
Protective Effects of Vitamin K Compounds on the Proteomic Profile of Osteoblasts under Oxidative Stress Conditions

 and
and 
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatment
4.2. SDS-PAGE and In-Gel Digestion
4.3. Liquid Chromatography-Mass Spectrometry (LC-MS)/MS Analysis
4.4. Protein Identification and Label-Free Quantification
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Rutkovskiy, A.; Stensløkken, K.O.; Vaage, I.J. Osteoblast Differentiation at a Glance. Med. Sci. Monit. Basic. Res. 2016, 22, 95–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchev, A.S.; Dimitrova, P.A.; Burns, A.J.; Kostov, R.V.; Dinkova-Kostova, A.T.; Georgiev, M.I. Oxidative stress and chronic inflammation in osteoarthritis: Can NRF2 counteract these partners in crime? Ann. N.Y. Acad. Sci. 2017, 1401, 114–135. [Google Scholar] [CrossRef] [PubMed]
- Domazetovic, V.; Marcucci, G.; Iantomasi, T.; Brandi, M.L.; Vincenzini, M.T. Oxidative stress in bone remodeling: Role of antioxidants. Clin. Cases Miner. Bone Metab. 2017, 14, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gavriilidis, C.; Miwa, S.; von Zglinicki, T.; Taylor, R.W.; Young, D.A. Mitochondrial dysfunction in osteoarthritis is associated with down-regulation of superoxide dismutase 2. Arthritis Rheum. 2013, 65, 378–387. [Google Scholar] [CrossRef]
- Basu, S.; Michaëlsson, K.; Olofsson, H.; Johansson, S.; Melhus, H. Association between oxidative stress and bone mineral density. Biochem. Biophys. Res. Commun. 2001, 288, 275–279. [Google Scholar] [CrossRef]
- Baltacıoğlu, E.; Kehribar, M.A.; Yuva, P.; Alver, A.; Atagün, Ö.S.; Karabulut, E.; Akalın, F.A. Total Oxidant Status and Bone Resorption Biomarkers in Serum and Gingival Crevicular Fluid of Patients with Periodontitis. J. Periodontol. 2014, 85, 317–326. [Google Scholar] [CrossRef]
- Koerdt, S.; Dax, S.; Grimaldi, H.; Ristow, O.; Kuebler, A.C.; Reuther, T. Histomorphologic characteristics of bisphosphonate-related osteonecrosis of the jaw. J. Oral. Pathol. Med. 2014, 43, 448–453. [Google Scholar] [CrossRef]
- Almeida, M.; O’Brien, C.A. Basic biology of skeletal aging: Role of stress response pathways. J. Gerontol. A Biol. Sci. Med. Sci. 2013, 68, 1197–1208. [Google Scholar] [CrossRef] [Green Version]
- Bai, X.C.; Lu, D.; Bai, J.; Zheng, H.; Ke, Z.Y.; Li, X.M.; Luo, S.Q. Oxidative stress inhibits osteoblastic differentiation of bone cells by ERK and NF-kappaB. Biochem. Piophys. Res. Commun. 2004, 314, 197–207. [Google Scholar] [CrossRef]
- Arai, M.; Shibata, Y.; Pugdee, K.; Abiko, Y.; Ogata, Y. Effects of reactive oxygen species (ROS) on antioxidant system and osteoblastic differentiation in MC3T3-E1 cells. IUBMB Life 2007, 59, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Jung, W.W. Protective effect of apigenin against oxidative stress-induced damage in osteoblastic cells. Int. J. Mol. Med. 2014, 33, 1327–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villa, J.K.D.; Diaz, M.A.N.; Virgínia Ramos Pizziolo, V.R.; Martino, H.S.D. Effect of vitamin K in bone metabolism and vascular calcification: A review of mechanisms of action and evidences. Crit. Rev. Food Sci. Nutr. 2017, 57, 3959–3970. [Google Scholar] [CrossRef] [PubMed]
- Jahanian, E.; Karimifar, M.; Rafieian-Kopaei, M. Antioxidants as a novel way to alleviate the adverse effects of oxidative stress in osteoporosis. J. Parathyr. Dis. 2016, 4, 60–65. [Google Scholar]
- Ivanova, D.; Zhelev, Z.; Getsov, P.; Nikolova, B.; Aoki, I.; Higashi, T.; Bakalova, R. Vitamin K: Redox-modulation, prevention of mitochondrial dysfunction and anticancer effect. Redox Biol. 2018, 16, 352–358. [Google Scholar] [CrossRef]
- Shiraki, M.; Tsugawa, N.; Okano, T. Recent advances in vitamin K-dependent Gla-containing proteins and vitamin K nutrition. Osteoporosis 2015, 1, 22–38. [Google Scholar] [CrossRef] [Green Version]
- Villa, M.M.; Wang, L.; Huang, J.; Rowe, D.W.; Wei, M. Bone tissue engineering with a collagen–hydroxyapatite scaffold and culture expanded bone marrow stromal cells. J. Biomed. Mater. Res. B Appl. Biomater. 2015, 103, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Martínez, J.; Moreno, J.J. Influence of superoxide radical and hydrogen peroxide on arachidonic acid mobilization. Arch. Biochem. Biophys. 1996, 336, 191–198. [Google Scholar] [CrossRef]
- Martínez, J.; Moreno, J.J. Role of Ca2+-independent phospholipase A2 on arachidonic acid release induced by reactive oxygen species. Arch. Biochem. Biophys. 2001, 392, 257–262. [Google Scholar] [CrossRef]
- Mukai, K.; Morimoto, H.; Kikuchi, S.; Nagaoka, S.-I. Kinetic study of free-radical-scavenging action of biological hydroquinones (reduced forms of ubiquinone, vitamin K and tocopherol quinone) in solution. Biochim. Biophys. Acta. 1993, 1157, 313–317. [Google Scholar] [CrossRef]
- Li, J.; Wang, H.; Rosenberg, P.A. Vitamin K prevents oxidative cell death by inhibiting activation of 12-lipoxygenase in developing oligodendrocytes. J. Neurosci. Res. 2009, 87, 1997–2005. [Google Scholar] [CrossRef] [Green Version]
- Cabral, M.; Martín-Venegas, R.; Moreno, J.J. Role of arachidonic acid metabolites on the control of non-differentiated intestinal epithelial cell growth. Int. J. Biochem. Cell Biol. 2013, 45, 1620–1628. [Google Scholar] [CrossRef] [PubMed]
- Cabral, M.; Martín-Venegas, R.; Moreno, J.J. Differential cell growth/apoptosis behavior of 13-hydroxyoctadecadienoic acid enantiomers in a colorectal cancer cell line. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G664–G671. [Google Scholar] [CrossRef] [Green Version]
- Ohsaki, Y.; Shirakawa, H.; Hiwatashi, K.; Furukawa, Y.; Mizutani, T.; Komai, M. Vitamin K Suppresses Lipopolysaccharide-Induced Inflammation in the Rat. Biosci. Biotechnol. Biochem. 2006, 70, 926–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Z.-W.; Zhang, J.; Townsend, D.M.; Tew, K.D. Oxidative Stress, Redox Regulation and Diseases of Cellular Differentiation. Biochim. Piophys. Acta. 2015, 1850, 1607–1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hotamisligil, G.S.; Davis, R.J. Cell Signaling and Stress Responses. Cold Spring Harb Perspect. Biol. 2016, 8, a006072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, R.R.; McDonald, L.T.; Jensen, N.R.; Sidles, S.J.; LaRue, A.C. Impacts of Psychological Stress on Osteoporosis: Clinical Implications and Treatment Interactions. Front. Psychiatry 2019, 10, 200. [Google Scholar] [CrossRef]
- Naot, D.; Bava, U.; Matthews, B.; Callon, K.E.; Gamble, G.D.; Black, M.; Song, S.; Pitto, R.P.; Cundy, T.; Cornish, J.; et al. Differential gene expression in cultured osteoblasts and bone marrow stromal cells from patients with Paget’s disease of bone. J. Bone Miner. Res. 2007, 22, 298–309. [Google Scholar] [CrossRef]
- Fan, Y.; Liu, J.; Wang, S.; Wang, H.; Shi, F.; Xiong, L.; He, W.; Peng, X. Functional proteome of bones in rats with osteoporosis following ovariectomy. Life Sci. 2005, 76, 2893–2901. [Google Scholar] [CrossRef]
- Iglesias-Gato, D.; Thysell, E.; Tyanova, S.; Crnalic, S.; Santos, A.; Lima, T.S.; Geiger, T.; Cox, J.; Widmark, A.; Bergh, A.; et al. The proteome of prostate cancer bone metastasis reveals heterogeneity with prognostic implications. Clin. Cancer Res. 2018, 24, 5433–5444. [Google Scholar] [CrossRef] [Green Version]
- Dowling, P.; Hayes, C.; Ting, K.R.; Hameed, A.; Meiller, J.; Mitsiades, C.; Anderson, K.C.; Clynes, M.; Clarke, C.; Richardson, P.; et al. Identification of proteins found to be significantly altered when comparing the serum proteome from Multiple Myeloma patients with varying degrees of bone disease. BMC Genomics 2014, 15, 904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soeta, S.; Higuchi, M.; Yoshimura, I.; Itoh, R.; Kimura, N.; Amasaki, H. Effects of Vitamin E on the Osteoblast Differentiation. J. Vet. Med. Sci. 2010, 72, 951–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, A.C.; Poulton, I.J.; Vrahnas, C.; Hausler, D.K.; Walkley, C.R.; Wu, J.Y.; Martin, T.J.; Gillespie, M.T.; Chandraratna, R.A.; Quinn, J.M.; et al. RARγ is a negative regulator of osteoclastogenesis. J. Steroid Biochem. Mol. Biol. 2015, 150, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Vester, H.; Holzer, N.; Neumaier, M.; Schyschka, L.; Nusser, A.K.; Seeliger, C. Green Tea Extract (GTE) improves differentiation in human osteoblasts during oxidative stress. J. Inflamm. 2014, 11, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambrożewicz, E.; Muszyńska, M.; Tokajuk, G.; Grynkiewicz, G.; Žarković, N.; Skrzydlewska, E. Beneficial Effects of Vitamins K and D3 on Redox Balance of Human Osteoblasts Cultured with Hydroxyapatite-Based Biomaterials. Cells 2019, 8, 325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Przekora, A.; Ginalska, G. Enhanced differentiation of osteoblastic cells on novel chitosan/β-1,3-glucan/bioceramic scaffolds for bone tissue regeneration. Biomed. Mater. 2015, 10, 015009. [Google Scholar] [CrossRef]
- Neve, A.; Corrado, A. Osteoblast physiology in normal and pathological conditions. Cell Tissue Res. 2011, 343, 289–302. [Google Scholar] [CrossRef]
- Nakashima, K.; de Crombrugghe, B. Transcriptional mechanisms in osteoblast differentiation and bone formation. Trends Genet. 2003, 19, 458–466. [Google Scholar] [CrossRef]
- Komori, T. Regulation of osteoblast differentiation by transcription factors. J. Cell Biochem. 2006, 99, 1233–1239. [Google Scholar] [CrossRef]
- Marie, P.J. Transcription factors controlling osteoblastogenesis. Arch. Biochem. Biophys. 2008, 473, 98–105. [Google Scholar] [CrossRef]
- Lu, H.; Kraut, D.; Gerstenfeld, L.C.; Graves, D.T. Diabetes interferes with the bone formation by affecting the expression of transcription factors that regulate osteoblast differentiation. Endocrinology 2003, 144, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Hung, L.-M.; Tsai, C.-H.; Chen, J.-K. TGF-β1 selectively suppresses PDGF receptor signaling pathways in MG-63 human osteosarcoma cell. Life Sci. 1997, 61, 685–693. [Google Scholar] [CrossRef]
- Casati, L.; Celotti, F.; Negri-Cesi, P.; Sacchi, M.C.; Castano, P.; Colciago, A. Platelet derived growth factor (PDGF) contained in Platelet Rich Plasma (PRP) stimulates migration of osteoblasts by reorganizing actin cytoskeleton. Cell Adh. Migr. 2014, 8, 595–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imai, S.; Kaksonen, M.; Raulo, E.; Kinnunen, T.; Fages, C.; Meng, X.; Lakso, M.; Rauvala, H. Osteoblast Recruitment and Bone Formation Enhanced by Cell Matrix–associated Heparin-binding Growth-associated Molecule (HB-GAM). J. Cell Biol. 1998, 43, 1113–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florencio-Silva, R.; da Silva Sasso, G.R.; Sasso-Cerri, E.; Simões, M.J.; Cerri, P.S. Biology of Bone Tissue: Structure, Function, and Factors That Influence Bone Cells. BioMed Res. Int. 2015, 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eaton, G.J.; Zhang, Q.S.; Diallo, C.; Matsuzawa, A.; Ichijo, H.; Steinbeck, M.J.; Freeman, T.A. Inhibition of apoptosis signal-regulating kinase 1 enhances endochondral bone formation by increasing chondrocyte survival. Cell Death Dis. 2014, 5, e1522. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Ying, J.; Li, H.; Zhang, Y.; Shu, X.; Fan, Y.; Tan, J.; Cao, Y.; Tsao, S.W.; Srivastava, G.; et al. The human cadherin 11 is a pro-apoptotic tumor suppressor modulating cell stemness through Wnt/β-catenin signaling and silenced in common carcinomas. Oncogene 2012, 31, 3901–3912. [Google Scholar] [CrossRef] [Green Version]
- Sunjic, S.B.; Cipak, A.; Rabuzin, F.; Wildburger, R.; Zarkovic, N. The influence of 4-hydroxy-2-nonenal on proliferation, differentiation and apoptosis of human osteosarcoma cells. Biofactors 2005, 24, 141–148. [Google Scholar] [CrossRef]
- Chang, J.K.; Li, C.J.; Liao, H.J.; Wang, C.K.; Wang, G.J.; Ho, M.L. Anti-inflammatory drugs suppress proliferation and induce apoptosis through altering expressions of cell cycle regulators and pro-apoptotic factors in cultured human osteoblasts. Toxicology 2009, 258, 148–156. [Google Scholar] [CrossRef]
- Sharma, A.R.; Choi, B.-S.; Park, J.-M.; Lee, D.-H.; Lee, J.-E.; Kim, H.-S.; Yoon, J.-K.; Song, D.-K.; Nam, J.-S.; Lee, S.-S. Rspo1 promotes osteoblast differentiation via Wnt signaling pathway. Indian J. Biochem. Biophys. 2013, I50, 19–25. [Google Scholar]
- Jiang, S.Y.; Shu, R.; Xie, Y.F.; Zhang, S.Y. Age-related changes in biological characteristics of human alveolar osteoblasts. Cell Prolif. 2010, 43, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Rishi, A.K.; Zhang, L.; Jiang, Y.; Nautiyal, J.; Wali, A.; Fontana, J.A.; Levi, E.; Majumdar, A.P. Cell cycle- and apoptosis-regulatory protein-1 is involved in apoptosis signaling by epidermal growth factor receptor. J. Biol. Chem. 2006, 281, 13188–13198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ou, C.-Y.; Kim, J.H.; Yang, C.K.; Stallcup, M.R. Requirement of Cell Cycle and Apoptosis Regulator 1 for Target Gene Activation by Wnt and β-Catenin and for Anchorage-independent Growth of Human Colon Carcinoma Cells*S. J. Biol. Chem. 2009, 284, 20629–20637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, V.; Bryant, H.U.; MacDougald, O.A. Regulation of bone mass by Wnt signaling. J. Clin. Invest. 2006, 116, 1202–1209. [Google Scholar] [CrossRef] [PubMed]
- Baron, R.; Rawadi, G. Wnt signaling and the regulation of bone mass. Curr. Osteoporosis Rep. 2007, 5, 73–80. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, B.; Zhang, A.; Zhao, Y.; Zhao, J.; Liu, J.; Gao, J.; Fang, D.; Rao, Z. Synergistic growth inhibition by sorafenib and vitamin K2 in human hepatocellular carcinoma cells. Clinics 2012, 67, 1093–1099. [Google Scholar] [CrossRef]
- Elefteriou, F. Neuronal signaling and the regulation of bone remodeling. Cell Mol. Life Sci. 2005, 62, 2339–2349. [Google Scholar] [CrossRef]
- Wu, X.; Itoh, N.; Taniguchi, T.; Nakanishi, T.; Tanaka, K. Requirement of calcium and phosphate ions in expression of sodium-dependent vitamin C transporter 2 and osteopontin in MC3T3-E1 osteoblastic cells. Biochim. Biophys. Acta. 2003, 1641, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Krueger, J.J.; Redmon, S.N.; Uppuganti, S.; Nyman, J.S.; Hahn, M.K.; Elefteriou, F. Extracellular norepinephrine clearance by the norepinephrine transporter is required for skeletal homeostasis. J. Biol. Chem. 2013, 288, 30105–30113. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.Q.; Liu, W.; Cui, R.R.; Wang, D.; Meng, J.C.; Xie, H.; Wu, X.P.; Zhou, H.D.; Lu, Y.; Liao, E.Y. Taurine inhibits osteoclastogenesis through the taurine transporter. Amino Acids 2010, 39, 89–99. [Google Scholar] [CrossRef]
- Seidlitz, E.P.; Sharma, M.K.; Singh, G. Extracellular glutamate alters mature osteoclast and osteoblast functions. Can. J. Physiol. Pharmacol. 2010, 88, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Sen, B.; Xie, Z.; Uzer, G.; Thompson, W.R.; Styner, M.; Wu, X.; Rubin, J. Intranuclear Actin Regulates Osteogenesis. Stem Cells 2015, 33, 3065–3076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dopie, J.; Skarp, K.-P.; Rajakyla, E.K.; Tanhuanpaa, K.; Vartiainen, M.K. Active maintenance of nuclear actin by importin 9 supports transcription. Proc. Natl. Acad. Sci. USA. 2012, 109, E544–E552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, W.-F.; Tang, F.L.; Xiong, L.; Xiong, S.; Jung, J.U.; Lee, D.-H.; Li, X.-S.; Feng, X.; Mei, L.; Xiong, W.-C. Vps35 loss promotes hyperresorptive osteoclastogenesis and osteoporosis via sustained RANKL signaling. J. Cell Biol. 2013, 200, 821–837. [Google Scholar] [CrossRef] [Green Version]
- Xiong, M.-Y.; Liu, L.-Q.; Liu, S.-Q.; Liu, Z.-H.; Gap, H.-F. Effects of osteoprotegerin, RANK and RANKL on bone destruction and collapse in avascular necrosis femoral head. Am. J. Transl. Res. 2016, 8, 3133–3140. [Google Scholar]
- Mollazadeh, S.; Bazzaz, B.S.F.; Kerachian, M.A. Role of apoptosis in pathogenesis and treatment of bone-related diseases. J Orthop. Surg. Res. 2015, 10, 15. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Moseman, E.A.; Farrar, M.A.; Bachanova, V.; Weisdorf, D.J.; Blazar, B.R.; Chen, W. Toll-like receptor 9 signaling by CpG-B oligodeoxynucleotides induces an apoptotic pathway in human chronic lymphocytic leukemia B cells. Blood 2010, 115, 5041–5052. [Google Scholar] [CrossRef] [Green Version]
- Westphal, D.; Dewson, G.; Czabotar, P.E.; Kluck, R.M. Molecular biology of Bax and Bak activation and action. Biochim. Biophys. Acta. 2011, 1813, 521–531. [Google Scholar] [CrossRef] [Green Version]
- Urayama, S.; Kawakami, A.; Nakashima, T.; Tsuboi, M.; Yamasaki, S.; Hida, A.; Ichinose, Y.; Nakamura, H.; Ejima, E.; Aoyagi, T.; et al. Effect of vitamin K2 on osteoblast apoptosis: Vitamin K2 inhibits apoptotic cell death of human osteoblasts induced by Fas, proteasome inhibitor, etoposide, and staurosporine. J. Lab. Clin. Med. 2000, 136, 181–193. [Google Scholar] [CrossRef]
- Ramirez, A.; Vazquez-Sanchez, A.Y.; Carrion-Robalino, N.; Camacho, J. Ion Channels and Oxidative Stress as a Potential Link for the Diagnosis or Treatment of Liver Diseases. Oxid. Mex. Cell Longev. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Stains, J.P.; Weber, J.A.; Gay, C.V. Expression of Na(+)/Ca(2+) exchanger isoforms (NCX1 and NCX3) and plasma membrane Ca(2+) ATPase during osteoblast differentiation. J. Cell Biochem. 2002, 84, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Prasad, V.; Hyung, S.-W.; Lee, Z.H.; Lee, S.-W.; Bhargava, A.; Pearce, D.; Lee, Y.; Kim, H.-H. Plasma membrane calcium ATPase regulates bone mass by fine-tuning osteoclast differentiation and survival. J. Cell Biol. 2012, 199, 1145–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czekanska, E.M.; Stoddart, M.J.; Richards, R.G.; Hayes, J.S. In search of an osteoblast cell model for in vitro research. Eur. Cell Mater. 2012, 24, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Tomas, H.; Havli, J.; Olsen, J.V.; Mann, M. In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 2006, 1, 2856–2860. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhu, G.; Yan, X.; Champion, M.M. Capillary zone electrophoresis for analysis of complex proteomes using an electrokinetically pumped sheath flow nanospray interface. Proteomics 2013, 14, 622–628. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds—phylloquinone (K1) and menaquinones (K2: MK4, MK7)...... are available from the authors. |



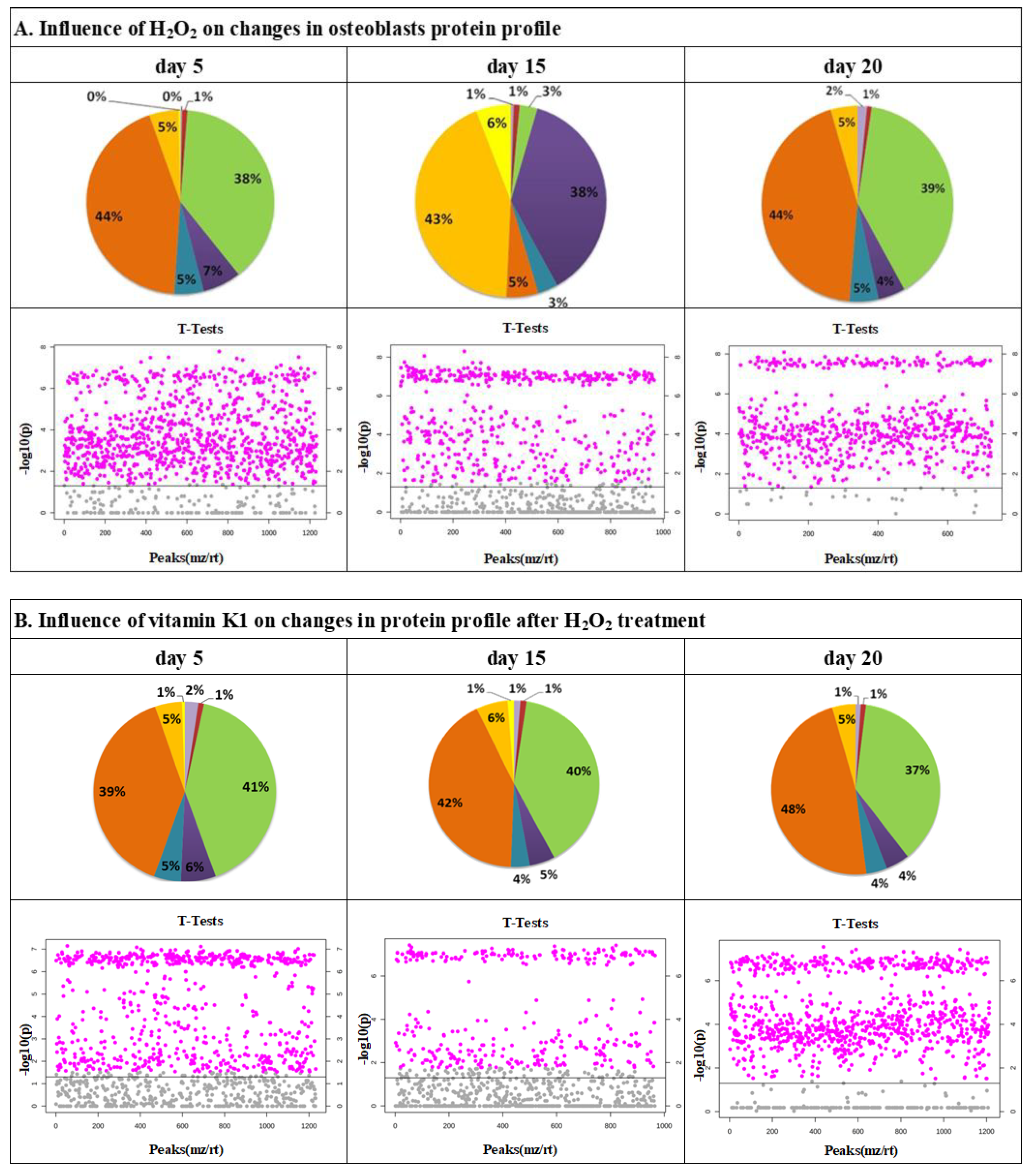{kind=link}
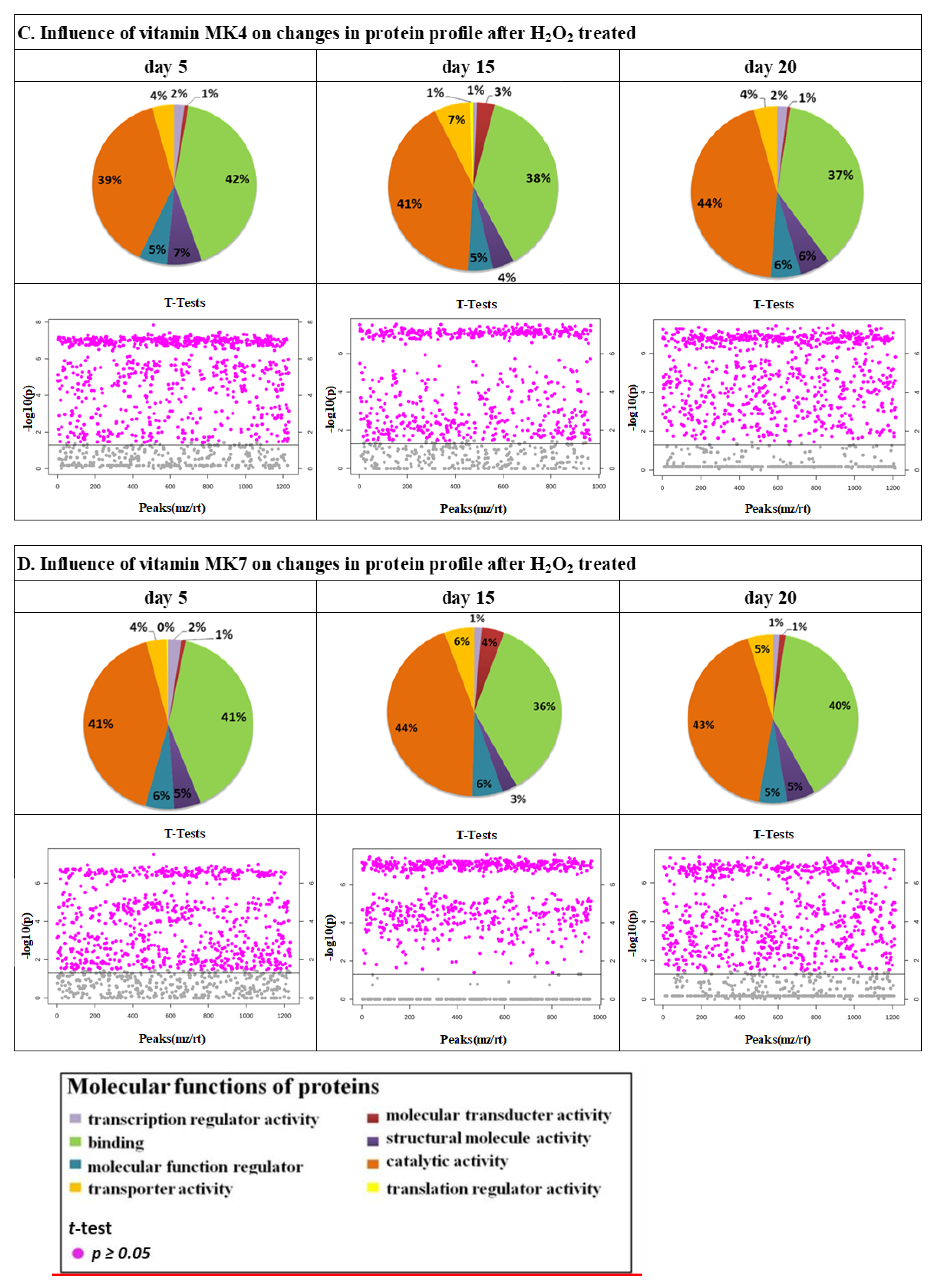{kind=link}
| ID | Protein Name | H2O2 vs. Control | H2O2 + K1 vs. H2O2 | H2O2 + MK4 vs. H2O2 | H2O2 + MK7 vs. H2O2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Days | |||||||||||||
| 5 | 15 | 20 | 5 | 15 | 20 | 5 | 15 | 20 | 5 | 15 | 20 | ||
| P05023 | Na/K-transporting ATPase subunit α1 | ↓ | ↓ | ||||||||||
| P38606 | v-type proton ATPase catalytic subunit A | ↓ | ↓ | ↓ | ↑ | ↑ | ↓ | ↑ | |||||
| O14983 | Sarcoplasmic/endoplasmic reticulum calcium ATPase 1 | ↑ | ↑ | ↑ | ↓ | ||||||||
| P20020 | Plasma membrane Ca-transporting ATPase | ↓ | ↑ | ||||||||||
| O95373 | Importin-7 | ↑ | ↓ | ↓ | |||||||||
| Q96P70 | Importin-9 | ↑ | ↓ | ↓ | |||||||||
| Q14974 | Importin subunit beta-1 | ↑ | ↓ | ||||||||||
| P55060 | Exportin-2 | ↑ | ↓ | ↑ | ↓ | ↓ | |||||||
| Q96QK1 | Vacuolar protein sorting-associated protein35 | ↑ | ↑ | ↓ | ↓ | ↓ | ↓ | ↓ | |||||
| Q15758 | Neutral amino acid transporter B(0) | ↑ | ↓ | ↓ | ↑ | ↑ | ↓ | ↓ | ↓ | ↓ | |||
| Q8WVM8 | Sec1 family domain-containing protein 1 | ↑ | ↓ | ↓ | ↓ | ↑ | ↑ | ↑ | ↓ | ↑ | |||
| O60518 | Ran-binding protein 6 | ↑ | ↓ | ↑ | ↓ | ↓ | ↓ | ||||||
| Q00325 | Phosphate carrier protein, mitochondrial | ↑ | ↓ | ↑ | ↑ | ||||||||
| Q00765 | Receptor expression-enhancing protein 5 | ↓ | ↓ | ↓ | ↓ | ||||||||
| Q02978 | Mitochondrial 2-oxoglutarate/malate carrier protein | ↓ | ↓ | ↓ | ↑ | ↑ | ↑ | ↑ | |||||
| Q07812 | Bax | ↑ | ↓ | ↓ | ↓ | ||||||||
| P60468 | Protein transport protein Sec61 subunit beta | ↑ | ↓ | ↑ | |||||||||
| O75746 | Ca-binding mitochondrial carrier protein Aralar1 | ↑ | ↑ | ↓ | |||||||||
| Q8N4V1 | Membrane magnesium transporter 1 | ↑ | |||||||||||
| Q99523 | Sortilin | ↑ | ↓ | ↑ | |||||||||
| ID | Protein Name | H2O2 vs. Control | H2O2 + K1 vs. H2O2 | H2O2 + MK4 vs. H2O2 | H2O2 + MK7 vs. H2O2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Days | |||||||||||||
| 5 | 15 | 20 | 5 | 15 | 20 | 5 | 15 | 20 | 5 | 15 | 20 | ||
| O75475 | PC4 and SFRS1-interacting protein | ↑ | ↓ | ↑ | ↓ | ||||||||
| Q14247 | Src substrate cortactin | ↑ | ↓ | ↑ | ↑ | ||||||||
| Q8N163 | Cell cycle and apoptosis regulator protein 1 | ↑ | ↓ | ↓ | ↓ | ||||||||
| B2RPK0 | Putative high mobility group protein B1 | ↓ | ↓ | ↓ | |||||||||
| P13489 | Ribonuclease inhibitor | ↓ | ↑ | ↑ | ↑ | ||||||||
| E9PA3 | Nascent polypeptide-associated complex | ↓ | ↑ | ↑ | ↓ | ||||||||
| Q13263 | Transcription intermediary factor 1-β | ↑ | ↑ | ↓ | ↓ | ↓ | ↑ | ||||||
| Q07666 | KH domain-containing, RNA-binding, signal transduction-associated protein 1 | ↓ | ↓ | ↓ | |||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muszyńska, M.; Ambrożewicz, E.; Gęgotek, A.; Grynkiewicz, G.; Skrzydlewska, E. Protective Effects of Vitamin K Compounds on the Proteomic Profile of Osteoblasts under Oxidative Stress Conditions. Molecules 2020, 25, 1990. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25081990
Muszyńska M, Ambrożewicz E, Gęgotek A, Grynkiewicz G, Skrzydlewska E. Protective Effects of Vitamin K Compounds on the Proteomic Profile of Osteoblasts under Oxidative Stress Conditions. Molecules. 2020; 25(8):1990. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25081990
Chicago/Turabian StyleMuszyńska, Marta, Ewa Ambrożewicz, Agnieszka Gęgotek, Grzegorz Grynkiewicz, and Elżbieta Skrzydlewska. 2020. "Protective Effects of Vitamin K Compounds on the Proteomic Profile of Osteoblasts under Oxidative Stress Conditions" Molecules 25, no. 8: 1990. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25081990