AT-MSCs Antifibrotic Activity is Improved by Eugenol through Modulation of TGF-β/Smad Signaling Pathway in Rats



Abstract
:1. Introduction
2. Results
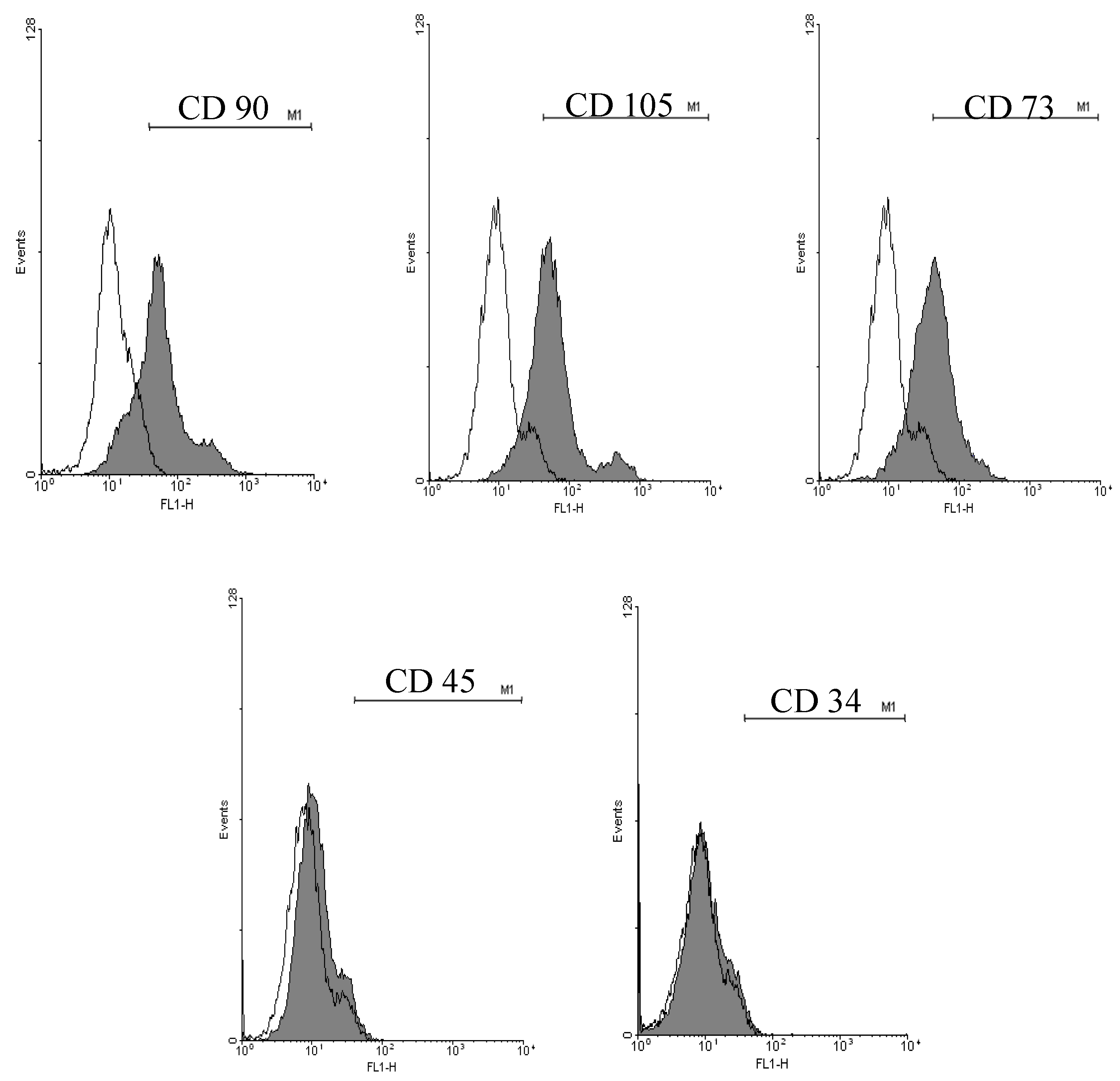
2.1. Cell Surface Markers Expression
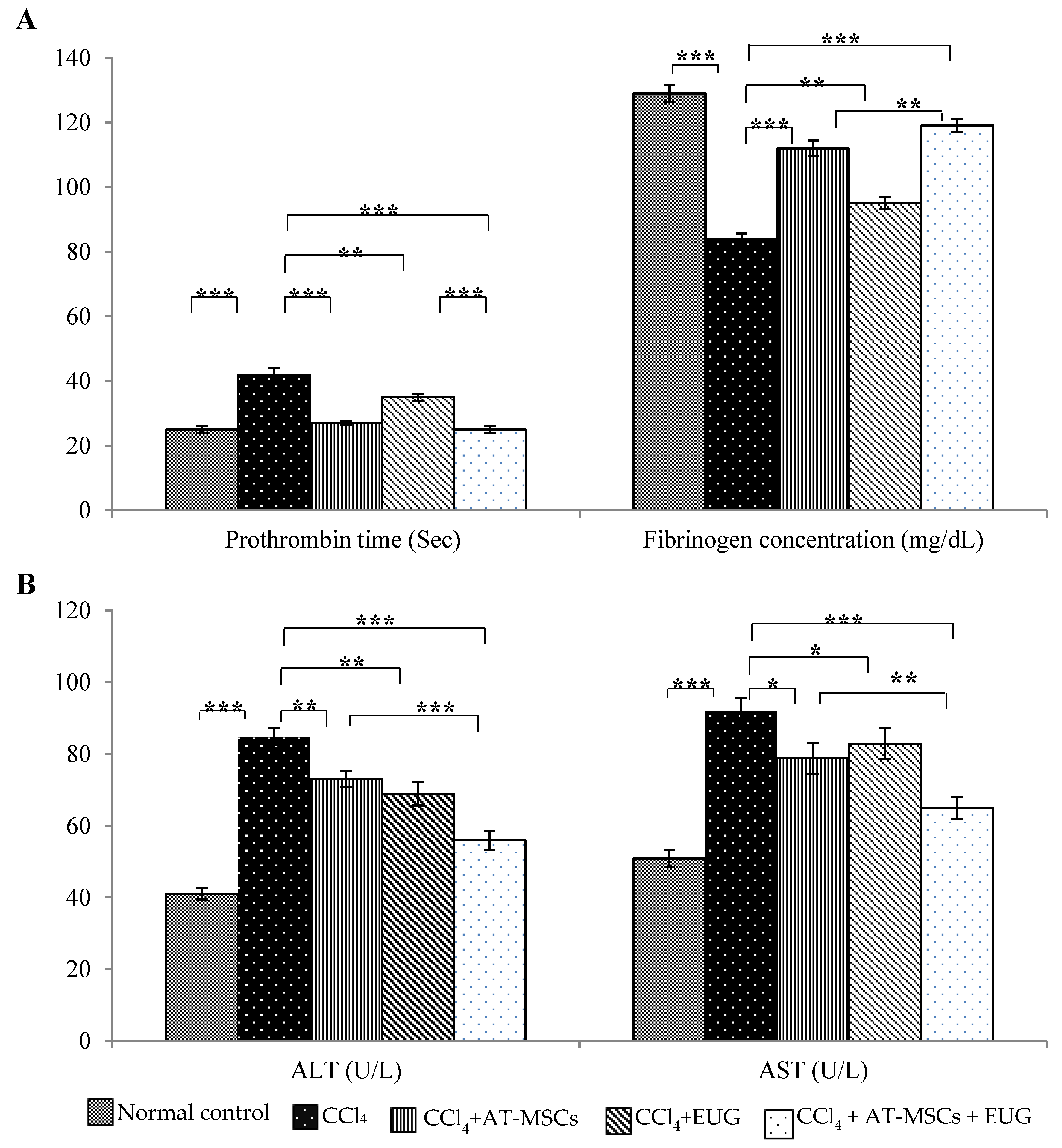
2.2. Prothrombin Time and Fibrinogen Concentration Assay
2.3. Liver Enzymes (ALT and AST) Levels
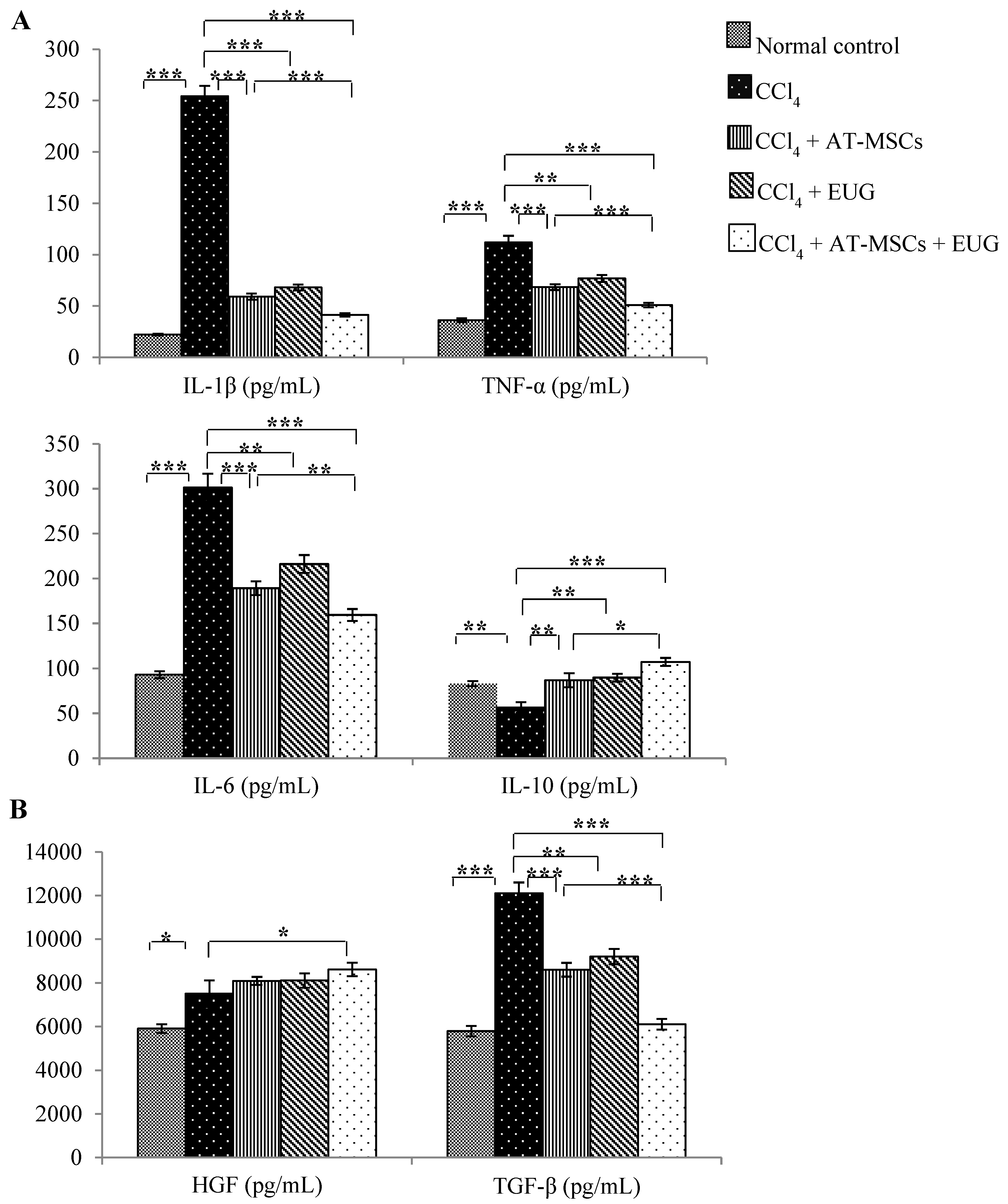
2.4. Serum Inflammatory Cytokines (IL-1β, TNF-α, IL-6 and IL-10) Levels
2.5. Serum Growth Factors (Hepatocyte Growth Factor (HGF) and Transforming Growth Factor-Beta (TGF-β)) Levels
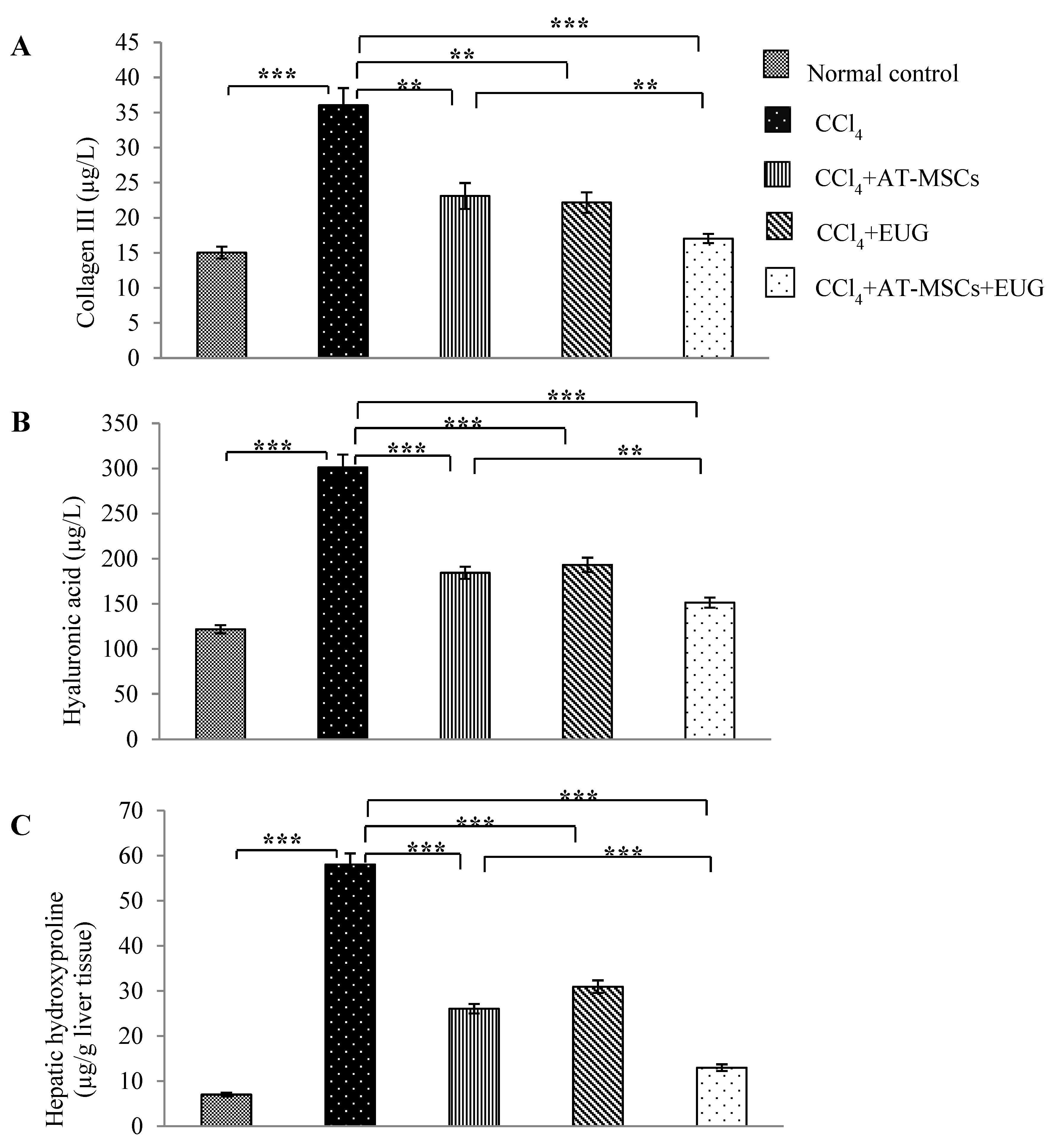
2.6. Levels of Type III Collagen, Hyaluronic Acid and Hepatic Hydroxyproline Content
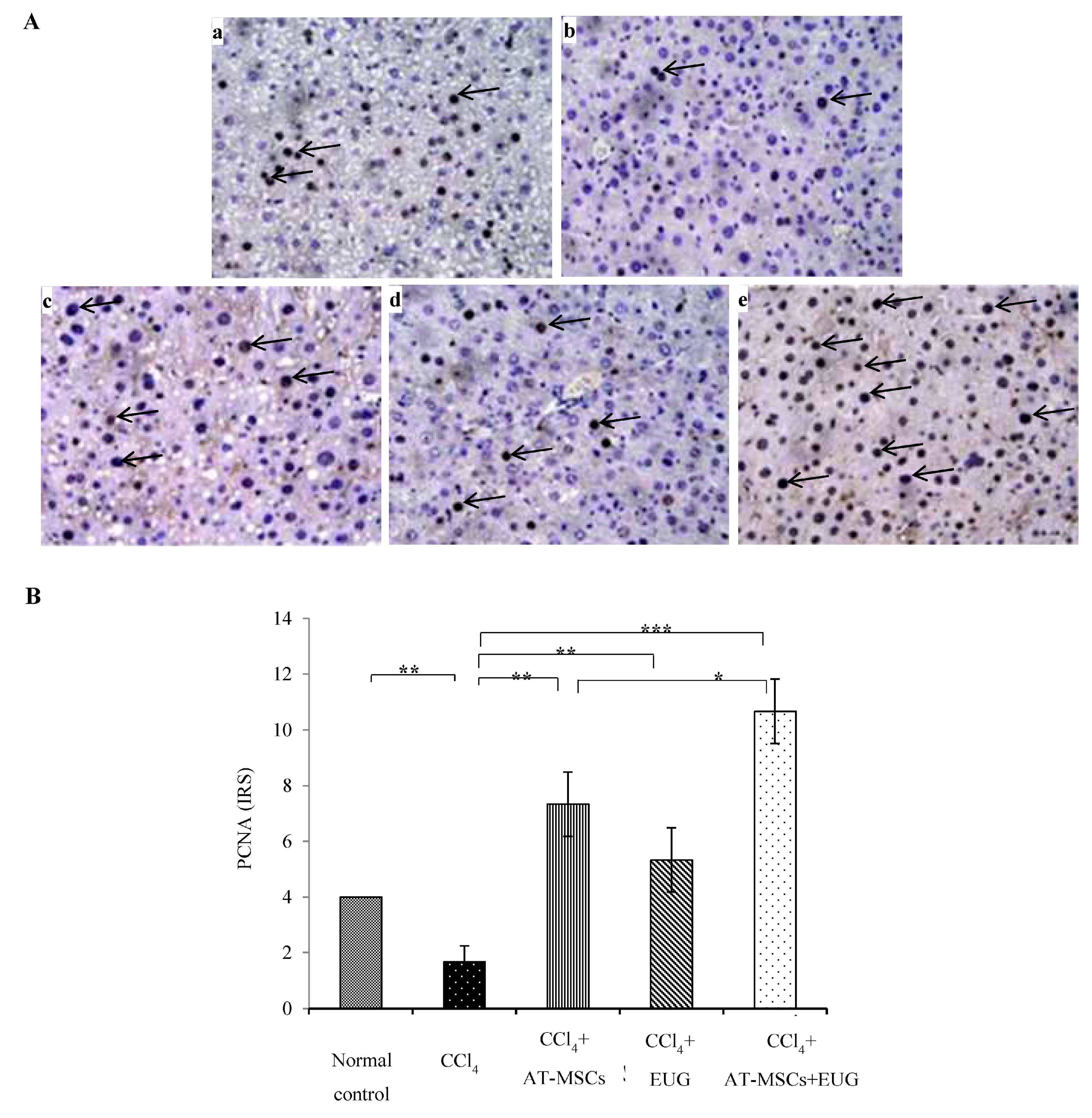
2.7. Expression of Proliferating Cell Nuclear Antigen (PCNA)
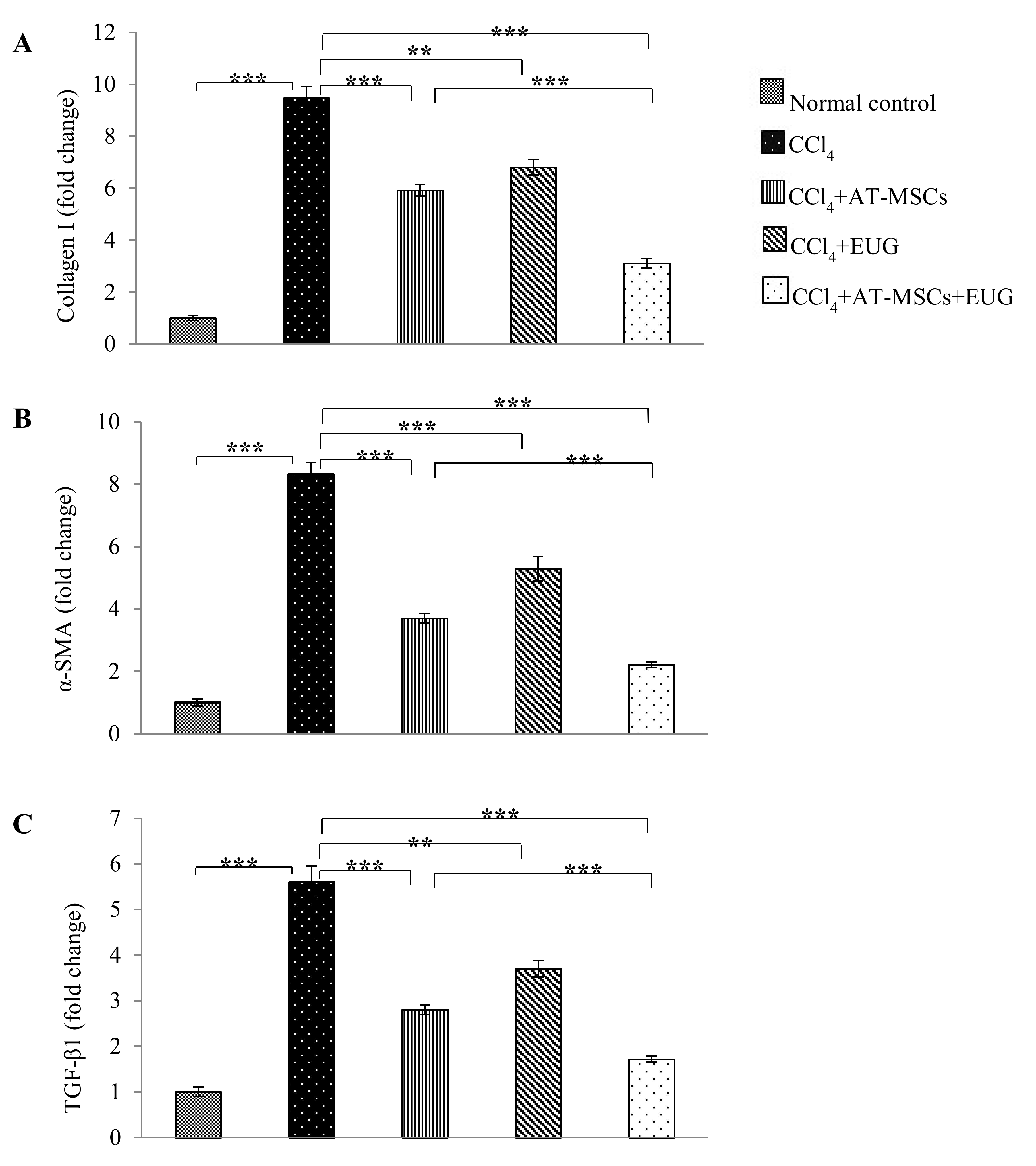
2.8. Type 1 Collagen, α-Smooth Muscle Actin (α-SMA) and TGF-β1 Genes Expression
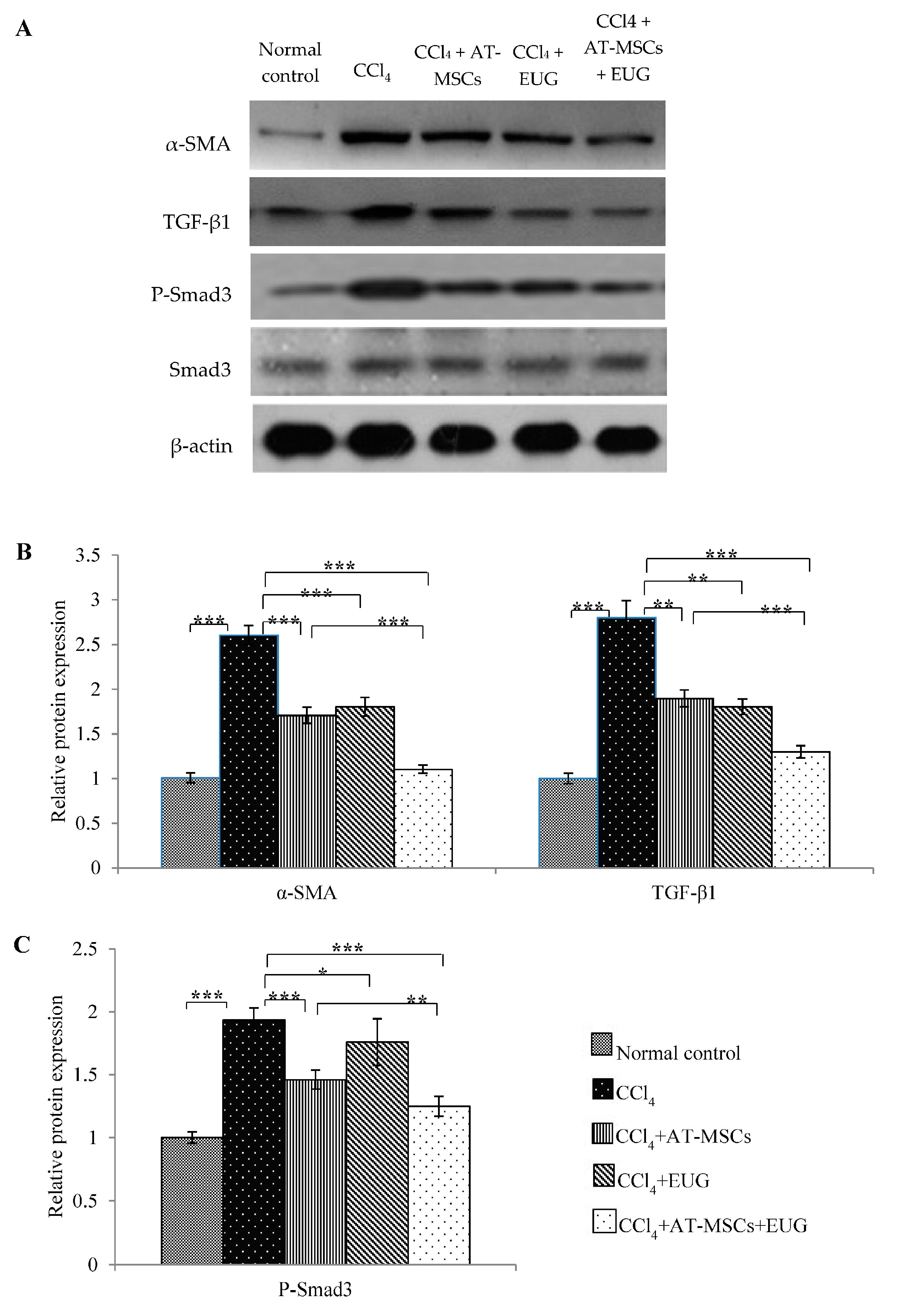
2.9. Expression of α-SMA, TGF-β1 and Phospho-Smad3 Proteins
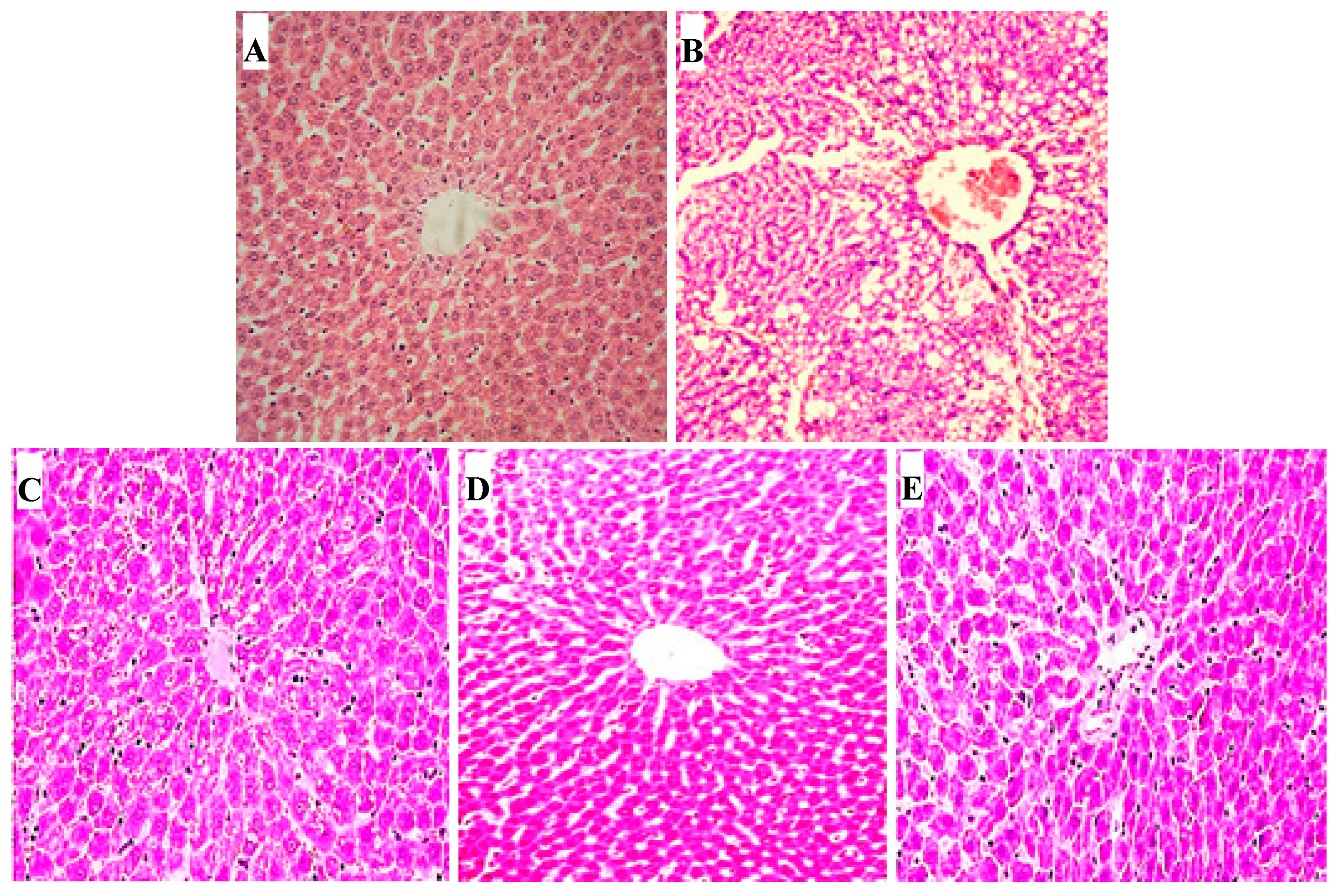
2.10. Hepatic Histopathological Changes
3. Discussion
4. Materials and Methods
4.1. Drugs and Reagents
4.2. Isolation, Culture and Characterization of AT-MSCs
4.3. Animals and the Experimental Design
4.4. Prothrombin Time and Fibrinogen Concentration Assay
4.5. Assay of Serum Liver Enzymes, Inflammatory Cytokines and Growth Factors Levels
4.6. Measurement of Serum Levels of Type III Collagen and Hyaluronic Acid and Hepatic Hydroxyproline Content
4.7. Proliferating Cell Nuclear Antigen Expression Determination
4.8. Quantitative Real-Time Polymerase Chain Reaction
4.9. Western Blotting Analysis for Hepatic α-SMA, TGF-β1, Smad3 and Phospho-Smad3 Proteins Expression
4.10. Histopathological Examination
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Koike, C.; Zhou, K.; Takeda, Y.; Fathy, M.; Okabe, M.; Yoshida, T.; Nakamura, Y.; Kato, Y.; Nikaido, T. Characterization of amniotic stem cells. Cell. Reprogram. 2014, 16, 298–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagura, S.; Otaka, S.; Koike, C.; Okabe, M.; Yoshida, T.; Fathy, M.; Fukahara, K.; Yoshimura, N.; Misaki, T.; Nikaido, T. Effect of exogenous Oct4 overexpression on cardiomyocyte differentiation of human amniotic mesenchymal cells. Cell. Reprogram. 2013, 15, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Otaka, S.; Nagura, S.; Koike, C.; Okabe, M.; Yoshida, T.; Fathy, M.; Yanagi, K.; Misaki, T.; Nikaido, T. Selective isolation of nanog-positive human amniotic mesenchymal cells and differentiation into cardiomyocytes. Cell. Reprogram. 2013, 15, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Parekkadan, B.; Milwid, J.M. Mesenchymal stem cells as therapeutics. Annu. Rev. Biomed. Eng. 2010, 12, 87–117. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Zhou, X.; Shi, Y.; Li, J.; Zheng, L.; Cui, L.; Zhang, J.; Wang, L.; Han, Z.; Han, Y.; et al. In vivo tracking and comparison of the therapeutic effects of MSCs and HSCs for liver injury. PLoS ONE 2013, 8, e62363. [Google Scholar] [CrossRef]
- Jang, Y.O.; Cho, M.Y.; Yun, C.O.; Baik, S.K.; Park, K.S.; Cha, S.K.; Chang, S.J.; Kim, M.Y.; Lim, Y.L.; Kwon, S.O. Effect of Function-Enhanced Mesenchymal Stem Cells Infected with Decorin-Expressing Adenovirus on Hepatic Fibrosis. Stem Cells Transl. Med. 2016, 5, 1247–1256. [Google Scholar] [CrossRef] [Green Version]
- Kern, S.; Eichler, H.; Stoeve, J.; Kluter, H.; Bieback, K. Comparative analysis of mesenchymal stem cells from bone marrow, umbilical cord blood, or adipose tissue. Stem Cells 2006, 24, 1294–1301. [Google Scholar] [CrossRef]
- Lotfy, A.; Salama, M.; Zahran, F.; Jones, E.; Badawy, A.; Sobh, M. Characterization of mesenchymal stem cells derived from rat bone marrow and adipose tissue: A comparative study. Int. J. Stem Cells 2014, 7, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Terai, S.; Ishikawa, T.; Omori, K.; Aoyama, K.; Marumoto, Y.; Urata, Y.; Yokoyama, Y.; Uchida, K.; Yamasaki, T.; Fujii, Y.; et al. Improved liver function in patients with liver cirrhosis after autologous bone marrow cell infusion therapy. Stem Cells 2006, 24, 2292–2298. [Google Scholar] [CrossRef]
- De Luna-Saldivar, M.M.; Marino-Martinez, I.A.; Franco-Molina, M.A.; Rivera-Morales, L.G.; Alarcon-Galvan, G.; Cordero-Perez, P.; Rojas-Martinez, A.; Rodriguez-Padilla, C.; Munoz-Espinosa, L.E. Advantages of adipose tissue stem cells over CD34(+) mobilization to decrease hepatic fibrosis in Wistar rats. Ann. Hepatol. 2019, 18, 620–626. [Google Scholar] [CrossRef]
- Bataller, R.; Brenner, D.A. Liver fibrosis. J. Clin. Investig. 2005, 115, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.L. Hepatic stellate cells: Protean, multifunctional, and enigmatic cells of the liver. Physiol. Rev. 2008, 88, 125–172. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Cen, P.; Chen, J.; Fan, L.; Li, J.; Cao, H.; Li, L. Role of mesenchymal stem cells, their derived factors, and extracellular vesicles in liver failure. Stem Cell Res. 2017, 8, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fathy, M.; Khalifa, E.; Fawzy, M.A. Modulation of inducible nitric oxide synthase pathway by eugenol and telmisartan in carbon tetrachloride-induced liver injury in rats. Life Sci. 2019, 216, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Saad Eldien, H.M.; Abdel-Aziz, H.O.; Sayed, D.; Mubarak, W.; Hareedy, H.H.G.; Mansor, S.G.; Yoshida, T.; Fathy, M. Periostin expression and characters of human adipose tissue-derived mesenchymal stromal cells were aberrantly affected by in vitro cultivation. Stem Cell Investig. 2019, 6, 33. [Google Scholar] [CrossRef]
- Higuchi, O.; Okabe, M.; Yoshida, T.; Fathy, M.; Saito, S.; Miyawaki, T.; Nikaido, T. Stemness of human Wharton’s jelly mesenchymal cells is maintained by floating cultivation. Cell. Reprogram. 2012, 14, 448–455. [Google Scholar] [CrossRef]
- Zhou, K.; Koike, C.; Yoshida, T.; Okabe, M.; Fathy, M.; Kyo, S.; Kiyono, T.; Saito, S.; Nikaido, T. Establishment and characterization of immortalized human amniotic epithelial cells. Cell. Reprogram. 2013, 15, 55–67. [Google Scholar] [CrossRef] [Green Version]
- Kharaziha, P.; Hellstrom, P.M.; Noorinayer, B.; Farzaneh, F.; Aghajani, K.; Jafari, F.; Telkabadi, M.; Atashi, A.; Honardoost, M.; Zali, M.R.; et al. Improvement of liver function in liver cirrhosis patients after autologous mesenchymal stem cell injection: A phase I-II clinical trial. Eur. J. Gastroenterol. Hepatol. 2009, 21, 1199–1205. [Google Scholar] [CrossRef]
- Kim, G.; Eom, Y.W.; Baik, S.K.; Shin, Y.; Lim, Y.L.; Kim, M.Y.; Kwon, S.O.; Chang, S.J. Therapeutic Effects of Mesenchymal Stem Cells for Patients with Chronic Liver Diseases: Systematic Review and Meta-analysis. J. Korean Med. Sci. 2015, 30, 1405–1415. [Google Scholar] [CrossRef] [Green Version]
- Fathy, M.; Fawzy, M.A.; Hintzsche, H.; Nikaido, T.; Dandekar, T.; Othman, E.M. Eugenol Exerts Apoptotic Effect and Modulates the Sensitivity of HeLa Cells to Cisplatin and Radiation. Molecules 2019, 24, 3979. [Google Scholar] [CrossRef] [Green Version]
- Ryeom, H.K.; Kim, S.H.; Kim, J.Y.; Kim, H.J.; Lee, J.M.; Chang, Y.M.; Kim, Y.S.; Kang, D.S. Quantitative evaluation of liver function with MRI Using Gd-EOB-DTPA. Korean J. Radiol. 2004, 5, 231–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, G.; Mohsin, S.; Khan, M.; Nasir, G.A.; Shams, S.; Khan, S.N.; Riazuddin, S. Nitric oxide augments mesenchymal stem cell ability to repair liver fibrosis. J. Transl. Med. 2012, 10, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.H.; Hwang, S.M. Cytokine interactions in mesenchymal stem cells from cord blood. Cytokine 2005, 32, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Huang, S.; Enhe, J.; Ma, K.; Yang, S.; Sun, T.; Fu, X. Bone marrow-derived mesenchymal stem cell attenuates skin fibrosis development in mice. Int. Wound J. 2014, 11, 701–710. [Google Scholar] [CrossRef]
- Dooley, S.; ten Dijke, P. TGF-beta in progression of liver disease. Cell Tissue Res. 2012, 347, 245–256. [Google Scholar] [CrossRef] [Green Version]
- Inagaki, Y.; Higashiyama, R.; Higashi, K. Novel antifibrotic modalities for liver fibrosis: Molecular targeting and regenerative medicine in fibrosis therapy. J. Gastroenterol. Hepatol. 2012, 27 (Suppl. 2), 85–88. [Google Scholar] [CrossRef]
- Rosenbloom, J.; Mendoza, F.A.; Jimenez, S.A. Strategies for antifibrotic therapies. Biochim. Biophys. Acta 2013, 1832, 1088–1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albright, C.D.; Borgman, C.; Craciunescu, C.N. Activation of a caspase-dependent oxidative damage response mediates TGFbeta1 apoptosis in rat hepatocytes. Exp. Mol. Pathol. 2003, 74, 256–261. [Google Scholar] [CrossRef]
- Wang, G.; Li, Z.; Li, H.; Li, L.; Li, J.; Yu, C. Metabolic Profile Changes of CCl(4)-Liver Fibrosis and Inhibitory Effects of Jiaqi Ganxian Granule. Molecules 2016, 21, 698. [Google Scholar] [CrossRef] [Green Version]
- Aithal, A.P.; Bairy, L.K.; Seetharam, R.N.; Kumar, N. Hemostatic potential of human bone marrow-derived mesenchymal stromal cells in Wistar rats with carbon tetrachloride induced liver cirrhosis. Stem. Cell. Investig. 2018, 5, 21. [Google Scholar] [CrossRef] [PubMed]
- Remmele, W.; Stegner, H.E. Recommendation for uniform definition of an immunoreactive score (IRS) for immunohistochemical estrogen receptor detection (ER-ICA) in breast cancer tissue. Pathologe 1987, 8, 138–140. [Google Scholar] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A (Percentage of Positive Cells) | B (Intensity of Staining) | IRS Score (A × B) |
|---|---|---|
| 0 = no positive cells | 0 = no color reaction | 0–1 = negative |
| 1 = <10% of positive cells | 1 = mild reaction | 2–3 = mild |
| 2 = 10%–50% positive cells | 2 = moderate reaction | 4–8 = moderate |
| 3 = 51%–80% positive cells | 3 = intense reaction | 9–12 = strongly positive |
| 4 = >80% positive cells |
| Primer | Sequence of the Primer |
|---|---|
| type 1 collagen | Forward: 5′-CAT GTT CAG CTT TGT GGA CCT-3′ |
| Reverse: 5′-GCA GCT GAC TTC AGG GAT GT-3′. | |
| α-SMA | Forward: 5′-CGA TAG AAC ACG GCA TCA TCA C-3′ |
| Reverse: 5′-GCA TAG CCC TCA TAG ATA GGC A-3′. | |
| TGF-β1 | Forward: 5′-GGA CTC TCC ACC TGC AAG AC-3′ |
| Reverse: 5′-GAC TGG CGA GCC TTA GTT TG-3′. | |
| GAPDH | Forward: 5′-AGA CAG CCG CAT CTT CTT GT-3′ |
| Reverse: 5′-TGA TGG CAA CAA TGT CCA CT-3′. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fathy, M.; Okabe, M.; Saad Eldien, H.M.; Yoshida, T. AT-MSCs Antifibrotic Activity is Improved by Eugenol through Modulation of TGF-β/Smad Signaling Pathway in Rats. Molecules 2020, 25, 348. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25020348
Fathy M, Okabe M, Saad Eldien HM, Yoshida T. AT-MSCs Antifibrotic Activity is Improved by Eugenol through Modulation of TGF-β/Smad Signaling Pathway in Rats. Molecules. 2020; 25(2):348. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25020348
Chicago/Turabian StyleFathy, Moustafa, Motonori Okabe, Heba M. Saad Eldien, and Toshiko Yoshida. 2020. "AT-MSCs Antifibrotic Activity is Improved by Eugenol through Modulation of TGF-β/Smad Signaling Pathway in Rats" Molecules 25, no. 2: 348. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25020348