
Free Amino Acids in Three Pleurotus Species Cultivated on Agricultural and Agro-Industrial By-Products

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Protein and FAAs Content in Mushroom Cultivation Substrates
2.2. FAAs Profiles in Mushrooms
2.2.1. Protein and Total FAAs Content
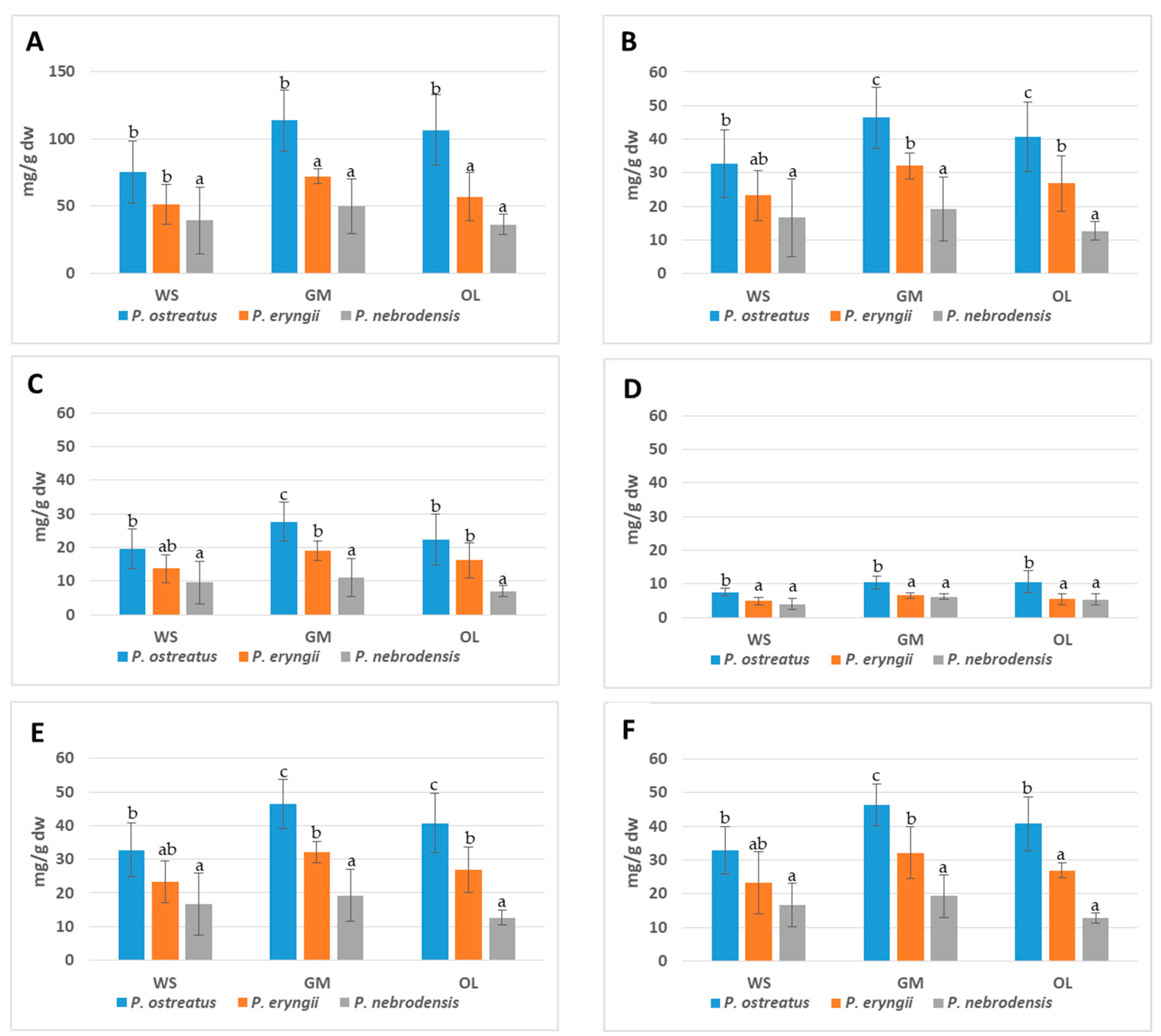
2.2.2. Essential FAAs
2.2.3. Branched Chain Amino Acids, GABA and Ornithine
2.2.4. Taste Attributes
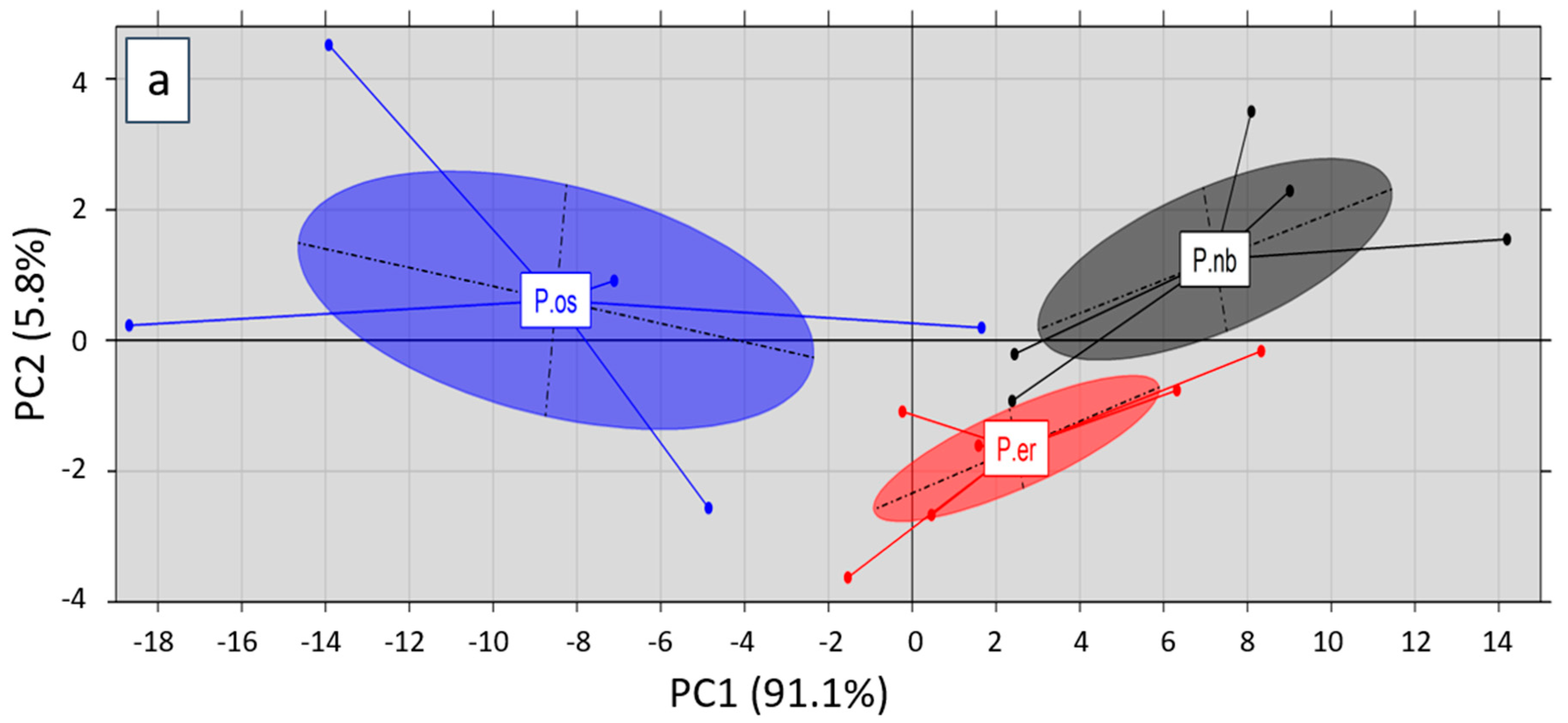
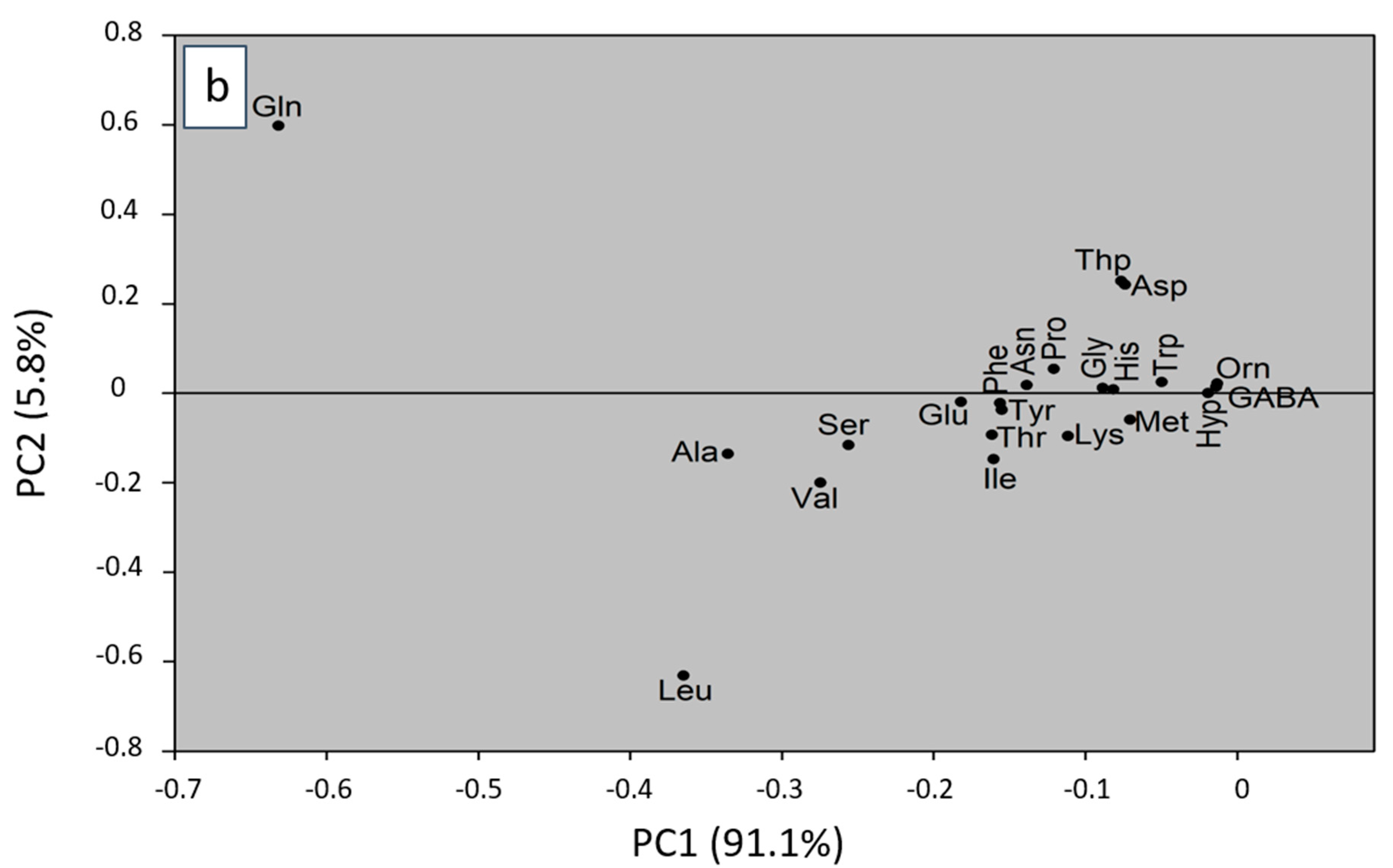
2.3. Principal Component Analysis
3. Materials and Methods
3.1. Biological Material and Mushroom Cultivation Substrates
3.2. Sample Pre-Treatment
3.3. Extraction of Free Amino Acids
3.4. Crude Protein Content
3.5. Derivatization of Free Amino Acids
3.6. GC/MS Analysis of Free Amino Acids
3.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Valverde, M.E.; Hernández-Pérez, T.; Paredes-López, O. Edible mushrooms: Improving human health and promoting quality life. Int. J. Microbiol. 2015, 2015, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Dicks, L.; Ellinger, S. Effect of the intake of oyster mushrooms (Pleurotus ostreatus) on cardiometabolic parameters—A systematic review of clinical trials. Nutrients 2020, 12, 1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gargano, M.L.; van Griensven, L.J.L.D.; Isikhuemhen, O.S.; Lindequist, U.; Venturella, G.; Wasser, S.P.; Zervakis, G.I. Medicinal mushrooms: Valuable biological resources of high exploitation potential. Plant Biosyst. 2017, 151, 548–565. [Google Scholar] [CrossRef]
- Royse, D.J.; Baars, J.; Tan, Q. Current overview of mushroom production in the world. In Edible and Medicinal Mushrooms: Technology and Applications; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2017; pp. 5–13. [Google Scholar] [CrossRef]
- Zervakis, G.I.; Ntougias, S.; Gargano, M.L.; Besi, M.I.; Polemis, E.; Typas, M.A.; Venturella, G. A reappraisal of the Pleurotus eryngii complex-New species and taxonomic combinations based on the application of a polyphasic approach, and an identification key to Pleurotus taxa associated with Apiaceae plants. Fungal Biol. 2014, 118, 814–834. [Google Scholar] [CrossRef]
- Ritota, M.; Manzi, P. Pleurotus spp. Cultivation on different agri-food by-products: Example of biotechnological application. Sustainability 2019, 11, 5049. [Google Scholar] [CrossRef] [Green Version]
- Zervakis, G.I.; Koutrotsios, G. Solid state fermentation of plant residues and agro-industrial wastes for the production of medicinal mushrooms. In Medicinal Plants and Fungi: Recent Advances in Research and Development; Agrawal, D.C., Tsay, H.-S., Shyur, L.-F., Wu, Y.-C., Wang, S.-Y., Eds.; Springer Nature Singapore: Singapore, 2017; pp. 365–396. [Google Scholar] [CrossRef]
- Koutrotsios, G.; Kalogeropoulos, N.; Kaliora, A.C.; Zervakis, G.I. Toward an increased functionality in oyster (Pleurotus) mushrooms produced on grape marc or olive mill wastes serving as sources of bioactive compounds. J. Agric. Food Chem. 2018, 66, 5971–5983. [Google Scholar] [CrossRef]
- Koutrotsios, G.; Mountzouris, K.C.; Chatzipavlidis, I.; Zervakis, G.I. Bioconversion of lignocellulosic residues by Agrocybe cylindracea and Pleurotus ostreatus mushroom fungi-Assessment of their effect on the final product and spent substrate properties. Food Chem. 2014, 161, 127–135. [Google Scholar] [CrossRef]
- Koutrotsios, G.; Patsou, M.; Mitsou, E.K.; Bekiaris, G.; Kotsou, M.; Tarantilis, P.A.; Pletsa, V.; Kyriacou, A.; Zervakis, G.I. Valorization of olive by-products as substrates for the cultivation of Ganoderma lucidum and Pleurotus ostreatus mushrooms with enhanced functional and prebiotic properties. Catalysts 2019, 9, 537. [Google Scholar] [CrossRef] [Green Version]
- Bao, X.; Feng, Z.; Yao, J.; Li, T.; Yin, Y. Roles of dietary amino acids and their metabolites in pathogenesis of inflammatory bowel disease. Mediators Inflamm. 2017, 2017, 1–9. [Google Scholar] [CrossRef]
- Mau, J.L.; Lin, Y.P.; Chen, P.T.; Wu, Y.H.; Peng, J.T. Flavor compounds in king oyster mushrooms Pleurotus eryngii. J. Agric. Food Chem. 1998, 46, 4587–4591. [Google Scholar] [CrossRef]
- Mau, J.-L.; Lin, H.-C.; Chen, C.-C. Non-volatile components of several medicinal mushrooms. Food Res. Int. 2001, 34, 521–526. [Google Scholar] [CrossRef]
- Tseng, Y.H.; Lee, Y.L.; Li, R.C.; Mau, J.L. Non-volatile flavour components of Ganoderma tsugae. Food Chem. 2005, 90, 409–415. [Google Scholar] [CrossRef]
- Yang, J.-H.; Lin, H.-C.; Mau, J.-L. Non-volatile taste components of several commercial mushrooms. Food Chem. 2001, 72, 465–471. [Google Scholar] [CrossRef]
- Mau, J.L. The umami taste of edible and medicinal mushrooms. Int. J. Med. Mushrooms 2005, 7, 119–125. [Google Scholar] [CrossRef]
- Patel, S.; Goyal, A. Recent developments in mushrooms as anti-cancer therapeutics: A review. 3 Biotech 2012, 2, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, C. Modern aspects of mushroom culture technology. Appl. Microbiol. Biotechnol. 2004, 64, 756–762. [Google Scholar] [CrossRef]
- EZ:faast Free (Physiological) Amino Acid Analysis by GC-MS, User’s Manual. Available online: https://phenomenex.blob.core.windows.net/documents/3eaba18b-9083-4760-bbe5-a511c0d9df21.pdf (accessed on 7 November 2019).
- Badawy, A.A.B.; Morgan, C.J.; Turner, J.A. Application of the Phenomenex EZ: faastTM amino acid analysis kit for rapid gas-chromatographic determination of concentrations of plasma tryptophan and its brain uptake competitors. Amino Acids 2008, 34, 587–596. [Google Scholar] [CrossRef] [Green Version]
- Baddi, G.A.; Alburquerque, J.A.; Gonzálvez, J.; Cegarra, J.; Hafidi, M. Chemical and spectroscopic analyses of organic matter transformations during composting of olive mill wastes. Int. Biodeterior. Biodegrad. 2004, 54, 39–44. [Google Scholar] [CrossRef]
- Feedipedia, an On-Line Encyclopedia of Animal Feeds. Available online: https://www.feedipedia.org (accessed on 2 July 2020).
- Li, W.; Gu, Z.; Yang, Y.; Zhou, S.; Liu, Y.; Zhang, J. Non-volatile taste components of several cultivated mushrooms. Food Chem. 2014, 143, 427–431. [Google Scholar] [CrossRef]
- Cohen, N.; Cohen, J.; Asatiani, M.D.; Varshney, V.K.; Yu, H.T.; Yang, Y.C.; Li, Y.H.; Mau, J.L.; Wasser, S.P. Chemical composition and nutritional and medicinal value of fruit bodies and submerged cultured mycelia of culinary-medicinal higher basidiomycetes mushrooms. Int. J. Med. Mushrooms 2014, 16, 273–291. [Google Scholar] [CrossRef]
- Bach, F.; Helm, C.V.; Bellettini, M.B.; Maciel, G.M.; Haminiuk, C.W.I. Edible mushrooms: A potential source of essential amino acids, glucans and minerals. Int. J. Food Sci. Technol. 2017, 52, 2382–2392. [Google Scholar] [CrossRef]
- Kim, M.Y.; Chung, M.; Lee, S.J.; Ahn, J.K.; Kim, E.H.; Kim, M.J.; Kim, S.L.; Moon, H.I.; Ro, H.M.; Kang, E.Y.; et al. Comparison of free amino acid, carbohydrates concentrations in Korean edible and medicinal mushrooms. Food Chem. 2009, 113, 386–393. [Google Scholar] [CrossRef]
- Guo, L.Q.; Lin, J.Y.; Lin, J.F. Non-volatile components of several novel species of edible fungi in China. Food Chem. 2007, 100, 643–649. [Google Scholar] [CrossRef]
- Yin, C.; Fan, X.; Fan, Z.; Shi, D.; Yao, F.; Gao, H. Comparison of non-volatile and volatile flavor compounds in six Pleurotus mushrooms. J. Sci. Food Agric. 2019, 99, 1691–1699. [Google Scholar] [CrossRef]
- Gao, S.; Huang, Z.; Feng, X.; Bian, Y.; Huang, W.; Liu, Y. Bioconversion of rice straw agro-residues by Lentinula edodes and evaluation of non-volatile taste compounds in mushrooms. Sci. Rep. 2020, 10, 1814. [Google Scholar] [CrossRef]
- Manzi, P.; Gambelli, L.; Marconi, S.; Vivanti, V.; Pizzoferrato, L. Nutrients in edible mushrooms: An inter-species comparative study. Food Chem. 1999, 65, 477–482. [Google Scholar] [CrossRef]
- Breene, W.M. Nutritional and medicinal value of specialty mushrooms. J. Food Prot. 1990, 53, 883–894. [Google Scholar] [CrossRef]
- Dundar, A.; Acay, H.; Yildiz, A. Yield performances and nutritional contents of three oyster mushroom species cultivated on wheat stalk. Afr. J. Biotechnol. 2008, 7, 3497–3501. [Google Scholar] [CrossRef]
- Miyazawa, N.; Matsuoka, H.; Ozawa, Y. Palatability characteristics of Pleurotus eryngii var. tuoliensis. Nippon Shokuhin Kagaku Kogaku Kaishi 2012, 59, 153–160. [Google Scholar] [CrossRef]
- Yoshida, H.; Sasaki, H.; Fujimoto, S.; Sugahara, T. The chemical components of the vegetative mycelia of Basidiomycetes. Nippon Shokuhin Kogyo Gakkai-Shi 1996, 43, 748–755. [Google Scholar] [CrossRef]
- Patil, S.S.; Ahmed, S.A.; Telang, S.M.; Baig, M.M.V. The nutritional value of Pleurotus ostreatus (Jacq.:Fr.) Kumm cultivated on different lignocellulosic agro-wastes. Innov. Rom. Food Biotechnol. 2010, 7, 66–76. [Google Scholar]
- Bamidele, O.P.; Fasogbon, B.M. Nutritional and functional properties of maize-oyster mushroom (Zea mays-Pleurotus ostreatus) based composite flour and its storage stability. Open Agric. 2020, 5, 40–49. [Google Scholar] [CrossRef] [Green Version]
- Oyetayo, F.L.; Akindahunsi, A.; Oyetayo, V.O. Chemical profile and amino acids composition of edible mushrooms Pleurotus sajor-caju. Nutr. Health 2007, 18, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Papada, E.; Amerikanou, C.; Torović, L.; Kalogeropoulos, N.; Tzavara, C.; Forbes, A.; Kaliora, A.C. Plasma free amino acid profile in quiescent Inflammatory Bowel Disease patients orally administered with Mastiha (Pistacia lentiscus); a randomised clinical trial. Phytomedicine 2019, 56, 40–47. [Google Scholar] [CrossRef] [Green Version]
- Kothari, D.; Patel, S.; Kim, S.K. Anticancer and other therapeutic relevance of mushroom polysaccharides: A holistic appraisal. Biomed. Pharmacother. 2018, 105, 377–394. [Google Scholar] [CrossRef]
- Khan, A.A.; Gani, A.; Khanday, F.A.; Masoodi, F.A. Biological and pharmaceutical activities of mushroom β-glucan discussed as a potential functional food ingredient. Bioact. Carbohydrates Diet. Fibre 2018, 16, 1–13. [Google Scholar] [CrossRef]
- Beluhan, S.; Ranogajec, A. Chemical composition and non-volatile components of Croatian wild edible mushrooms. Food Chem. 2011, 124, 1076–1082. [Google Scholar] [CrossRef]
- Diana, M.; Quílez, J.; Rafecas, M. Gamma-aminobutyric acid as a bioactive compound in foods: A review. J. Funct. Foods 2014, 10, 407–420. [Google Scholar] [CrossRef]
- Sivashanmugam, M.; Jaidev, J.; Umashankar, V.; Sulochana, K.N. Ornithine and its role in metabolic diseases: An appraisal. Biomed. Pharmacother. 2017, 86, 185–194. [Google Scholar] [CrossRef]
- Lin, S.Y.; Chen, Y.K.; Yu, H.T.; Barseghyan, G.S.; Asatiani, M.D.; Wasser, S.P.; Mau, J.L. Comparative study of contents of several bioactive components in fruiting bodies and mycelia of culinary-medicinal mushrooms. Int. J. Med. Mushrooms 2013, 15, 315–323. [Google Scholar] [CrossRef]
- Park, Y.-J.; Oh, T.-S.; Jang, M.-J. Effect of adding amino acids on the production of Gamma-Aminobutyric Acid (GABA) by mycelium of Lentinula edodes. Int. J. Food Eng. 2019, 15, 1–6. [Google Scholar] [CrossRef]
- Narayan, V.S.; Nair, P.M. Metabolism, enzymology and possible roles of 4-aminobutyrate in higher plants. Phytochemistry 1990, 29, 367–375. [Google Scholar] [CrossRef]
- Yamaguchi, S. Basic properties of umami and effects on humans. Physiol. Behav. 1991, 49, 833–841. [Google Scholar] [CrossRef]
- Kirimura, J.; Shimizu, A.; Kimizuka, A.; Ninomiya, T.; Katsuya, N. Contribution of peptides and amino acids to the taste of foods. J. Agric. Food Chem. 1969, 17, 689–695. [Google Scholar] [CrossRef]
- Dong, M.; Qin, L.; Xue, J.; Du, M.; Lin, S.-Y.; Xu, X.-B.; Zhu, B.-W. Simultaneous quantification of free amino acids and 5′-nucleotides in shiitake mushrooms by stable isotope labeling-LC-MS/MS analysis. Food Chem. 2018, 268, 57–65. [Google Scholar] [CrossRef]
- Elmore, S.J.; Koutsidis, G.; Dodson, A.T.; Mottram, D.S.; Wedzicha, B.L. Measurement of acrylamide and its precursors in potato, wheat, and rye model systems. J. Agric. Food Chem. 2005, 53, 1286–1293. [Google Scholar] [CrossRef]
- Kalač, P. Chemical composition and nutritional value of European species of wild growing mushrooms: A review. Food Chem. 2009, 113, 9–16. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |




{kind=link}
{kind=link}
{kind=link}
| Essential AAs | BCAAs | MSG-Like AAs | Bitter Taste AAs | Sweet Taste AAs |
|---|---|---|---|---|
| Valine | Valine | Aspartic acid | Valine | Threonine |
| Leucine | Leucine | Glutamic acid | Methionine | Serine |
| Isoleucine | Isoleucine | Leucine | Glycine | |
| Threonine | Isoleucine | Alanine | ||
| Methionine | Phenylalanine | Proline | ||
| Phenylalanine | Histidine | |||
| Lysine | Tryptophan | |||
| Histidine | ||||
| Tryptophan |
| Amino acid | Abbreviation | WS | GM | OL |
|---|---|---|---|---|
| Alanine | Ala | 0.003 ± 0.001 a | 0.065 ± 0.007 c | 0.023 ± 0.005 b |
| Glycine | Gly | 0.002 ± 0.001 a | 0.022 ± 0.004 c | 0.009 ± 0.000 b |
| Valine | Val | 0.002 ± 0.001 a | 0.010 ± 0.002 b | 0.004 ± 0.000 a |
| Leucine | Leu | 0.008 ± 0.005 a | 0.032 ± 0.006 b | 0.008 ± 0.001 a |
| Isoleucine | Ile | 0.004 ± 0.002 a | 0.019 ± 0.004 b | 0.006 ± 0.001 a |
| Threonine | Thr | 0.004 ± 0.002 a | 0.020 ± 0.005 b | 0.006 ± 0.000 a |
| Serine | Ser | 0.005 ± 0.002 a | 0.043 ± 0.003 c | 0.012 ± 0.001 b |
| Proline | Pro | 0.004 ± 0.001 a | 0.091 ± 0.020 b | 0.022 ± 0.001 a |
| Asparagine | Asn | 0.003 ± 0.001 a | 0.137 ± 0.002 c | 0.061 ± 0.015 b |
| Thioproline | Thp | 0.013 ± 0.005 a | 0.105 ± 0.003 c | 0.147 ± 0.004 b |
| Aspartic acid | Asp | 0.012 ± 0.003 a | 0.097 ± 0.006 c | 0.138 ± 0.003 b |
| Methionine | Met | 0.003 ± 0.002 a | 0.001 ± 0.001 a | nd |
| Hydroxyproline | Hyp | nd | 0.003 ± 0.001 | nd |
| Glutamic acid | Glu | 0.155 ± 0.026 a | 0.204 ± 0.002 c | 0.064 ± 0.002 b |
| Phenylalanine | Phe | 0.006 ± 0.003 a | 0.024 ± 0.005 b | 0.006 ± 0.000 a |
| Glutamine | Gln | 0.037 ± 0.010 a | 0.093 ± 0.003 b | 0.037 ± 0.004 a |
| Ornithine | Orn | 0.022 ± 0.000 a | 0.025 ± 0.001 b | 0.022 ± 0.000 a |
| Lysine | Lys | 0.033 ± 0.001 a | 0.057 ± 0.006 b | 0.035 ± 0.001 a |
| Histidine | His | 0.0003 ± 0.000 a | 0.023 ± 0.005 b | 0.002 ± 0.000 a |
| Tyrosine | Tyr | 0.006 ± 0.002 a | 0.018 ± 0.003 b | 0.008 ± 0.001 a |
| Tryptophan | Trp | 0.001 ± 0.001 a | 0.085 ± 0.001 b | 0.093 ± 0.009 b |
| γ-Aminobutyric acid | GABA | nd | 0.058 ± 0.003 a | 0.008 ± 0.002 b |
| Sum of amino acids | 0.319 ± 0.070 | 1.231 ± 0.077 | 0.711 ± 0.033 | |
| Crude protein | 85.20 ± 4.34 a | 116.60 ± 5.04 b | 131.12 ± 0.09 c |
| Amino Acid | Substrate | P. ostreatus | P. eryngii | P. nebrodensis | |||
|---|---|---|---|---|---|---|---|
| LGAM 14 | LGAM 11 | UPA 12 | LGAM 212 | UPA 6 | LGAM 162 | ||
| Alanine | WS | 7.22 ± 1.10 a | 9.77 ± 2.29 a | 4.74 ± 0.32 a | 7.27 ± 1.08 a | 2.82 ± 0.19 a | 7.20 ± 1.47 |
| GM | 10.34 ± 2.31 ab | 13.33 ± 2.71 a | 8.26 ± 1.11 b | 8.54 ± 1.60 a | 4.27 ± 0.69 a | 7.53 ± 3.49 | |
| OL | 11.72 ± 2.69 b | 10.42 ± 3.83 a | 4.74 ± 1.99 a | 7.61 ± 1.34 a | 4.08 ± 1.04 a | * | |
| Glycine | WS | 1.03 ± 0.05 a | 1.94 ± 0.97 a | 1.03 ± 0.10 a | 1.21 ± 0.56 a | 0.29 ± 0.04 a | 1.51 ± 0.17 |
| GM | 1.85 ± 0.20 a | 2.99 ± 0.35 a | 1.88 ± 0.18 b | 1.65 ± 0.11 a | 0.73 ± 0.19 b | 1.61 ± 0.55 | |
| OL | 3.29 ± 1.13 b | 2.12 ± 0.82 a | 0.98 ± 0.41 a | 1.45 ± 0.10 a | 0.50 ± 0.06 ab | * | |
| Valine | WS | 4.24 ± 0.34 a | 6.94 ± 2.08 a | 3.24 ± 0.12 a | 4.81 ± 1.12 a | 1.05 ± 0.14 a | 3.97 ± 0.61 |
| GM | 6.94 ± 0.84 ab | 9.49 ± 1.05 a | 5.26 ± 0.56 b | 6.24 ± 0.53 a | 1.76 ± 0.80 a | 3.98 ± 1.54 | |
| OL | 8.44 ± 2.19 b | 7.13 ± 2.01 a | 3.35 ± 1.25 a | 5.59 ± 0.83 a | 1.56 ± 0.26 a | * | |
| Leucine | WS | 8.59 ± 1.32 a | 12.24 ± 3.27 a | 5.11 ± 0.50 a | 8.88 ± 1.81 a | 2.24 ± 0.15 a | 8.29 ± 1.89 |
| GM | 12.31 ± 2.50 a | 16.45 ± 2.33 a | 8.81 ± 1.11 a | 11.24 ± 1.51 a | 4.51 ± 1.22 b | 7.81 ± 3.37 | |
| OL | 9.66 ± 8.19 a | 9.95 ± 6.70 a | 7.30 ± 3.07 a | 10.10 ± 1.84 a | 4.34 ± 1.10 b | * | |
| Isoleucine | WS | 2.80 ± 0.29 a | 4.13 ± 0.96 a | 2.15 ± 0.06 a | 3.15 ± 0.77 a | 0.67 ± 0.03 a | 2.81 ± 0.53 |
| GM | 4.35 ± 0.66 ab | 5.79 ± 0.81 a | 3.40 ± 0.33 a | 4.13 ± 0.28 a | 1.25 ± 0.54 a | 2.70 ± 1.07 | |
| OL | 5.09 ± 1.21 b | 4.38 ± 1.29 a | 2.32 ± 0.92 a | 3.62 ± 0.48 a | 1.09 ± 0.24 a | * | |
| Threonine | WS | 2.29 ± 0.13 a | 3.96 ± 1.56 a | 1.93 ± 0.06 a | 2.85 ± 0.87 a | 0.61 ± 0.06 a | 2.45 ± 0.31 |
| GM | 3.83 ± 0.40 ab | 5.65 ± 0.65 a | 3.31 ± 0.26 b | 3.68 ± 0.00 a | 1.17 ± 0.44 a | 2.47 ± 0.93 | |
| OL | 5.23 ± 1.27 b | 4.19 ± 1.08 a | 1.93 ± 0.77 a | 3.22 ± 0.33 a | 1.02 ± 0.20 a | * | |
| Serine | WS | 3.93 ± 0.14 a | 6.17 ± 2.57 a | 3.30 ± 0.24 a | 4.81 ± 1.62 a | 0.91 ± 0.21 a | 4.17 ± 0.31 |
| GM | 6.38 ± 0.50 b | 8.81 ± 0.32 a | 5.77 ± 0.64 b | 5.97 ± 0.37 a | 2.13 ± 0.60 b | 4.19 ± 1.37 | |
| OL | 8.83 ± 1.29 c | 6.95 ± 1.81 a | 3.34 ± 1.29 a | 5.11 ± 0.35 a | 1.84 ± 0.24 b | * | |
| Proline | WS | 1.21 ± 0.06 a | 2.30 ± 1.22 a | 1.04 ± 0.09 a | 1.22 ± 0.57 a | 0.55 ± 0.06 a | 1.37 ± 0.15 |
| GM | 2.19 ± 0.25 a | 3.77 ± 0.47 a | 2.00 ± 0.26 b | 1.67 ± 0.02 a | 0.43 ± 0.20 a | 1.35 ± 0.44 | |
| OL | 4.22 ± 1.59 b | 2.69 ± 1.37 a | 0.88 ± 0.31 a | 1.57 ± 0.23 a | 0.37 ± 0.06 a | * | |
| Asparagine | WS | 1.75 ± 0.10 a | 2.82 ± 1.55 a | 1.18 ± 0.06 a | 1.77 ± 0.65 a | 0.40 ± 0.05 a | 1.63 ± 0.14 |
| GM | 3.02 ± 0.35 b | 4.52 ± 0.68 a | 2.19 ± 0.23 b | 2.26 ± 0.01 a | 0.74 ± 0.24 b | 1.57 ± 0.52 | |
| OL | 4.39 ± 1.03 c | 3.25 ± 1.25 a | 1.22 ± 0.49 a | 1.95 ± 0.20 a | 0.70 ± 0.10 ab | * | |
| Thioproline | WS | 2.48 ± 0.24 a | 2.10 ± 0.38 a | 0.82 ± 0.43 a | 0.59 ± 0.44 a | 0.71 ± 0.27 a | 0.75 ± 0.11 |
| GM | 3.57 ± 0.75 a | 2.93 ± 0.93 a | 0.93 ± 0.30 a | 1.12 ± 0.03 a | 2.69 ± 0.30 b | 1.60 ± 0.80 | |
| OL | 3.31 ± 0.84 a | 3.17 ± 2.14 a | 1.17 ± 0.78 a | 1.02 ± 0.78 a | 1.97 ± 0.46 c | * | |
| Aspartic acid | WS | 2.51 ± 0.09 a | 2.20 ± 0.03 a | 0.98 ± 0.26 a | 0.81 ± 0.57 a | 0.76 ± 0.19 a | 0.81 ± 0.10 |
| GM | 3.51 ± 0.48 b | 2.65 ± 0.80 a | 1.14 ± 0.28 a | 1.06 ± 0.12 a | 2.59 ± 0.40 b | 1.69 ± 0.62 | |
| OL | 3.55 ± 0.62 b | 3.32 ± 2.12 a | 1.22 ± 0.69 a | 1.27 ± 0.85 a | 1.94 ± 0.52 b | * | |
| Methionine | WS | 1.14 ± 0.12 a | 1.86 ± 0.68 a | 0.66 ± 0.21 a | 1.36 ± 0.31 a | 0.09 ± 0.01 a | 1.31 ± 0.23 |
| GM | 1.66 ± 0.21 ab | 2.47 ± 0.14 a | 1.08 ± 0.04 a | 1.62 ± 0.08 a | 0.58 ± 0.19 b | 1.15 ± 0.42 | |
| OL | 2.10 ± 0.57 b | 1.93 ± 0.42 a | 0.90 ± 0.31 a | 1.50 ± 0.23 a | 0.51 ± 0.10 b | * | |
| Hydroxyproline | WS | 0.39 ± 0.04 a | 0.48 ± 0.15 a | 0.22 ± 0.08 a | 0.40 ± 0.09 a | 0.11 ± 0.10 a | 0.34 ± 0.02 |
| GM | 0.60 ± 0.09 b | 0.71 ± 0.12 a | 0.44 ± 0.08 b | 0.44 ± 0.04 a | 0.25 ± 0.08 a | 0.36 ± 0.09 | |
| OL | 0.66 ± 0.11 b | 0.58 ± 0.13 a | 0.25 ± 0.07 a | 0.38 ± 0.05 a | 0.24 ± 0.01 a | * | |
| Glutamic acid | WS | 4.81 ± 0.92 a | 5.68 ± 1.19 a | 3.22 ± 0.48 a | 4.92 ± 0.63 a | 2.07 ± 0.09 a | 4.42 ± 1.21 |
| GM | 6.76 ± 2.08 a | 7.84 ± 2.44 a | 5.29 ± 0.70 b | 5.73 ± 1.35 a | 3.58 ± 0.62 a | 4.59 ± 2.15 | |
| OL | 7.39 ± 2.32 a | 6.76 ± 2.55 a | 3.60 ± 1.47 ab | 4.77 ± 0.81 a | 3.46 ± 1.18 a | * | |
| Phenylalanine | WS | 2.95 ± 0.24 a | 4.30 ± 1.51 a | 1.71 ± 0.20 a | 2.94 ± 0.71 a | 0.97 ± 0.03 a | 3.05 ± 0.54 |
| GM | 4.14 ± 0.55 ab | 6.01 ± 0.11 a | 2.91 ± 0.19 b | 3.57 ± 0.01 a | 1.74 ± 0.41 b | 2.81 ± 1.10 | |
| OL | 5.33 ± 1.16 b | 4.68 ± 1.45 a | 2.27 ± 0.83 ab | 3.31 ± 0.57 a | 1.98 ± 0.47 b | * | |
| Glutamine | WS | 7.80 ± 0.69 a | 10.04 ± 6.97 a | 3.35 ± 0.82 a | 7.19 ± 0.87 a | 0.84 ± 0.61 a | 7.02 ± 0.69 |
| GM | 14.08 ± 2.32 b | 21.69 ± 6.93 a | 8.28 ± 1.61 b | 7.52 ± 1.04 a | 4.22 ± 0.69 b | 6.99 ± 1.17 | |
| OL | 20.03 ± 1.09 c | 13.06 ± 8.46 a | 4.66 ± 2.40 a | 6.96 ± 1.74 a | 6.42 ± 1.23 c | * | |
| Ornithine | WS | 0.65 ± 0.08 a | 0.63 ± 0.24 a | 0.65 ± 0.04 b | 0.36 ± 0.08 a | 0.42 ± 0.01 a | 1.51 ± 0.29 |
| GM | 0.93 ± 0.15 b | 0.90 ± 0.25 a | 0.82 ± 0.09 b | 0.50 ± 0.19 a | 0.76 ± 0.08 b | 1.63 ± 0.67 | |
| OL | 1.10 ± 0.14 b | 0.56 ± 0.28 a | 0.31 ± 0.14 a | 0.43 ± 0.09 a | 0.32 ± 0.12 a | * | |
| Lysine | WS | 2.47 ± 0.27 a | 3.23 ± 0.83 a | 1.83 ± 0.09 a | 2.77 ± 0.32 a | 0.53 ± 0.03 a | 2.79 ± 0.49 |
| GM | 3.33 ± 0.78 a | 4.29 ± 0.67 a | 2.77 ± 0.41 a | 3.42 ± 0.45 a | 1.36 ± 0.44 b | 2.61 ± 1.02 | |
| OL | 4.20 ± 1.30 a | 3.59 ± 1.10 a | 2.21 ± 1.02 a | 2.96 ± 0.51 a | 1.44 ± 0.30 b | * | |
| Histidine | WS | 0.88 ± 0.17 a | 1.67 ± 1.24 a | 0.52 ± 0.07 a | 0.88 ± 0.50 a | 0.10 ± 0.05 a | 0.98 ± 0.21 |
| GM | 1.63 ± 0.27 ab | 2.59 ± 0.57 a | 1.14 ± 0.26 b | 1.14 ± 0.33 a | 0.32 ± 0.21 a | 0.86 ± 0.21 | |
| OL | 2.46 ± 1.03 b | 1.66 ± 0.66 a | 0.63 ± 0.20 a | 1.04 ± 0.30 a | 0.33 ± 0.06 a | * | |
| Tyrosine | WS | 2.66 ± 0.08 a | 4.25 ± 1.73 a | 1.54 ± 0.12 a | 2.59 ± 0.62 a | 0.95 ± 0.13 a | 3.05 ± 0.46 |
| GM | 3.99 ± 0.44 ab | 5.93 ± 0.33 a | 2.67 ± 0.24 b | 3.18 ± 0.23 a | 1.70 ± 0.42 b | 2.81 ± 1.04 | |
| OL | 4.96 ± 1.08 b | 4.34 ± 1.42 a | 1.95 ± 0.70 ab | 3.11 ± 0.54 a | 1.40 ± 0.23 ab | * | |
| Tryptophan | WS | 1.07 ± 0.12 a | 0.81 ± 0.71 a | 0.59 ± 0.15 a | 1.22 ± 0.68 a | 0.18 ± 0.04 a | 1.16 ± 0.34 |
| GM | 1.80 ± 0.33 a | 1.76 ± 0.00 a | 1.14 ± 0.09 b | 0.55 ± 0.78 a | 0.43 ± 0.14 b | 1.01 ± 0.33 | |
| OL | 1.70 ± 0.00 a | 1.44 ± 0.22 a | 0.76 ± 0.28 a | 0.76 ± 0.66 a | 0.42 ± 0.09 b | * | |
| γ-Aminobutyric acid (GABA) | WS | 0.21 ± 0.01 a | 0.31 ± 0.20 a | 0.08 ± 0.05 a | 0.18 ± 0.13 a | 0.09 ± 0.03 a | 0.56 ± 0.26 |
| GM | 0.43 ± 0.21 a | 0.70 ± 0.41 a | 0.38 ± 0.12 b | 0.18 ± 0.03 a | 0.18 ± 0.05 a | 0.86 ± 0.37 | |
| OL | 0.46 ± 0.21 a | 0.35 ± 0.21 a | 0.06 ± 0.02 a | 0.23 ± 0.08 a | 0.36 ± 0.12 b | * | |
| Total free amino acids | WS | 63.10 ± 4.22 a | 87.84 ± 29.53 a | 62.19 ± 13.43 a | 39.91 ± 1.58 a | 17.37 ± 1.48 a | 61.18 ± 9.42 |
| GM | 97.03 ± 14.44 b | 130.12 ± 17.16 a | 75.44 ± 4.42 a | 69.86 ± 5.81 b | 37.42 ± 7.09 b | 62.16 ± 22.76 | |
| OL | 116.98 ± 19.46 b | 96.05 ± 31.79 a | 67.89 ± 8.88 a | 46.03 ± 19.17 a | 36.30 ± 7.68 b | * | |
| Crude protein | WS | 164.07 ± 1.65 a | 177.36 ± 3.55 a | 154.63 ± 9.25 a | 144.70 ± 2.90 b | 121.10 ± 1.20 a | 175.20 ± 5.26 |
| GM | 221.18 ± 11.05 b | 284.77 ± 2.85 b | 187.83 ± 1.85 b | 179.60 ± 5.40 c | 175.23 ± 8.73 b | 172.24 ± 10.33 | |
| OL | 293.20 ± 17.60 c | 299.20 ± 9.00 c | 167.23 ± 8.35 a | 130.77 ± 7.84 a | 130.50 ± 5.20 a | * | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tagkouli, D.; Kaliora, A.; Bekiaris, G.; Koutrotsios, G.; Christea, M.; Zervakis, G.I.; Kalogeropoulos, N. Free Amino Acids in Three Pleurotus Species Cultivated on Agricultural and Agro-Industrial By-Products. Molecules 2020, 25, 4015. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25174015
Tagkouli D, Kaliora A, Bekiaris G, Koutrotsios G, Christea M, Zervakis GI, Kalogeropoulos N. Free Amino Acids in Three Pleurotus Species Cultivated on Agricultural and Agro-Industrial By-Products. Molecules. 2020; 25(17):4015. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25174015
Chicago/Turabian StyleTagkouli, Dimitra, Andriana Kaliora, Georgios Bekiaris, Georgios Koutrotsios, Margarita Christea, Georgios I. Zervakis, and Nick Kalogeropoulos. 2020. "Free Amino Acids in Three Pleurotus Species Cultivated on Agricultural and Agro-Industrial By-Products" Molecules 25, no. 17: 4015. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25174015