
Melanogenic Inhibition and Toxicity Assessment of Flavokawain A and B on B16/F10 Melanoma Cells and Zebrafish (Danio rerio)

 , ,
, ,
Abstract
:1. Introduction
2. Results
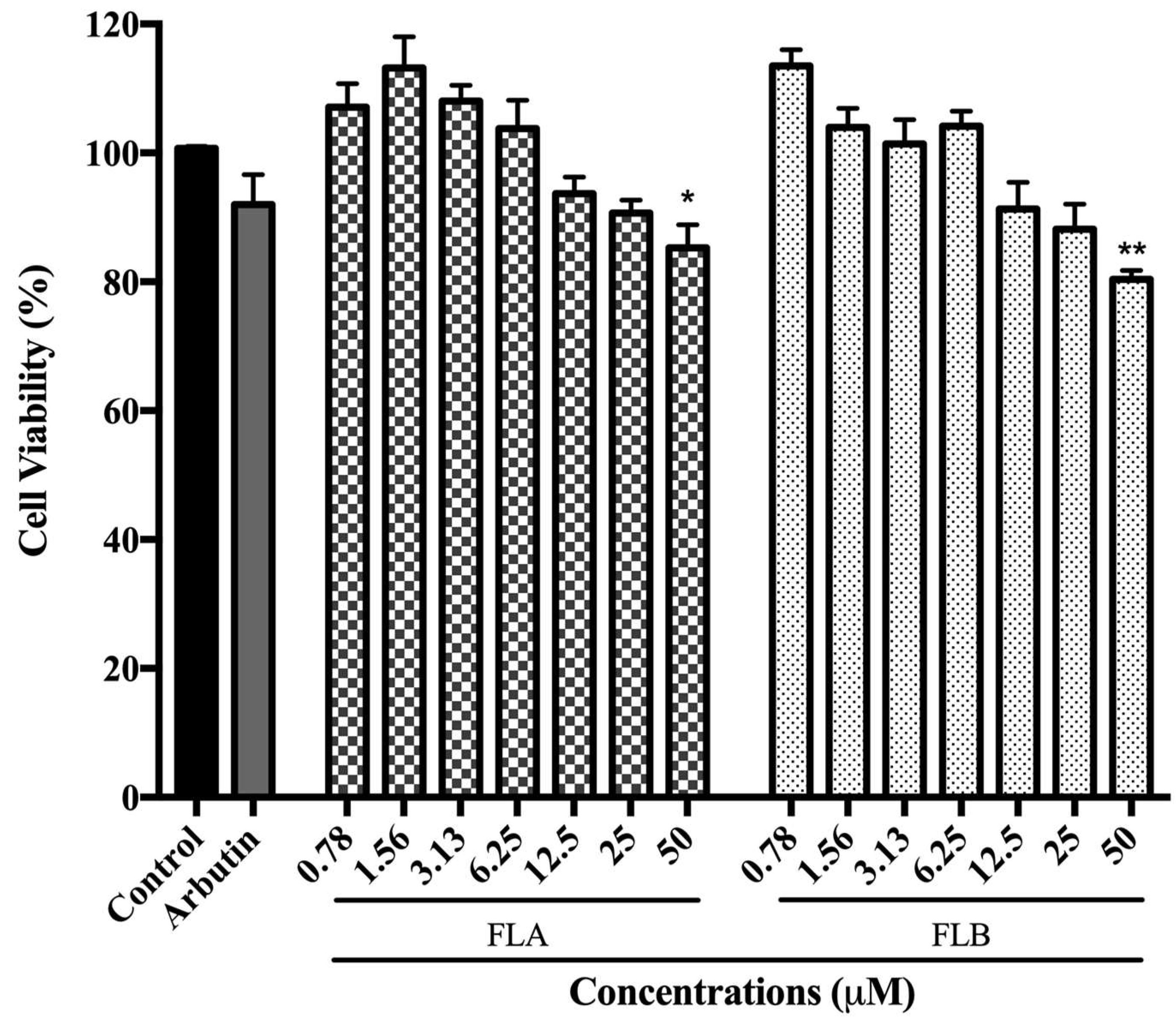
2.1. Cytotoxic Effect of Chalcone Derivatives on B16/F10 Cells
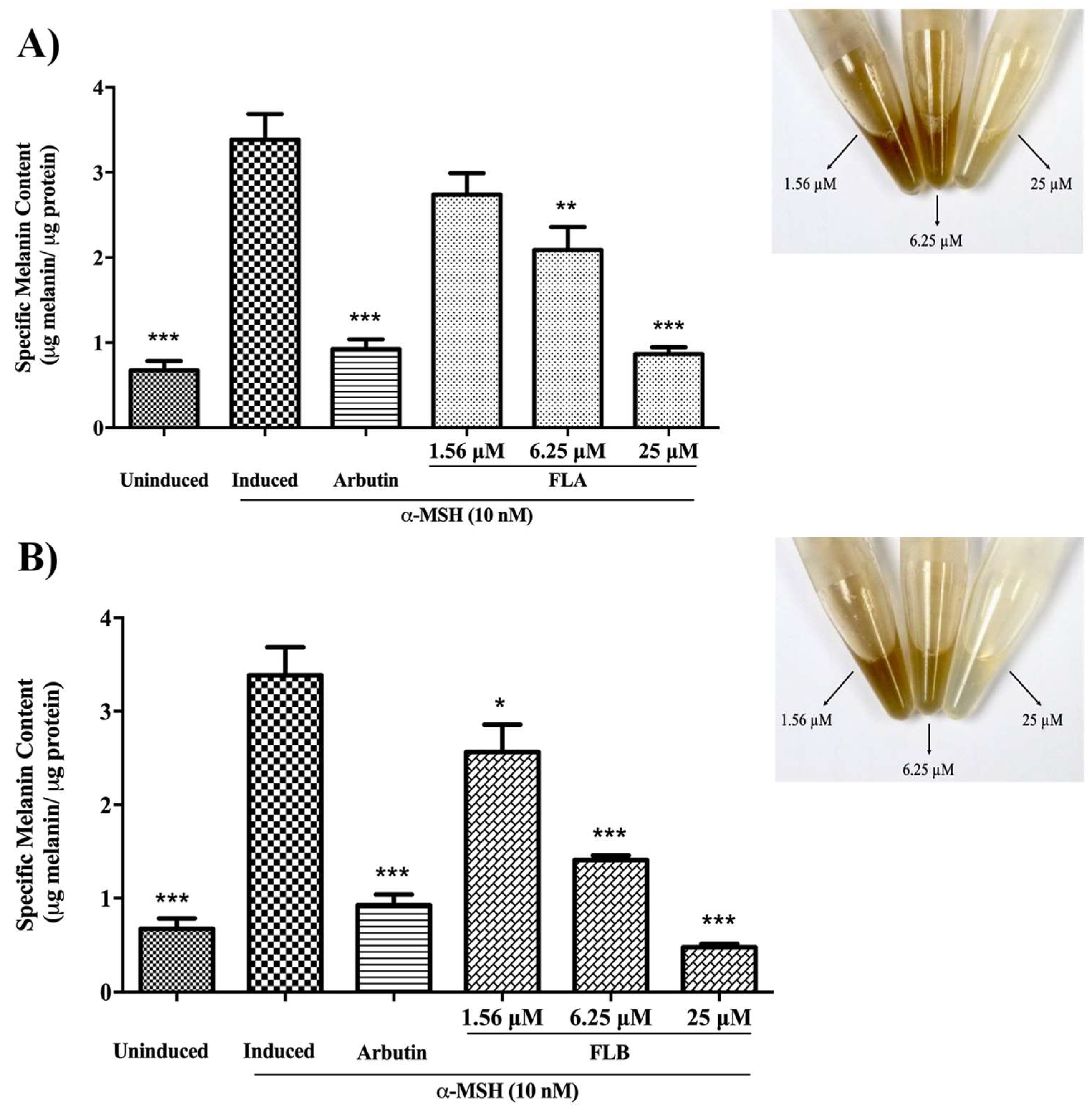
2.2. Inhibitory Effect of Chalcone Derivatives on Cellular Melanin Content
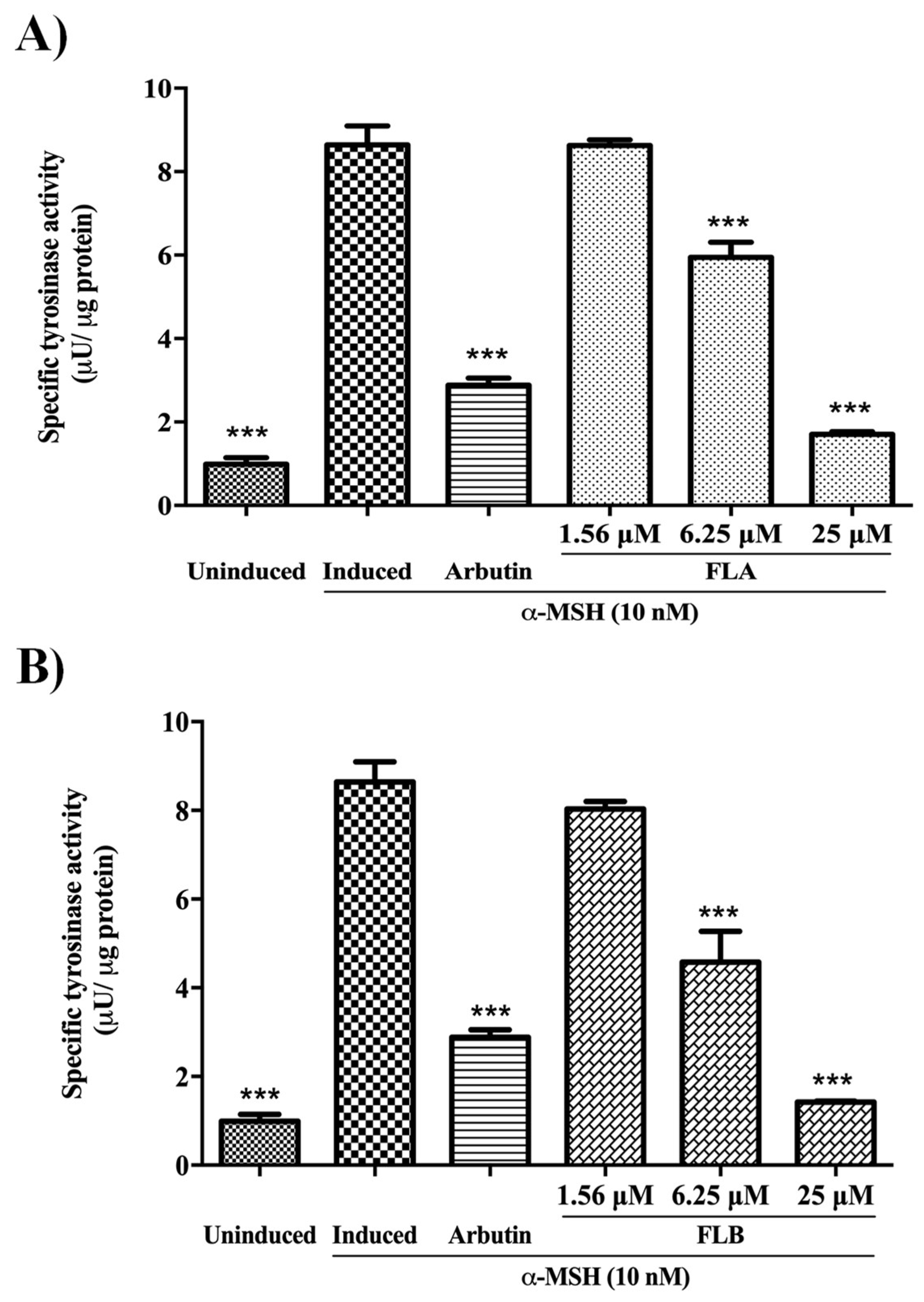
2.3. Inhibitory Effect of Chalcone Derivatives on Cellular Tyrosinase Activity
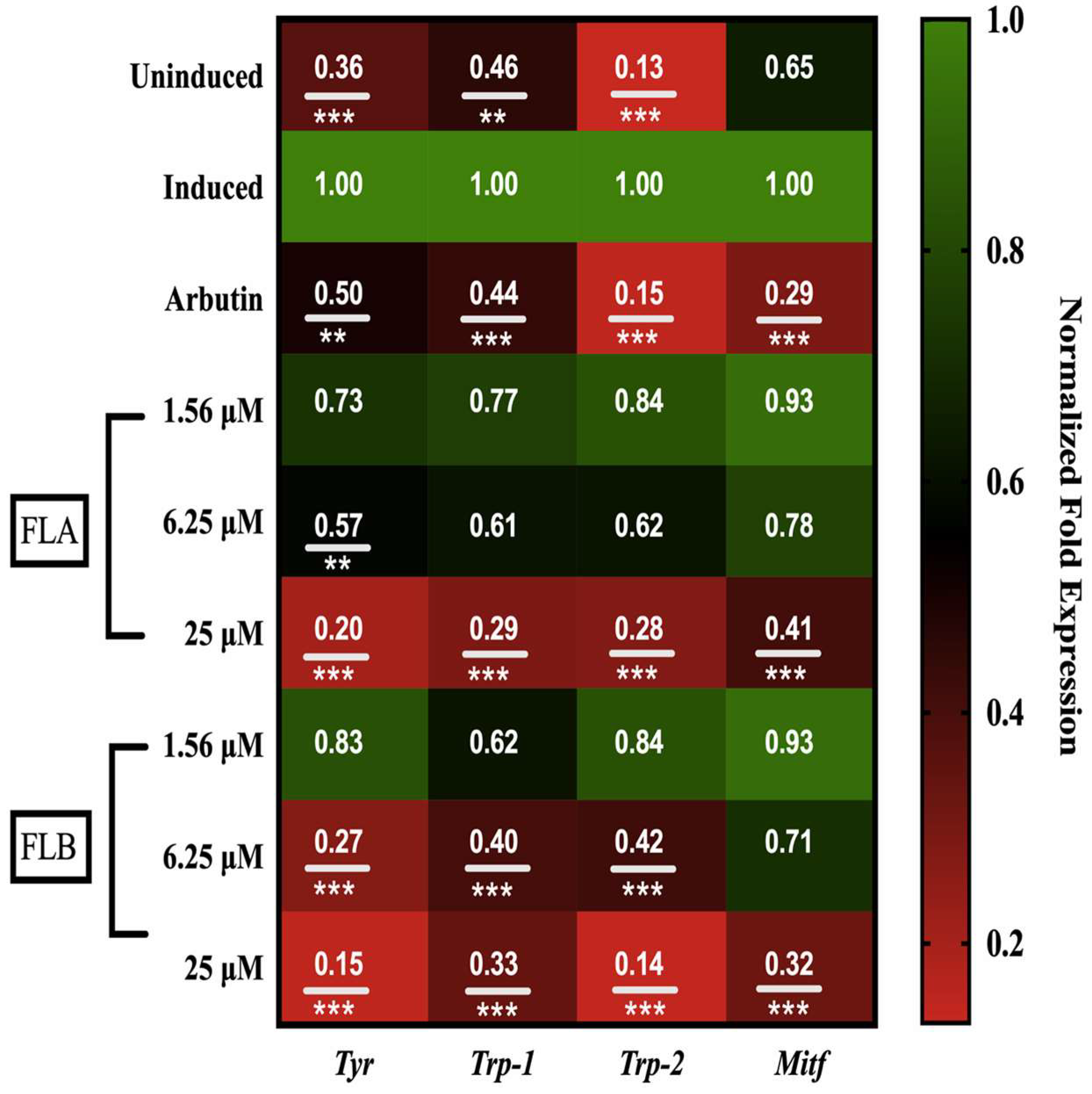
2.4. Down-Regulation Effect of Chalcone Derivatives on Tyr, Trp-1, Trp-2 and Mitf Genes Expression in B16/F10 Cells
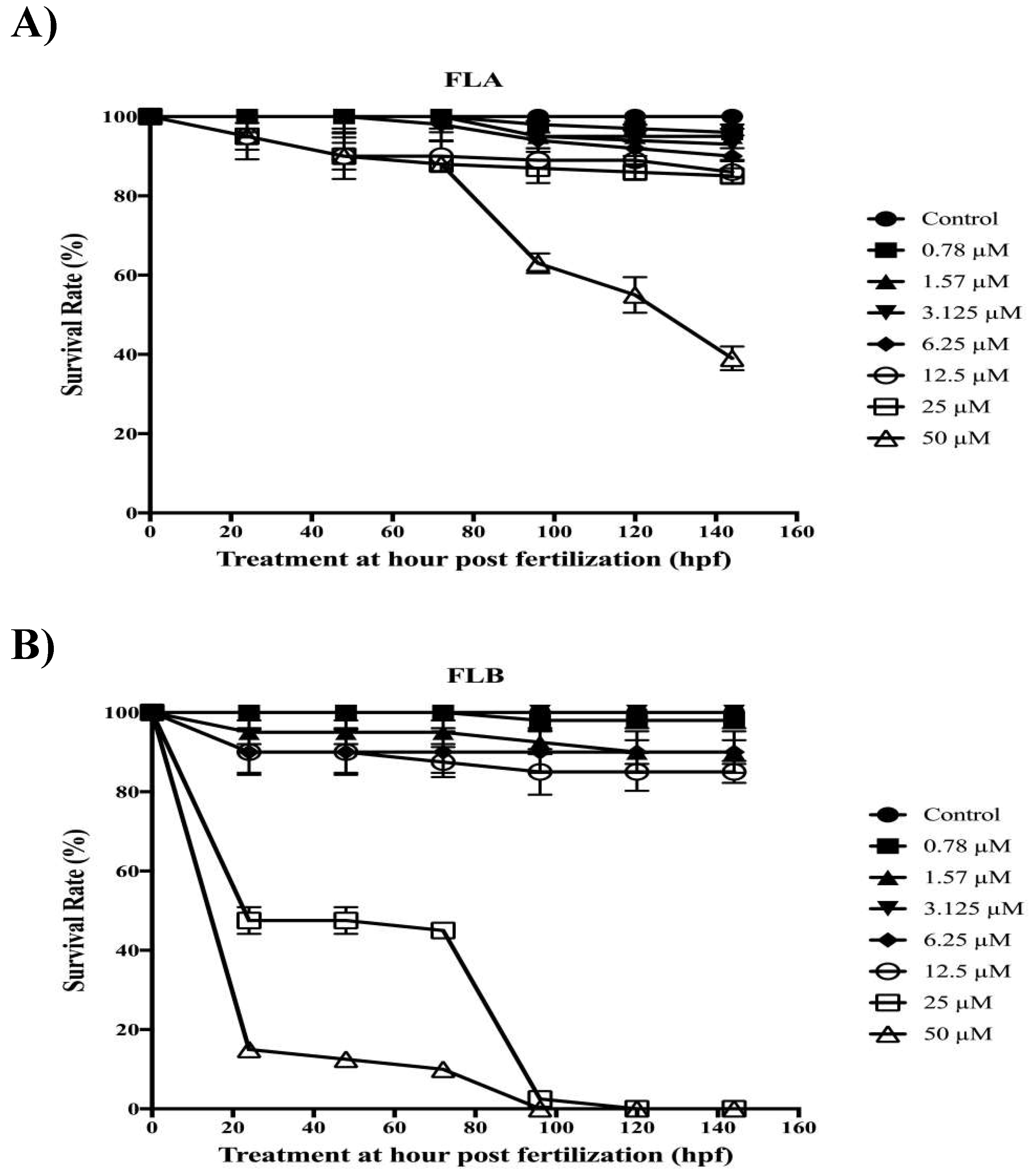
2.5. Zebrafish Toxicity Assessment
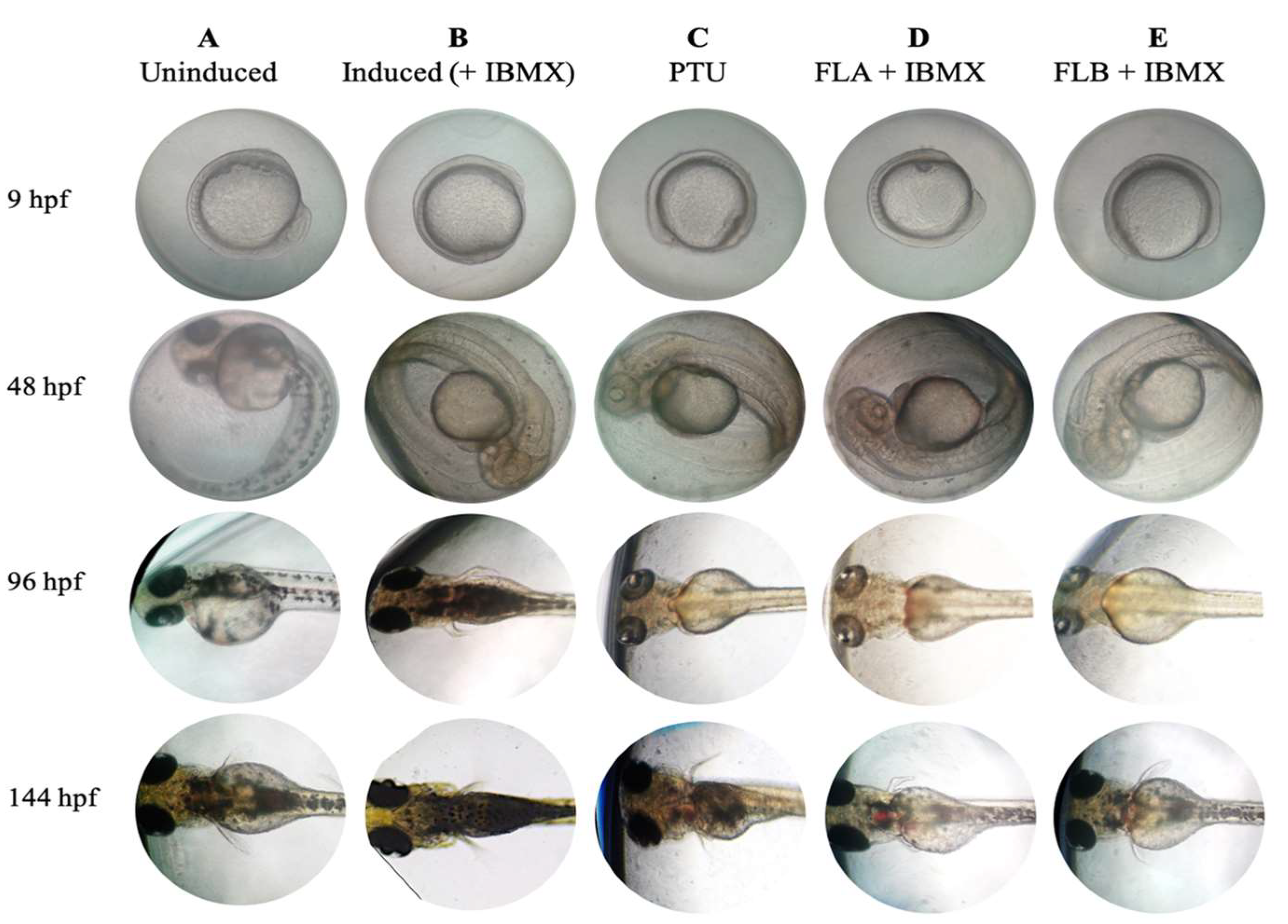
2.6. Zebrafish Phenotype-Based of Melanogenic Inhibition by FLA and FLB
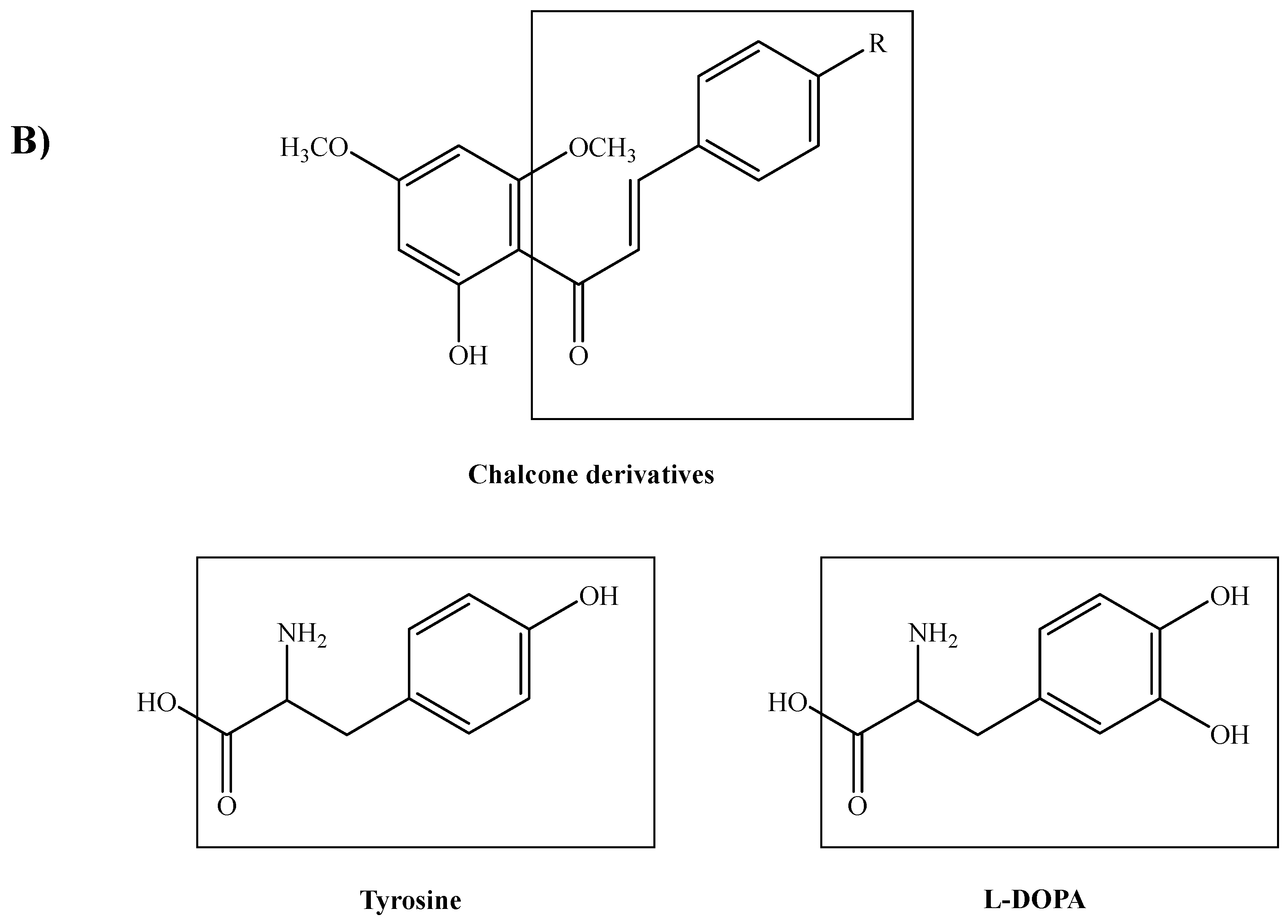
3. Discussion
4. Materials and Methods
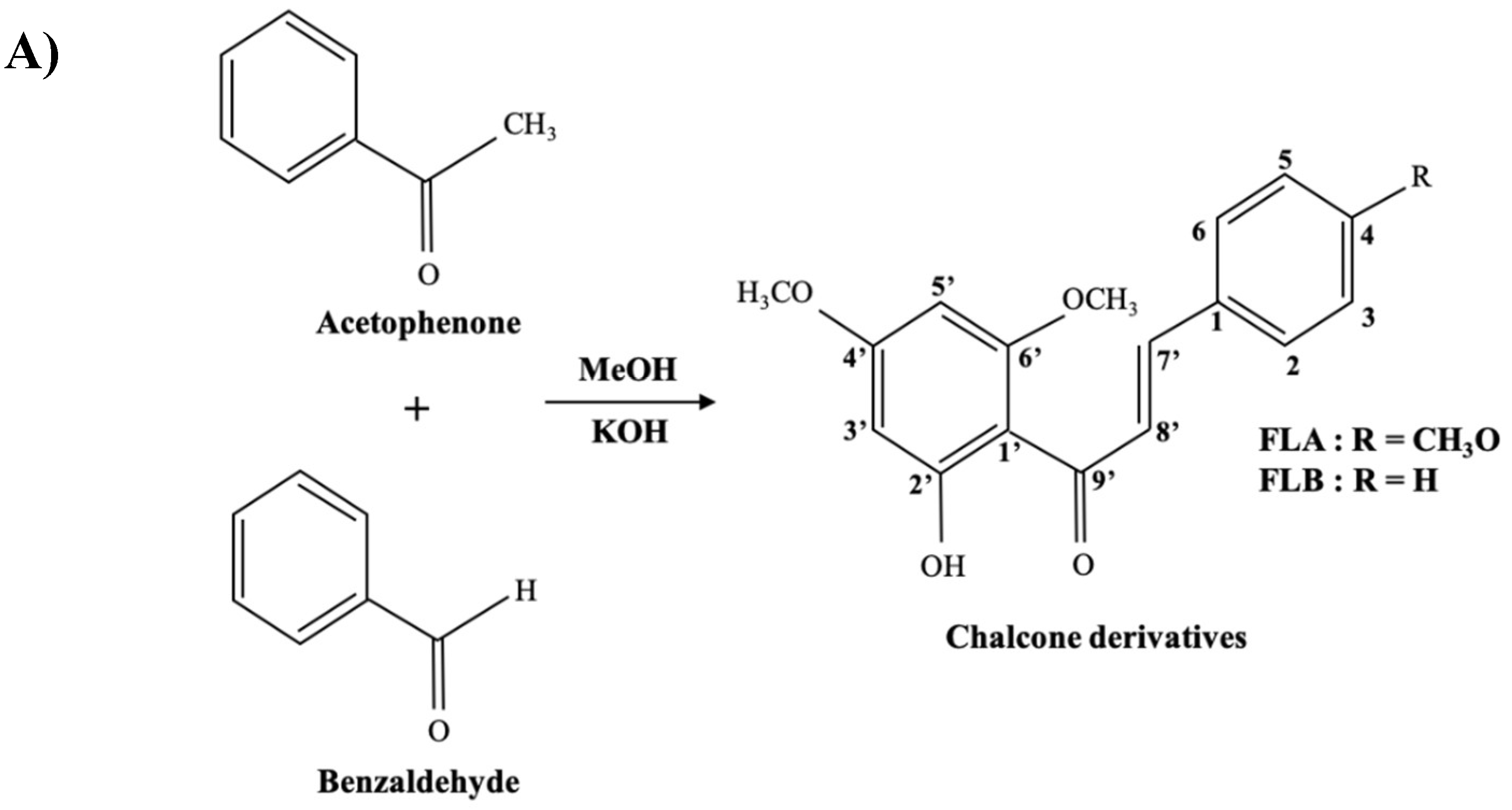
4.1. General Procedure for Preparation of Analogues
4.1.1. (E)-1-(2′-Hydroxy-4′,6′-dimethoxyphenyl)-3-(4-methoxyphenylprop-2-en-1-one: (FLA)
4.1.2. (E)-1-(2′-Hydroxy-4′,6′-dimethoxyphenyl)-3-phenylprop-2-en-1-one:(FLB)
4.2. Cell Culture and Treatment
4.3. Cell Viability Test
4.4. Determination of Cellular Melanin Content
4.5. Determination of Cellular Tyrosinase
4.6. Quantitative Real-Time PCR (RT-qPCR) of Tyr, Trp-1, Trp-2 and Mitf
4.7. Zebrafish Husbandry and Embryos Collection
4.8. Zebrafish Toxicity Assay of FLA and FLB
4.9. Phenotype-Based Depigmenting Test of FLA and FLB on Zebrafish
4.10. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Ethical Standard
Abbreviations
| ACTH | adrenocorticotropic hormone |
| α-MSH | alpha-melanocyte stimulating hormone |
| bHLH-LZ | basic-helix-loop-helix-leucine-zipper |
| BSA | Bovine serum albumin |
| DHI | Dihydroxyindole |
| DHICA | Dihydroxyindole carboxylic acid |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| DMSO | Dimethyl sulfoxide |
| EDTA | Ethylenediaminetetraacetic acid |
| FBS | Foetal bovine serum |
| FLA | (E)-1-(2′-Hydroxy-4′,6′-dimethoxyphenyl)-3-(4-methoxyphenylprop-2-en-1-one |
| FLB | (E)-1-(2′-Hydroxy-4′,6′-dimethoxyphenyl)-3-phenylprop-2-en-1-one |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| IBMX | 3-isobutyl-1-methylxanthine |
| l-DOPA | l-3,4-dihydroxyphenylalanine |
| Mitf | Microphthalmia associated transcription factor |
| MTT | 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyl tetrazolium bromide |
| NMR | Nuclear magnetic resonance |
| PTU | Phenylthiourea |
| qPCR | Quantitative real-time polymerase chain reaction |
| Trp-1 | Tyrosinase related protein-1 |
| Trp-2 | Tyrosinase related protein-2 |
| Tyr | Tyrosinase |
| UVR | Ultraviolet radiation |
References
- Sorg, H.; Tilkorn, D.J.; Hager, S.; Hauser, J.; Mirastschijski, U. Skin wound healing: An update on the current knowledge and concepts. Eur. Surg. Res. 2017, 58, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Passeron, T.; Nouveau, S.; Duval, C.; Cardot-Leccia, N.; Piffaut, V.; Bourreau, E.; Queille-Roussel, C.; Bernerd, F. Development and validation of a reproducible model for studying post-inflammatory hyperpigmentation. Pigm. Cell Melanoma Res. 2018, 31, 649–652. [Google Scholar] [CrossRef]
- Yang, Y.; Jang, G.B.; Yang, X.; Wang, Q.; He, S.; Li, S.; Quach, C.; Zhao, S.; Li, F.; Yuan, Z.; et al. Central role of autophagic UVRAG in melanogenesis and the suntan response. Proc. Natl. Acad. Sci. USA 2018, 115, E7728–E7737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shekhar, S.; Dharmshaktu, P. On the palms of his hands: ACTH-induced hyperpigmentation. Am. J. Med. 2018, 131, 144–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.Y.; Fisher, D.E. Melanocyte biology and skin pigmentation. Nature 2007, 445, 843. [Google Scholar] [CrossRef] [PubMed]
- Cone, R.D.; Lu, D.; Koppula, S.; Vage, D.I.; Klungland, H.; Boston, B.; Chen, W.; Orth, D.N.; Pouton, C.; Kesterson, R.A. The melanocortin receptors: Agonists, antagonists, and the hormonal control of pigmentation. Recent Prog. Horm. Res. 1996, 51, 287–317. [Google Scholar] [PubMed]
- D’Mello, S.; Finlay, G.; Baguley, B.; Askarian-Amiri, M. Signaling pathways in melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aksan, I.; Goding, C. Targeting the microphthalmia basic helix-loop-helix–leucine zipper transcription factor to a subset of E-box elements in vitro and in vivo. Mol. Cell. Biol. 1998, 18, 6930–6938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertolotto, C.; Abbe, P.; Hemesath, T.J.; Bille, K.; Fisher, D.E.; Ortonne, J.; Ballotti, R. Microphthalmia gene product as a signal transducer in cAMP-induced differentiation of melanocytes. J. Cell Biol. 1998, 142, 827–835. [Google Scholar] [CrossRef] [Green Version]
- Ito, S. A chemist’s view of melanogenesis. Pigm. Cell Res. 2003, 16, 230–236. [Google Scholar] [CrossRef] [Green Version]
- Tsukamoto, K.; Jackson, I.J.; Urabe, K.; Montague, P.M.; Hearing, V. A second tyrosinase-related protein, Trp-2, is a melanogenic enzyme termed DOPAchrome tautomerase. EMBO J. 1992, 11, 519. [Google Scholar] [CrossRef] [PubMed]
- Del Marmol, V.; Ito, S.; Jackson, I.; Vachtenheim, J.; Berr, P.; Ghanem, G.; Morandini, R.; Wakamatsu, K.; Huez, G. Trp-1 expression correlates with eumelanogenesis in human pigment cells in culture. FEBS Lett. 1993, 327, 307–310. [Google Scholar] [CrossRef]
- Kobayashi, T.; Urabe, K.; Winder, A.; Jimenez-Cervantes, C.; Imokawa, G.; Brewington, T.; Solano, F.; Garcia-Borron, J.C.; Hearing, V. Tyrosinase related protein 1 (Trp-1) functions as a DHICA oxidase in melanin biosynthesis. EMBO J. 1994, 13, 5818. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Akinseye, O.; Tovar-Garza, A.; Pandya, A.G. The effect of melasma on self-esteem: A pilot study. Int. J. Women’s Dermatol. 2018, 4, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Dabas, G.; Vinay, K.; Parsad, D.; Kumar, A.; Kumaran, M.S. Psychological disturbances in patients with pigmentary disorders: A cross-sectional study. J. Eur. Acad. Dermatol. Venereol. 2019, 34, 392–399. [Google Scholar] [CrossRef]
- Sanchez, J.L.; Vazquez, M. A hydroquinone solution in the treatment of melasma. Int. J. Dermatol. 1982, 21, 55–59. [Google Scholar] [CrossRef]
- Palumbo, A.; d’Ischia, M.; Misuraca, G.; Prota, G. Mechanism of inhibition of melanogenesis by hydroquinone. BBA-Gen. Subjects 1991, 1073, 85–90. [Google Scholar] [CrossRef]
- Nordlund, J.; Grimes, P.; Ortonne, J. The safety of hydroquinone. J. Eur. Acad. Dermatol. Venereol. 2006, 20, 781–787. [Google Scholar] [CrossRef]
- Draelos, Z.D.; Yatskayer, M.; Bhushan, P.; Pillai, S.; Oresajo, C. Evaluation of a kojic acid, emblica extract, and glycolic acid formulation compared with hydroquinone 4% for skin lightening. Cutis 2010, 86, 153–158. [Google Scholar]
- Maeda, K.; Fukuda, M. In vitro effectiveness of several whitening cosmetic components in human melanocytes. J. Soc. Cosmet. Chem. 1991, 42, 261–268. [Google Scholar]
- Cabanes, J.; Chazarra, S.; Garcia-Carmona, F. Kojic acid, a cosmetic skin whitening agent, is a slow-binding inhibitor of catecholase activity of tyrosinase. J. Pharm. Pharmacol. 1994, 46, 982–985. [Google Scholar] [CrossRef] [PubMed]
- Battaini, G.; Monzani, E.; Casella, L.; Santagostini, L.; Pagliarin, R. Inhibition of the catecholase activity of biomimetic dinuclear copper complexes by kojic acid. J. Biol. Inorg. Chem. 2000, 5, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, M.; Kawai, K.; Kawai, K. Contact allergy to kojic acid in skin care products. Contact Dermat. 1995, 32, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Chusiri, Y.; Wongpoomchai, R.; Kakehashi, A.; Wei, M.; Wanibuchi, H.; Vinitketkumnuan, U.; Fukushima, S. Non-genotoxic mode of action and possible threshold for hepatocarcinogenicity of kojic acid in F344 rats. Food Chem. Toxicol. 2011, 49, 471–476. [Google Scholar] [CrossRef]
- Nohynek, G.J.; Kirkland, D.; Marzin, D.; Toutain, H.; Leclerc-Ribaud, C.; Jinnai, H. An assessment of the genotoxicity and human health risk of topical use of kojic acid [5-hydroxy-2-(hydroxymethyl)-4H-pyran-4-one]. Food Chem. Toxicol. 2004, 42, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Cassar, S.; Adatto, I.; Freeman, J.L.; Gamse, J.T.; Iturria, I.; Lawrence, C.; Muriana, A.; Peterson, R.T.; Van Cruchten, S.; Zon, L.I. Use of zebrafish in drug discovery toxicology. Chem. Res. Toxicol. 2020, 33, 95–118. [Google Scholar] [CrossRef] [Green Version]
- He, J.H.; Guo, S.Y.; Zhu, F.; Zhu, J.J.; Chen, Y.X.; Huang, C.J.; Gao, J.M.; Dong, Q.X.; Xuan, Y.X.; Li, C.Q. A zebrafish phenotypic assay for assessing drug-induced hepatotoxicity. J. Pharmacol. Toxicol. Methods 2013, 67, 25–32. [Google Scholar] [CrossRef]
- Ramlan, N.F.; Sata, N.S.A.M.; Hassan, S.N.; Bakar, N.A.; Ahmad, S.; Zulkifli, S.Z.; Abdullah, C.A.C.; Ibrahim, W.N.W. Time dependent effect of chronic embryonic exposure to ethanol on zebrafish: Morphology, biochemical and anxiety alterations. Behav. Brain Res. 2017, 332, 40–49. [Google Scholar] [CrossRef]
- Bakar, N.A.; Sata, N.S.A.M.; Ramlan, N.F.; Ibrahim, W.N.W.; Zulkifli, S.Z.; Abdullah, C.A.C.; Ahmad, S.; Amal, M.N.A. Evaluation of the neurotoxic effects of chronic embryonic exposure with inorganic mercury on motor and anxiety-like responses in zebrafish (Danio rerio) larvae. Neurotoxicol. Teratol. 2017, 59, 53–61. [Google Scholar] [CrossRef]
- Lajis, A.A. Zebrafish embryo as an animal model for the treatment of hyperpigmentation in cosmetic dermatology medicine. Medicina 2018, 54, 35. [Google Scholar] [CrossRef] [Green Version]
- Colanesi, S.; Taylor, K.L.; Temperley, N.D.; Lundegaard, P.R.; Liu, D.; North, T.E.; Ishizaki, H.; Kelsh, R.N.; Patton, E.E. Small molecule screening identifies targetable zebrafish pigmentation pathways. Pigm. Cell Melanoma Res. 2012, 25, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Choi, T.Y.; Kim, J.H.; Ko, D.H.; Kim, C.H.; Hwang, J.S.; Ahn, S.; Kim, S.Y.; Kim, C.D.; Lee, J.H.; Yoon, T.J. Zebrafish as a new model for phenotype-based screening of melanogenic regulatory compounds. Pigm. Cell Res. 2007, 20, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.T.; Fong, T.H.; Chen, H.M.; Chang, C.Y.; Wang, Y.H.; Chern, C.Y.; Chen, Y.H. Toxicity assessments of chalcone and some synthetic chalcone analogues in a zebrafish model. Molecules 2014, 19, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Dharmaratne, R.W.; Dhammika Nanayakkara, N.; Khan, I.A. Kavalactones from Piper methysticum, and their 13C NMR spectroscopic analyses. Phytochemistry 2002, 59, 429–433. [Google Scholar] [CrossRef]
- Abu Bakar, A.; Akhtar, M.N.; Mohd Ali, N.; Yeap, S.K.; Quah, C.K.; Loh, W.S.; Alitheen, N.B.; Zareen, S.; Ul-Haq, Z.; Shah, S.A.A. Design, synthesis and docking studies of flavokawain b type chalcones and their cytotoxic effects on MCF-7 and MDA-MB-231 cell lines. Molecules 2018, 23, 616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, N.M.; Akhtar, M.N.; Ky, H.; Lim, K.L.; Abu, N.; Zareen, S.; Ho, W.Y.; Alan-Ong, H.K.; Tan, S.W.; Alitheen, N.B.; et al. Flavokawain derivative FLS induced G2/M arrest and apoptosis on breast cancer MCF-7 cell line. Drug Des. Dev. Ther. 2016, 10, 1897. [Google Scholar]
- Kamaldin, M.N.; Akhtar, M.N.; Mohamad, A.S.; Lajis, N.; Perimal, E.K.; Akira, A.; Ming-Tatt, L.; Israf, D.A.; Sulaiman, M.R. Peripheral antinociception of a chalcone, flavokawin B and possible involvement of the nitric oxide/cyclic guanosine monophosphate/potassium channels pathway. Molecules 2013, 18, 4209–4220. [Google Scholar] [CrossRef]
- Kwon, D.; Ju, S.M.; Youn, G.S.; Choi, S.Y.; Park, J. Suppression of iNOS and COX-2 expression by flavokawain a via blockade of NF-κB and AP-1 activation in RAW 264.7 macrophages. Food Chem. Toxicol. 2013, 58, 479–486. [Google Scholar] [CrossRef]
- Abu, N.; Mohamed, N.E.; Tangarajoo, N.; Yeap, S.K.; Akhtar, M.N.; Abdullah, M.P.; Omar, A.R.; Alitheen, N.B. In vitro toxicity and in vivo immunomodulatory effects of Flavokawain A and Flavokawain B in Balb/C Mice. Nat. Prod. Commun. 2015, 10. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, M.N.; Sakeh, N.M.; Zareen, S.; Gul, S.; Lo, K.M.; Ul-Haq, Z.; Shah, S.A.A.; Ahmad, S. Design and synthesis of chalcone derivatives as potent tyrosinase inhibitors and their structural activity relationship. J. Mol. Struct. 2015, 1085, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Jin, M.L.; Park, S.Y.; Kim, Y.H.; Park, G.; Son, H.J.; Lee, S.J. Suppression of α-MSH and IBMX-induced melanogenesis by cordycepin via inhibition of CREB and Mitf, and activation of PI3K/Akt and ERK-dependent mechanisms. Int. J. Mol. Med. 2012, 29, 119–124. [Google Scholar]
- Molagoda, I.M.N.; Choi, Y.H.; Lee, S.; Sung, J.; Lee, C.R.; Lee, H.G.; Lim, J.; Lee, K.J.; Jeon, Y.J.; Ma, J.; et al. Ethanolic extract of Hippocampus abdominalis exerts anti-melanogenic effects in B16F10 melanoma cells and zebrafish larvae by activating the erk signaling pathway. Cosmetics 2020, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Briganti, S.; Camera, E.; Picardo, M. Chemical and instrumental approaches to treat hyperpigmentation. Pigm. Cell Res. 2003, 16, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Parvez, S.; Kang, M.; Chung, H.; Bae, H. Naturally occurring tyrosinase inhibitors: Mechanism and applications in skin health, cosmetics and agriculture industries. Phytother. Res. 2007, 21, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Kubo, I.; Kinst-Hori, I.; Chaudhuri, S.K.; Kubo, Y.; Saánchez, Y.; Ogura, T. Flavonols from Heterotheca inuloides: Tyrosinase inhibitory activity and structural criteria. Bioorg. Med. Chem. 2000, 8, 1749–1755. [Google Scholar] [CrossRef]
- Saewan, N.; Koysomboon, S.; Chantrapromma, K. Anti-tyrosinase and anti-cancer activities of flavonoids from Blumea balsamifera DC. J. Med. Plants Res. 2011, 5, 1018–1025. [Google Scholar]
- Venturelli, S.; Burkard, M.; Biendl, M.; Lauer, U.M.; Frank, J.; Busch, C. Prenylated chalcones and flavonoids for the prevention and treatment of cancer. Nutrition 2016, 32, 1171–1178. [Google Scholar] [CrossRef]
- Khatib, S.; Nerya, O.; Musa, R.; Shmuel, M.; Tamir, S.; Vaya, J. Chalcones as potent tyrosinase inhibitors: The importance of a 2,4-substituted resorcinol moiety. Bioorg. Med. Chem. 2005, 13, 433–441. [Google Scholar] [CrossRef]
- Chaves, O.A.; de Barros, L.S.; de Oliveira, M.C.; Sant’Anna, C.M.R.; Ferreira, A.B.; da Silva, F.A.; Cesarin-Sobrinho, D.; Netto-Ferreira, J.C. Biological interactions of fluorinated chalcones: Stimulation of tyrosinase activity and binding to bovine serum albumin. J. Fluor. Chem. 2017, 199, 30–38. [Google Scholar] [CrossRef]
- Singh, P.; Anand, A.; Kumar, V. Recent developments in biological activities of chalcones: A mini review. Eur. J. Med. Chem. 2014, 85, 758–777. [Google Scholar] [CrossRef]
- Nerya, O.; Musa, R.; Khatib, S.; Tamir, S.; Vaya, J. Chalcones as potent tyrosinase inhibitors: The effect of hydroxyl positions and numbers. Phytochemistry 2004, 65, 1389–1395. [Google Scholar] [CrossRef] [PubMed]
- Hearing, V.J.; Tsukamoto, K. Enzymatic control of pigmentation in mammals. FASEB J. 1991, 5, 2902–2909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, R.; Hearing, V.; Urabe, K.; Aroca, P.; Spritz, R. Mutational mapping of the catalytic activities of human tyrosinase. J. Biol. Chem. 1992, 267, 23707–23712. [Google Scholar]
- Jeong, H.J.; Lee, C.S.; Choi, J.; Hong, Y.D.; Shin, S.S.; Park, J.S.; Lee, J.H.; Lee, S.; Yoon, K.D.; Ko, J. Flavokawains B and C, melanogenesis inhibitors, isolated from the root of Piper methysticum and synthesis of analogs. Bioorg. Med. Chem. Lett. 2015, 25, 799–802. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Zhao, B.; Liu, Y.; Wang, R.; Yang, Y.; Yang, L.; Dong, C. Mitf-M regulates melanogenesis in mouse melanocytes. J. Dermatol. Sci. 2018, 90, 253–262. [Google Scholar] [CrossRef] [Green Version]
- Hridya, H.; Amrita, A.; Mohan, S.; Gopalakrishnan, M.; Dakshinamurthy, T.K.; Doss, G.P.; Siva, R. Functionality study of santalin as tyrosinase inhibitor: A potential depigmentation agent. Int. J. Biol. Macromol. 2016, 86, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.K.; Funasaka, Y.; Slominski, A.; Ermak, G.; Hwang, J.; Pawelek, J.M.; Ichihashi, M. Production and release of proopiomelanocortin (POMC) derived peptides by human melanocytes and keratinocytes in culture: Regulation by ultraviolet B. BBA Mol. Cell Res. 1996, 1313, 130–138. [Google Scholar] [CrossRef] [Green Version]
- Ballotti, R. Cyclic AMP a key messenger in the regulation of skin pigmentation. Pigm. Cell Res. 2000, 13, 60–69. [Google Scholar]
- Hill, C.; Dunbar, J.C. The effects of acute and chronic alpha melanocyte stimulating hormone (αMSH) on cardiovascular dynamics in conscious rats. Peptides 2002, 23, 1625–1630. [Google Scholar] [CrossRef]
- Feichtinger, R.G.; Pétervári, E.; Zopf, M.; Vidali, S.; Aminzadeh-Gohari, S.; Mayr, J.A.; Kofler, B.; Balaskó, M. Effects of alpha-melanocyte-stimulating hormone on mitochondrial energy metabolism in rats of different age-groups. Neuropeptides 2017, 64, 123–130. [Google Scholar] [CrossRef]
- Lim, J.; Nam, S.; Jeong, J.H.; Kim, M.J.; Yang, Y.; Lee, M.S.; Lee, H.G.; Ryu, J.H.; Lim, J.S. Kazinol U inhibits melanogenesis through the inhibition of tyrosinase-related proteins via AMP kinase activation. Br. J. Pharmacol. 2019, 176, 737–750. [Google Scholar] [CrossRef] [Green Version]
- Zenger, K.; Dutta, S.; Wolff, H.; Genton, M.G.; Kraus, B. In vitro structure-toxicity relationship of chalcones in human hepatic stellate cells. Toxicology 2015, 336, 26–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thieury, C.; Lebouvier, N.; Le Guével, R.; Barguil, Y.; Herbette, G.; Antheaume, C.; Hnawia, E.; Asakawa, Y.; Nour, M.; Guillaudeux, T. Mechanisms of action and structure-activity relationships of cytotoxic flavokawain derivatives. Bioorg. Med. Chem. 2017, 25, 1817–1829. [Google Scholar] [CrossRef] [PubMed]
- Caballero, M.V.; Candiracci, M. Zebrafish as screening model for detecting toxicity and drugs efficacy. J. Unexplored Med. Data 2018, 3, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Chan, Y.; Kim, K.; Cheah, S. Inhibitory effects of Sargassum polycystum on tyrosinase activity and melanin formation in B16F10 murine melanoma cells. J. Ethnopharmacol. 2011, 137, 1183–1188. [Google Scholar] [CrossRef] [PubMed]
- Lin, V.C.H.; Ding, H.Y.; Kuo, S.Y.; Chin, L.W.; Wu, J.Y.; Chang, T.S. Evaluation of in vitro and in vivo depigmenting activity of raspberry ketone from Rheum officinale. Int. J. Mol. Sci. 2011, 12, 4819–4835. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Matthews, M.; Trevarrow, B.; Matthews, J.A. Virtual tour of the guide for zebrafish users. Resource 2002, 31, 34–40. [Google Scholar]
- Nasiadka, A.; Clark, M.D. Zebrafish breeding in the laboratory environment. ILAR J. 2012, 53, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Padilla, S.; Hunter, D.L.; Padnos, B.; Frady, S.; MacPhail, R.C. Assessing locomotor activity in larval zebrafish: Influence of extrinsic and intrinsic variables. Neurotoxicol. Teratol. 2011, 33, 624–630. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dynam. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- OECD. Test No. 236: Fish Embryo Acute Toxicity (FET) test. In OECD Guidelines for the Testing of Chemicals, Section 2; OECD Publishing: Paris, France, 2013; pp. 1–22. [Google Scholar]
- Ellis, L.D.; Soo, E.C.; Achenbach, J.C.; Morash, M.G.; Soanes, K.H. Use of the zebrafish larvae as a model to study cigarette smoke condensate toxicity. PLoS ONE 2014, 9, e115305. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, X.; Yin, X.; Shi, M.; Dahlgren, R.A.; Wang, H. Toxicity assessment of combined fluoroquinolone and tetracycline exposure in zebrafish (Danio rerio). Environ. Toxicol. 2016, 31, 736–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas, Q.; Ashraf, Z.; Hassan, M.; Nadeem, H.; Latif, M.; Afzal, S.; Seo, S.Y. Development of highly potent melanogenesis inhibitor by in vitro, in vivo and computational studies. Drug Des. Dev. Ther. 2017, 11, 2029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, K.D.; Chen, H.J.; Wang, C.S.; Lum, C.C.; Wu, S.P.; Lin, S.P.; Cheng, K.C. Extract of Ganoderma formosanum mycelium as a highly potent tyrosinase inhibitor. Sci. Rep. 2016, 6, 32854. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.C.; Kim, S.; Hwang, K.-S.; Kim, C.H. p-coumaric acid potently down-regulates zebrafish embryo pigmentation: Comparison of in vivo assay and computational molecular modeling with phenylthiourea. Biomed. Sci. Lett. 2017, 23, 8–16. [Google Scholar] [CrossRef] [Green Version]
- Renquist, B.J.; Zhang, C.; Williams, S.Y.; Cone, R.D. Development of an assay for high-throughput energy expenditure monitoring in the zebrafish. Zebrafish 2013, 10, 343–352. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Samples of the compounds FLA and FLB are available from Assoc. Muhammad Nadeem Akhtar. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences | Accession Number |
|---|---|---|
| Tyr | Forward: TTG CCA CTT CAT GTC ATC ATA GAA TAT T Reverse: TTT ATC AAA GGT GTG ACT GCT ATA CAA AT | NM_011661.5 |
| Trp-1 | Forward: GCT GCA GGA GCC TTC TTT CTC Reverse: AAG ACG CTG CAC TGC TGG TCT | NM_031202.3 |
| Trp-2 | Forward: GGA TGA CCG TGA GCA ATG GCC Reverse: CGG TTG TGA CCA ATG GGT GCC | NM_010024.3 |
| Mitf | Forward: TAC AGA AAG TAG AGG GAG GAG GAC TAA G Reverse: CAC AGT TGG AGT TAA GAG TGA GCA TAG CC | NM_008601.3 |
| GAPDH | Forward: ACC ACA GTC CAT GCC ATC AC Reverse: TCC ACC ACC CTG TTG CTG TA | NM_008084.3 |
| ß-actin | Forward: ACC GTG AAA AGA TGA CCC AG Reverse: TAC GGA TGT CAA CGT CAC AC | NC_007393.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohd Sakeh, N.; Md Razip, N.N.; Mohd Ma’in, F.I.; Abdul Bahari, M.N.; Latif, N.; Akhtar, M.N.; Balia Yusof, Z.N.; Ahmad, S. Melanogenic Inhibition and Toxicity Assessment of Flavokawain A and B on B16/F10 Melanoma Cells and Zebrafish (Danio rerio). Molecules 2020, 25, 3403. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25153403
Mohd Sakeh N, Md Razip NN, Mohd Ma’in FI, Abdul Bahari MN, Latif N, Akhtar MN, Balia Yusof ZN, Ahmad S. Melanogenic Inhibition and Toxicity Assessment of Flavokawain A and B on B16/F10 Melanoma Cells and Zebrafish (Danio rerio). Molecules. 2020; 25(15):3403. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25153403
Chicago/Turabian StyleMohd Sakeh, Nurshafika, Nurliyana Najwa Md Razip, Farah Idayu Mohd Ma’in, Mohammad Nazri Abdul Bahari, Naimah Latif, Muhammad Nadeem Akhtar, Zetty Norhana Balia Yusof, and Syahida Ahmad. 2020. "Melanogenic Inhibition and Toxicity Assessment of Flavokawain A and B on B16/F10 Melanoma Cells and Zebrafish (Danio rerio)" Molecules 25, no. 15: 3403. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25153403