The Effect of Rutin and Extracts of Uncaria guianensis (Aubl.) J. F. Gmeland on Primary Endometriotic Cells: A 2D and 3D Study

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
2.1. Identification of Alkaloids and Phenolics in Extracts of U. guianensis
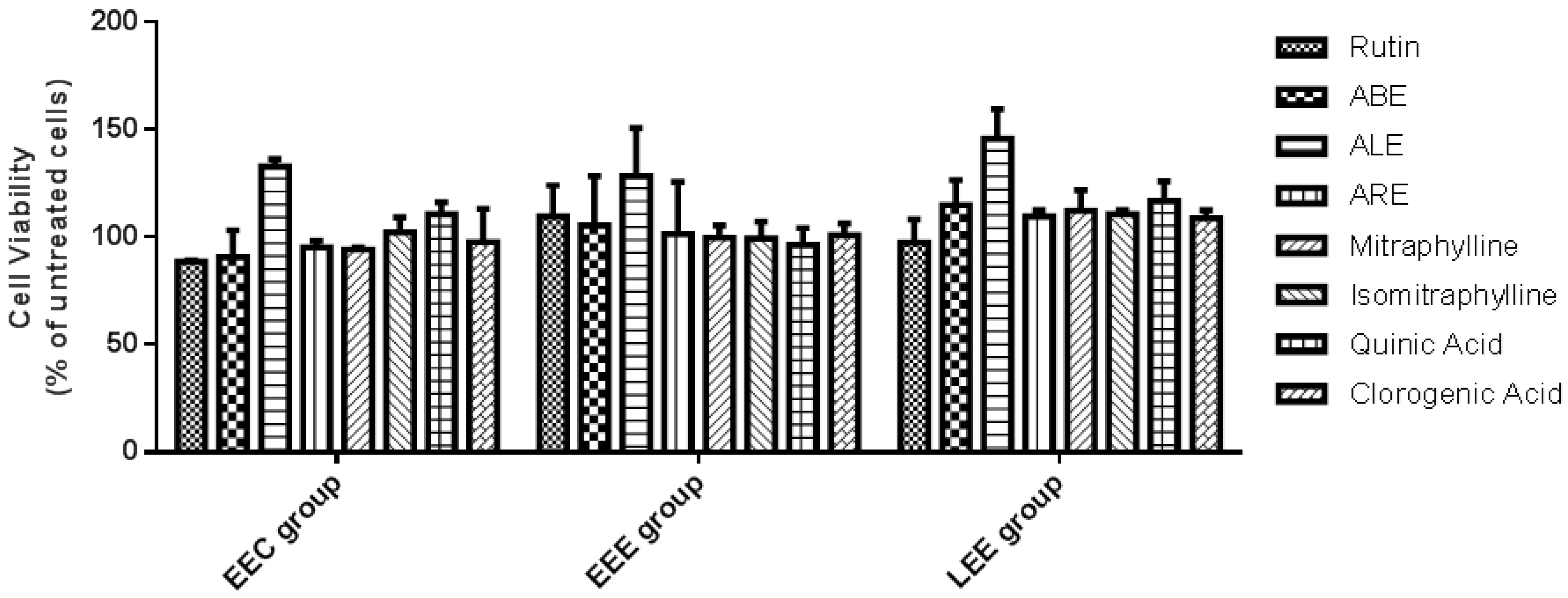
2.2. Viability of Endometrial Stromal Cells
2.3. Susceptibility of Endometrial Stromal Cells
2.4. Production of Reactive Oxygen Species (ROS) in Endometrial Stromal Cells
2.5. Production of Inflammatory Markers in Endometrial Stromal Cells
3. Discussion
4. Material and Methods
4.1. Preparation of Aqueous Extracts of Uncaria guianensis
4.2. Extraction and Quantification of Mitraphylline and Isomitraphylline
4.3. Analysis of Quinic and Chlorogenic Acids in Aqueous Extracts from U. guianensis
4.4. Patients and Sampling
4.5. Isolation and Culture of Primary Stromal Cells from Eutopic Endometrium and Lesions
4.6. Cell Characterization
4.7. Assessment of Cell Viability Using a 2D Culture Model
4.8. Assessment of Cell Susceptibility Using a 3D Culture Model
4.9. Quantification of ROS Using 2D and 3D Culture Models
4.10. Quantification of Inflammatory Markers Using 2D and 3D Culture Models
4.11. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Halis, G.; Mechsner, S.; Ebert, A.D. The diagnosis and treatment of deep infiltrating endometriosis. Dtsch. Arztebl. Int. 2010, 107, 446–455. [Google Scholar] [CrossRef] [PubMed]
- Andres, M.P.; Mendes, R.F.P.; Hernandes, C.; Araújo, S.E.A.; Podgaec, S. Hormone treatment as first line therapy is safe and relieves pelvic pain in women with bowel endometriosis. Einstein (São Paulo) 2019, 17, eAO4583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nisolle, M.; Donnez, J. Peritoneal endometriosis, ovarian endometriosis, and adenomyotic nodules of the rectovaginal septum are three different entities. Fertil. Steril. 1997, 68, 585–596. [Google Scholar] [CrossRef]
- Dunselman, G.A.J.; Vermeulen, N.; Becker, C.; Calhaz-Jorge, C.; D’Hooghe, T.; De Bie, B.; Heikinheimo, O.; Horne, A.W.; Kiesel, L.; Nap, A.; et al. ESHRE guideline: Management of women with endometriosis. Hum. Reprod. 2014, 29, 400–412. [Google Scholar] [CrossRef]
- Khan, K.N.; Fujishita, A.; Masumoto, H.; Muto, H.; Kitajima, M.; Masuzaki, H.; Kitawaki, J. Molecular detection of intrauterine microbial colonization in women with endometriosis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 199, 69–75. [Google Scholar] [CrossRef]
- Bellelis, P.; Frediani Barbeiro, D.; Gueuvoghlanian-Silva, B.Y.; Kalil, J.; Abrão, M.S.; Podgaec, S. Interleukin-15 and interleukin-7 are the major cytokines to maintain endometriosis. Gynecol. Obstet. Invest. 2019, 84, 435–444. [Google Scholar] [CrossRef]
- Sampson, J.A. Metastatic or embolic endometriosis, due to the menstrual dissemination of endometrial tissue into the venous circulation. Am. J. Pathol. 1927, 3, 93–110. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pmc/articles/PMC1931779/pdf/amjpathol00375-0001.pdf (accessed on 9 December 2019).
- Scutiero, G.; Iannone, P.; Bernardi, G.; Bonaccorsi, G.; Spadaro, S.; Volta, C.A.; Greco, P.; Nappi, L. Oxidative stress and endometriosis: A systematic review of the literature. Oxidative Med. Cell. Long. 2017, 2017. [Google Scholar] [CrossRef]
- Khan, K.N.; Fujishita, A.; Hiraki, K.; Kitajima, M.; Nakashima, M.; Fushiki, S.; Kitawaki, J. Bacterial contamination hypothesis: A new concept in endometriosis. Reprod. Med. Biol. 2018, 17, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Gueuvoghlanian-Silva, B.Y.; Bellelis, P.; Barbeiro, D.F.; Hernandes, C.; Podgaec, S. Treg and NK cells related cytokines are associated with deep rectosigmoid endometriosis and clinical symptoms related to the disease. J. Reprod. Immunol. 2018, 126, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, K.; Nishioka, K.; Khan, K.N.; Tanaka, Y.; Mori, T.; Nakaya, T.; Kitawaki, J. Molecular detection of microbial colonization in cervical mucus of women with and without endometriosis. Am. J. Reprod. Immunol. 2019, 82, 1–9. [Google Scholar] [CrossRef]
- Bellelis, P.; Podgaec, S.; Abrão, M.S. Fatores ambientais e endometriose: Um ponto de vista. Rev. Bras. Ginecol. Obs. 2014, 36, 433–435. [Google Scholar] [CrossRef] [Green Version]
- Agostinis, C.; Zorzet, S.; De Leo, R.; Zauli, G.; De Seta, F.; Bulla, R. The combination of N-acetyl cysteine, alpha-lipoic acid, and bromelain shows high anti-inflammatory properties in novel in vivo and in vitro models of endometriosis. Mediat. Inflamm. 2015, 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malvezzi, H.; Hernandes, C.; Piccinato, C.A.; Podgaec, S. Interleukin in endometriosis-associated infertility-pelvic pain: Systematic review and meta-analysis. Reproduction. 2019, 158, 1–12. [Google Scholar] [CrossRef]
- Nogueira-Neto, J.; Coelho, T.M.; Aguiar, G.C.; Carvalho, L.R.; de Araujo, A.G.P.; Girão, M.J.B.C.; Schor, E. Experimental endometriosis reduction in rats treated with Uncaria tomentosa (cat’s claw) extract. Eur. J. Obstet. Gynecol. Reprod. Biol. 2011, 154, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Casper, R.F. Progestin-only pills may be a better first-line treatment for endometriosis than combined estrogen-progestin contraceptive pills. Fertil. Steril. 2017, 107, 533–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zito, G.; Luppi, S.; Giolo, E.; Martinelli, M.; Venturin, I.; Di Lorenzo, G.; Ricci, G. Medical treatments for endometriosis-associated pelvic pain. Biomed. Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asante, A.; Taylor, R.N. Endometriosis: The role of neuroangiogenesis. Annu. Rev. Physiol. 2011, 73, 163–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urdanibia, I.; Michelangeli, F.; Ruiz, M.-C.; Milano, B.; Taylor, P. Anti-inflammatory and antitumoural effects of Uncaria guianensis bark. J. Ethnopharmacol. 2013, 150, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, M.; Okuhama, N.N.; Zhang, X.-J.; Condezo, L.A.; Lao, J.; Angeles, F.M.; Musah, R.A.; Bobrowski, P.; Miller, M.J.S. Anti-inflammatory and antioxidant activities of cat’s claw (Uncaria tomentosa and Uncaria guianensis) are independent of their alkaloid content. Phytomedicine 2002, 9, 325–337. [Google Scholar] [CrossRef] [Green Version]
- Burney, R.O.; Giudice, L.C. Pathogenesis and pathophysiology of endometriosis. Fertil. Steril. 2012, 98, 511–519. [Google Scholar] [CrossRef] [Green Version]
- Khan, K.N.; Kitajima, M.; Inoue, T.; Tateishi, S.; Fujishita, A.; Nakashima, M.; Masuzaki, H. Additive effects of inflammation and stress reaction on Toll-like receptor 4-mediated growth of endometriotic stromal cells. Hum. Reprod. 2013, 28, 2794–2803. [Google Scholar] [CrossRef] [Green Version]
- Podgaec, S.; Rizzo, L.V.; Fernandes, L.F.; Baracat, E.C.; Abrao, M.S. CD4(+) CD25(high) Foxp3(+) cells increased in the peritoneal fluid of patients with endometriosis. Am. J. Reprod. Immunol. 2012, 68, 301–308. [Google Scholar] [CrossRef]
- Rosa e Silva, J.C.; Fortunato, G.G.; Meola, J. Aspectos gerais da etiopatogenia da endometriose. In Endometriose, 1st ed.; Podgaec, S., Ed.; Elsevier Saúde: Rio de Janeiro, Brazil, 2014. [Google Scholar]
- Samimi, M.; Pourhanifeh, M.H.; Mehdizadehkashi, A.; Eftekhar, T.; Asemi, Z. The role of inflammation, oxidative stress, angiogenesis, and apoptosis in the pathophysiology of endometriosis: Basic science and new insights based on gene expression. J. Cell. Physiol. 2019, 234, 19384–19392. [Google Scholar] [CrossRef] [PubMed]
- Cherrak, S.A.; Mokhtari-Soulimane, N.; Berroukeche, F.; Bensenane, B.; Cherbonnel, A.; Merzouk, H.; Elhabiri, M. In vitro antioxidant versus metal ion chelating properties of flavonoids: A structure-activity investigation. PLoS ONE 2016, 11, e0165575. [Google Scholar] [CrossRef] [PubMed]
- Abarikwu, S.O.; Olufemi, P.D.; Lawrence, C.J.; Wekere, F.C.; Ochulor, A.C.; Barikuma, A.M. Rutin, an antioxidant flavonoid, induces glutathione and glutathione peroxidase activities to protect against ethanol effects in cadmium-induced oxidative stress in the testis of adult rats. Andrologia 2017, 49, e12696. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.N.; Kitajima, M.; Yamaguchi, N.; Fujishita, A.; Nakashima, M.; Ishimaru, T.; Masuzaki, H. Role of prostaglandin E2 in bacterial growth in women with endometriosis. Hum. Reprod. 2012, 27, 3417–3424. [Google Scholar] [CrossRef] [Green Version]
- Gautam, R.; Singh, M.; Gautam, S.; Rawat, J.K.; Saraf, S.A.; Kaithwas, G. Rutin attenuates intestinal toxicity induced by methotrexate linked with anti-oxidative and anti-inflammatory effects. BMC Complement. Altern. Med. 2016, 16, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosco, D.; Failla, P.; Costa, N.; Pullano, S.; Fiorillo, A.; Mollace, V.; Fresta, M.; Paolino, D. Rutin-loaded chitosan microspheres: Characterization and evaluation of the anti-inflammatory activity. Carbohydr. Polym. 2016, 152, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Na, J.Y.; Song, K.; Kim, S.; Kwon, J. Rutin protects rat articular chondrocytes against oxidative stress induced by hydrogen peroxide through SIRT1 activation. Biochem. Biophys. Res. Commun. 2016, 473, 1301–1308. [Google Scholar] [CrossRef] [PubMed]
- Guruvayoorappan, C.; Kuttan, G. Antiangiogenic effect of rutin and its regulatory effect on the production of VEGF, IL-1β and TNF-α in tumor associated macrophages. J. Biol. Sci. 2007, 7, 1511–1519. [Google Scholar] [CrossRef] [Green Version]
- Aguilar, J.L.; Rojas, P.; Marcelo, A.; Plaza, A.; Bauer, R.; Reininger, E.; Klaas, C.A.; Merfort, I. Anti-inflammatory activity of two different extracts of Uncaria tomentosa (Rubiaceae). J. Ethnopharmacol. 2002, 81, 271–276. [Google Scholar] [CrossRef]
- Malvezzi, H.; Viana, B.G.; Dobo, C.; Filippi, R.Z.; Podgaec, S.; Piccinato, C.A. Depleted lamin B1: A possible marker of the involvement of senescence in endometriosis? Arch. Gynecol. Obstet. 2018, 297, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Duval, K.; Grover, H.; Han, L.-H.; Mou, Y.; Pegoraro, A.F.; Fredberg, J.; Chen, Z. Modeling physiological events in 2D vs. 3D cell culture. Physiology 2017, 32, 266–277. [Google Scholar] [CrossRef]
- Chen, C.; Zhou, Y.; Hu, C.; Wang, Y.; Yan, Z.; Li, Z.; Wu, R. Mitochondria and oxidative stress in ovarian endometriosis. Free Rad. Biol. Med. 2019, 136, 22–34. [Google Scholar] [CrossRef]
- Ngô, C.; Chéreau, C.; Nicco, C.; Weill, B.; Chapron, C.; Batteux, F. Reactive oxygen species controls endometriosis progression. Am. J. Pathol. 2009, 175, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.-J.; Sun, H.-T.; Zhang, Z.-F.; Shi, R.-X.; Liu, L.-B.; Shang, W.-Q.; Wei, C.-Y.; Chang, K.-K.; Shao, J.; Wang, M.-Y.; et al. IL15 promotes growth and invasion of endometrial stromal cells and inhibits killing activity of NK cells in endometriosis. Reproduction 2016, 152, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Hendarto, H.; Widyanugraha, M.Y.A.W.; Widjiati, W. Curcumin improves growth factors expression of bovine cumulus-oocyte complexes cultured in peritoneal fluid of women with endometriosis. Int. J. Reprod. Biomed. 2018, 16, 775–782. [Google Scholar] [CrossRef]
- Invitti, A.L.; Schor, E.; Parreira, R.M.; Kopelman, A.; Kamergorodsky, G.; Gonçalves, G.A.; Girão, M.J.B.C. Inflammatory cytokine profile of co-cultivated primary cells from the endometrium of women with and without endometriosis. Mol. Med. Rep. 2018, 18, 1287–1296. [Google Scholar] [CrossRef] [Green Version]
- Sekiguchi, K.; Ito, Y.; Hattori, K.; Inoue, T.; Hosono, K.; Honda, M.; Numao, A.; Amano, H.; Shibuya, M.; Unno, N.; et al. VEGF receptor 1-expressing macrophages recruited from bone marrow enhances angiogenesis in endometrial tissues. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.-J.; Yang, H.-L.; Shao, J.; Mei, J.; Chang, K.-K.; Zhu, R.; Li, M.-Q. Anti-inflammatory cytokines in endometriosis. Cell. Mol. Life Sci. 2019, 76, 2111–2132. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, R.; Sirohi, V.K.; Gupta, K.; Dwivedi, A. Naringenin ameliorates progression of endometriosis by modulating Nrf2/Keap1/HO1 axis and inducing apoptosis in rats. J. Nutr. Biochem. 2019, 70, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Ferella, L.; Bastón, J.I.; Bilotas, M.A.; Singla, J.J.; González, A.M.; Olivares, C.N.; Meresman, G.F. Active compounds present in Rosmarinus officinalis leaves and Scutellaria baicalensis root evaluated as new therapeutic agents for endometriosis. Reprod. Biomed. Online 2018, 37, 769–782. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Lim, W.; Bazer, F.W.; Song, G. Apigenin induces ROS-dependent apoptosis and ER stress in human endometriosis cells. J. Cell. Physiol. 2018, 233, 3055–3065. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Lim, W.; You, S.; Song, G. Ameliorative effects of luteolin against endometriosis progression in vitro and in vivo. J. Nutr. Biochem. 2019, 67, 161–172. [Google Scholar] [CrossRef]
- Gentilini, D.; Busacca, M.; Di Francesco, S.; Vignali, M.; Viganò, P.; Di Blasio, A.M. PI3K/Akt and ERK1/2 signalling pathways are involved in endometrial cell migration induced by 17β-estradiol and growth factors. Mol. Hum. Reprod. 2007, 13, 317–322. [Google Scholar] [CrossRef]
- Schwenke, M.; Knöfler, M.; Velicky, P.; Weimar, C.H.E.; Kruse, M.; Samalecos, A.; Wolf, A.; Macklon, N.S.; Bamberger, A.-M.; Gellersen, B. Control of human endometrial stromal cell motility by PDGF-BB, HB-EGF and trophoblast-secreted factors. PLoS ONE 2013, 8, e54336. [Google Scholar] [CrossRef]
- Rakhila, H.; Al-Akoum, M.; Bergeron, M.-E.; Leboeuf, M.; Lemyre, M.; Akoum, A.; Pouliot, M. Promotion of angiogenesis and proliferation cytokines patterns in peritoneal fluid from women with endometriosis. J. Reprod. Immunol. 2016, 116, 1–6. [Google Scholar] [CrossRef]
- Falkiewicz, B.; Łukasiak, J. Vilcacora [Uncaria tomentosa (Willd.) DC. and Uncaria guianensis (Aublet) Gmell.]—a review of published scientific literature. Case Rep. Clin. Pract. Rev. 2001, 2, 305–316. Available online: https://www.amjcaserep.com/download/index/idArt/475352 (accessed on 9 December 2019).
- Luna-Palencia, G.R.; Huerta-Heredia, A.A.; Cerda-García-Rojas, C.M.; Ramos-Valdivia, A.C. Differential alkaloid profile in Uncaria tomentosa micropropagated plantlets and root cultures. Biotechnol. Lett. 2013, 35, 791–797. [Google Scholar] [CrossRef]
- Laus, G.; Brössner, D.; Keplinger, K. Alkaloids of peruvian Uncaria tomentosa. Phytochemistry 1997, 45, 855–860. [Google Scholar] [CrossRef]
- Wink, M. Evolutionary advantage and molecular modes of action of multi-component mixtures used in phytomedicine. Curr. Drug. Metab. 2008, 9, 996–1009. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Modes of action of herbal medicines and plant secondary metabolites. Medicines 2015, 2, 251–286. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, B.C.; Roxo, M.; Borges, M.C.; Peixoto, H.; Crevelin, E.J.; Bertoni, B.W.; Contini, S.H.T.C.; Lopes, A.A.; França, S.C.; Pereira, A.M.S.; et al. Antioxidant activity of an aqueous leaf extract from Uncaria tomentosa and its major alkaloids mitraphylline and isomitraphylline in Caenorhabditis elegans. Molecules 2019, 24, 3299. [Google Scholar] [CrossRef] [Green Version]
- Honório, I.C.G.; Bertoni, B.W.; Pereira, A.M.S. Uncaria tomentosa and Uncaria guianensis an agronomic history to be written. Cienc. Rural 2016, 46, 1401–1410. [Google Scholar] [CrossRef] [Green Version]
- Kapałczyńska, M.; Kolenda, T.; Przybyła, W.; Zajączkowska, M.; Teresiak, A.; Filas, V.; Ibbs, M.; Bliźniak, R.; Łuczewski, L.; Lamperska, K. 2D and 3D cell cultures—A comparison of different types of cancer cell cultures. Arch. Med. Sci 2018, 14, 910–919. [Google Scholar] [CrossRef]
- Bertol, G.; Franco, L.; Oliveira, B.H. HPLC analysis of oxindole alkaloids in Uncaria tomentosa: Sample preparation and analysis optimization by factorial design. Phytochem. Anal. 2012, 23, 143–151. [Google Scholar] [CrossRef]
- Peñaloza, E.M.C.; Kaiser, S.; Resende, P.E.; Pittol, V.; Carvalho, Â.R.; Ortega, G.G. Chemical composition variability in the Uncaria tomentosa (cat’s claw) wild population. Quim. Nova 2015, 38, 378–386. [Google Scholar] [CrossRef]
- Agência Nacional de Vigilância Sanitária. Resolução-RE no. 899, de 29 de maio de 2003. Guia para Validação de Métodos Analíticos e Bioanalíticos. Diário Oficial da União: Brasília, Brasil. 2003. Available online: http://portal.anvisa.gov.br/documents/10181/2718376/RE_899_2003_COMP.pdf/ff6fdc6b-3ad1-4d0f-9af2-3625422e6f4b (accessed on 22 November 2019).
- Khan, K.N.; Kitajima, M.; Hiraki, K.; Fujishita, A.; Nakashima, M.; Ishimaru, T.; Masuzaki, H. Cell proliferation effect of GnRH agonist on pathological lesions of women with endometriosis, adenomyosis and uterine myoma. Hum. Reprod. 2010, 25, 2878–2890. [Google Scholar] [CrossRef]
- Piccinato, C.A.; Neme, R.M.; Torres, N.; Sanches, L.R.; Cruz Derogis, P.B.; Brudniewski, H.F.; Rosa e Silva, J.C.; Ferriani, R.A. Increased expression of CYP1A1 and CYP1B1 in ovarian/peritoneal endometriotic lesions. Reproduction 2016, 151, 683–692. [Google Scholar] [CrossRef] [Green Version]
- Faraway, J.J. Extending the Linear Model. with R: Generalized Linear, Mixed Effects and Nonparametric Regression Models, 2nd ed.; Chapman and Hall/CRC press: Boca Raton, FL, USA, 2016. [Google Scholar]
Sample Availability: Samples of the compounds are not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | Mitraphylline (mg g−1 Dry Weight) | Isomitraphylline (mg g−1 Dry Weight) |
|---|---|---|
| ALE | 0.10 ± 0.010 | 0.15 ± 0.020 |
| ABE | < LoD and LoQ | 0.06 ± 0.003 |
| ARE | < LoD and LoQ | 0.03 ± 0.001 |
| Analyte | Estimated Mean pg mL−1 (95% CI) | Estimated Mean Difference 1 pg mL−1 (95% CI) | p-Value * |
|---|---|---|---|
| IL-15 | |||
| Untreated EEE cells | 0.56 (0.26, 1.23) | - | - |
| ABE (100 μg mL−1) treated EEE cells | 1.72 (0.97, 3.06) | 1.16 (0.04, 2.28) | 0.038 |
| IL-17A | |||
| Untreated EEE cells | 0.14 (−0.10, 0.38) | - | - |
| ABE (100 μg mL−1) treated EEE cells | 0.63 (0.39, 0.86) | 0.48 (0.16, 0.81) | 0.001 |
| IL-4 | |||
| Untreated EEE cells | 2.71 (0.83, 8.86) | - | - |
| ABE (100 μg mL−1) treated EEE cells | 13.25 (8.20, 21.41) | 10.55 (1.65, 19.44) | 0.013 |
| IL-6 | |||
| Untreated EEE cells | 62.21 (−4.17, 128.58) | - | - |
| ABE (100 μg mL−1) treated EEE cells | 161.68 (95.31, 228.05) | 99.48 (47.71, 155.25) | <0.001 |
| TNF-α | |||
| Untreated EEE cells | 1.00 (−1.06, 3.06) | - | - |
| ABE (100 μg mL−1) treated EEE cells | 3.75 (1.69, 5.81) | 2.75 (0.29, 5.20) | 0.021 |
| VEGF | |||
| Untreated EEE cells | 0.00 (−4.55, 4.55) | - | - |
| ABE (100 μg mL−1) treated EEE cells | 11.08 (6.53, 15.63) | 11.08 (2.99, 19.18) | 0.003 |
| EGF | |||
| Untreated LEE cells | 3.52 (1.52, 5.53) | - | - |
| ALE (100 μg mL−1) treated LEE cells | 7.07 (5.06, 9.07) | 3.54 (0.85, 6.24) | 0.005 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernandes, C.; de Oliveira, R.N.; de Souza Santos, A.H.; Malvezzi, H.; de Azevedo, B.C.; Gueuvoghlanian-Silva, B.Y.; Pereira, A.M.S.; Podgaec, S. The Effect of Rutin and Extracts of Uncaria guianensis (Aubl.) J. F. Gmeland on Primary Endometriotic Cells: A 2D and 3D Study. Molecules 2020, 25, 1325. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25061325
Hernandes C, de Oliveira RN, de Souza Santos AH, Malvezzi H, de Azevedo BC, Gueuvoghlanian-Silva BY, Pereira AMS, Podgaec S. The Effect of Rutin and Extracts of Uncaria guianensis (Aubl.) J. F. Gmeland on Primary Endometriotic Cells: A 2D and 3D Study. Molecules. 2020; 25(6):1325. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25061325
Chicago/Turabian StyleHernandes, Camila, Renata Nascimento de Oliveira, Artur Henrique de Souza Santos, Helena Malvezzi, Bruna Cestari de Azevedo, Bárbara Yasmin Gueuvoghlanian-Silva, Ana Maria Soares Pereira, and Sergio Podgaec. 2020. "The Effect of Rutin and Extracts of Uncaria guianensis (Aubl.) J. F. Gmeland on Primary Endometriotic Cells: A 2D and 3D Study" Molecules 25, no. 6: 1325. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25061325