Inositol Pyrophosphate Pathways and Mechanisms: What Can We Learn from Plants?

Department of Biochemistry, Virginia Tech, Blacksburg, VA 24061, USA
*
Author to whom correspondence should be addressed.
Molecules 2020, 25(12), 2789; https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25122789
Submission received: 1 May 2020
/
Revised: 11 June 2020
/
Accepted: 13 June 2020
/
Published: 17 June 2020
(This article belongs to the Special Issue Bioactivity of Inositol Phosphates)
Abstract
:The ability of an organism to maintain homeostasis in changing conditions is crucial for growth and survival. Eukaryotes have developed complex signaling pathways to adapt to a readily changing environment, including the inositol phosphate (InsP) signaling pathway. In plants and humans the pyrophosphorylated inositol molecules, inositol pyrophosphates (PP-InsPs), have been implicated in phosphate and energy sensing. PP-InsPs are synthesized from the phosphorylation of InsP6, the most abundant InsP. The plant PP-InsP synthesis pathway is similar but distinct from that of the human, which may reflect differences in how molecules such as Ins(1,4,5)P3 and InsP6 function in plants vs. animals. In addition, PP-InsPs can potentially interact with several major signaling proteins in plants, suggesting PP-InsPs play unique signaling roles via binding to protein partners. In this review, we will compare the biosynthesis and role of PP-InsPs in animals and plants, focusing on three central themes: InsP6 synthesis pathways, synthesis and regulation of the PP-InsPs, and function of a specific protein domain called the Syg1, Pho1, Xpr1 (SPX ) domain in binding PP-InsPs and regulating inorganic phosphate (Pi) sensing. This review will provide novel insights into the biosynthetic pathway and bioactivity of these key signaling molecules in plant and human systems.
{kind=link}
{kind=link}
{kind=link}
1. Introduction
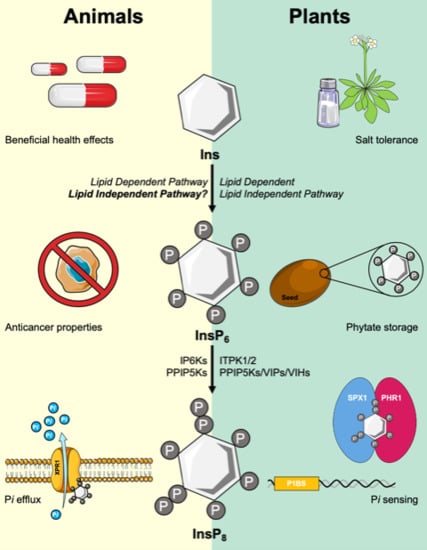
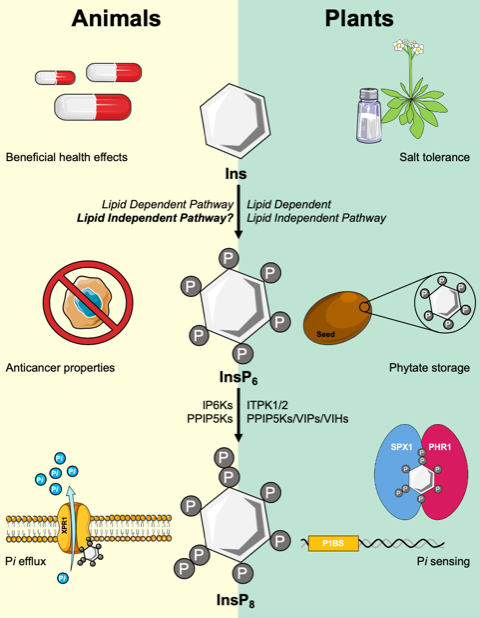
Inositol phosphates (InsPs) are a widely known family of signaling molecules, with conserved functions in numerous cellular processes in eukaryotes, many of which are related to human health [1,2,3]. The InsP family of molecules consists of monophosphorylated inositol (InsP) to the fully phosphorylated InsP6 (inositol hexakisphosphate), and are generated by InsP kinases that reversibly add phosphates to specific positions of the 6-carbon inositol ring backbone [2,4]. The specific number and position of phosphates on the inositol ring is thought to confer specific information within the cell, thus InsPs are thought to function as a type of chemical signaling language first described in [5], and elaborated on in plants [6]. In addition, InsPs can be further phosphorylated to produce the high-energy inositol pyrophosphates (PP-InsPs), which while only found in low concentrations in the cell, have been implicated in an array of developmental, metabolic and signaling processes [7]. Given the emerging role of PP-InsPs in controlling eukaryotic signaling pathways, it is important to consider how disrupting Ins and InsP biosynthesis for therapeutic uses impacts PP-InsPs and the processes they mediate.
One significant difference between plants and animals is the large phosphate (P) storage capacity of InsP6 present in seeds. In the model plant Arabidopsis thaliana, for example, InsP6 comprises ~1% of seed dry weight [8]. This large pool of stored P is important for maintaining metabolism between the phases of seed germination and for the onset of efficient photosynthesis [9]. A second potentially critical difference is the presence of a known InsP6 transporter in plant cells called Multidrug Resistance Protein 5 (MRP5) [10]. The MRP5 gene product of Arabidopsis encodes an ABC transporter that most likely functions to transport InsP6 across the vacuolar membrane during seed development, allowing high amounts of InsP6 to be stored in the seed [10]. At present we have a good understanding of the synthesis and transport of InsP6 within plants, due to the presence of key Arabidopsis mutants that are defective in InsP kinases [11] or the Atmrp5 gene [10]. A third major difference is that plants have no known Ins(1,4,5)P3 receptor that could serve to release intracellular calcium stores in response to signaling-stimulated hydrolysis of phosphatidylinositol(4,5)P2 (PtdIns(4,5)P2) [12]. These key differences highlight the fact that plants and animals likely had different pressures driving evolution of central genes in the InsP pathway. The purpose of this review is to compare knowledge on the InsP and inositol pyrophosphate (PP-InsP) pathways in humans and plants, and highlight how new information on the plant InsP pathway may provide insights into the function of this pathway in other eukaryotes. In this review, we will concentrate on three central themes for our comparison of plants and humans: InsP6 synthesis pathways, synthesis and regulation of the PP-InsPs, and function of a specific protein domain called the SPX domain in binding PP-InsPs and regulating inorganic phosphate (Pi) sensing.
2. Myo-Inositol: The Signaling Scaffold and Precursor for the Lipid Independent Pathway
Similar to other eukaryotes, the scaffold for InsPs is the most abundant inositol isomer in plants, the so-called myo-inositol isomer. Herein, we will refer to myo-inositol as “inositol”. Plants also contain D-chiro, epi-, neo-, muco-, and allo-inositol isomers in minor amounts [13], and these may be important for various physiological events. For example, the presence of D-chiro-inositol has been linked to salt stress tolerance in plants [14,15]. Intriguingly, a lack of D-chiro-inositol has been correlated with various diseases in humans, such as polycystic ovary syndrome and type 2 diabetes [16,17,18,19,20,21]. It is also interesting to note that groups have proposed supplementing patient diets with D-chiro-inositol rich foods and supplements as a form of treatment [22].
3. Is the Lipid-Independent InsP6 Synthetic Pathway Plant Specific?
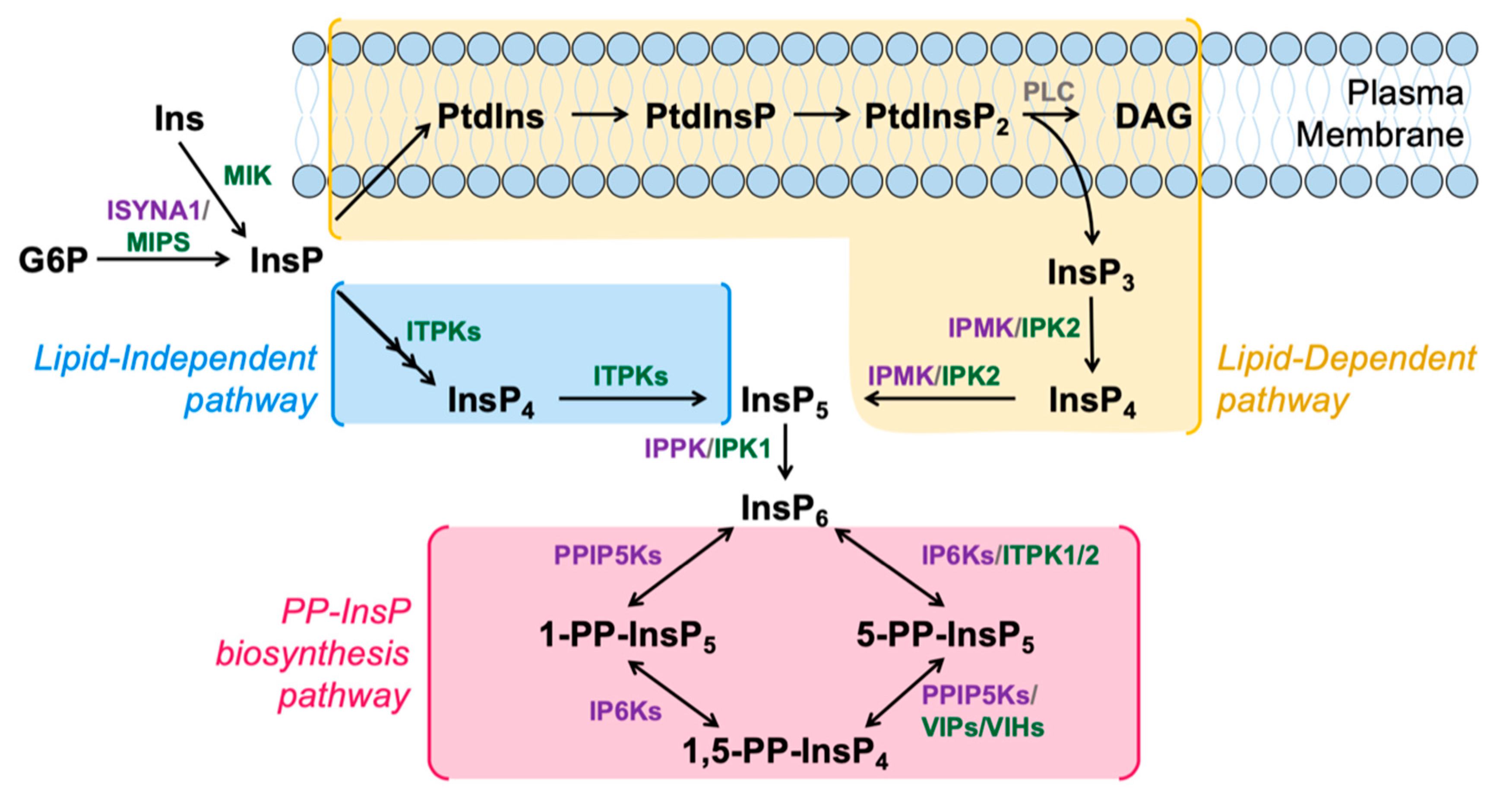
A canonical InsP6 synthesis pathway is shared between plants and animals, and is usually referred to as the “Lipid-Dependent” pathway, as the first step is initiated by the hydrolysis of PtdIns(4,5)P2 into Ins(1,4,5)P3 and diacylglycerol (DAG) [23,24,25] (Figure 1). This Ins(1,4,5)P3 product can be sequentially phosphorylated by two sets of enzymes in both plants and animals, namely the Inositol polyphosphate Multikinases (IPMK; EC 2.7.1.151), also known as IPK2 in plants, and the Inositol Pentakisphosphate 2-Kinase (IPK1; EC 2.7.1.158) [23,24,26].
InsP6 can also be synthesized from the phosphorylation of inositol by a series of InsP kinases in a pathway previously thought to be unique to plants and slime molds: the “Lipid Independent” pathway of InsP6 synthesis (Figure 1). In this pathway, inositol is first acted on by an enzyme named myo-Inositol Kinase (MIK; EC 2.7.1.64). The Arabidopsis genome encodes one copy of the Mik gene, and the MIK enzyme is known to phosphorylate the 3-position of inositol [27,28]. Atmik mutants have a significant reduction in InsP6 seed levels, indicating that a portion of seed InsP6 requires the function of MIK. However, InsP6 levels in vegetative tissues from Atmik mutants have not yet been reported [27]. In maize (corn), mik knockout mutants accumulate 50% less seed InsP6, and contain a reduction in other InsPs [28], so the function of MIK in seed InsP6 synthesis is thought to be conserved across various plant species. The human genome, in contrast, does not contain a highly homologous MIK orthologue although one human ribokinase and several adenokinases contain limited regions of homology to MIK. We note that animals may not require MIK for a lipid -independent pathway as the myo-inositol phosphate synthase (MIPS; EC 5.5.1.4) can convert Glucose-6-P to Inositol 3-P [29], bypassing the need for MIK (Figure 1). An alternative is that there is a human MIK that is distinctly different compared to the plant MIK.
In plants, the enzyme thought to be responsible for phosphorylating InsP to InsP2, which was first isolated as a low phytic acid (lpa) mutant in rice and named LPA1, also lacks a clear human orthologue. Exciting new data indicate that both plants and animals have a way to directly phosphorylate Ins 3-P, without using an LPA1 enzyme [30]. Specifically, the Inositol 1,3,4-Trisphosphate 5/6-Kinases (ITPKs; EC 2.7.1.159), are enzymes that are well-conserved in plants and humans and have been examined previously for their ability to convert InsP3 isomers to InsP4 and InsP5 [31,32]. New data from Archaea suggested that other eukaryotes might be able to synthesize InsPs via a Lipid-Independent pathway using ITPK1 as a highly promiscuous InsP kinase [30]. In this study the authors identified 4 genes homologous to ITPK1 in the Archaea Lokiarchaeota candidatus, sharing an ATP-grasp domain and conserved regions in the InsP- and ATP-binding sites, yet containing a more restricted InsP binding site [30]. Using both enzyme assays and yeast complementation assays, the authors showed HsITPK1 and AtITPK1 can phosphorylate Ins(1)P and Ins(3)P to InsP5 in vitro and in vivo [30]. Plants also contain four ITPK genes, which exist as part of a multigene family. It is interesting to note that only two of the four Arabidopsis itpk loss-of-function mutants have reduced seed InsP6 levels, suggesting redundancy of the plant genes [27].
The last step in the lipid-independent pathway, conversion of Ins(1,3,4,5,6)P5 to InsP6 is catalyzed by another very well-conserved enzyme in eukaryotes, named Inositol Pentakisphosphate 2-Kinase (IPK1; EC 2.7.1.158) in both plants and animals [33,34]. Atipk1 loss-of-function mutants have an 83% reduction in InsP6 levels in seeds, in addition to a reduction in InsP7 and InsP8 [24,35]. The human IPK1, named HsIPPK, can be knocked down, and these mutants accumulate InsP4 at the expense of InsP6 levels [34], which is a notable difference from the situation in Atipk1 mutants.
4. Catalytic Flexibility in Plant and Human ITPKs
We have already described how the plant and human ITPKs are capable of accepting multiple InsP substrates. This class of enzymes also has another unexpected role in plants. In non-plant eukaryotes, InsP6 kinases (inositol-hexakisphosphate kinase; IP6K; EC 2.7.4.21) catalyze the synthesis of InsP6 and InsP7 from InsP5 and InsP6, respectively [36]. However, orthologous genes encoding IP6Ks are notably absent in investigated plant genomes [37]. When searching for the enzymes responsible for the initial step in PP-InsP synthesis, conversion of InsP6 to InsP7, our group and others focused on the ITPKs, which are able to catalyze the sequential conversion of InsP3 to InsP4 and InsP5. The Arabidopsis genome contains four ITPK genes (Itpk1, Itpk2, Itpk3, and Itpk4), which encode ATP-grasp domain-containing proteins [31,38]. In humans, a sole HsItpk gene encodes an ATP-grasp containing ITPK enzyme, which shares 30% sequence similarity to its Arabidopsis counterpart [31,39]. HsITPK1 is responsible for the production of Ins(1,3,4,6)P4 and Ins(1,3,4,5)P4 in a ratio of 3:1, whereas AtITPKs produce the same products in the 1:3 ratio, hinting at a functional divergence in the InsP synthetic pathways in animals and plants [31,38]. Furthermore, both HsITPK1 and AtITPKs can phosphorylate InsP4 at the 1-position to produce InsP5.
Our biochemical analysis has demonstrated that both AtITPK1 and AtITPK2 are able to phosphorylate InsP6 in vitro to produce a more phosphorylated product [40]. This novel activity of AtITPKs is also reported in other recent studies, identifying the product of InsP6 phosphorylation by AtITPK1/2 as 5PP-InsP5 [41]. The high catalytic flexibility of plant ITPKs has likely evolved due to the paucity of canonical InsP3 3-kinases (IP3Ks) [42] and IP6Ks. In animals, dedicated IP3K and IP6K families are responsible for the phosphorylation of InsP3 and InsP6, respectively [43]. Given the notable lack of IP6Ks and the closely related IP3K in plants, it is conceivable to hypothesize that AtITPKs have evolved to expand their substrate specificity to accommodate several diverse substrates. We note here that the kinase domains of the AtITPKs share structural similarity to IP3Ks and to the diphosphoinositol-pentakisphosphate kinases (PPIP5Ks; EC 2.7.4.21), despite limited sequence identity [39,44].
5. The PPIP5K/VIP/VIH Dual Domains are Arbiters of Biology
One focus of our lab is on the highly conserved PPIP5Ks, which are also known as VIP or VIH enzymes in plants. Until recently it was suspected that the PPIP5Ks in plants functioned in two steps to convert InsP6 to the first inositol pyrophosphate, called 5PP-InsP5 or InsP7, and also to convert InsP7 to 1,5PP2-InsP4 or InsP8. As mentioned before, recent work has shown the likelihood that this is not true, and that plant ITPKs are responsible for conversion of InsP6 to InsP7 [40]. This leaves the plant PPIP5K enzymes as functioning in a very similar role as the animal PPIP5Ks. Indeed both the human PPIP5Ks and the plant PPIP5Ks contain a conserved structure: an N-terminal ATP-grasp kinase domain (KD) and a C-terminal phosphatase domain (PD) [45]. Like humans, Arabidopsis contains two Ppip5k genes orthologous to the mammalian Ppip5ks, and both AtPpip5k genes encode catalytically active enzymes, sharing a conserved Asp residue in the KD, which is necessary for kinase activity [37,45,46]. In addition, AtPPIP5Ks and HsPPIP5Ks both share a conserved Pleckstrin-homology (PH) domain, which interrupts the C-terminal PD [47]. PH domains are found in signaling proteins and bind phospholipids and molecules derived from phospholipid head groups [48]. The human PPIP5K1 PH domain is known to bind PtdIns(3,4,5)P3 preferentially, and can also bind InsP6 and PtdInsP2 [49]. PH domain-containing proteins are known to interact with InsP7, with the inositol pyrophosphate competing with PtdInsP2 and PtdInsP3 molecules for binding [47,50,51]. A conserved arginine residue (Arg417 in HsPPIP5K1) is required for ligand binding in the PH domain in human PPIP5Ks [47]. This arginine residue is not conserved in Arabidopsis PPIP5Ks, however, the substituted lysine residue carries a similar charge [47]. This could indicate that the AtPPIP5K PH domain has a similar phospholipid-binding affinity, however, analysis on ligand binding has not yet been assessed in plants. In addition to sharing a conserved dual domain structure, the AtPPIP5Ks and HsPPIP5Ks also share a C-terminal intrinsically disordered region (IDR) [52,53]. IDRs are involved in protein-protein interactions, and the HsPPIP5K IDRs have been implicated in protein scaffolding and interacting with proteins involved in lipid metabolism, vesicle-mediated transport and actin cytoskeleton organization [52,53]. While a number of proteins interacting with the disordered regions in the PPIP5K family have been identified, the function of these protein-protein interactions and their involvement in PP-InsP metabolism remains unknown.
The presence of two competing domains (i.e. the KD and PD) is a unique feature of the PPIP5K enzymes, which has been discussed in detail [52]. In metabolism this is usually referred to as a futile cycle, in that the substrate, InsP7, for one domain (the KD), is also the product of the other domain (the PD). This dual domain structure is found in all eukaryotic PPIP5K enzymes except for those from the ancestral slime mold, Dictyostelium discoideum [52], which lacks a PD, either within the protein or as an independent gene, explaining the slime mold’s ability to synthesize unusually high levels of InsP7 and InsP8 [54]. While the protein structures of the HsPPIP5Ks, AtPPIP5Ks or their orthologues have not yet been elucidated, we do know that the PD of HsPPIP5Ks and the yeast orthologue, Asp1, have 1-phosphatase activity, acting on the 1PP in an InsP7 or an InsP8 substrate [55,56,57,58]. Therefore, it seems likely that the function of the PPIP5K enzymes is to interconvert InsP7 and InsP8 signaling molecules, and thus control specific information within the cell. Gu et al., performed a detailed analysis of the kinetics of the human PPIP5Ks [56]. To discern the balance between activities of the KD and PD, these authors mutated each domain within the full-length enzyme, and tested activity of each with respect to physiologically relevant substrate concentrations. Their results indicated that the PD can restrict the maximal activity of the KD under certain conditions. Interestingly, a human mutation in the PD associated with elevated PPIP5K kinase activity and autosomal recessive nonsyndromic hearing loss indicates that the KD-PD balance is required in certain instances [56]. Gu et al. also found that inorganic phosphate (Pi) is a strong inhibitor of the PD of the HsPPIP5K2 isoform [56]. This last part is of enormous importance as PP-InsPs are known to serve as the proxy for Pi sensing in different eukaryotes, including plants [59,60].
Recent work illustrates that the balance between the KD and PD is crucial for controlling growth, development and Pi sensing in plants. Zhu et al. (2019) constructed transgenic Arabidopsis plants carrying a copy of the full-length PPIPK (VIP1/VIH2) protein harboring a known mutation either in the KD or PD that eliminates activity of that domain [61]. The authors compared these plants to those overexpressing the intact PPIPK (VIP1/VIH2) protein–(active KD and PD), finding that the intact PPIPK plants did not show greatly perturbed growth, development or Pi sensing, as measured by the accumulation of Pi within the plant [61]. In contrast, transgenic plants overexpressing PPIPK (VIP1/VIH2) with a mutated KD are compromised in plant growth and have greatly increased Pi accumulation, consistent with an elevated phosphate starvation response or PSR [61]. Transgenic plants overexpressing PPIPK (VIP1/VIH2) with a mutated PD also have less overall plant growth, accompanied by less Pi accumulation in tissues, suggesting these plants have a repressed PSR. Thus, an unbalanced PD activity appears to turn off Pi sensing, and an unbalanced KD activity appears to turn up Pi sensing. These data strongly support the idea that plants require InsP8 for sensing environmental Pi, and the PPIP5Ks function to increase or decrease InsP8 levels in response to fluctuating environmental Pi. Given this, it is imperative to understand whether or not Pi can also regulate the plant PPIP5K PD, as is the case for the human PPIP5Ks [56]. We note that the plant PPIP5K PD has been recalcitrant to enzyme studies, although the KD has been nicely characterized [40,41]. Notably, the single report of PD activity from plant PPIP5Ks required incubation for 16 hours to visualize even very modest activity, thus more work is needed to nail down the substrate specificity of this important domain [61].
Another powerful approach in plants, the use of genetic mutants, underscores the importance of InsP8 in Pi sensing. Arabidopsis mutants devoid of AtPPIP5K activity display significant growth phenotypes and defects in Pi sensing [61,62]. These mutants have increased levels of InsP7 and a reduction in InsP8 levels, and exhibit severely stunted growth with shorter roots and smaller leaves [61,62]. In addition, these mutants exhibit Pi-stress related phenotypes, including increased Pi accumulation and upregulation of PSR genes even under sufficient Pi growth conditions [62], Additionally, other mutants defective in InsP6 synthesis, itpk1, ipk1 and itpk4 mutants, exhibit an upregulation of the PSR under Pi-replete conditions [35,63]. Together, these data support a critical role for InsP8 in the regulation of Pi homeostasis in plants, a role which may be shared in humans. In humans, PP-InsPs are known to regulate certain transmembrane Pi-transport proteins, with ip6k1 ip6k2 double mutant HCT116 cells having a decreased rate of cellular Pi-efflux, strongly suggesting that PP-InsPs are required for Pi efflux [64].
6. Do Human Signaling Pathways Share a Common Mechanism for Regulation by InsP8?
Up to this point we have discussed new and critical data from plants regarding regulation of Pi sensing and the PSR by InsP8. It is important to describe the mechanism by which PP-InsPs are thought to influence signaling pathways, and to ask whether this mechanism is shared by humans.
PP-InsPs have been likened to a “molecular glue”, acting to facilitate protein-protein interactions and initiate energy- and nutrient-mediated responses. One protein domain found throughout eukaryotes that PP-InsPs bind to is named the SPX domain, named after the yeast proteins, Syg1 and Pho1, and the mammalian Xpr1. SPX-domains contain a basic surface of lysine residues that are known to coordinate the binding of PP-InsPs [65,66]. For many years it was known that the SPX domain basic surface could bind Pi. This, along with the identification of yeast and plant mutants defective in a variety of SPX domain-containing proteins, prompted the hypothesis that the SPX domain, itself, was the receptor for Pi sensing [67,68,69,70]. Wild et al. disrupted the thinking on this issue in 2016 by discovering that PP-InsPs can also serve as ligands for SPX domains, binding with high affinity (50 nM-100 μM) [65], thus making them better candidates for a so-called Pi sensing receptor. This group nicely showed that while isolated SPX domains can recognize 5-InsP7 and InsP6 with similar binding affinities, in the presence of their target transcription factor, plant SPX domains show a binding preference for PP-InsPs [65].
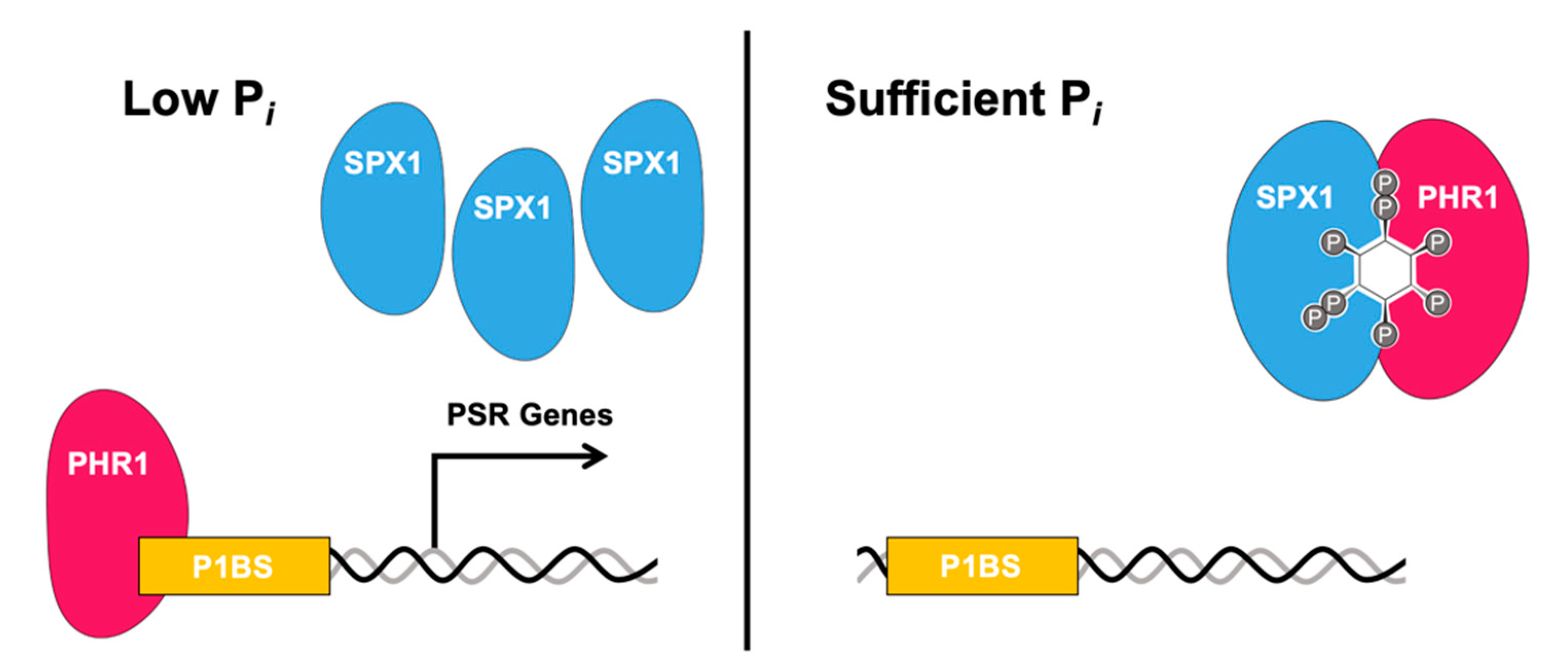
In plants, one SX domain-containing protein named SPX1 binds to PSR transcription factors, including PHR1 (Phosphate Starvation Response 1) and its homologs, which prevent the transcription factor from binding its target promoters under sufficient Pi conditions. Under low Pi conditions, the SPX1-PHR1 complex is unable to form, allowing the transcription factor to bind to its promoters and upregulate PSR genes [59,71] (Figure 2). PHR1 and PHL1 control the expression of a majority of the PSR genes, controlling numerous metabolic and development physiological adaptations to Pi deficiency. In rice, SPX proteins can associate with OsPHR2 in the presence of InsPs and PP-InsPs [65]. While initially Pi was proposed as the ligand for facilitating SPX-PHR complex formation, studies have identified InsPs, and more so PP-InsPs as bona fide ligands enabling this complex formation. The dissociation factor for Pi in the OsSPX4-OsPHR2 complex is over a factor greater than that of InsP6, while 5-InsP7 has the highest affinity of molecules tested, with a KD in the low micromolar range [65,72]. Recently, two groups have characterized PPIPK double mutants (vih1/vih2) mutants, which as previously described, have significantly reduced levels of InsP8 and compromised Pi signaling pathways under Pi-sufficient levels [64,71]. In vivo analysis indicated that even though the intracellular Pi concentration was high in vih1/vih2 mutants, the SPX1-PHR1 interaction was compromised [62]. Additionally, in vitro binding analysis showed both InsP7 and InsP8 bind SPX1 in the micromolar range, with InsP8 exhibiting a higher binding affinity than InsP7, further indicating the importance of PP-InsP binding in mediating SPX-protein interactions [62].
Xpr1 (Xenotropic and polytropic retrovirus receptor 1) is the only SPX-domain containing protein identified in the human genome [52,65]. Xpr1 mediates Pi export in humans, and mutations within the SPX-domain of Xpr1 are responsible for primary familial brain calcification (PFBC) [59]. PFBC is characterized by calcification, predominantly of calcium phosphate, in the basal ganglia [59]. The mutations within the SPX-domain of Xpr1 cause a reduction in Pi export efficiency [73]. Li et al. has recently described the role of InsP8 in XPR1-mediated Pi efflux, providing a novel understanding of the genetic factors involved in defective bone maintenance and ectopic mineralization [73]. This study nicely showed how a reduction in InsP8 reduces Pi export by XPR1 and accelerates calcification and mineralization [73].
While the mechanism of InsP8-mediated XPR1 regulation in humans differs from the SPX-PHR1 interactions observed in plants, we can speculate that InsP8, and possibly PP-InsPs in general, may play common roles, facilitating protein-protein interactions or protein-PP-InsP interaction in various signaling pathways in animals and plants. It is important to note that Arabidopsis encodes 20 SPX-domain containing proteins, and rice encodes 15 predicted SPX-containing proteins, with a majority predicted to be involved in Pi homeostasis [59], although not all SPX proteins bind to transcription factors. The greatly expanded family of SPX genes present in plants in comparison to SPX genes in humans, and their involvement in Pi metabolism is likely due to the importance of Pi sensing and homeostasis in plant health. Given the important role Pi plays in cellular metabolism, understanding the regulation of Pi homeostasis via PP-InsPs is a critical aspect of how bioenergetic responses are mediated under various stimuli.
7. Concluding Remarks
Elucidating the biosynthesis, regulation, and function of PP-InsPs is crucial to furthering our understanding of energy metabolism and nutrient sensing in eukaryotes. In this review, we addressed four major questions remaining in the InsP and PP-InsP fields in both humans and plants; do animals have a Lipid-Independent InsP6 pathway, what is the catalytic flexibility of ITPKs, how is the biosynthesis of PP-InsPs regulated and what role PP-InsPs play in signaling pathways. ITPK1 has been recently characterized as a highly flexible enzyme, capable of catalyzing the phosphorylation of InsP3, InsP4, and InsP6 in Arabidopsis, and InsP1 through InsP4 in both humans and plants [30,40,41]. The recent advances in characterizing ITPK1, a key enzyme in the eukaryotic PP-InsP pathway, and the identification of ITPK1 homologs in Archaea, has provided insights into the evolutionary development and catalytic flexibility of InsP/PP-InsP kinases [30]. Further genetic and biochemical work on the multigene families that encode plant InsP kinases may continue to shed light on the evolution of InsP and PP-InsP pathways in other eukaryotes. The production of InsP8 via the unique bifunctional PPIP5Ks is a highly regulated process, with both the KD and PD acting interdependently to regulate InsP7 and InsP8 levels [52]. Further understanding of the activity and regulation of the PD, along with whether the IDR facilitates protein-protein interactions, will be crucial to understanding Pi sensing and homeostasis in plants and humans alike. The interaction between PP-InsPs and SPX-domain containing proteins is a phenomenon shared in humans and plants. While plants have an expanded group of SPX proteins, humans have only one SPX-domain containing protein identified to date [52,65]. Given the shared PP-InsP:SPX interactions observed in humans and plants, we speculate that PP-InsPs interact and facilitate protein-protein interactions with yet to be identified SPX proteins or structurally similar protein families. Identifying and characterizing these putative protein families could lead to novel insights into energy metabolism in humans. Given the known role of Ins and InsPs in numerous human diseases, it is important to consider how novel therapeutics and drugs targeting Ins and InsP biosynthetic pathways may impact PP-InsPs and their cellular functions.
Author Contributions
Conceptualization, C.C. and G.G.; writing—original draft preparation, C.C., G.G.; writing—review and editing, C.C. and G.G. All authors have read and agree to the published version of the manuscript.
Funding
We gratefully acknowledge the NSF for funding to GG (MCB 1616038). This work is supported in part by the USDA National Institute of Food and Agriculture, Hatch project VA-136334, and the Institute for Critical and Applied Technology at Virginia Tech.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chakraborty, A.; Kim, S.; Snyder, S.H. Inositol pyrophosphates as mammalian cell signals. Sci. Signal. 2011, 4. [Google Scholar] [CrossRef] [Green Version]
- Hatch, A.J.; York, J.D. SnapShot: Inositol phosphates. Cell 2010, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Irvine, R.F. Inositide evolution-Towards turtle domination?: Inositide evolution. J. Physiol. 2005, 566, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Shears, S.B.; Wang, H. Inositol phosphate kinases: Expanding the biological significance of the universal core of the protein kinase fold. Adv. Biol. Regul. 2019, 71, 118–127. [Google Scholar] [CrossRef] [PubMed]
- York, J.D. Regulation of nuclear processes by inositol polyphosphates. Biochim. Biophys. Acta 2006, 1761, 552–559. [Google Scholar] [CrossRef]
- Gillaspy, G.E. The cellular language of myo-inositol signaling. New Phytol. 2011, 192, 823–839. [Google Scholar] [CrossRef]
- Shears, S.B. Intimate connections: Inositol pyrophosphates at the interface of metabolic regulation and cell signaling. J. Cell. Physiol. 2018, 233, 1897–1912. [Google Scholar] [CrossRef]
- Raboy, V. Seeds for a better future: “low phytate” grains help to overcome malnutrition and reduce pollution. Trends Plant Sci. 2001, 6, 458–462. [Google Scholar] [CrossRef]
- White, P.J.; Veneklaas, E.J. Nature and nurture: The importance of seed phosphorus content. Plant Soil 2012, 357, 1–8. [Google Scholar] [CrossRef]
- Nagy, R.; Grob, H.; Weder, B.; Green, P.; Klein, M.; Frelet-Barrand, A.; Schjoerring, J.K.; Brearley, C.; Martinoia, E. The Arabidopsis ATP-binding cassette protein AtMRP5/AtABCC5 is a high affinity inositol hexakisphosphate transporterinvolved in guard cell signaling and phytate storage. J. Biol. Chem. 2009, 284, 33614–33622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freed, C.; Adepoju, O.; Gillaspy, G. Can inositol pyrophosphates inform strategies for developing low phytate crops? Plants 2020, 9, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krinke, O.; Novotná, Z.; Valentová, O.; Martinec, J. Inositol trisphosphate receptor in higher plants: Is it real? J. Exp. Bot. 2007, 58, 361–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michell, R.H. Inositol derivatives: Evolution and functions. Nat. Rev. Mol. Cell Biol. 2008, 9, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.E.; Rammesmayer, G.; Bohnert, H.J. Regulation of cell-specific inositol metabolism and transport in plant salinity tolerance. Plant Cell 1998, 10, 753–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishitani, M.; Majumder, A.L.; Bornhouser, A.; Michalowski, C.B.; Jensen, R.G.; Bohnert, H.J. Coordinate transcriptional induction of myo-inositol metabolism during environmental stress. Plant J. 1996, 9, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Carlomagno, G.; Unfer, V.; Roseff, S. The D-chiro-inositol paradox in the ovary. Fertil. Steril. 2011, 95, 2515–2516. [Google Scholar] [CrossRef]
- Galazis, N.; Galazi, M.; Atiomo, W. D-Chiro-inositol and its significance in polycystic ovary syndrome: A systematic review. Gynecol. Endocrinol. 2011, 27, 256–262. [Google Scholar] [CrossRef]
- Davis, A.; Christiansen, M.; Horowitz, J.F.; Klein, S.; Hellerstein, M.K.; Ostlund, R.E. Effect of pinitol treatment on insulin action in subjects with insulin resistance. Diabetes Care 2000, 23, 1000–1005. [Google Scholar] [CrossRef] [Green Version]
- Larner, J. D-chiro-inositol-its functional role in insulin action and its deficit in insulin resistance. Int. J. Exp. Diabetes Res. 2002, 3, 47–60. [Google Scholar] [CrossRef]
- Kang, M.-J.; Kim, J.-I.; Yoon, S.-Y.; Kim, J.C.; Cha, I.-J. Pinitol from soybeans reduces postprandial blood glucose in patients with type 2 diabetes mellitus. J. Med. Food 2006, 9, 182–186. [Google Scholar] [CrossRef]
- Nissen, P.M.; Nebel, C.; Oksbjerg, N.; Bertram, H.C. Metabolomics reveals relationship between plasma inositols and birth weight: Possible markers for fetal programming of type 2 diabetes. J. Biomed. Biotechnol. 2011, 2011. [Google Scholar] [CrossRef]
- Pintaudi, B.; Di Vieste, G.; Bonomo, M. The Effectiveness of Myo-Inositol and D-Chiro Inositol Treatment in Type 2 Diabetes. Int. J. Endocrinol. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Stevenson-Paulik, J.; Odom, A.R.; York, J.D. Molecular and biochemical characterization of two plant inositol polyphosphate 6-/3-/5-kinases. J. Biol. Chem. 2002, 277, 42711–42718. [Google Scholar] [CrossRef] [Green Version]
- Stevenson-Paulik, J.; Bastidas, R.J.; Chiou, S.T.; Frye, R.A.; York, J.D. Generation of phytate-free seeds in Arabidopsis through disruption of inositol polyphosphate kinases. Proc. Natl. Acad. Sci. USA 2005, 102, 12612–12617. [Google Scholar] [CrossRef] [Green Version]
- Berridge, M.J.; Irvine, R.F. Inositol phosphates and cell signalling. Nature 1989, 341, 197–205. [Google Scholar] [CrossRef]
- Saiardi, A.; Erdjument-Bromage, H.; Snowman, A.M.; Tempst, P.; Snyder, S.H. Synthesis of diphosphoinositol pentakisphosphate by a newly identified family of higher inositol polyphosphate kinases. Curr. Biol. 1999, 9, 1323–1326. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.I.; Tai, T.H. Identification of genes necessary for wild-type levels of seed phytic acid in Arabidopsis thaliana using a reverse genetics approach. Mol. Genet. Genom. 2011, 286, 119–133. [Google Scholar] [CrossRef]
- Shi, J.; Wang, H.; Hazebroek, J.; Ertl, D.S.; Harp, T. The maize low-phytic acid 3 encodes a myo-inositol kinase that plays a role in phytic acid biosynthesis in developing seeds. Plant J. 2005, 42, 708–719. [Google Scholar] [CrossRef]
- Funkhouser, E.A.; Loewus, F.A. Purification of myo-Inositol 1-Phosphate Synthase from Rice Cell Culture by Affinity Chromatography. Plant Physiol. 1975, 56, 786–790. [Google Scholar] [CrossRef] [Green Version]
- Desfougères, Y.; Wilson, M.S.C.; Laha, D.; Miller, G.J.; Saiardi, A. ITPK1 mediates the lipid-independent synthesis of inositol phosphates controlled by metabolism. Proc. Natl. Acad. Sci. USA 2019, 116, 24551–24561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, M.P.; Majerus, P.W. Characterization of a cDNA encoding Arabidopsis thaliana inositol 1,3,4-trisphosphate 5/6-kinase. Biochem. Biophys. Res. Commun. 1997, 232, 678–681. [Google Scholar] [CrossRef]
- Yang, X.; Shears, S.B. Multitasking in signal transduction by a promiscuous human Ins(3,4,5,6)P(4) 1-kinase/Ins(1,3,4)P(3) 5/6-kinase. Biochem. J. 2000, 351, 551–555. [Google Scholar] [CrossRef]
- Sweetman, D.; Johnson, S.; Caddick, S.E.K.; Hanke, D.E.; Brearley, C.A. Characterization of an Arabidopsis inositol 1,3,4,5,6-pentakisphosphate 2-kinase(AtIPK1). Biochem. J. 2006, 394, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Verbsky, J.W.; Wilson, M.P.; Kisseleva, M.V.; Majerus, P.W.; Wente, S.R. The synthesis of inositol hexakisphosphate: Characterization of human inositol 1,3,4,5,6-pentakisphosphate 2-kinase. J. Biol. Chem. 2002, 277, 31857–31862. [Google Scholar] [CrossRef] [Green Version]
- Kuo, H.F.; Hsu, Y.Y.; Lin, W.C.; Chen, K.Y.; Munnik, T.; Brearley, C.A.; Chiou, T.J. Arabidopsis inositol phosphate kinases IPK1 and ITPK1 constitute a metabolic pathway in maintaining phosphate homeostasis. Plant J. 2018, 95, 613–630. [Google Scholar] [CrossRef] [Green Version]
- Draskovic, P.; Saiardi, A.; Bhandari, R.; Burton, A.; Ilc, G.; Kovacevic, M.; Snyder, S.H.; Podobnik, M. Inositol hexakisphosphate kinase products contain diphosphate and triphosphate groups. Chem. Biol. 2008, 15, 274–286. [Google Scholar] [CrossRef] [Green Version]
- Desai, M.; Rangarajan, P.; Donahue, J.L.; Williams, S.P.; Land, E.S.; Mandal, M.K.; Phillippy, B.Q.; Perera, I.Y.; Raboy, V.; Gillaspy, G.E. Two inositol hexakisphosphate kinases drive inositol pyrophosphate synthesis in plants. Plant J. 2014, 80, 642–653. [Google Scholar] [CrossRef]
- Sweetman, D.; Stavridou, I.; Johnson, S.; Green, P.; Caddick, S.E.K.; Brearley, C.A. Arabidopsis thaliana inositol 1,3,4-trisphosphate 5/6-kinase 4 (AtITPK4) is an outlier to a family of ATP-grasp fold proteins from Arabidopsis. FEBS Lett. 2007, 581, 4165–4171. [Google Scholar] [CrossRef] [Green Version]
- Qian, X.; Mitchell, J.; Wei, S.J.; Williams, J.; Petrovich, R.M.; Shears, S.B. The Ins(1,3,4)P3 5/6-kinase/Ins(3,4,5,6)P4 1-kinase is not a protein kinase. Biochem. J. 2005, 389, 389–395. [Google Scholar] [CrossRef] [Green Version]
- Adepoju, O.; Williams, S.P.; Craige, B.; Cridland, C.A.; Sharpe, A.K.; Brown, A.M.; Land, E.; Perera, I.Y.; Mena, D.; Sobrado, P.; et al. Inositol Trisphosphate Kinase and Diphosphoinositol Pentakisphosphate Kinase Enzymes Constitute the Inositol Pyrophosphate Synthesis Pathway in Plants. bioRxiv 2019, 724914. [Google Scholar] [CrossRef] [Green Version]
- Laha, D.; Parvin, N.; Hofer, A.; Giehl, R.F.H.; Fernandez-Rebollo, N.; Von Wirén, N.; Saiardi, A.; Jessen, H.J.; Schaaf, G. Arabidopsis ITPK1 and ITPK2 Have an Evolutionarily Conserved Phytic Acid Kinase Activity. ACS Chem. Biol. 2019, 14, 2127–2133. [Google Scholar] [CrossRef] [Green Version]
- Schell, M.J. Inositol trisphosphate 3-kinases: Focus on immune and neuronal signaling. Cell. Mol. Life Sci. 2010, 67, 1755–1778. [Google Scholar] [CrossRef]
- Wang, H.; Derose, E.F.; London, R.E.; Shears, S.B. IP6K structure and the molecular determinants of catalytic specificity in an inositol phosphate kinase family. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Falck, J.R.; Hall, T.M.T.; Shears, S.B. Structural basis for an inositol pyrophosphate kinase surmounting phosphate crowding. Nat. Chem. Biol. 2012, 8, 111–116. [Google Scholar] [CrossRef] [Green Version]
- Mulugu, S.; Bai, W.; Fridy, P.C.; Bastidas, R.J.; Otto, J.C.; Dollins, D.E.; Haystead, T.A.; Ribeiro, A.A.; York, J.D. A conserved family of enzymes that phosphorylate inositol hexakisphosphate. Science 2007, 316, 106–109. [Google Scholar] [CrossRef]
- Choi, J.H.; Williams, J.; Cho, J.; Falck, J.R.; Shears, S.B. Purification, sequencing, and molecular identification of a mammalian PP-InsP5 kinase that is activated when cells are exposed to hyperosmotic stress. J. Biol. Chem. 2007, 282, 30763–30775. [Google Scholar] [CrossRef] [Green Version]
- Gokhale, N.A.; Zaremba, A.; Shears, S.B. Receptor-dependent compartmentalization of PPIP5K1, a kinase with a cryptic polyphosphoinositide binding domain. Biochem. J. 2011, 434, 415–426. [Google Scholar] [CrossRef] [Green Version]
- Scheffzek, K.; Welti, S. Pleckstrin homology (PH) like domains-versatile modules in protein-protein interaction platforms. FEBS Lett. 2012, 586, 2662–2673. [Google Scholar] [CrossRef] [Green Version]
- Nair, V.S.; Gu, C.; Janoshazi, A.K.; Jessen, H.J.; Wang, H.; Shears, S.B. Inositol pyrophosphate synthesis by diphosphoinositol pentakisphosphate kinase-1 is regulated by phosphatidylinositol(4,5)bisphosphate. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef] [Green Version]
- Luo, H.R.; Huang, Y.E.; Chen, J.C.; Saiardi, A.; Iijima, M.; Ye, K.; Huang, Y.; Nagata, E.; Devreotes, P.; Snyder, S.H. Inositol pyrophosphates mediate chemotaxis in Dictyostelium via pleckstrin homology domain-Ptdins(3,4,5)P3 interactions. Cell 2003, 114, 559–572. [Google Scholar] [CrossRef] [Green Version]
- Shears, S.B. A short historical perspective of methods in inositol phosphate research. In Methods in Molecular Biology; Miller, G., Ed.; Humana Press Inc.: Totowa, NJ, USA, 2020; Volume 2091, pp. 1–28. [Google Scholar]
- Randall, T.A.; Gu, C.; Li, X.; Wang, H.; Shears, S.B. A two-way switch for inositol pyrophosphate signaling: Evolutionary history and biological significance of a unique, bifunctional kinase/phosphatase. Adv. Biol. Regul. 2019, 75. [Google Scholar] [CrossRef] [PubMed]
- Machkalyan, G.; Trieu, P.; Pétrin, D.; Hébert, T.E.; Miller, G.J. PPIP5K1 interacts with the exocyst complex through a C-terminal intrinsically disordered domain and regulates cell motility. Cell. Signal. 2016, 28, 401–411. [Google Scholar] [CrossRef]
- Pisani, F.; Livermore, T.; Rose, G.; Chubb, J.R.; Gaspari, M.; Saiardi, A. Analysis of Dictyostelium discoideum inositol pyrophosphate metabolism by gel electrophoresis. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [Green Version]
- Dollins, D.E.; Bai, W.; Fridy, P.C.; Otto, J.C.; Neubauer, J.L.; Gattis, S.G.; Mehta, K.P.M.; York, J.D. Vip1 is a kinase and pyrophosphatase switch that regulates inositol diphosphate signaling. Proc. Natl. Acad. Sci. USA 2020, 117, 9356–9364. [Google Scholar] [CrossRef] [Green Version]
- Gu, C.; Nguyen, H.N.; Ganini, D.; Chen, Z.; Jessen, H.J.; Gu, Z.; Wang, H.; Shears, S.B. KO of 5-InsP7 kinase activity transforms the HCT116 colon cancer cell line into a hypermetabolic, growth-inhibited phenotype. Proc. Natl. Acad. Sci. USA 2017, 114, 11968–11973. [Google Scholar] [CrossRef] [Green Version]
- Pascual-Ortiz, M.; Saiardi, A.; Walla, E.; Jakopec, V.; Künzel, N.A.; Span, I.; Vangala, A.; Fleig, U. Asp1 Bifunctional Activity Modulates Spindle Function via Controlling Cellular Inositol Pyrophosphate Levels in Schizosaccharomyces pombe. Mol. Cell. Biol. 2018, 38. [Google Scholar] [CrossRef] [Green Version]
- Pöhlmann, J.; Risse, C.; Seidel, C.; Pohlmann, T.; Jakopec, V.; Walla, E.; Ramrath, P.; Takeshita, N.; Baumann, S.; Feldbrügge, M.; et al. The Vip1 Inositol Polyphosphate Kinase Family Regulates Polarized Growth and Modulates the Microtubule Cytoskeleton in Fungi. PLoS Genet. 2014, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azevedo, C.; Saiardi, A. Eukaryotic Phosphate Homeostasis: The Inositol Pyrophosphate Perspective. Trends Biochem. Sci. 2017, 42, 219–231. [Google Scholar] [CrossRef]
- Jung, J.-Y.; Ried, M.K.; Hothorn, M.; Poirier, Y. Control of plant phosphate homeostasis by inositol pyrophosphates and the SPX domain. Curr. Opin. Biotechnol. 2018, 49, 156–162. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Lau, K.; Puschmann, R.; Harmel, R.K.; Zhang, Y.; Pries, V.; Gaugler, P.; Broger, L.; Dutta, A.K.; Jessen, H.J.; et al. Two bifunctional inositol pyrophosphate kinases/phosphatases control plant phosphate homeostasis. eLife 2019, 8, 1–25. [Google Scholar] [CrossRef]
- Dong, J.; Ma, G.; Sui, L.; Wei, M.; Satheesh, V.; Zhang, R.; Ge, S.; Li, J.; Zhang, T.-E.; Wittwer, C.; et al. Inositol Pyrophosphate InsP8 Acts as an Intracellular Phosphate Signal in Arabidopsis. Mol. Plant 2019, 12, 1463–1473. [Google Scholar] [CrossRef] [PubMed]
- Kuo, H.F.; Chang, T.Y.; Chiang, S.F.; Wang, W.D.; Charng, Y.Y.; Chiou, T.J. Arabidopsis inositol pentakisphosphate 2-kinase, AtIPK1, is required for growth and modulates phosphate homeostasis at the transcriptional level. Plant J. 2014, 80, 503–515. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.S.; Henning, X.; Jessen, J.; Saiardi, A. The inositol hexakisphosphate kinases IP6K1 and-2 regulate human cellular phosphate homeostasis, including XPR1-mediated phosphate export Downloaded from. J. Biol. Chem 2019, 294, 11597–11608. [Google Scholar] [CrossRef] [Green Version]
- Wild, R.; Gerasimaite, R.; Jung, J.-Y.; Truffault, V.; Pavlovic, I.; Schmidt, A.; Saiardi, A.; Jessen, H.J.; Poirier, Y.; Hothorn, M.; et al. Control of eukaryotic phosphate homeostasis by inositol polyphosphate sensor domains. Science 2016, 352, 986–990. [Google Scholar] [CrossRef] [Green Version]
- Gerasimaite, R.; Pavlovic, I.; Capolicchio, S.; Hofer, A.; Schmidt, A.; Jessen, H.J.; Mayer, A. Inositol Pyrophosphate Specificity of the SPX-Dependent Polyphosphate Polymerase VTC. ACS Chem. Biol. 2017, 12, 648–653. [Google Scholar] [CrossRef] [PubMed]
- Secco, D.; Wang, C.; Arpat, B.A.; Wang, Z.; Poirier, Y.; Tyerman, S.D.; Wu, P.; Shou, H.; Whelan, J. The emerging importance of the SPX domain-containing proteins in phosphate homeostasis. New Phytol. 2012, 193, 842–851. [Google Scholar] [CrossRef] [Green Version]
- Secco, D.; Wang, C.; Shou, H.; Whelan, J. Phosphate homeostasis in the yeast Saccharomyces cerevisiae, the key role of the SPX domain-containing proteins. FEBS Lett. 2012, 586, 289–295. [Google Scholar] [CrossRef] [Green Version]
- Hürlimann, H.C.; Pinson, B.; Stadler-Waibel, M.; Zeeman, S.C.; Freimoser, F.M. The SPX domain of the yeast low-affinity phosphate transporter Pho90 regulates transport activity. EMBO Rep. 2009, 10, 1003–1008. [Google Scholar] [CrossRef]
- Zhou, Z.; Wang, Z.; Lv, Q.; Shi, J.; Zhong, Y.; Wu, P.; Mao, C. SPX proteins regulate Pi homeostasis and signaling in different subcellular level. Plant Signal. Behav. 2015, 10. [Google Scholar] [CrossRef]
- Puga, M.I.; Mateos, I.; Charukesi, R.; Wang, Z.; Franco-Zorrilla, J.M.; de Lorenzo, L.; Irigoyen, M.L.; Masiero, S.; Bustos, R.; Rodrigues, J.; et al. SPX1 is a phosphate-dependent inhibitor of PHOSPHATE STARVATION RESPONSE 1 in Arabidopsis. PNAS 2014, 111, 14947–14952. [Google Scholar] [CrossRef] [Green Version]
- Ried, M.K.; Wild, R.; Zhu, J.; Broger, L.; Harmel, R.K.; Hothorn, L.A.; Fiedler, D.; Hothorn, M. Inositol pyrophosphates promote the interaction of SPX domains with the coiled-coil motif of PHR transcription factors to regulate plant phosphate homeostasis. BioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Gu, C.; Hostachy, S.; Sahu, S.; Wittwer, C.; Jessen, H.J.; Fiedler, D.; Wang, H.; Shears, S.B. Control of XPR1-dependent cellular phosphate efflux by InsP 8 is an exemplar for functionally-exclusive inositol pyrophosphate signaling. Proc. Natl. Acad. Sci. USA 2020, 117. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic model of InsP and PP-InsP metabolism in humans and plants. InsPs are synthesized through the Lipid-Dependent (yellow) or Lipid-Independent (blue) pathways. PP-InsPs are synthesized from InsP6 via the PP-InsP biosynthesis pathway (pink). Key enzymes in the InsP and PP-InsP pathways conserved in animals and plants (grey), specific to animals (purple) or specific to plants (green) are shown next to their respective reactions. Enzyme names are defined in the text.
Figure 1.
Schematic model of InsP and PP-InsP metabolism in humans and plants. InsPs are synthesized through the Lipid-Dependent (yellow) or Lipid-Independent (blue) pathways. PP-InsPs are synthesized from InsP6 via the PP-InsP biosynthesis pathway (pink). Key enzymes in the InsP and PP-InsP pathways conserved in animals and plants (grey), specific to animals (purple) or specific to plants (green) are shown next to their respective reactions. Enzyme names are defined in the text.

Figure 2.
Model of PP-InsP regulation of the plant Pi starvation response. Under low Pi conditions (left), transcription factor PHR1 binds to P1BS-containing promoters. Under sufficient Pi conditions, SPX1 interacts with PHR1 via PP-InsPs, preventing PHR1 from binding to P1BS-containing promoters. Adapted from [71].
Figure 2.
Model of PP-InsP regulation of the plant Pi starvation response. Under low Pi conditions (left), transcription factor PHR1 binds to P1BS-containing promoters. Under sufficient Pi conditions, SPX1 interacts with PHR1 via PP-InsPs, preventing PHR1 from binding to P1BS-containing promoters. Adapted from [71].

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cridland, C.; Gillaspy, G. Inositol Pyrophosphate Pathways and Mechanisms: What Can We Learn from Plants? Molecules 2020, 25, 2789. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25122789
AMA Style
Cridland C, Gillaspy G. Inositol Pyrophosphate Pathways and Mechanisms: What Can We Learn from Plants? Molecules. 2020; 25(12):2789. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25122789
Chicago/Turabian StyleCridland, Caitlin, and Glenda Gillaspy. 2020. "Inositol Pyrophosphate Pathways and Mechanisms: What Can We Learn from Plants?" Molecules 25, no. 12: 2789. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25122789