 mg/kg | In vivo intestinal I/R-injured rats | Significantly attenuated rat intestinal I/R injury by decreasing pro-inflammatory cytokines, reducing oxidative stress, and restoring intestinal barrier function | [92] |
| Anti-depressive | Genipin | 100 mg/kg | CUMS rat model | Decreases in serum trimetlylamine oxide (TMAO) and β-hydroxybutyric acid (β-HB) | [93] |
| Wound healing properties | Methylcatalpol | 50 mg/kg | Male rabbits model | Protective activity against increased skin vascular permeability | [94] |
| Antiallergic | 3,4-Dihydroxy catalpol | 30 mg/kg | Asthmatic mouse model | Exhibited an antiasthmatic effect by the suppression of elevated IgE, IL-4 and IL-13 levels and eosinophilia in the plasma | [95] |
| Analgesic activity | Bis-iridoid | 50, 100 mg/kg | Male mice | Bis-iridoid inhibited TNF-α-induced NF-κB-dependent promoter activity in a dose-dependent manner, The release of several proinflammatory cytokines and mediators (including TNF-a andPGE2) contributes to nociceptor sensitization and a reduction | [7] |
| Anti-HIV-1 activity | 2′-O-(4-Methoxycinnamoyl) mussaenosidic acid | 0.1 μg/mL | Real-time polymerase chain reaction (PCR) assay and HIV-1 p24 antigen kit | The expression level of C-C chemokine receptor type 5 (CCR5) and chemokine receptor type 4 (CXCR4) on CD4+ Tcellswere decreased in cells treated with this iridoid glycoside, demonstrated that this iridoid glycoside restricts HIV-1 replication on the early stage of HIV infection | [35] |
| Antiproliferative activities | Deacetyl asperuloside | 806.4 μg/mL | K562 chronic myelogenous leukemic cells | Significantly increased caspase 3 activity (p < 0.05) | [16] |
| AChE inhibitory | Lupulinoside | 134.0 μM | GST, AChE | Lupulinoside exhibited different levels of GST, AChE inhibitory | [41] |
| Anti-obesity | Genipin | 20 mg/kg | High-fat diet–fed obese mice | Regulating miR-142a-5p/SREBP-1c axis, led to the inhibition of lipogenesis | [96] |
| Anti-osteoporosis activity | Aucubin | 1, 2.5, 5 μM | MG63 cells | Improved osteoblast differentiation and enhanced the levels of BMP2 (bone morphogenetic proteins-2) in MG63 cells | [97] |
| Nuezhenelenoliciside | - | MC3T3-E1 cells | Increased the proliferation of pre-osteoblast MC3T3-E1 cells, possessed anti-osteoporosis activity | [11] | |
| Inhibition of gastric lesions | Genipin | 50, 100 mg/kg (p.o.) | HCl/ethanol-induced gastric lesions rat | Increasing ROS and ROS-induced NAPDH-oxidase (NOX) production and enabling gastric cancer cells to start the tumor cell apoptosis process via Egr1/p21 signaling pathway | [98] |
| Anti- Alzheimer’s disease | Gardenoside | - | Fruit-fly Alzheimer’s disease model induced by human Abeta protein over-expression | Suppressed the expression of immune-related genes in the brain | [99] |
| Anti-angiogenic | Geniposide | 25–100 μM | NIH3T3 cell line | Dose-dependently inhibiting the growth of the transformed N1H3T3 cell line | [100] |
| Inhibition of rheumatoid arthritis | Geniposide | 50 μM | MH7A fibroblast-like synoviocytes in patients with rheumatoid arthritis | Inhibition of TNF-α-stimulated cell proliferation and activation of the Ras-Erk1/2 pathway via upregulating miRNA-124a expression | [101] |
| α-Glucosidase inhibitory activities | Aldosecologanin | 0.5 mM | Lipopolysaccharide (LPS)-induced nitric oxide (NO) production and α-glucosidase reagent | Inhibiting α-glucosidase with IC50 values of 1.08 ± 0.70 and 0.62 ± 0.14 mM | [102] |
| Anti-anxiety properties | Geniposide | 20, 40 mg/kg | Male ddY mice 4 weeks of age | Increased the social interaction time and demonstrated to exert an anxiolytic effect in a dose- dependent manner | [103] |
| Antiviral | 6-O-trans-p-coumaroyl-8-O-acetylshanzhiside methyl ester | 42.2 μg/mL | The viral CPE assay | Have potent in vitro activity against respiratory syncytial virus (EC50 2.46 μg/mL, IC50 42.2 μg/mL) | [104] |
| Angiogenic properties | Aucubin | 10 mg/kg | Female C57BL/6 mice | Induced angiogenesis via vascular endothelial cell growth factor (VEGF)/Akt/endothelial nitric oxide synthase (eNOS) signaling pathway | [105] |
| Sedative effect | Total iridoids | 0.3, 0.6, 0.9 g/kg | Male mice | Reduce the number of autonomous activities, prolong the sleep time of mice, and strengthen the hypnotic effect of pentobarbital sodium | [106] |
| Anticonvulsant | Total iridoids | 1.0, 1.5, 2.0 g/kg | Pentetrazol-induced mice epilepsy model | Reduce the mortality of mice and prolong the latent period of convulsion | [106] |
| Improving irritable bowel syndrome | Total iridoids | 0.3 mg/kg | Irritable bowel syndrome model rats | Regulating TPH1 and MAO-A factor, reducing 5-HT expression in serum and visceral sensitivity in rats with irritable bowel syndrome | [107] |
| Antibacterial activity | Phloyoside I, Phlomiol, Pulchelloside I | - | 12 different strains | Exhibited from low to moderate levels (MIC = 0.05–0.50 mg/mL) of antibacterial activity | [108] |
| Antimalarial activity | Epoxygaertneroside, Methoxygaertneroside, Gaertneroside, Acetylgaertneroside, Gaertneric acid | 1.3, 2.3, 4.3, 5.4, and 7.1 mg/mL | MT-4 cells | Displayed antiamoebic activity with IC50 values of 1.3, 2.3, 4.3, 5.4, and 7.1 mg/mL | [109] |
| DNA polymerase inhibitory | Catalpol, 8-O-acetylharpagide, Harpagide | 47.8 mM | Adequate primer/template DNA and nucleotide analogues | Inhibition of tag DNA polymerase activity with an IC50 value of 47.8 mM | [110] |
| Anti-melanogenesis activity | 9-epi-6α–methoxy geniposidic acid, Asperulosidic acid, Deacetyl asperulosidic acid, Scandoside methyl ester | 100 mM | B16 melanoma cells induced by a-melanocyte-stimulating hormone (a-MSH) | Exhibited anti-melanogenesis activity with 40–50% reduction of melanin content at 100 mM | [111] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Gong, X.; Bo, A.; Zhang, L.; Zhang, M.; Zang, E.; Zhang, C.; Li, M. Iridoids: Research Advances in Their Phytochemistry, Biological Activities, and Pharmacokinetics. Molecules 2020, 25, 287. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25020287
Wang C, Gong X, Bo A, Zhang L, Zhang M, Zang E, Zhang C, Li M. Iridoids: Research Advances in Their Phytochemistry, Biological Activities, and Pharmacokinetics. Molecules. 2020; 25(2):287. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25020287
Chicago/Turabian StyleWang, Congcong, Xue Gong, Agula Bo, Lei Zhang, Mingxu Zhang, Erhuan Zang, Chunhong Zhang, and Minhui Li. 2020. "Iridoids: Research Advances in Their Phytochemistry, Biological Activities, and Pharmacokinetics" Molecules 25, no. 2: 287. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25020287