
Selected Fungal Natural Products with Antimicrobial Properties


1
Institute of Bioorganic Chemistry, Polish Academy of Sciences, 61-704 Poznań, Poland
2
Department of Agriculture, Horticulture and Engineering Science, Scotland’s Rural College (SRUC), Edinburgh EH9 3JG, UK
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Molecules 2020, 25(4), 911; https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25040911
Submission received: 31 January 2020
/
Revised: 13 February 2020
/
Accepted: 14 February 2020
/
Published: 18 February 2020
(This article belongs to the Special Issue Antimicrobial Properties of Natural Products)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Fungal natural products and their effects have been known to humankind for hundreds of years. For example, toxic ergot alkaloids produced by filamentous fungi growing on rye poisoned thousands of people and livestock throughout the Middle Ages. However, their later medicinal applications, followed by the discovery of the first class of antibiotics, penicillins and other drugs of fungal origin, such as peptidic natural products, terpenoids or polyketides, have altered the historically negative reputation of fungal “toxins”. The development of new antimicrobial drugs is currently a major global challenge, mainly due to antimicrobial resistance phenomena. Therefore, the structures, biosynthesis and antimicrobial activity of selected fungal natural products are described here.
1. Introduction
Natural products (NPs) are a very rich source of antimicrobial drugs. They constitute more than two-thirds of clinically used antibiotics and half of anticancer drugs [1]. Molecules biosynthesised by fungi are a diverse and useful group of NPs. Plant endophytic and pathogenic fungi produce many secondary metabolites that play important roles in virulence and competition against other microbes. Due to their broad-spectrum activity, some of these NPs can also exhibit high biocidal activity against human pathogenic microbes. In recent years, marine fungi have emerged as a novel source of fungal NPs and may be a potential game changer in drug discovery; however, marine fungi still constitute an underrepresented resource of diverse NPs.
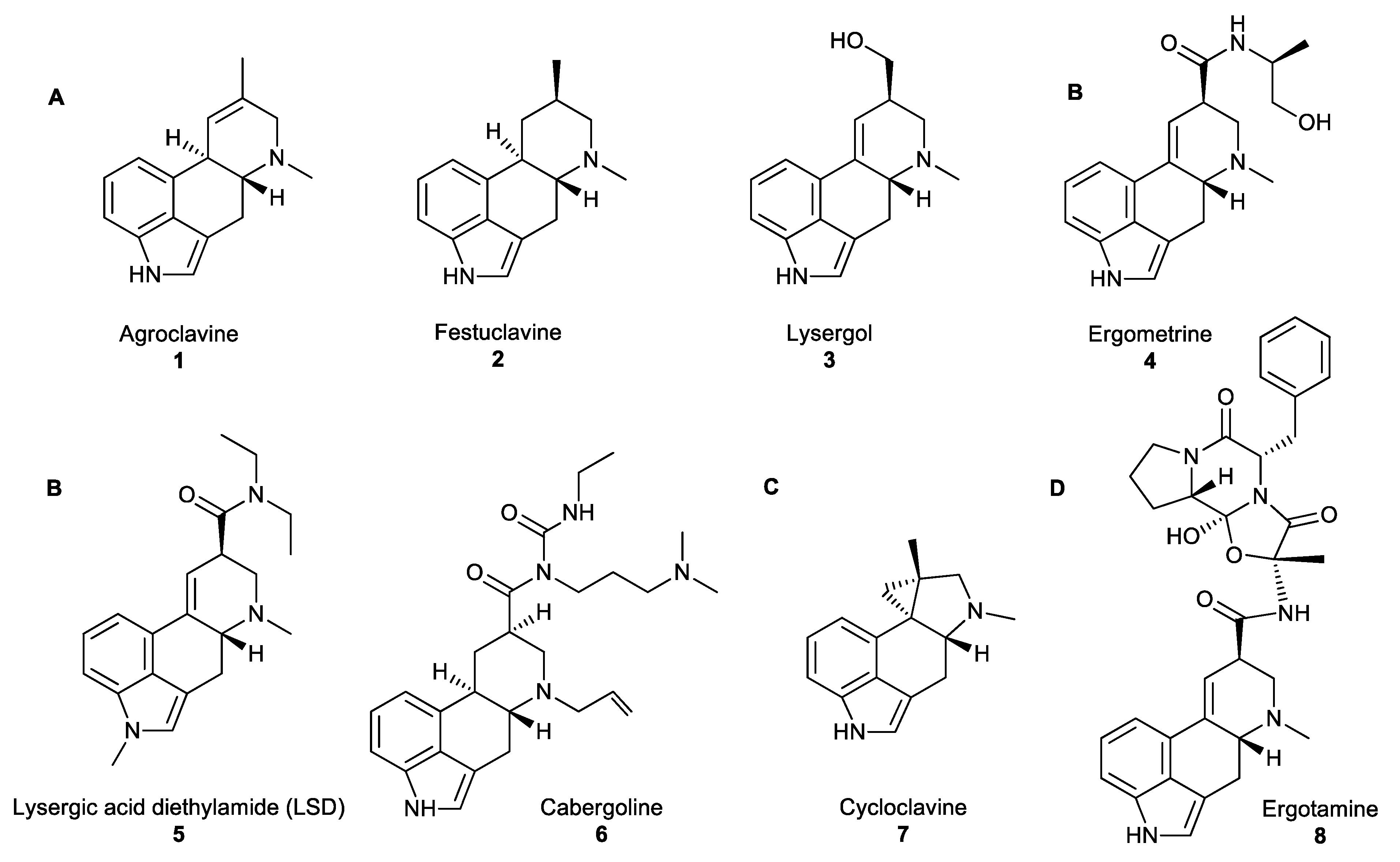
Fungal NPs have an important place in human history. For instance, ergot alkaloids (EAs) which are produced by the filamentous fungi of the genus Claviceps, have been referenced in ancient historical texts. References to grain diseases have been found in the Bible, in the Old Testament (850–550 BC). In the Middle Ages, the first reported ergotism epidemic was recorded in 944–1000 AD when almost half the population of the Aquitaine region of France (about 60,000 people) died of ergot poisoning [2,3]. The gangrenous form of the disease (medically known under the name Ergotismus gangraenosus) was commonly known as ‘‘ergotism’’, ‘‘holy fire’’ or ‘‘St. Anthony’s fire’’. Symptoms include delirium, hallucinations, muscle spasms, convulsions and gangrene of the limbs. The gruesome history of ergots has overshadowed their beneficial medicinal properties. However, the use of ergots as medicinal compounds was first documented in 1582, as they were administered for ‘‘quickening childbirth’’. Further research and screening of ergot analogues for oxytocic drugs that stimulate uterine contractions to hasten childbirth resulted, in 1938, in the synthesis of lysergic acid diethylamide (LSD) 5 (Figure 1B), a hallucinogenic compound that has become infamous for its use as an illicit “recreational drug” [4]. Currently, EAs are the inspiration for numerous semi-synthetic derivatives, such as cabergoline 6 or ergotamine 8 (Figure 1B,D, respectively) that have been applied in a wide range of medicinal treatments, such as the treatment of migraines, Parkinson’s disease, reduction of tumour growth, and other lesser-known synergistic antimicrobial activities.
Another example of the historical importance of fungal NPs was the discovery of penicillin 32 by Sir Alexander Fleming in 1928. The identification of this first antibiotic compound from the mould Penicillium notatum was a breakthrough and a revolution in drug discovery [5]. Ever since penicillin was discovered, a completely new era of chemotherapy started, thereby changing the quality of human life. The importance of this life-saving discovery cannot be understated, as penicillin was used to cure countless people of bacterial diseases. The development of penicillin initiated the golden era of natural antibiotics. The search for bioactive NPs led to the discovery of a multitude of antibacterial compounds, many of which were isolated from Streptomyces species [6]. The genus Streptomyces regroups species of actinobacteria that share morphological traits with fungi, such as filamentous hyphae and spore production. To date, over 350 agents derived from diverse chemical classes of metabolites isolated from Streptomyces have reached the world market as antimicrobial compounds [7]. These bioactive agents include NPs, semi-synthetic antibiotics and synthetic compounds [7,8]. However, the wide use of antibiotics has resulted in the development of resistant microbes due to the evolutionary selective pressure driven by antibiotics [9]. The number of effective therapeutics against life-threatening bacterial and fungal infections has fallen dramatically because of emerging multidrug-resistant (MDR) pathogens.
Antimicrobial resistance (AMR) is a major concern of modern medicine and it has now become one of the key research areas of the European Union Commission [10]. AMR occurs when microbes, such as bacteria, fungi, viruses and parasites acquire resistance to one or more drugs. Drug resistance is the biggest obstacle to success during the treatment of infectious diseases, and has been observed following the introduction of numerous antimicrobial agents into clinical practice. It is difficult to quantify the global human burden posed by AMR but, in the European Union alone, 25,000 people die every year due to drug-resistant bacterial infections [10]. Resistance to antimicrobial compounds also has a major impact on food production worldwide. Since the Green Revolution (ca. 1950–1970), food production and agriculture has been reliant on chemical input to control pathogenic microbes, either in animal or plant production [11]. However, the overuse of these chemicals, combined with the lack of diversity in their modes of action, has driven the appearance of resistance to these compounds. As a result, the control of pathogenic microbes has become increasingly difficult in the past few decades, contributing to the increased volatility of food production and food insecurity. Drug resistance is driving the constant need for new drug discoveries. To manage the risk of resistance to antimicrobial compounds arising, efforts are being made to better understand the mechanisms underlying host-microbe interactions, pathogen population evolution and drug modes of action. Herein, the structures, biosynthesis and antimicrobial activities of selected natural products derived from fungi are presented.
2. Selected Examples of Antimicrobial Natural Products from Fungi
2.1. Ergot Alkaloids: Fungal Natural Products Derived from Amino Acids
All naturally occurring EAs share a common tetracyclic scaffold, the so-called ‘‘ergoline scaffold”, derived from L-tryptophan. EAs are divided into three major classes based on the substituents decorating this scaffold: clavines (festuclavine and agroclavine derivatives), simple lysergic acid derivatives and ergopeptides (Figure 1A,B,D, accordingly) [12]. Clavines include the partially or fully saturated ring species D, such as agroclavine 1, festuclavine 2 or lysergol 3 (Figure 1A). Simple lysergic acid derivatives consist of the basic D-lysergic acid structure as an alkyl amide (Figure 1B), and ergopeptides also based on D-lysergic acid and a cyclic tripeptide moiety (Figure 1D). Cycloclavine 7 is a newly characterised ergot alkaloid which has been reproduced in vitro, and has an unusual ring system, where ring D has been transformed into a new five- and three-membered ring fusion [13].
EAs are produced by fungi occupying distinct ecological niches. Clavicipitaceous species, such as Claviceps purpurea and Neotyphodium lolii from the order Eurotiales are plant pathogenic and symbiotic fungi, respectively. Aspergillus fumigatus from the same order, Eurotiales, is an opportunistic pathogen of mammals which also produces EAs, such as festuclavine 2 [15,16]. Cycloclavine 7 is biosynthesised in nature by Aspergillus japonicus, which is frequently responsible for the post-harvest decay of fresh fruit (apples, pears, peaches, citrus, grapes, figs, strawberries, tomatoes or melons) and some vegetables (especially onions, garlic, and yams) [17].
The biosynthetic pathways of EAs have been well studied and are described in depth elsewhere [14,18]; however, a brief overview of EAs biosynthesis is given here. First, the prenylation of L-tryptophan by dimethylallyl pyrophosphate (DMAPP) yields 4-(γ,γ-dimethylallyl)tryptophan (DMAT) and is followed by the N-methylation of DMAT to 4-dimethyl-L-abrine (N-Me-DMAT). Subsequently, a series of successive oxidation steps catalyses the intramolecular cyclization of the prenyl and indole moieties to form ring C in tricyclic chanoclavine-I, which, in turn, is oxidised to form chanoclavine-I-aldehyde. At this branch point, chanoclavine-I-aldehyde undergoes intramolecular cyclization to form either ring D of tetracyclic agroclavine 1 (C. purpurea, N. lolii) or festuclavine 2 (A. fumigatus). Subsequently festuclavine 2 is further biotransformed into fumigaclavines. The new branch of this pathway is an unusual oxidation of the cyclic iminium form of chanoclavine-I-aldehyde catalysed by non-heme iron and α-ketoglutarate dependent oxidase EasH, to yield a unique cyclopropyl ring moiety which is fused to a five-membered ring, in cycloclavine 7. The structure of EasH and possible mechanism of cycloclavine 7 formation has been published recently [19,20].
Ergot-derived medicines, such as ergometrine 4, were used to facilitate obstetric deliveries or to treat postpartum haemorrhage. The high bioactivity of EAs is correlated with the ability of these compounds to act as agonists or antagonists toward neuroreceptors. Although some EAs, such as the hallucinogenic compound LSD 5 have been used as recreational drugs, most EAs were associated with medicinal applications, including treatments against migraine and tumour (ergotamine 8), Parkinson’s disease or restless leg syndrome (cabergoline 6; Figure 1B). However, their synergistic antimicrobial activity is a less commonly known fact. Lysergol 3 is a synthetic EA that exhibits a synergistic antibiotic pharmaceutical activity as a bioactive enhancer and a bioavailability facilitator for broad-spectrum antibiotics. This property facilitates the absorption of antibiotics across the cell membrane in animal cells resulting in increased action against Gram-positive and -negative bacteria [12]. With the recommended dosage of lysergol of 10 μg/mL, the improved activity of antimicrobial effect is in the range of 2–12 folds, against a wide spectrum of both Gram-positive and -negative bacteria including Escherichia. coli, Bacillus subtilis, Mycobacterium smegmatis and other similar microorganisms.
2.2. Fungal Polyketides
The polyketide pathway constitutes one of the major biosynthetic pathways leading to the production of fungal NPs. Polyketides are polymers synthesised from simple carboxylic acid derivatives (e.g., acetyl-CoA, malonyl-CoA, and methylmalonyl-CoA) into linear chains by iterative Claisen condensation, followed, in some cases, by reductive modification of the resulting β-keto groups. These compounds are synthesised in fungi (and other organisms) by enzymes called polyketide synthases (PKSs). Polyketides are extremely diverse and include compounds such as polyesters, polyphenols, macrolides (macrocyclic esters), polyenes and enediynes.
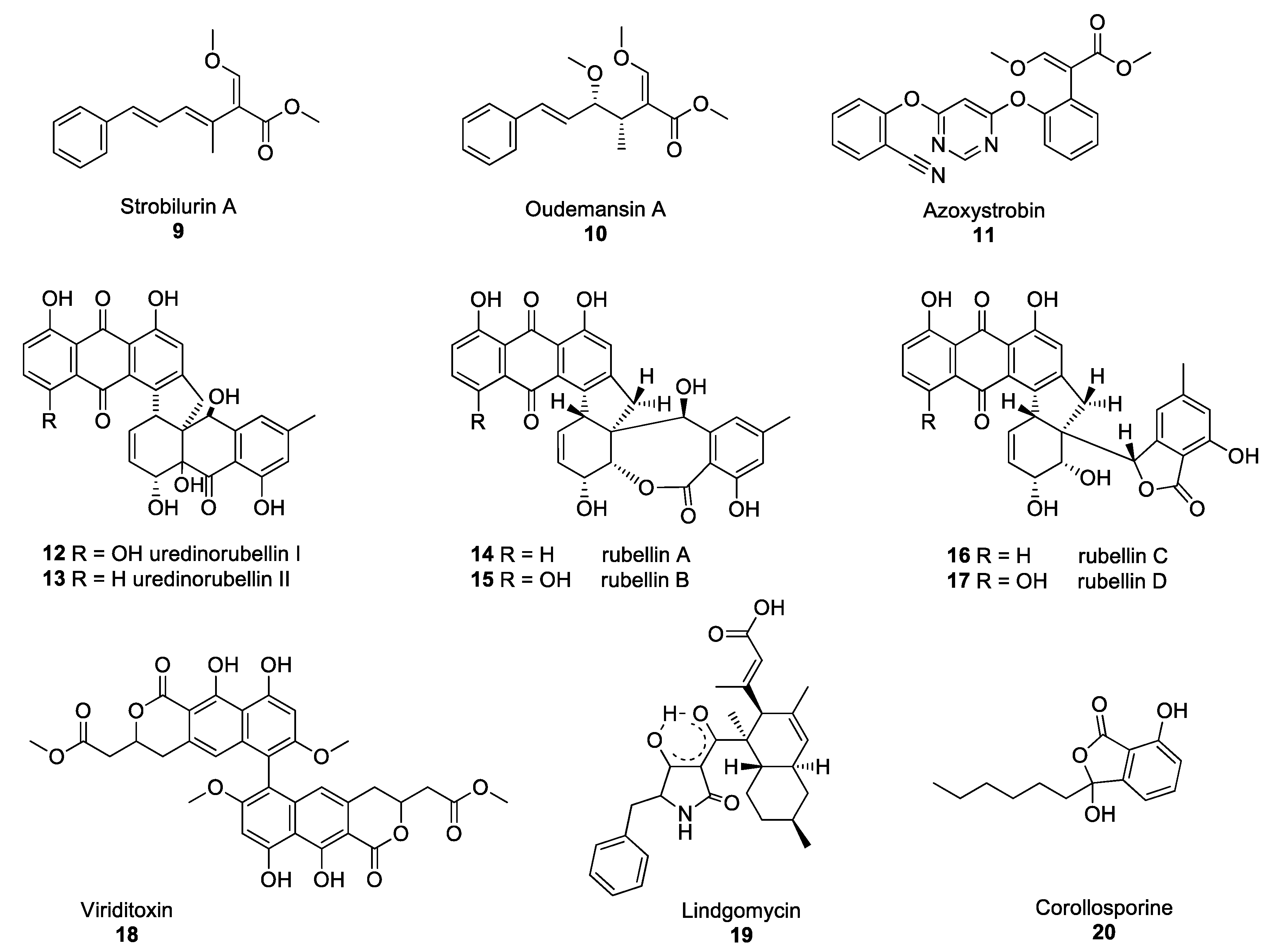
Strobilurins are an important group of polyketide-derived fungal NPs which have yielded one of the major classes of fungicides currently in use to protect agricultural crops from fungal diseases. The discovery of these compounds occurred after the observation that Strobilurus tenacellus and Oudemansiella mucida, two agaricomycetes growing on decaying wood in European forests, were able to defend themselves against other fungi. Their antifungal activity was associated with the production of the compounds strobilurin A 9 and oudemansin A 10 in S. tenacellus and O. mucida, respectively (Figure 2) [21,22]. These two compounds inhibit the transfer of electrons between complexes II and III of the electron transport chain in the mitochondria, resulting in impaired cell respiration and ATP synthesis [23]. Despite strobilurin A 9 and oudemansin A 10 exhibiting high antifungal activity, these NPs are quickly degraded by light, rendering them unsuitable for use in crop protection. Many attempts were made to modify the chemical structures of natural strobilurins to increase photo-stability while maintaining antifungal activity [24]. After several years of research, azoxystrobin 11 was synthesised and became the first photo-stable strobilurin-derived active ingredient registered for use in crop protection (Figure 2) [25], paving the way for the synthesis of a multitude of fungicides belonging to the quinone outside inhibitor (QoI) class of fungicides. However, resistance to QoI fungicides arose after a few years of use in fields. The single point mutation which confers resistance to QoI leads to the substitution of the amino acid glycine for alanine at position 143 (G143A). This mutation is now widespread in many fungal species, including Zymoseptoria tritici, Botrytis cinerea and Cercospora beticola, the agents responsible for Septoria leaf blotch in wheat, Botrytis grey mould and Cercospora leaf spot in beets, respectively [26,27,28]. Despite numerous resistance issues, QoIs are still used to control some of the most devastating rust fungi, such as Puccinia striiformis and Phakopsora pachyrhizi, the causative agents of the yellow rust of cereals and soybean rust diseases, respectively [29].
Some of the polyketide NPs synthesised by pathogenic fungi exhibit dimeric structures. Such is the case of lesser-known compounds produced by the Torrubiella species. The Torrubiella species are arthropod-pathogenic fungi that parasitise spiders, scale-insects and hoppers and are known to synthesise derivatives of uredinorubellin I 12 and II 13 (Figure 2). These compounds exhibited photodynamic activity, influencing cell viability in three mammalian cell lines, such as HIG82, HT29 and J774A.1, as well as antibacterial activity against Staphylococcus aureus [30]. The genus Torrubiella which belongs to the Clavicipitaceae family, is related to the genus Ramularia which also contains fungi that produce polyketide NPs with antimicrobial activity.
Ramularia collo-cygni is an ascomycete fungus responsible for the important plant disease Ramularia leaf spot (RLS) [31]. RLS is primarily a disease of barley but the fungus can infect other grain crops, such as wheat and oats as well as wild grasses. R. collo-cygni produces a range of secondary metabolites, including rubellin anthraquinones 14–17 (Figure 2). Rubellins are non-host-specific phytotoxins with photodynamic properties [32]. Miethbauer et al. showed that rubellins are biosynthesised via a polyketide pathway, by demonstrating the incorporation of both [1-13C]-acetate and [2-13C]-acetate into the rubellins during their formation [33]. McGrann and co-workers have recently sequenced and analysed the genome of R. collo-cygni and found that it contains the genetic architecture to synthesise a wide range of secondary metabolites, including rubellins [34]. In a later study, it was suggested that the co-expression of genes coding for PKSs and hybrid PKS/nonribosomal peptide synthetases (NRPSs) may be associated with the competitive colonisation of the host plant and early symptom development [35]. However, no exact determination of the biosynthetic pathway of rubellins has been elucidated yet and the role of these metabolites remains unclear. Despite the phytotoxic properties of rubellins, these compounds may have potential pharmaceutical applications. Miethbauer et al. have observed initial activities against Gram-positive bacteria, including MDR strains, such as B. subtilis (ATCC) 6633, S. aureus (SG) 511, S. aureus 134/94 (MRSA), Enterococcus faecalis 1528 (VRE) or Mycobacterium vaccae (IMET) 10670 [36]. Rubellins also exhibit antimicrobial, antiproliferative, cytotoxic and tau aggregation inhibitory activity in vitro tests [36]. Minimal inhibitory concentrations (MICs) were determined with and without illumination, showing a light-dependent increase in the antibacterial activity of compounds 14–17 (except against M. vaccae), with rubellin D 17 being the most active.
PKSs are not only involved in the biosynthesis of anthraquinone derivatives, such as rubellins, but also that of other dimers, such as viriditoxin 18 or lindgomycin 19 (Figure 2). Viriditoxin 18 belongs to the group of xanthoradones produced by Penicillium radicum FKI-3765-2. Xanthoradones exhibit activity against methicillin-resistant S. aureus (MRSA) by inhibiting FtsZ, the bacterial tubulin homolog which is crucial in septum formation [37].
Lindgomycin 19, an unusual antibiotic polyketide, contains two distinct structural domains, a bicyclic hydrocarbon and a tetramic acid that are connected by a carbonyl functional group. Naturally occurring tetramic acid derivatives, originating from a variety of marine and terrestrial fungi (Arctic fungus of the Lindgomycetaceae family), have attracted great interest due to the breadth of the spectrum of their biological activities, as well as their challenging structural complexity [38,39]. The majority of the compounds isolated to date have exhibited mostly antibiotic or antiviral activity. Lindgomycin 19 revealed good antibiotic activity against a number of Gram-positive bacteria (IC50: 2–6 μM), as well as the yeast Candida albicans and the plant pathogenic fungus Z. tritici (IC50: 5–10 μM) [40]. This compound also showed antibiotic activity against an MRSA strain with IC50 values of 5.1 μM. Another example of polyketide-derived NP from marine fungi is corollosporine 20 (Figure 2). Corollosporine 20 is an antibacterial phthalide derivative produced by the fungus Corollospora maritima which was isolated from driftwood found near the island of Heligoland, Germany [41].
2.3. Peptidic Fungal Natural Products
Two distinct pathways in fungi are responsible for the production of peptidic NPs. Enzymes in the nonribosomal peptide (NRP) pathway produce the majority of peptide metabolites. These are highly specific, multimodular enzymes called NRPSs, which utilise both proteinogenic and non-proteinogenic amino acids to synthesise the peptidic backbones. The genes encoding these enzymes are usually located within a biosynthetic cluster, comprising several genes that are co-regulated. The other pathway is that of ribosomally synthesised and post-translationally modified peptides (RiPPs); very large peptidic NPs with molecular weights typically around 1000 Da are synthesised through this pathway [42]. In this section we review selected groups of peptidic fungal NPs which have yielded valuable compounds with regards to cytotoxic properties and antibiotic drug discovery.
2.3.1. Ribosomally Synthesised and Post-Translationally Modified Peptides (RiPPs)
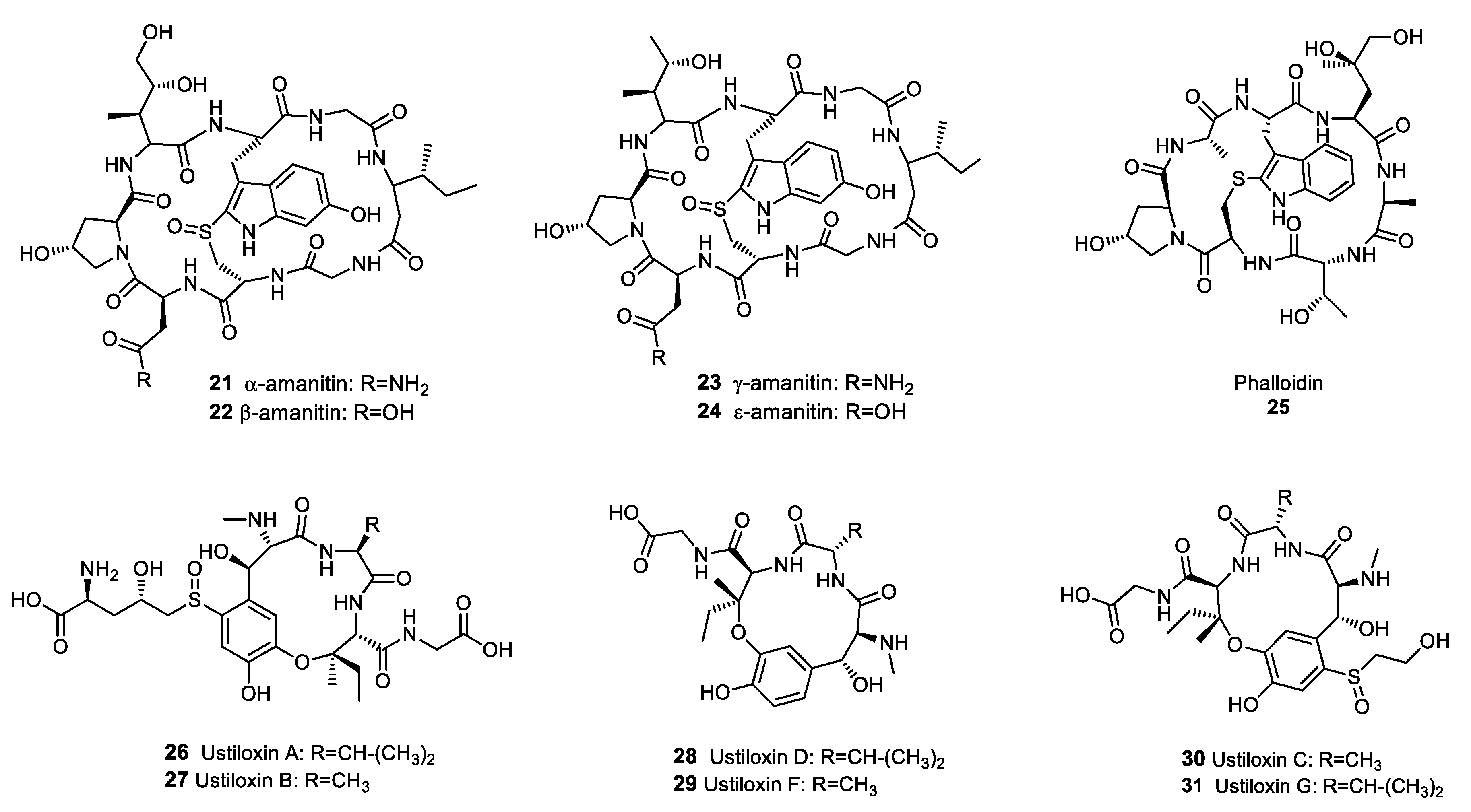
Although many RiPPs isolated from bacteria exhibit antibiotic activity [43], at the time of writing only a limited number of fungal RiPPs have been identified. The main groups of fungal RiPPs were isolated from agaricomycetes in the genus Amanita and belong to the amatoxin and phallotoxin families [44]. Amatoxins and phallotoxins are highly toxic compounds exhibiting cyclic octo- and hepta-peptide structures, respectively [45]. Despite the cytotoxicity of these metabolites, amatoxins and phallotoxins have yielded valuable compounds, although admittedly not antibiotics. Amanitins 21–24 (Figure 3), which belong to the amatoxin family, are potent inhibitors of RNA polymerase II and were investigated as potential anti-cancer drugs. Amanitin derivatives suppressed the multiplication of several cancer cell lines, including pancreatic, colorectal, and breast cancer cells [46]. The ability of phallotoxins, such as phalloidin 25, to bind actin filaments with a high affinity was used extensively to study cell biology, and phalloidin staining is still considered a gold standard for actin localisation in cells [47,48]. Ustiloxins 26–31 (Figure 3) were not originally identified as RiPPs when they were first isolated from the rice pathogen Ustilaginoidea virens, but the elucidation of their biosynthetic pathway has led to the reclassification of these metabolites as RiPPs [49]. Ustiloxins inhibit the assembly of microtubules during cell division, resulting in antimitotic properties and cytotoxicity against various cancer cells lines, including stomach, lung, breast, colon and kidney cancer cells [50]. Therefore, ustiloxins are of interest in the development of anti-cancer drugs.
2.3.2. Nonribosomal Peptide Natural Products
β-lactam Antibiotics
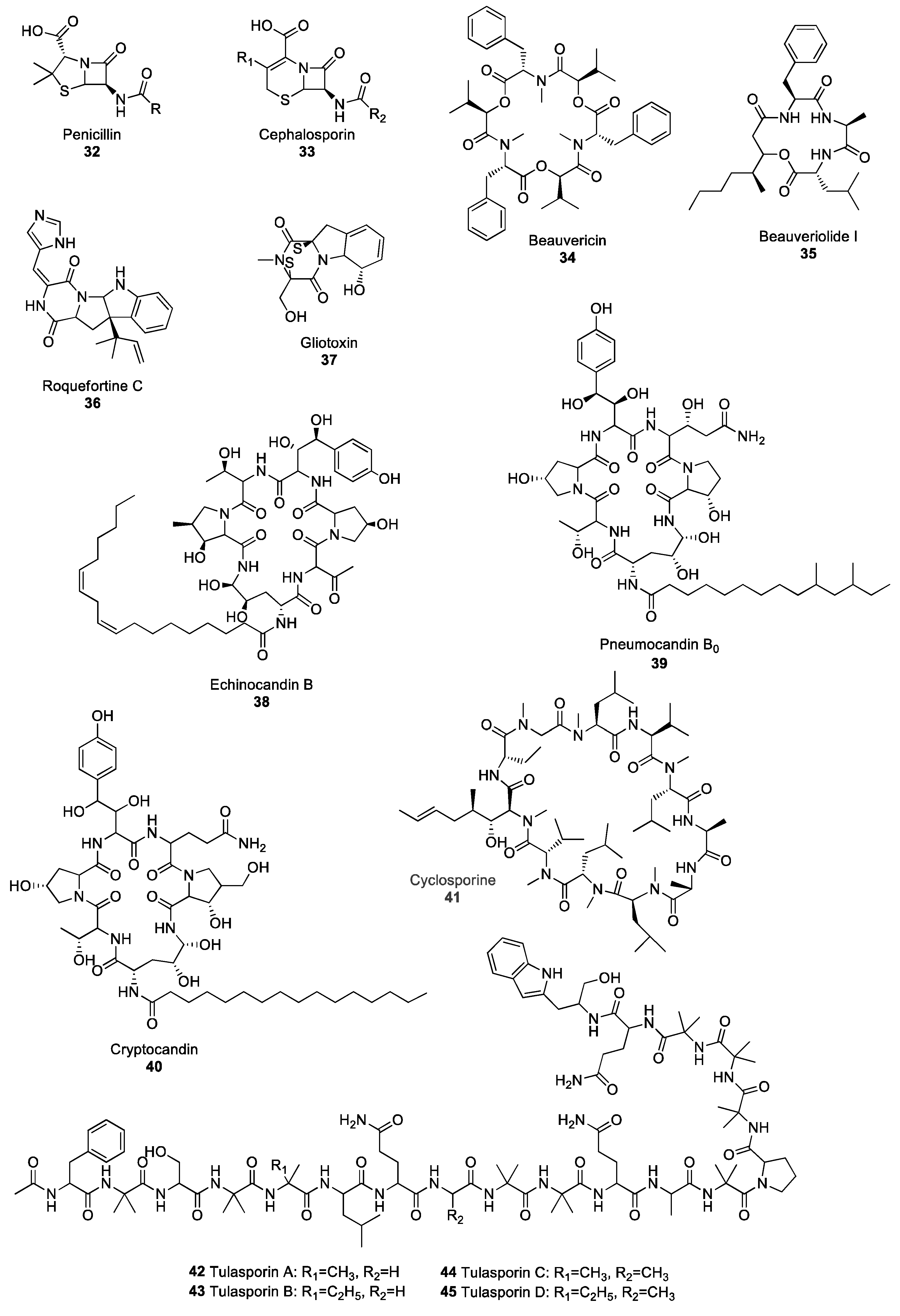
The discovery of the β-lactam class of antibiotics is, arguably, one of the greatest advancements of modern medicine, as it has given rise to a myriad of bioactive molecules. Compounds belonging to the β-lactam family all contain a four-member cyclic amide group. The two major families of products in the β-lactams are penicillins and cephalosporins, originally isolated from the Penicillium and Acremonium species, respectively [51,52]. The initial step in the biosynthesis of penicillin and cephalosporin requires the condensation of the three amino acids L-α-aminoadipate, L-cysteine and L-valine. The reaction is catalysed by the NRPS δ-(L-α-aminoadipyl)-L-cysteinyl-D-valine (ACV) synthetase [53]. Subsequently, ACV is transformed into isopenicillin N, which serves as a starting compound for either penicillin 32 or cephalosporin 33-derivative synthesis (Figure 4). β-lactams are the longest-serving antibiotics used in modern medicine and act against a broad spectrum of bacteria, including the pathogenic Streptococcus, Staphylococcus, Enterococcus, Clostrodium and Treponema species [54] by inhibiting the synthesis of bacterial cell wall peptidoglycans [55]. However, due to over-usage of β-lactams, many cases of resistance have been reported resulting in difficulties in controlling certain bacterial strains, including MRSA [56,57,58]. One of the most-studied mechanisms responsible for microbial resistance to antibiotics is the acquisition of genes encoding for β-lactamase enzymes by bacterial strains [59]. These genes, which can be integrated in the bacterial genome through horizontal gene transfer events, code for enzymes that are able to hydrolyse the β-lactam ring. The loss of the β-lactam ring is known to result in antibiotics inactivation. In addition to microbial resistance to antibiotics, cases of allergies to β-lactams have arisen, with about 10% of the population exhibiting allergic reactions to penicillins, rendering the control of bacterial diseases even more difficult [60]. Although only 1% of the population suffers from cephalosporin allergies, and allergies to other families of β-lactam are even less frequent, allergies are a major concern in clinical use. They are a particularly important consideration, as the ability to use different families of antibiotics including several β-lactams is crucial to prevent the development of multi-drug resistance [61,62].
Depsipeptides
Depsipeptides are peptides in which some amide groups are replaced by ester groups, resulting in lower susceptibility to proteolytic degradation [63]. Fungal depsipeptides have recently been reviewed at length [64], therefore, this section will focus on a few selected depsipeptides. Depsipeptides have been isolated from many ascomycete fungi, including plant pathogenic species in the genera Fusarium, Alternaria, Calonectria and Cochliobolus [65] as well as entomopathogenic fungal species, such as Cordyceps cardinalis and Ophiocordyceps communis [66,67]. Many depsipeptides exhibit both antimicrobial and insecticidal activity, and have, therefore, attracted attention for potential use in the development of new drug and crop protection products. Beauvericin 34 (Figure 4) is a family of natural products that were first isolated from Beauveria bassiana, the agent responsible for the white muscardine disease of arthropods. Many beauvericin derivatives have been identified and named alphabetically from A to J (including three G and H derivatives) and almost all of them exhibit biological activity including antimicrobial, antiviral and insecticidal activity; they can also inhibit cancer cell migration and be toxic to cancer cells [68,69,70,71,72]. The cytotoxicity of beauvericin is associated with the ability of this metabolite to form calcium channels in cell membranes, thus changing the intracellular cation levels and resulting in apoptosis, a form of programmed cell death [73,74]. Beauvericin was also reported to act as a potentiator of azole fungicides, resulting in reduced MIC against various strains of C. albicans, including azole-resistant strains [75]. Beauveriolide I 35, which was isolated from the entomopathogenic fungus Cordyceps militaris, exhibits anti-aging activity in the baker’s yeast Saccharomyces cerevisiae [76], as treatment with beauveriolide I 35 extends the chronological lifespan of the yeast. The chronological lifespan of yeast, which is commonly used as a model for the ageing of cells, corresponds to the amount of time a yeast cell stays alive in a non-dividing state [77]. The many biological properties of fungal depsispeptides and the potential they represent for medicinal use have led to a reclassification of these compounds from mycotoxins to drug candidates [78,79].
Piperazines
Piperazines are marked by the presence of a six-membered ring containing two nitrogen atoms in opposite positions, and are the starting point for the biosynthesis of compounds in the diketopiperazine and epipolythiodioxopiperazine (ETP) families. Roquefortine C 36 (Figure 4) is one of the most famous and well-studied fungal diketopiperazines. It has been isolated from several Penicillium species, including P. roqueforti, the fungus responsible for the blue veins in some cheeses, such as Roquefort, Gorgonzola or Stilton [80]. The role and impact of this secondary metabolite on human and animal health has been subject to debate for many years. Several authors reported acute toxicity from roquefortine C in mice and dogs [81,82,83], and postulated that, as roquefortine is consistently present in silages, it posed a potential threat to cattle [84,85]. However, a consensus has emerged that doses found in human and animal diets are far below the level required for roquefortine C toxicity [86,87]. Interestingly, diketopiperazines play an important role in communication between fungi. The production of a set of two diketopiperazines by Epichloë typhnia, an endophyte of timothy (Phleum pratense), induces the production of the antifungal perylenequinone metabolite phleichrome by Cladosporium phlei, a pathogen of timothy [88]. Gliotoxin 37 (Figure 4) is an ETP produced by the opportunistic fungus A. fumigatus, which is responsible for aspergillosis in immunodeficient patients and plays a crucial role as a virulence factor during host colonisation [89]. Gliotoxin 37 exhibits immunosuppressive activity by inhibiting the activation of certain transcription factors involved in the mounting of the immune response, including lymphocyte B and T activation [90]. Although gliotoxin demonstrates potent antifungal activity against C. albicans and several Aspergillus species [91], its high toxicity and adverse effect on human health made it unsuitable for use as a potential drug.
Lipopetides
Several fungal species are known to produce secondary metabolites belonging to the lipopeptide class of NPs. Lipopeptides are molecules synthesised via the NRP pathway and consist of a lipid backbone fused to a peptidic moiety. The echinocandin family, which regroups lipidated cyclic hexapeptides, has been an important source of antifungal agents since the turn of the century. Echinocandin B 38 (Figure 4) was first isolated from Aspergillus nidulans in 1974 [92] but has since been reported in other Aspergillus species of the Nidulantes section [93]. Echinocandins A, B, C, D, and H are highly potent antifungal agents that act by inhibiting β-(1,3)-glucan synthase, the enzyme responsible for the biosynthesis of β-(1,3)-glucan, one of the main components of the fungal cell wall [94]. Within the echinocandin family, pneumocandins are also important antifungal agents. Pneumocandin B0 39 (Figure 4) was first isolated from the saprophytic fungus Glarea lozoyensis [95] and exhibited high antifungal activity against C. albicans and Pneumocystis carinii, the agents responsible for candidiasis and pneumonia diseases [96]. Pneumocandin B0 39 is now used as the starting compound in the production of the semi-synthetic antifungal drug caspofungin acetate; pneumocandin B0 is chemically modified to increase its molecule stability and bioactivity [97]. Cryptocandin 40 (Figure 4) is another powerful antifungal agent that also belongs to the lipopetide class of NPs, and has also attracted attention for its potential use as a medicinal drug. Cryptocandin 40 was isolated from Cryptosporiopsis quercina, a common fungal endophyte of hardwood species [98]. Cryptocandin 40 has shown antifungal activity against a broad range of fungal species, including the agent responsible for skin mycosis, Tricophyton rubrum, as well as the plant pathogens Sclerotinia sclerotiorum and B. cinerea, responsible for white and grey mould, respectively [99].
Other Peptides
Fungi have also yielded several other peptide metabolites of interest. One of the most important is, arguably, cyclosporine 41 (Figure 4). This cyclic undecapeptide was first isolated from Toplypocladium inflatum, an entomopathogenic fungus, and is synthesised by the NRPS cyclosporine synthetase [100]. Although cyclosporine 41 exhibits broad spectrum antifungal activity [101] its most valuable contribution is as an immunosuppressive drug administrated after organ transplant to avoid rejection. Cyclosporine 41 prevents the mounting of an immune response by inhibiting the activity of calcineurin, a calcium/calmodulin-dependent serine threonine phosphatase which activates transcription factors, therefore regulating the expression of cytokine genes involved in immunity [102].
Peptaibols are linear peptide fungal NPs composed of a short chain of amino acids, generally shorter than 20 residues. Several peptaibols exhibit promising antibiotic activity against a broad spectrum of pathogens. A set of peptaibols isolated from Trichoderma reesei, a fungus used industrially to produce cellulase enzymes, are extremely potent growth inhibitors of several fungal species, including the plant pathogens Alternaria alternata, Phoma cucurbitaceum, Fusarium spp., as well as the human pathogen A. fumigatus [103]. Similarly, trichokonin VI, a peptaibol isolated from Trichoderma pseudokoningii, triggers programmed cell death in a broad range of fungal phytopathogens, including B. cinerea, F. oxysporum, Aschochyta citrullina and the oomycete Phytophtora parasitica, resulting in growth inhibition [104]. Tulasporins 42–45 (Figure 4) are a group of four 19-mer peptaibols isolated from Sepedonium tulasneanum, a mycoparasitic fungus. All four tulasporins exhibit antifungal activity against B. cinerea and P. infestans, the pathogens responsible for Botrytis grey mould and potato late blight, respectively [105]. In addition to inhibiting the growth of many fungal plant pathogens, some peptaibols also have the ability to induce the expression of defence-related genes in plants, therefore conferring increased resistance to pathogens. Application of an 18-mer peptaibol produced by T. virens induces the expression of genes encoding key enzymes in different defence signalling pathways, with up-regulation of hydroxyperoxide lyase and phenylalanine ammonia lyase in the oxylipin and salicylic acid pathways, respectively, which highlights the potential of these compounds to elicit plant immune response [106].
2.4. Terpenoid Compounds
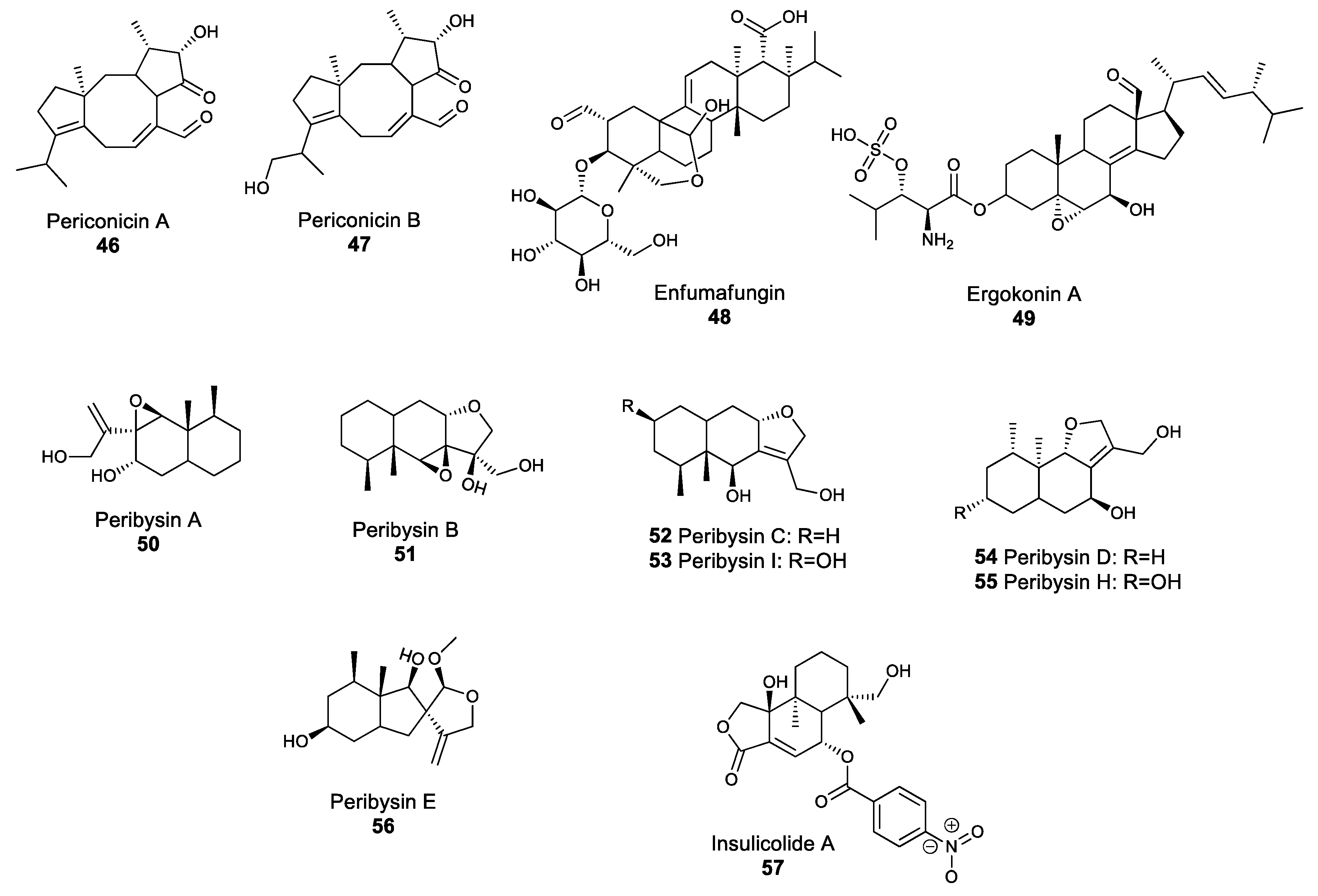
Terpenoids are a large class of natural products related to terpenes, and are made of condensed isoprene units. Terpenoids are often considered mostly as plant NPs; however, fungal species are also known to produce terpene-derived metabolites. Periconicin A 46 and B 47 (Figure 5) are secondary metabolites belonging to the diterpene family, which comprises compounds with four isoprene units. Both periconicin A and B exhibit antibacterial activity against several bacterial human pathogens, such as S. aureus and Salmonella typhimurium [107]. Similarly, enfumafungin 48 and ergokonin A 49 (Figure 5) are acidic terpenoids isolated from Hormonema and Trichoderma species, and which demonstrate antibiotic activity against several bacterial, yeast and fungal species, including B. subtilis, Cryptococcus neoformans, C. albicans and A. fumigatus [108,109]. The broad-spectrum activity of these two compounds was linked with their action as inhibitors of glucan biosynthesis [110]. The majority of bioactive fungal terpenes and terpenoids discovered in the past decade were isolated from marine fungi and fungi associated with algae [111]. For instance, peribysins 50–56 (Figure 5) are sesquiterpenes which were isolated from the dothideomycete Periconia byssoides, and are able to inhibit the development of leukemia cancer cells even at low concentrations, making compounds in this family potential anti-cancer drugs [112]. Similarly, the sesquiterpenoid insulicolide A 57, which was isolated from the marine Aspergillus ochraceus Jcma1F17 strain, is a promising natural product that showed biological activity against viruses, such as the influenza virus H3N2 and inhibited the growth of diverse human cancer cell lines [113]. The screening of marine fungi for natural products with potential pharmaceutical applications has only recently begun, but has the potential to yield several new drugs.
3. Concluding Remarks
The production of antimicrobial compounds by fungal species has been suspected and utilised by humankind for centuries. For instance, healers in Egypt ca. 2650 BC were known to use mouldy bread to treat superficial wounds; whereas, in central Asia, clergymen used a mouldy mixture of barley and apples to dress wounds [114]. Although they did not understand the mode of action of their remedies, healers were using fungal-derived antimicrobial compounds to prevent and combat any potential wound infections. However, the later identification of these compounds paved the way for major advances in therapeutic medicine by allowing for the scaling-up of their production and increasing the diversity of active compounds through the use of semi-synthetic chemistry. The discovery that a myriad of bioactive molecules could be found in nature acted as a catalyst in research efforts to study fungal organisms and identify new bioactive compounds.
However, not all the compounds that exhibit interesting biological properties are made into active pharmaceutical ingredients. Although it is difficult to estimate the exact numbers, a rough approximation is that, for every new drug being released onto the market, at least 10,000 compounds showing potential biological activity were screened [115]. The bioactivities of NPs are always first tested only against specific fungal or bacterial strains, or against specific cancer lines in laboratory conditions; however, due to their high toxicity, many of these molecules also affect non-target cells, rendering the compound unsuitable for medicinal use. Although highly toxic NPs are unlikely to reach the pharmaceutical market, the diversity of their chemical structures can serve as a source of inspiration for chemists to synthesise molecules with a more targeted activity.
Another potential barrier to commercialisation is the ability to increase production of bioactive NPs. Secondary metabolites are generally synthesised by fungi in response to specific environmental conditions, which are difficult to reproduce artificially. Furthermore, these metabolites are often synthesised in small quantities. However, several methods of strain improvement have been used to increase the ability of a fungus to consistently produce NPs in large amounts. For instance, untargeted mutagenesis and alteration of the growing medium has increased the production of pneumocandins in G. lozoyensis by over 100% and 300%, respectively [116]. In recent years, synthetic biology methods, such as the heterologous expression of fungal biosynthetic gene clusters and targeted mutagenesis, have become prominent methods to significantly increase metabolite production. The antibiotic pleuromutilin was originally isolated from the basidiomycete Clitopilus passeckerianus [117]. The reconstruction of the pleuromutilin biosynthetic gene cluster in an engineered strain of A. oryzae resulted in over 1000% increase of antibiotic titration [118]. To support the production of medicinal drugs derived from fungal NPs, commercial-scale synthesis must be achieved, potentially through optimising fungal strains, growth conditions and incorporating techniques, such as pathway engineering.
The present review mostly focused on antimicrobial properties of fungal NPs, but the value of these secondary metabolites extends far above that of the antibiotic activity. For instance, placlitaxel (commercialised under the named Taxol®), which is one of the most-used anticancer drugs, is a fungal NP originally isolated form Taxomyces andreanae, which is an endophyte of the pacific yew tree (Taxus brevifolia) [119]. Similarly, statins, such as lovastatin produced by A. terreus, are fungal NPs with hypolipidemic properties that inhibit the 3-hydroxy-3-methylglutaryl-CoA reductase, an enzyme involved in cholesterol biosynthesis [120]. Statins are, at present, the most important class of cholesterol-lowering pharmaceuticals, further highlighting the importance of fungal NPs. Considering that, (i) relative to the large diversity of fungal species, only a handful of fungi have been studied and cultivated and, (ii) based on genome mining studies, each fungus probably has the ability to produce several NPs, after years of research into fungal NPs, humankind has only started to scratch the very surface of this diverse world. Therefore, the vast majority of bioactive compounds remain yet to be identified, and perhaps within these molecules lie the foundations of the next pharmaceutical drug families.
Author Contributions
Both authors contributed equally to this manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
F.D. was partially funded by the Scottish Government Rural and Environment Science and Analytical Services Division (RESAS). D.J. was partially supported by Institute of Bioorganic Chemistry, Polish Academy of Sciences in Poznań, Poland.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiff, P.L. Ergot and its alkaloids. Am. J. Pharm. Educ. 2006, 70, 98. [Google Scholar] [CrossRef] [PubMed]
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach; John Wiley & Sons: West Sussex, UK, 2002; ISBN 0471496413. [Google Scholar]
- Mukherjee, J.; Menge, M. Progress and prospects of ergot alkaloid research. In New Products and New Areas of Bioprocess Engineering; Springer: Berlin/Heidelberg, Germany, 2000; pp. 1–20. [Google Scholar]
- Ligon, B.L. Penicillin: Its discovery and early development. Semin. Pediatr. Infect. Dis. WB Saunders 2004, 15. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Swierstra, J.; Wu, C.; Girard, G.; Choi, Y.H.; van Wamel, W.; Sandiford, S.K.; van Wezel, G.P. Eliciting antibiotics active against the ESKAPE pathogens in a collection of actinomycetes isolated from mountain soils. Microbiology 2014, 160, 1714–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Versluis, D. Antibiotic Resistance Reservoirs. 2016. Available online: https://research.wur.nl/en/publications/antibiotic-resistance-reservoirs-the-cases-of-sponge-and-human-gu (accessed on 20 January 2020).
- Hamad, B. The antibiotics market. Nat. Rev. Drug Discov. 2010, 9, 675–676. [Google Scholar] [CrossRef]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.; Lichten, C.A.; Taylor, J.; MacLure, C.; Lepetit, L.; Harte, E.; Martin, A.; Ghiga, I.; Pitchforth, E.; Sussex, J.; et al. Evaluation of the EC Action Plan Against the Rising Threats from Antimicrobial Resistance. RAND Corp. RB-9930-EC. 2018. Available online: https//www.rand.org/pubs/research_briefs/RB9930.html (accessed on 20 January 2020).
- Pimentel, D. Green revolution agriculture and chemical hazards. Sci. Total Environ. 1996, 188, S86–S98. [Google Scholar] [CrossRef]
- Schardl, C.L.; Panaccione, D.G.; Tudzynski, P. Ergot alkaloids–biology and molecular biology. Alkaloids Chem. Biol. 2006, 63, 45–86. [Google Scholar]
- Jakubczyk, D.; Caputi, L.; Hatsch, A.; Nielsen, C.A.F.; Diefenbacher, M.; Klein, J.; Molt, A.; Schröder, H.; Cheng, J.Z.; Naesby, M.; et al. Discovery and Reconstitution of the Cycloclavine Biosynthetic Pathway—Enzymatic Formation of a Cyclopropyl Group. Angew. Chem. Int. Ed. 2015, 54, 5117–5121. [Google Scholar] [CrossRef] [Green Version]
- Bräse, S. Privileged Scaffolds in Medicinal Chemistry; The Royal Society of Chemistry: Cambridge, UK, 2016; ISBN 978-1-78262-030-3. [Google Scholar]
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal secondary metabolism—From biochemistry to genomics. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar] [CrossRef]
- Brookman, J.L.; Denning, D.W. Molecular genetics in Aspergillus fumigatus. Curr. Opin. Microbiol. 2000, 3, 468–474. [Google Scholar] [CrossRef]
- Perrone, G.; Susca, A.; Cozzi, G.; Ehrlich, K.; Varga, J.; Frisvad, J.C.; Meijer, M.; Noonim, P.; Mahakarnchanakul, W.; Samson, R.A. Biodiversity of Aspergillus species in some important agricultural products. Stud. Mycol. 2007, 59, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Florea, S.; Panaccione, D.G.; Schardl, C.L. Ergot Alkaloids of the Family Clavicipitaceae. Phytopathology 2017, 107, 504–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakubczyk, D.; Caputi, L.; Stevenson, C.E.M.; Lawson, D.M.; O’Connor, S.E. Structural characterization of EasH (Aspergillus japonicus)—an oxidase involved in cycloclavine biosynthesis. Chem. Commun. 2016, 52, 14306–14309. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Liu, Y. Insights into the Mechanism and Enantioselectivity in the Biosynthesis of Ergot Alkaloid Cycloclavine Catalyzed by Aj_EasH from Aspergillus japonicus. Inorg. Chem. 2019. [Google Scholar] [CrossRef]
- Anke, T.; Oberwinkler, F.; Steglich, W.; Schramm, G. The strobilurins—New antifungal antibiotics from the basidiomycete strobilurus tenacellus (Pers. ex Fr.) Sing. J. Antibiot. (Tokyo) 1977, 30, 806–810. [Google Scholar] [CrossRef]
- Anke, T.; Hecht, H.; Schramm, G.; Steglich, W. Antibiotics from basidiomycetes. IX Oudemansin, an antifungal antibiotic from Oudemansiella mucida (Schrader ex Fr.) Hoehnel (agaricales). J. Antibiot. (Tokyo) 1979, 32, 1112–1117. [Google Scholar] [CrossRef] [Green Version]
- Brandt, U.; Haase, U.; Schaegger, H.; von Jagow, G. Species specificity and mechanism of action of strobilurins. Dechema Monogr. 1993, 129, 27–38. [Google Scholar]
- Clough, J. The strobilurins, oudemansins, and myxothiazols, fungicidal derivatives of β-methoxyacrylic acid. Nat. Prod. Rep. 1993, 10, 565–574. [Google Scholar] [CrossRef]
- Godwin, J.; Anthony, V.; Clough, J.; Godfrey, C. ICIA5504: A novel, broad spectrum, systemic beta-methoxyacrylate fungicide. In Proceedings of the Brighton Crop Protection Conference—Pests and Diseases; British Crop Protection Council: Farnham, Surrey, UK, 23–26 November 1992; pp. 435–442. [Google Scholar]
- Torriani, S.F.F.; Brunner, P.C.; McDonald, B.A.; Sierotzki, H. QoI resistance emerged independently at least 4 times in European populations of Mycosphaerella graminicola. Pest Manag. Sci. 2009, 65, 155–162. [Google Scholar] [CrossRef]
- Leroux, P.; Gredt, M.; Leroch, M.; Walker, A.S. Exploring mechanisms of resistance to respiratory inhibitors in field strains of botrytis cinerea, the causal agent of gray mold. Appl. Environ. Microbiol. 2010, 76, 6615–6630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piszczek, J.; Pieczul, K.; Kiniec, A. First report of G143A strobilurin resistance in Cercospora beticola in sugar beet (Beta vulgaris) in Poland. J. Plant Dis. Prot. 2018, 125, 99–101. [Google Scholar] [CrossRef] [Green Version]
- Oliver, R.P. A reassessment of the risk of rust fungi developing resistance to fungicides. Pest Manag. Sci. 2014, 70, 1641–1645. [Google Scholar] [CrossRef] [PubMed]
- Isaka, M.; Palasarn, S.; Tobwor, P.; Boonruangprapa, T.; Tasanathai, K. Bioactive anthraquinone dimers from the leafhopper pathogenic fungus Torrubiella sp. BCC 28517. J. Antibiot. (Tokyo) 2012, 65, 571. [Google Scholar] [CrossRef]
- Miethbauer, S.; Heiser, I.; Liebermann, B. The phytopathogenic fungus Ramularia collo-cygni produces biologically active rubellins on infected barley leaves. J. Phytopathol. 2003, 151, 665–668. [Google Scholar] [CrossRef]
- Heiser, I.; Heß, M.; Schmidtke, K.-U.; Vogler, U.; Miethbauer, S.; Liebermann, B. Fatty acid peroxidation by rubellin B, C and D, phytotoxins produced by Ramularia collo-cygni (Sutton et Waller). Physiol. Mol. Plant Pathol. 2004, 64, 135–143. [Google Scholar] [CrossRef]
- Miethbauer, S.; Haase, S.; Schmidtke, K.-U.; Günther, W.; Heiser, I.; Liebermann, B. Biosynthesis of photodynamically active rubellins and structure elucidation of new anthraquinone derivatives produced by Ramularia collo-cygni. Phytochemistry 2006, 67, 1206–1213. [Google Scholar] [CrossRef]
- McGrann, G.R.D.; Andongabo, A.; Sjokvist, E.; Trivedi, U.; Dussart, F.; Kaczmarek, M.; Mackenzie, A.; Fountaine, J.M.; Taylor, J.M.G.; Paterson, L.J.; et al. The genome of the emerging barley pathogen Ramularia collo-cygni. BMC Genom. 2016, 17, 584. [Google Scholar] [CrossRef] [Green Version]
- Dussart, F.; Douglas, R.; Sjökvist, E.; Hoebe, P.N.; Spoel, S.H.; McGrann, G.R.D. Genome-Based Discovery of Polyketide-Derived Secondary Metabolism Pathways in the Barley Pathogen Ramularia collo-cygni. Mol. Plant-Microbe Interact. 2018, 31, 962–975. [Google Scholar] [CrossRef] [Green Version]
- Miethbauer, S.; Gaube, F.; Mollmann, U.; Dahse, H.-M.; Schmidtke, M.; Gareis, M.; Pickhardt, M.; Liebermann, B. Antimicrobial, antiproliferative, cytotoxic, and tau inhibitory activity of rubellins and caeruleoramularin produced by the phytopathogenic fungus Ramularia collo-cygni. Planta Med. 2009, 75, 1523–1525. [Google Scholar] [CrossRef] [Green Version]
- Schueffler, A.; Anke, T. Fungal natural products in research and development. Nat. Prod. Rep. 2014, 31, 1425–1448. [Google Scholar] [CrossRef] [PubMed]
- Royles, B.J.L. Naturally Occurring Tetramic Acids: Structure, Isolation, and Synthesis. Chem. Rev. 1995, 95, 1981–2001. [Google Scholar] [CrossRef]
- Marfori, E.C.; Bamba, T.; Kajiyama, S.; Fukusaki, E.; Kobayashi, A. Biosynthetic studies of the tetramic acid antibiotic trichosetin. Tetrahedron 2002, 58, 6655–6658. [Google Scholar] [CrossRef]
- Imhoff, J.F. Natural products from marine fungi—Still an underrepresented resource. Mar. Drugs 2016, 14, 19. [Google Scholar] [CrossRef]
- Liberra, K.; Jansen, R.; Lindequist, U. Corollosporine, a new phthalide derivative from the marine fungus Corollospora maritima Werderm. 1069. Pharmazie 1998, 53, 578–581. [Google Scholar] [CrossRef]
- Arnison, P.G.; Bibb, M.J.; Bierbaum, G.; Bowers, A.A.; Bugni, T.S.; Bulaj, G.; Camarero, J.A.; Campopiano, D.J.; Challis, G.L.; Clardy, J.; et al. Ribosomally synthesized and post-translationally modified peptide natural products: Overview and recommendations for a universal nomenclature. Nat. Prod. Rep. 2013, 30, 108–160. [Google Scholar] [CrossRef]
- Hudson, G.A.; Mitchell, D.A. RiPP antibiotics: Biosynthesis and engineering potential. Curr. Opin. Microbiol. 2018, 45, 61–69. [Google Scholar] [CrossRef]
- Hallen, H.E.; Luo, H.; Scott-craig, J.S.; Walton, J.D. Gene family encoding the major toxins of lethal Amanita mushrooms. Proc. Natl. Acad. Sci. USA 2007, 104, 19097–19101. [Google Scholar] [CrossRef] [Green Version]
- Wong, J.H. Fungal Toxins. In Handbook of Biologically Active Peptides, 2nd ed.; Elsevier Inc.: San Diego, CA, USA, 2013; ISBN 9780123850959. [Google Scholar]
- Moldenhauer, G.; Salnikov, A.V.; Lüttgau, S.; Herr, I.; Anderl, J.; Faulstich, H. Therapeutic potential of amanitin-conjugated anti-epithelial cell adhesion molecule monoclonal antibody against pancreatic carcinoma. J. Natl. Cancer Inst. 2012, 104, 622–634. [Google Scholar] [CrossRef] [Green Version]
- Wulf, E.; Deboben, A.; Bautz, F.A.; Faulstich, H.; Wieland, T. Fluorescent phallotoxin, a tool for the visualization of cellular actin. Proc. Natl. Acad. Sci. USA 1979, 76, 4498–4502. [Google Scholar] [CrossRef] [Green Version]
- Anderl, J.; Echner, H.; Faulstich, H. Chemical modification allows phallotoxins and amatoxins to be used as tools in cell biology. Beilstein J. Org. Chem. 2012, 8, 2072–2084. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Minami, A.; Igarashi, Y.; Izumikawa, M.; Umemura, M.; Nagano, N.; Machida, M.; Kawahara, T.; Shin-ya, K.; Gomi, K.; et al. Unveiling the biosynthetic pathway of the ribosomally synthesized and post-translationally modified peptide ustiloxin B in filamentous fungi. Angew. Chem. Int. Ed. 2016, 55, 8072–8075. [Google Scholar] [CrossRef] [PubMed]
- Koiso, Y.; Li, Y.; Iwasaki, S.; Hanaka, K.; Kobayashi, T.; Sonoda, R.; Fujita, Y.; Yaegashi, H.; Sato, Z. Ustiloxins, antimitotic cyclic peptides from false smut balls on rice panicles caused by ustilaginoidea virens. J. Antibiot. (Tokyo) 1994, 47, 765–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaynes, R. The discovery of penicillin—New insights after more than 75 years of clinical use. Emerg. Infect. Dis. 2017, 23, 849–853. [Google Scholar] [CrossRef]
- Bo, G. Giuseppe Brotzu and the discovery of cephalosporins. Clin. Microbiol. Infect. 2000, 6, 6–8. [Google Scholar] [CrossRef] [Green Version]
- Byford, M.F.; Baldwin, J.E.; Shiau, C.Y.; Schofield, C.J. The mechanism of ACV synthetase. Chem. Rev. 1997, 97, 2631–2649. [Google Scholar] [CrossRef]
- Miller, E.L. The penicillins: A review and update. J. Midwifery Women’s Heal. 2002, 47, 426–434. [Google Scholar] [CrossRef]
- Tipper, D.J. Mode of action of beta-lactam antibiotics. Pharmacol. Ther. 1985, 27, 1–35. [Google Scholar] [CrossRef]
- Sabath, L. Mechanisms of resistance to beta-lactam antibiotics in strains of Staphylococcus aureus. Ann. Intern. Med. 1982, 97, 339–344. [Google Scholar] [CrossRef] [Green Version]
- Preheim, L.C.; Penn, R.G.; Sanders, C.C.; Goering, R.V.; Giger, D.K. Emergence of resistance to β-lactam and aminoglycoside antibiotics during moxalactam therapy of Pseudomonas aeruginosa infections. Antimicrob. Agents Chemother. 1982, 22, 1037–1041. [Google Scholar] [CrossRef] [Green Version]
- Hiramatsu, K.; Cui, L.; Kuroda, M.; Ito, T. The emergence and evolution of methicillin-resistant Staphylococcus aureus. Trends Microbiol. 2001, 9, 486–493. [Google Scholar] [CrossRef]
- Frère, J. Beta-lactamases and bacterial resistance to antibiotics. Mol. Microbiol. 1995, 16, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Solensky, R. Hypersensitivity reactions to beta-lactam antibiotics. Clin. Rev. Allergy Immunol. 2003, 24, 201–219. [Google Scholar] [CrossRef]
- Jarvis, M. Preventing the emergence of multidrug-resistant microorganisms through antimicrobial use controls: The complexity of the problem. Infect. Control Hosp. Epidemiol. 1996, 17, 490–495. [Google Scholar] [CrossRef]
- Macy, E. Penicillin and Beta-Lactam Allergy: Epidemiology and Diagnosis. Curr. Allergy Asthma Rep. 2014, 14, 1–7. [Google Scholar] [CrossRef]
- Stawikowski, M.; Cudic, P. Depsipeptide synthesis. Methods Mol. Biol. 2007, 386, 321–339. [Google Scholar]
- Wang, X.; Gong, X.; Li, P.; Lai, D.; Zhou, L. Structural diversity and biological activities of cyclic depsipeptides from fungi. Molecules 2018, 23, 169. [Google Scholar] [CrossRef] [Green Version]
- Anke, H.; Laatsch, H. Cyclic Peptides and Depsipeptides from Fungi. In Physiology and Genetics. The Mycota (A Comprehensive Treatise on Fungi as Experimental Systems for Basic and Applied Research); Anke, T., Weber, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 15, pp. 331–335. [Google Scholar]
- Umeyama, A.; Takahashi, K.; Grudniewska, A.; Shimizu, M.; Hayashi, S.; Kato, M.; Okamoto, Y.; Suenaga, M.; Ban, S.; Kumada, T.; et al. In Vitro antitrypanosomal activity of the cyclodepsipeptides, cardinalisamides A-C, from the insect pathogenic fungus Cordyceps cardinalis NBRC 103832. J. Antibiot. (Tokyo) 2014, 67, 163–166. [Google Scholar] [CrossRef]
- Haritakun, R.; Sappan, M.; Suvannakad, R.; Tasanathai, K.; Isaka, M. An antimycobacterial cyclodepsipeptide from the entomopathogenic fungus Ophiocordyceps communis BCC 16475. J. Nat. Prod. 2010, 73, 75–78. [Google Scholar] [CrossRef]
- Nilanonta, C.; Isaka, M.; Kittakoop, P.; Trakulnaleamsai, S.; Tanticharoen, M.; Thebtaranonth, Y. Precursor-directed biosynthesis of beauvericin analogs by the insect pathogenic fungus Paecilomyces tenuipes BCC 1614. Tetrahedron 2002, 58, 3355–3360. [Google Scholar] [CrossRef]
- Shin, C.G.; An, D.G.; Song, H.H.; Lee, C. Beauvericin and enniatins H, i and MK1688 are new potent inhibitors of human immunodeficiency virus type-1 integrase. J. Antibiot. (Tokyo) 2009, 62, 687–690. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhan, J.; Wijeratne, E.M.K.; Burns, A.M.; Gunatilaka, A.A.L.; Molnár, I. Cytotoxic and antihaptotactic beauvericin analogues from precursor-directed biosynthesis with the insect pathogen Beauveria bassiana ATCC 7159. J. Nat. Prod. 2007, 70, 1467–1471. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yan, K.; Zhang, Y.; Huang, R.; Bian, J.; Zheng, C.; Sun, H.; Chen, Z.; Sun, N.; An, R.; et al. High-throughput synergy screening identifies microbial metabolites as combination agents for the treatment of fungal infections. Proc. Natl. Acad. Sci. USA 2007, 104, 4606–4611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhang, D.M.; Jia, J.F.; Peng, Q.L.; Tian, H.Y.; Wang, L.; Ye, W.C. Cyclodepsipeptides from the ascocarps and insect-body portions of fungus Cordyceps cicadae. Fitoterapia 2014, 97, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Patocka, J.; Nepovimova, E.; Kuca, K. A review on the synthesis and bioactivity aspects of beauvericin, a Fusarium mycotoxin. Front. Pharmacol. 2018, 9, 1338. [Google Scholar] [CrossRef] [Green Version]
- Mallebrera, B.; Prosperini, A.; Font, G.; Ruiz, M.J. In Vitro mechanisms of Beauvericin toxicity: A review. Food Chem. Toxicol. 2018, 111, 537–545. [Google Scholar] [CrossRef]
- Fukuda, T.; Tomoda, H.; Omura, S. New beauvericins, potentiators of antifungal miconazole activity, produced by Beauveria sp. FKI-1366. II. Structure elucidation. J. Antibiot. (Tokyo) 2004, 57, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Nakaya, S.; Mizuno, S.; Ishigami, H.; Yamakawa, Y.; Kawagishi, H.; Ushimaru, T. New rapid screening method for anti-aging compounds using budding yeast and identification of beauveriolide I as a potent active compound. Biosci. Biotechnol. Biochem. 2012, 76, 1226–1228. [Google Scholar] [CrossRef]
- Fabrizio, P.; Longo, V.D. The chronological life span of Saccharomyces cerevisiae. Aging Cell 2003, 2, 73–81. [Google Scholar] [CrossRef]
- Taevernier, L.; Wynendaele, E.; De Vreese, L.; Burvenich, C.; De Spiegeleer, B. The mycotoxin definition reconsidered towards fungal cyclic depsipeptides. J. Environ. Sci. Heal. Part C Environ. Carcinog. Ecotoxicol. Rev. 2016, 34, 114–135. [Google Scholar] [CrossRef]
- Tedjiotsop Feudjio, F.; Dornetshuber, R.; Lemmens, M.; Hoffmann, O.; Lemmens-Gruber, R.; Berger, W. Beauvericin and enniatin: Emerging toxins and/or remedies? World Mycotoxin J. 2010, 3, 415–430. [Google Scholar] [CrossRef]
- Scott, P.M.; Merrien, M.A.; Polonsky, J. Roquefortine and isofumigaclavine A, metabolites from Penicillium roqueforti. Experientia 1976, 32, 140–142. [Google Scholar] [CrossRef]
- Arnold, D.; Scott, P.; McGuire, P.; Harwig, J.; Nera, E. Acute toxicity studies on roquefortine and PR toxin, metabolites of the Penicillium roqueforti in the mouse. Food Cosmet. Toxicol. 1978, 16, 369–371. [Google Scholar] [CrossRef]
- Lowes, N.R.; Smith, R.A.; Beck, B.E. Roquefortine in the stomach contents of dogs suspected of strychnine poisoning in Alberta. Can. Vet. J. 1992, 33, 535–538. [Google Scholar] [PubMed]
- Eriksen, G.S.; Jäderlund, K.H.; Moldes-Anaya, A.; Schönheit, J.; Bernhoft, A.; Jæger, G.; Rundberget, T.; Skaar, I. Poisoning of dogs with tremorgenic Penicillium toxins. Med. Mycol. 2010, 48, 188–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auerbach, H.; Oldenburg, E.; Weissbach, F. Incidence of penicillium roqueforti and roquefortine C in silages. J. Sci. Food Agric. 1998, 76, 565–572. [Google Scholar] [CrossRef]
- Driehuis, F.; Spanjer, M.C.; Scholten, J.M.; Te Giffel, M.C. Occurrence of mycotoxins in feedstuffs of dairy cows and estimation of total dietary intakes. J. Dairy Sci. 2008, 91, 4261–4271. [Google Scholar] [CrossRef]
- Bünger, J.; Westphal, G.; Mönnich, A.; Hinnendahl, B.; Hallier, E.; Müller, M. Cytotoxicity of occupationally and environmentally relevant mycotoxins. Toxicology 2004, 202, 199–211. [Google Scholar] [CrossRef]
- Finoli, C.; Vecchio, A.; Galli, A.; Dragoni, I. Roquefortine C occurrence in blue cheese. J. Food Prot. 2001, 64, 246–251. [Google Scholar] [CrossRef]
- Seto, Y.; Kogami, Y.; Shimanuki, T.; Takahashi, K.; Matsuura, H.; Yoshihara, T. Production of phleichrome by Cladosporium phlei as stimulated by diketopiperadines of Epichloe typhina. Biosci. Biotechnol. Biochem. 2005, 69, 1515–1519. [Google Scholar] [CrossRef] [Green Version]
- Sutton, P.; Waring, P.; Mullbacher, A. Exacerbation of invasive aspergillosis by the immunosuppressive fungal metabolite, gliotoxin. Immunol Cell Biol 1996, 74, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Pahl, B.H.L.; Kraub, B.; Schulze-osthoff, K.; Decker, T.; Traenckner, E.B.; Myersfl, C.; Parksfl, T.; Warring, P.; Miihlbacher, I.I.A.; Czernilofiky, A.; et al. The immunosuppressive fungal metabolite gliotoxin specifically inhibits transcription factor NF-kB. J. Exp. Med. 1996, 183, 1829–1840. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.J.; Ghosh, S.; Okoli, I.; Mylonakis, E. Antifungal activity of microbial secondary metabolites. PLoS ONE 2011, 6, e25321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyfeler, R.; Keller-Schierlein, W. Metabolites of microorganisms. 143. Echinocandin B, a novel polypeptide-antibiotic from Aspergillus nidulans var. echinulatus: Isolation and structural components. Helv. Chim. Acta 1974, 57, 2459–2477. [Google Scholar] [PubMed]
- De La Cruz, M.; Martín, J.; González-Menéndez, V.; Pérez-Victoria, I.; Moreno, C.; Tormo, J.R.; El Aouad, N.; Guarro, J.; Vicente, F.; Reyes, F.; et al. Chemical and physical modulation of antibiotic activity in Emericella species. Chem. Biodivers. 2012, 9, 1095–1113. [Google Scholar] [CrossRef] [PubMed]
- Denning, D. Echinocandins: A new class of antifungal. J. Antimicrob. Chemother. 2002, 49, 889–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, R.E.; Sesin, D.F.; Joshua, H.; Wilson, K.E.; Kempf, A.J.; Goklen, K.A.; Kuehner, D.; Gailliot, P.; Gleason, C.; White, R.; et al. Pneumocandins from Zalerion arboricola I. Discovery and isolation. J. Antibiot. (Tokyo) 1992, 45, 1853–1866. [Google Scholar] [CrossRef] [Green Version]
- Schmatz, D.M.; Abruzzo, G.; Powles, M.A.; Balkovec, J.M.; Black, R.M.; Nollstadt, K.; Mcfadden, D.C.; Bartizal, K. Pneumocandins from Zalerion arboricola IV. Biological evaluation of natural and semisynthetic pneumocandins for activity against Pneumocystis carinii and Candida species. J. Antibiot. (Tokyo) 1992, 45, 1886–1891. [Google Scholar] [CrossRef] [Green Version]
- Balkovec, J.; Hughes, D.; Masurekar, P.; Sable, C.; Schwartz, R.; Singh, S. Discovery and development of first in class antifungal caspofungin (CANCIDAS®)—A case study. Nat. Prod. Rep. 2014, 31, 15–34. [Google Scholar] [CrossRef]
- Strobel, G.A.; Miller, R.V.; Martinez-Miller, C.; Condron, M.M.; Teplow, D.B.; Hess, W.M. Cryptocandin, a potent antimycotic from the endophytic fungus Cryptosporiopsis cf. quercina. Microbiology 1999, 145, 1919–1926. [Google Scholar] [CrossRef] [Green Version]
- Strobel, G.; Daisy, B. Bioprospecting for microbial endophytes and their natural products. Microbiol. Mol. Biol. Rev. 2003, 67, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, G.; Schörgendorfer, K.; Schneider-Scherzer, E.; Leitner, E. The peptide synthetase catalyzing cyclosporine production in Tolypocladium niveum is encoded by a giant 45.8-kilobase open reading frame. Curr. Genet. 1994, 26, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Feng, P.; Yin, Y.; Bushley, K.; Spatafora, J.W.; Wang, C. Cyclosporine biosynthesis in Tolypocladium inflatum benefits fungal adaptation to the environment. MBio 2018, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuda, S.; Koyasu, S. Mechanisms of action of cyclosporine. Immunopharmacology 2000, 47, 119–125. [Google Scholar] [CrossRef]
- Marik, T.; Tyagi, C.; Balázs, D.; Urbán, P.; Szepesi, Á.; Bakacsy, L.; Endre, G.; Rakk, D.; Szekeres, A.; Andersson, M.A.; et al. Structural diversity and bioactivities of peptaibol compounds from the longibrachiatum clade of the filamentous fungal genus trichoderma. Front. Microbiol. 2019, 10, 1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, M.; Chen, L.; Wang, X.W.; Zhang, T.; Zhao, P.B.; Song, X.Y.; Sun, C.Y.; Chen, X.L.; Zhou, B.C.; Zhang, Y.Z. Antimicrobial peptaibols from Trichoderma pseudokoningii induce programmed cell death in plant fungal pathogens. Microbiology 2012, 158, 166–175. [Google Scholar] [CrossRef] [Green Version]
- Otto, A.; Laub, A.; Haid, M.; Porzel, A.; Schmidt, J.; Wessjohann, L.; Arnold, N. Tulasporins A-D, 19-residue peptaibols from the mycoparasitic fungus sepedonium tulasneanum. Nat. Prod. Commun. 2016, 11, 1821–1824. [Google Scholar] [CrossRef] [Green Version]
- Viterbo, A.; Wiest, A.; Brotman, Y.; Chet, I.; Kenerley, C. The 18mer peptaibols from Trichoderma virens elicit plant defence responses. Mol. Plant Pathol. 2007, 8, 737–746. [Google Scholar] [CrossRef]
- Kim, S.; Shin, D.S.; Lee, T.; Oh, K.B. Periconicins, two new fusicoccane diterpenes produced by an endophytic fungus Periconia sp. with antibacterial activity. J. Nat. Prod. 2004, 67, 448–450. [Google Scholar] [CrossRef]
- Pelaez, F.; Cabello, A.; Platas, G.; Díez, M.T.; Del Val, A.G.; Basilio, A.; Martán, I.; Vicente, F.; Bills, G.F.; Giacobbe, R.A.; et al. The discovery of enfumafungin, a novel antifungal compound produced by an endophytic Hormonema species biological activity and taxonomy of the producing organisms. Syst. Appl. Microbiol. 2000, 23, 333–343. [Google Scholar] [CrossRef]
- Vicente, M.F.; Cabello, A.; Platas, G.; Basilio, A.; Díez, M.T.; Dreikorn, S.; Giacobbe, R.A.; Onishi, J.C.; Meinz, M.; Kurtz, M.B.; et al. Antimicrobial activity of ergokonin A from Trichoderma longibrachiatum. J. Appl. Microbiol. 2001, 91, 806–813. [Google Scholar] [CrossRef] [Green Version]
- Onishi, J.; Meinz, M.; Thompson, J.; Curotto, J.; Dreikorn, S.; Rosenbach, M.; Douglas, C.; Abruzzo, G.; Flattery, A.; Kong, L.; et al. Discovery of novel antifungal (1,3)-β-D-glucan synthase inhibitors. Antimicrob. Agents Chemother. 2000, 44, 368–377. [Google Scholar] [CrossRef] [Green Version]
- Elissawy, A.M.; El-Shazly, M.; Ebada, S.S.; Singab, A.N.B.; Proksch, P. Bioactive terpenes from marine-derived fungi. Mar. Drugs 2015, 13, 1966–1992. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Doi, M.; Miura, A.; Harada, W.; Hiramura, M.; Minoura, K.; Tanaka, R.; Numata, A. Absolute stereostructures of cell-adhesion inhibitors, peribysins A, E, F and G, produced by a sea hare-derived Periconia sp. J. Antibiot. (Tokyo) 2005, 58, 185–191. [Google Scholar] [CrossRef]
- Fang, W.; Lin, X.; Zhou, X.; Wan, J.; Lu, X.; Yang, B.; Ai, W.; Lin, J.; Zhang, T.; Tu, Z.; et al. Cytotoxic and antiviral nitrobenzoyl sesquiterpenoids from the marine-derived fungus Aspergillus ochraceus Jcma1F17. Med. Chem. Commun. 2014, 5, 701–705. [Google Scholar] [CrossRef]
- Wainwright, M. Moulds in ancient and more recent medicine. Mycologist 1989, 3, 21–23. [Google Scholar] [CrossRef]
- Torjesen, I. Drug development: The journey of a medicine from lab to shelf. Pharm. J. 2015. [Google Scholar]
- Masurekar, P.S.; Fountoulakis, J.M.; Hallada, T.C.; Sosa, M.S.; Kaplan, L. Pneumocandins from zalerion arboricola II. Modification of product spectrum by mutation and medium manipulation. J. Antibiot. (Tokyo) 1992, 45, 1867–1874. [Google Scholar] [CrossRef] [Green Version]
- Kavanagh, F.; Hervey, A.; Robbins, W.J. Antibiotic Substances from Basidiomycetes: VIII. Pleurotus multilus (Fr.) Sacc. and Pleurotus passeckerianus Pilat. Proc. Natl. Acad. Sci. USA 1951, 37, 570–574. [Google Scholar]
- Bailey, A.M.; Alberti, F.; Kilaru, S.; Collins, C.M.; De Mattos-Shipley, K.; Hartley, A.J.; Hayes, P.; Griffin, A.; Lazarus, C.M.; Cox, R.J.; et al. Identification and manipulation of the pleuromutilin gene cluster from Clitopilus passeckerianus for increased rapid antibiotic production. Sci. Rep. 2016, 6, 25202. [Google Scholar] [CrossRef] [Green Version]
- Stierle, A.; Strobel, G.; Stierle, D. Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew. Science 1993, 260, 214–216. [Google Scholar] [CrossRef]
- Manzoni, M.; Rollini, M. Biosynthesis and biotechnological production of statins by filamentous fungi and application of these cholesterol-lowering drugs. Appl. Microbiol. Biotechnol. 2002, 58, 555–564. [Google Scholar]
Figure 1.
(A) Examples of clavines. (B) Simple lysergic acid derivatives. (C) The unusual ergoline scaffold of cycloclavine. (D) Ergopeptides consisting of D-lysergic acid with a cyclic tripeptide moiety [14].
Figure 1.
(A) Examples of clavines. (B) Simple lysergic acid derivatives. (C) The unusual ergoline scaffold of cycloclavine. (D) Ergopeptides consisting of D-lysergic acid with a cyclic tripeptide moiety [14].

Figure 2.
Structures of the selected fungal polyketides: Strobilurin A 9, oudemansin A 10, azoxystrobin 11, uredinorubellins I and II (12–13) rubellins A–D (14–17), viriditoxin 18, lindgomycin 19, corollosporine 20.
Figure 2.
Structures of the selected fungal polyketides: Strobilurin A 9, oudemansin A 10, azoxystrobin 11, uredinorubellins I and II (12–13) rubellins A–D (14–17), viriditoxin 18, lindgomycin 19, corollosporine 20.

Figure 3.
Structures of selected fungal ribosomally synthesised and post-translationally modified peptides (RiPPs).
Figure 3.
Structures of selected fungal ribosomally synthesised and post-translationally modified peptides (RiPPs).

Figure 4.
Diverse structures of selected non-ribosomal peptide (NRP)-derived fungal natural products.
Figure 4.
Diverse structures of selected non-ribosomal peptide (NRP)-derived fungal natural products.

Figure 5.
Structures of selected fungal terpenoids.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jakubczyk, D.; Dussart, F. Selected Fungal Natural Products with Antimicrobial Properties. Molecules 2020, 25, 911. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25040911
AMA Style
Jakubczyk D, Dussart F. Selected Fungal Natural Products with Antimicrobial Properties. Molecules. 2020; 25(4):911. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25040911
Chicago/Turabian StyleJakubczyk, Dorota, and Francois Dussart. 2020. "Selected Fungal Natural Products with Antimicrobial Properties" Molecules 25, no. 4: 911. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25040911