Characterizations and Assays of α-Glucosidase Inhibition Activity on Gallic Acid Cocrystals: Can the Cocrystals be Defined as a New Chemical Entity During Binding with the α-Glucosidase?

Abstract
:1. Introduction
2. Results and Discussion
2.1. Characterizations of the Cocrystals
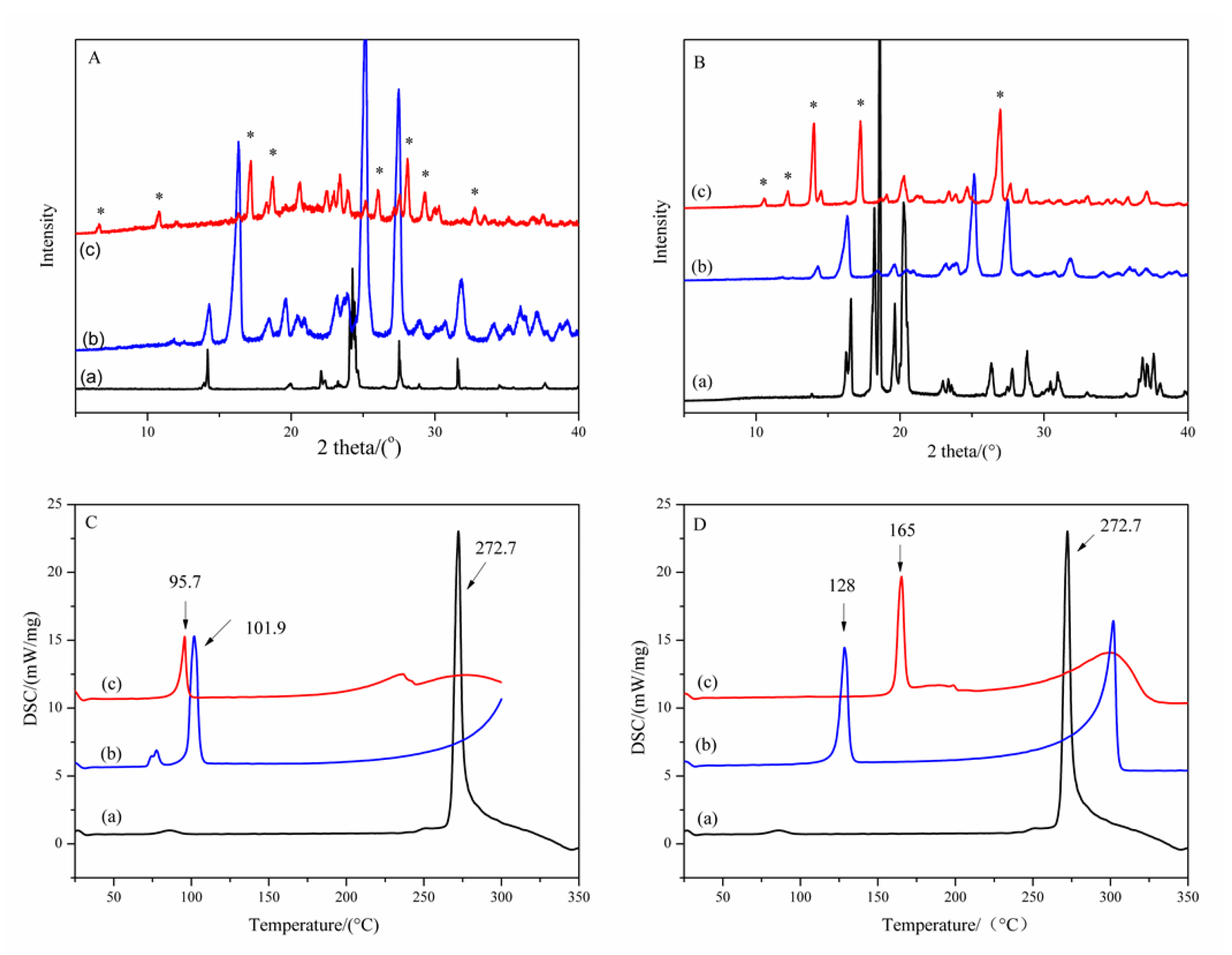
2.1.1. PXRD Patterns and DSC Curves
2.1.2. SEM Observation
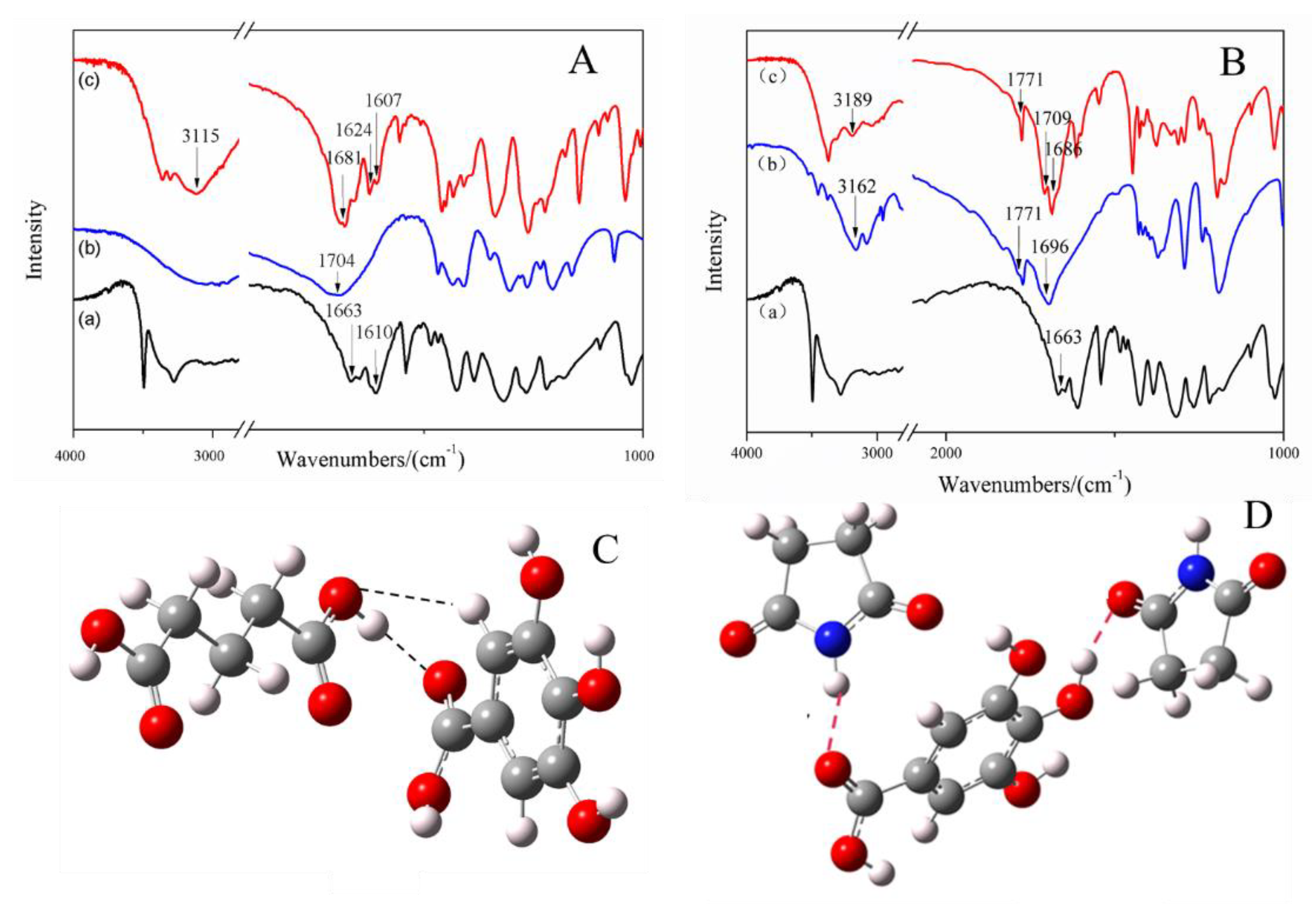
2.1.3. FT-IR Spectroscopy and DFT Simulation for Hydrogen Bonds in Cocrystals
2.2. Evaluations on Solubility, Dissolution Rate and Bioavailability
2.3. α-Glucosidase Inhibitory Activity
2.3.1. In Vitro Evaluation
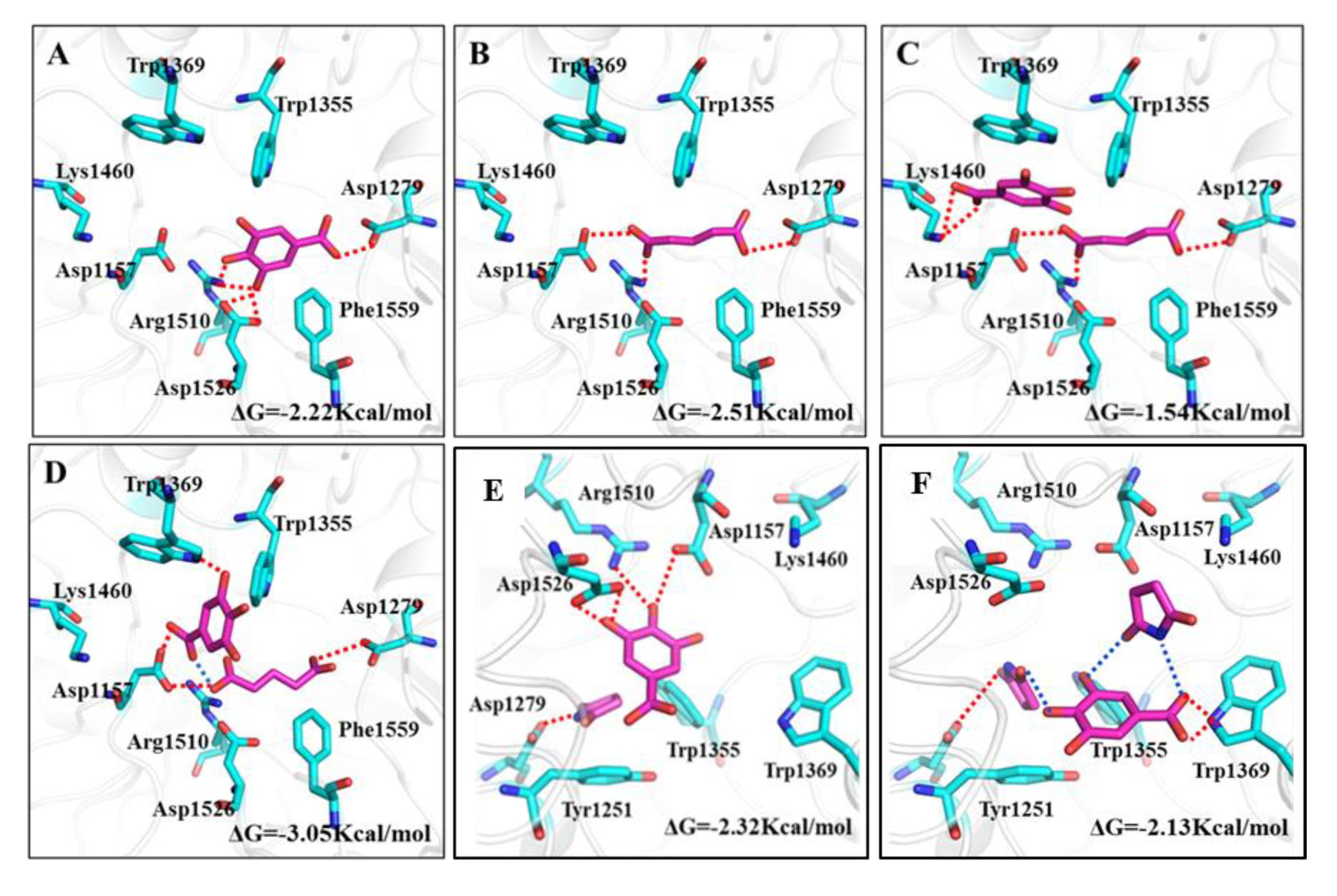
2.3.2. Molecular Docking
3. Materials and Methods
3.1. Materials
3.2. Methods
3.2.1. Preparation of Cocrystals
3.2.2. Characterizations
3.3. Assays on Solubility, Dissolution Rate and Bioavailability
3.4. Evaluation of α-Glucosidase Inhibitory Activity
3.4.1. Experimental Evaluation
3.4.2. Simulation Calculations
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Thorat, S.H.; Sahu, S.K.; Gonnade, R.G. Crystal structures of the pyrazinamide-p-aminobenzoic acid (1/1) cocrystal and the transamidation reaction product 4-(pyrazine-2-carboxamido)benzoic acid in the molten state. Acta Cryst. Struct Chem. 2015, 71, 1010–1016. [Google Scholar] [CrossRef] [PubMed]
- Bolla, G.; Nangia, A. Pharmaceutical cocrystals: Walking the talk. Chem. Commun. 2016, 52, 8342–8360. [Google Scholar] [CrossRef] [PubMed]
- Hiendrawan, S.; Veriansyah, B.; Widjojokusumo, E.; Soewandhi, S.N.; Wikarsa, S.; Tjandrawinata, R.R. Physicochemical and mechanical properties of paracetamol cocrystal with 5-nitroisophthalic acid. Int. J. Pharm. 2016, 497, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.S.; Kim, J.S.; Kim, M.S.; Alhalaweh, A.; Cho, W.; Hwang, S.J.; Velaga, S.P. Bioavailability of indomethacin-saccharin cocrystals. J. Pharm. Pharmacol. 2010, 62, 1560–1568. [Google Scholar] [CrossRef] [PubMed]
- Putra, O.D.; Umeda, D.; Nugraha, Y.P.; Furuishi, T.; Nagase, H.; Fukuzawa, K.; Yonemochi, E. Solubility improvement of epalrestat by layered structure formation via cocrystallization. CrystEngComm 2017, 19, 2614–2622. [Google Scholar] [CrossRef]
- Brittain, H.G. Cocrystal systems of pharmaceutical interest: 2010. Cryst. Growth Des. 2012, 12, 1046–1054. [Google Scholar] [CrossRef]
- Brittain, H.G. Cocrystal systems of pharmaceutical interest: 2011. Cryst. Growth Des. 2012, 12, 5823–5832. [Google Scholar] [CrossRef]
- Ferretti, V.; Dalpiaz, A.; Bertolasi, V.; Ferraro, L.; Beggiato, S.; Spizzo, F.; Pavan, B. Indomethacin co-crystals and their parent mixtures: Does the intestinal barrier recognize them differently? Mol. Pharm. 2015, 12, 1501–1511. [Google Scholar] [CrossRef]
- Li, K.; Guan, G.; Zhu, J.; Wu, H.; Sun, Q. Antibacterial activity and mechanism of a laccase-catalyzed chitosan-gallic acid derivative against escherichia coli and staphylococcus aureus. Food Control. 2019, 96, 234–243. [Google Scholar] [CrossRef]
- Karimi-Khouzani, O.; Heidarian, E.; Amini, S.A. Anti-inflammatory and ameliorative effects of gallic acid on fluoxetine-induced oxidative stress and liver damage in rats. Pharmacol. Rep. 2017, 69, 830–835. [Google Scholar] [CrossRef]
- Tanaka, M.; Kishimoto, Y.; Sato, A.; Sasaki, M.; Kamiya, T.; Kondo, K.; Iida, K. Molecular mechanisms underlying anti-Inflammatory and antioxidant activities of terminalia bellirica extract and gallic acid in LPS-stimulated macrophages. Atheroscler. Suppl. 2018, 32, 126. [Google Scholar] [CrossRef]
- Yigitturk, G.; Acara, A.C.; Erbas, O.; Oltulu, F.; Yavasoglu, N.U.K.; Uysal, A.; Yavasoglu, A. The antioxidant role of agomelatine and gallic acid on oxidative stress in STZ induced type I diabetic rat testes. Biomed. Pharmacother. 2017, 87, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Salas, M.G.; Estilla, A.R.; Chávez, J.M.; Sepúlveda, S.L.; Herrera, R.R.; González, C.A. P420 gallic acid has antiviral effect against hepatitis C virus (HCV), which is mediated by its antioxidant activity. J. Hepatol. 2014, 60, S208. [Google Scholar] [CrossRef]
- Ajibade, T.O.; Oyagbemi, A.A.; Omobowale, T.O.; Asenuga, E.R.; Afolabi, J.M.; Adedapo, A.A. Mitigation of diazinon-induced cardiovascular and renal dysfunction by gallic acid. Interdiscip Toxicol. 2016, 9, 66–77. [Google Scholar] [CrossRef]
- Abdel-Moneim, A.; Yousef, A.I.; El-Twab, S.M.A.; Reheim, E.S.A.; Ashour, M.B. Gallic acid and p-coumaric acid attenuate type 2 diabetes-induced neurodegeneration in rats. Metab. Brain Dis. 2017, 32, 1279–1286. [Google Scholar] [CrossRef]
- Abdel-Moneim, A.; El-Twab, S.M.A.; Yousef, A.I.; Reheim, E.S.A.; Ashour, M.B. Modulation of hyperglycemia and dyslipidemia in experimental type 2 diabetes by gallic acid and p-coumaric acid: The role of adipocytokines and PPARγ. Biomed. Pharmacother. 2018, 105, 1091–1097. [Google Scholar] [CrossRef]
- El-Lakkany, N.M.; El-Maadawy, W.H.; Seifel-Din, S.H.; Saleh, S.; Safar, M.M.; Ezzat, S.M.; Hammam, O.A. Antifibrotic effects of gallic acid on hepatic stellate cells: In vitro and in vivo mechanistic study. JTCM 2019, 9, 45–53. [Google Scholar] [CrossRef]
- Guimaraes, T.A.; Farias, L.C.; Fraga, C.A.; Feltenberger, J.D.; Melo, G.A.; Coletta, R.D.; Santos, S.H.S.; de Paula, A.M.B.; Guimaraes, A.L. Evaluation of the antineoplastic activity of gallic acid in oral squamous cell carcinoma under hypoxic conditions. Anti-Cancer Drugs 2016, 27, 407–416. [Google Scholar] [CrossRef]
- Oboh, G.; Ogunsuyi, O.B.; Ogunbadejo, M.D.; Adefegha, S.A. Influence of gallic acid on α-amylase and α-glucosidase inhibitory properties of acarbose. J. Food Drug Anal. 2016, 24, 627–634. [Google Scholar] [CrossRef]
- Huang, D.W.; Chang, W.C.; Wu, J.S.B.; Shih, R.W.; Shen, S.C. Gallic acid ameliorates hyperglycemia and improves hepatic carbohydrate metabolism in rats fed a high-fructose diet. Nutr. Res. 2016, 36, 150–160. [Google Scholar] [CrossRef]
- Variya, B.C.; Bakrania, A.K.; Madan, P.; Patel, S.S. Acute and 28-days repeated dose sub-acute toxicity study of gallic acid in albino mice. Regul. Toxicol. Pharm. 2019, 101, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Jacques, A.C.; Pertuzatti, P.B.; Barcia, M.T.; Zambiazi, R.C.; Chim, J.F. Estabilidade de compostos bioativos em polpa congelada de amora-preta (Rubus fruticosus) cv Tupy. Quim. Nova 2010, 33, 1720–1725. [Google Scholar] [CrossRef] [Green Version]
- Purbowatiningrum, N.O.; Fachriyah, E.; Eviana, I.; Eldiana, O.; Amaliyah, N.; Sektianingrum, A.N. Antidiabetic Activity from Gallic Acid Encapsulated Nanochitosan. Mater. Sci. Eng. 2017, 172, 012042. [Google Scholar] [CrossRef] [Green Version]
- Kaur, R.; Cherukuvada, S.; Managutti, P.B.; Row, T.N.G. A gallic acid-succinimide co-crystal landscape: Polymorphism, pseudo polymorphism, variable stoichiometry co-crystals and concomitant growth of non-solvated and solvated co-crystals. CrystEngComm 2016, 18, 3191–3203. [Google Scholar] [CrossRef]
- Dabir, T.O.; Gaikar, V.G.; Jayaraman, S.; Mukherjee, S. Thermodynamic modeling studies of aqueous solubility of caffeine, gallic acid and their cocrystal in the temperature range of 303 K-363 K. Fluid Phase Equilib. 2018, 456, 65–76. [Google Scholar] [CrossRef]
- Kaur, R.; Perumal, S.S.R.R.; Bhattacharyya, A.J.; Yashonath, S.; Guru Row, T.N. Structural insights into proton conduction in gallic acid-isoniazid cocrystals. Cryst. Growth Des. 2014, 14, 423–426. [Google Scholar] [CrossRef]
- Surov, A.O.; Churakov, A.V.; Proshin, A.N.; Dai, X.L.; Lu, T.; Perlovich, G.L. Cocrystals of a 1, 2, 4-thiadiazole-based potent neuroprotector with gallic acid: Solubility, thermodynamic stability relationships and formation pathways. Phys. Chem. Chem. Phys. 2018, 20, 14469–14481. [Google Scholar] [CrossRef]
- Yuliandra, Y.; Zaini, E.; Syofyan, S.; Pratiwi, W.; Putri, L.; Pratiwi, Y.; Arifin, H. Cocrystal of ibuprofen-nicotinamide: Solid-state characterization and in vivo analgesic activity evaluation. Sci. pharm. 2018, 86, 23. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.M.; Wu, D.M.; Zhang, L.L.; Xu, M.; Chen, J.H. Preparation and certification of reference material of gallic acid. Biomass Chem. Eng. 2013, 47, 49–53. [Google Scholar]
- Uno, T.; Machida, K. Infrared Spectra of Succinimide and Maleimide in the Crystalline State. B Chem Soc. JPN. 1962, 35, 276–283. [Google Scholar] [CrossRef]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B. Gaussian 09, Revision A. 02; Gaussian, Inc.: Wallingford, CT, USA, 2009. [Google Scholar]
- Zhang, B.W.; Sang, Y.B.; Sun, W.L.; Yu, H.S.; Ma, B.P.; Xiu, Z.L.; Dong, Y.S. Combination of flavonoids from Oroxylum indicum seed extracts and acarbose improves the inhibition of postprandial blood glucose: In vivo and in vitro study. Biomed. Pharm. 2017, 91, 890–898. [Google Scholar] [CrossRef] [PubMed]
- Proença, C.; Freitas, M.; Ribeiro, D.; Oliveira, E.F.T.; Sousa, J.; Tomé, S.; Ramos, M.J.; Silva, A.M.S.; Fernandes, P.; Fernandes, E. α-Glucosidase Inhibition by Flavonoids: An In Vitro and In Silico Structure-Activity Relationship Study. J. Enzym. Inhib. Med. Chem. 2017, 32, 1216–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Limanto, A.; Simamora, A.; Santoso, A.W.; Timotius, K.H. Antioxidant, a-Glucosidase Inhibitory Activity and Molecular Docking Study of Gallic Acid, Quercetin and Rutin: A Comparative Study. Mol. Cell Biomed. Sci. 2019, 3, 67–74. [Google Scholar] [CrossRef]
- Paulsen, P.J.; Cooke, W.D. Determination of Active Hydrogen by Nuclear Magnetic Resonance Spectrometry. Anal. Chem. 1964, 36, 1721–1723. [Google Scholar] [CrossRef]
- Tavani, C.; Bianchi, L.; De Palma, A.; Passeri, G.I.; Punzi, G.; Pierri, C.L.; Lovece, A.; Cavalluzzi, M.M.; Franchini, C.; Lentini, G.; et al. Nitro-Substituted Tetrahydroindolizines and Homologs: Design, Kinetics, and Mechanism of α-Glucosidase Inhibition. Bioorg Med. Chem Lett. 2017, 27, 3980–3986. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | IC50 (mmoL·L−1) |
|---|---|
| GA | 2.79 ± 0.09 |
| Glutaric acid | 1.51 ± 0.04 |
| GA-glutaric acid **△△ | 1.04 ± 0.04 |
| GA-succinimide cocrystal ** | 1.84 ± 0.30 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, N.; Jia, Y.; Li, C.; He, B.; Yang, C.; Wang, J. Characterizations and Assays of α-Glucosidase Inhibition Activity on Gallic Acid Cocrystals: Can the Cocrystals be Defined as a New Chemical Entity During Binding with the α-Glucosidase? Molecules 2020, 25, 1163. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25051163
Xue N, Jia Y, Li C, He B, Yang C, Wang J. Characterizations and Assays of α-Glucosidase Inhibition Activity on Gallic Acid Cocrystals: Can the Cocrystals be Defined as a New Chemical Entity During Binding with the α-Glucosidase? Molecules. 2020; 25(5):1163. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25051163
Chicago/Turabian StyleXue, Na, Yutao Jia, Congwei Li, Binnan He, Caiqin Yang, and Jing Wang. 2020. "Characterizations and Assays of α-Glucosidase Inhibition Activity on Gallic Acid Cocrystals: Can the Cocrystals be Defined as a New Chemical Entity During Binding with the α-Glucosidase?" Molecules 25, no. 5: 1163. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25051163