
Bioguided Fractionation of Local Plants against Matrix Metalloproteinase9 and Its Cytotoxicity against Breast Cancer Cell Models: In Silico and In Vitro Study

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Results
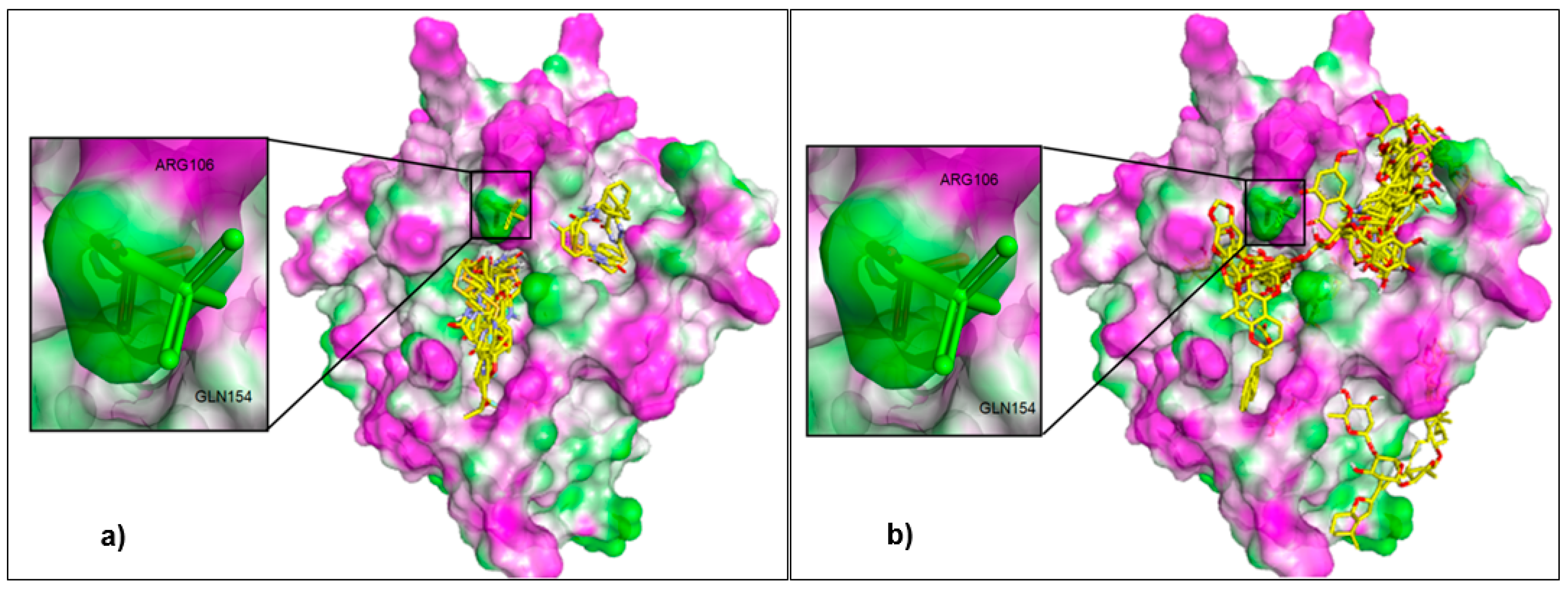
2.1. Control Docking and External Validation
2.2. Virtual Screening
2.3. Plants Extraction
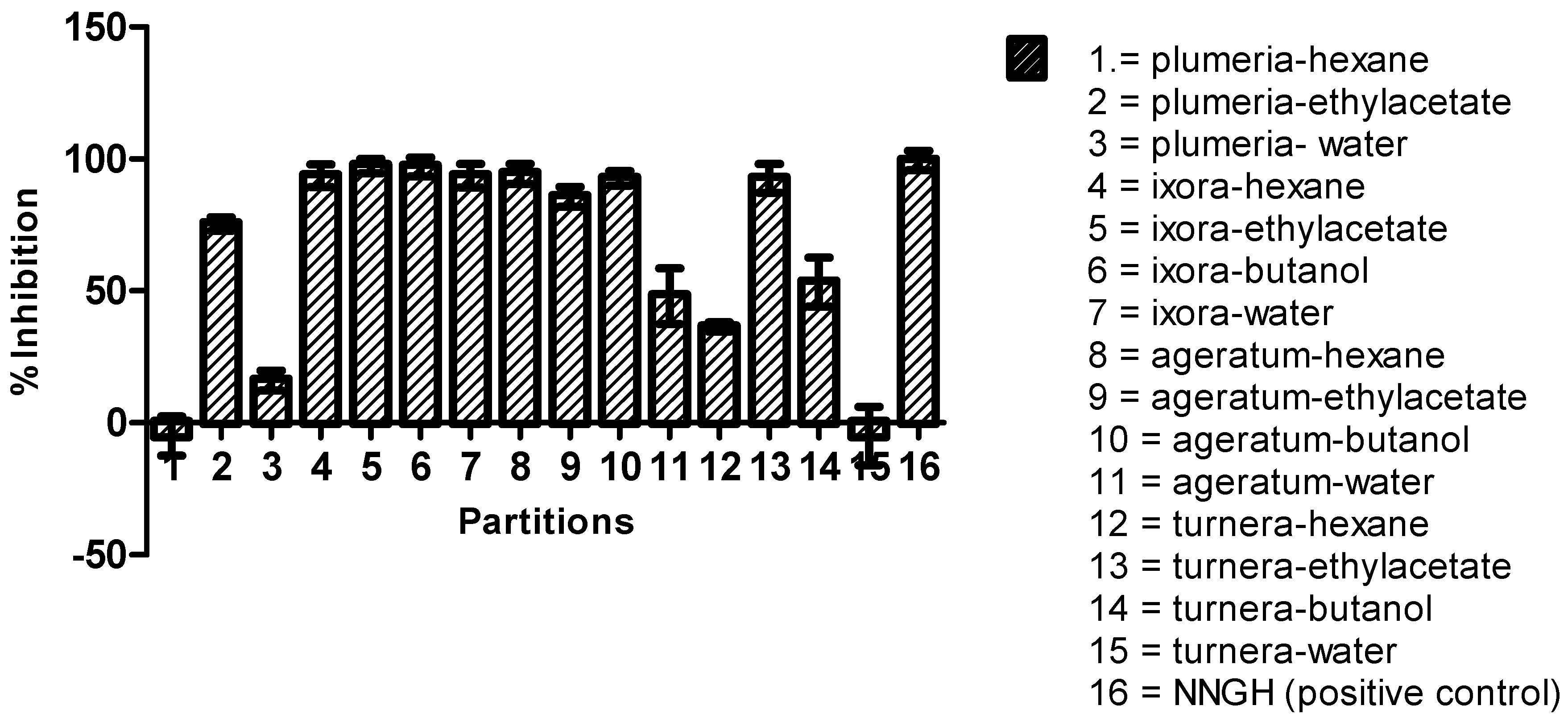
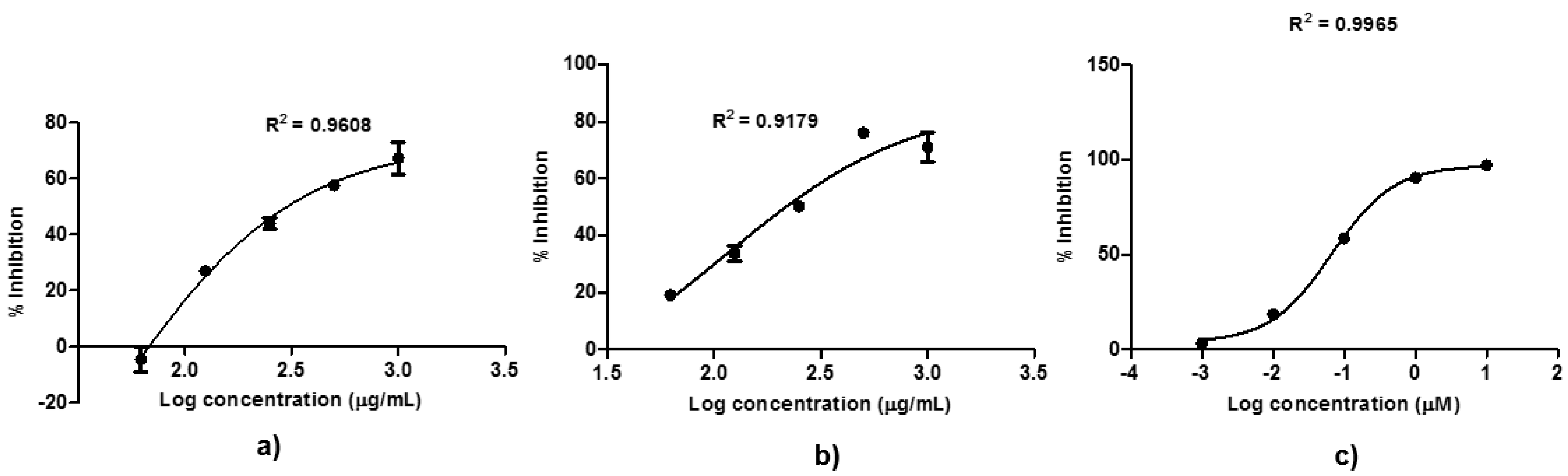
2.4. Bioguided Fractionation against MMP9
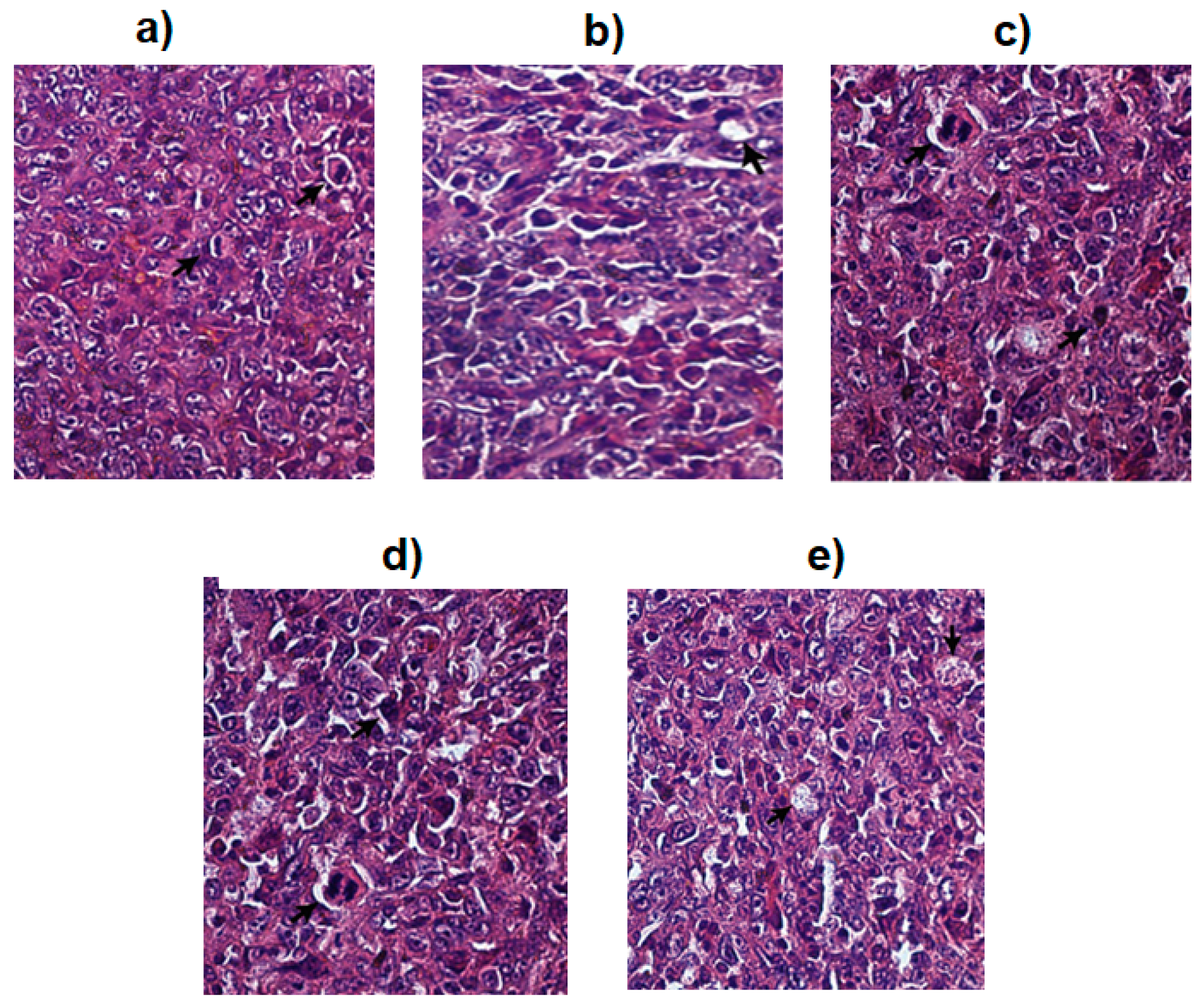
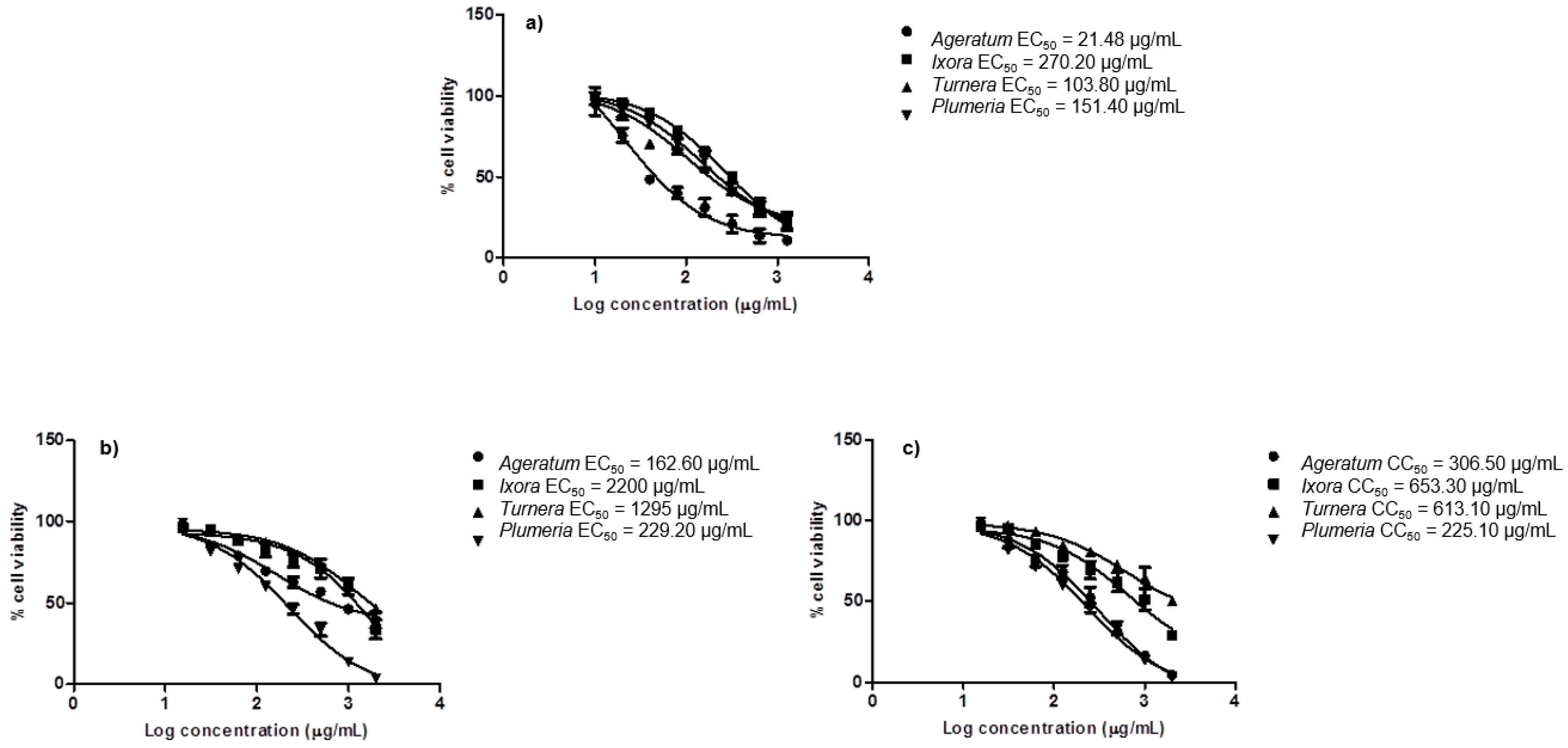
2.5. In Vitro Cytotoxicity Assay
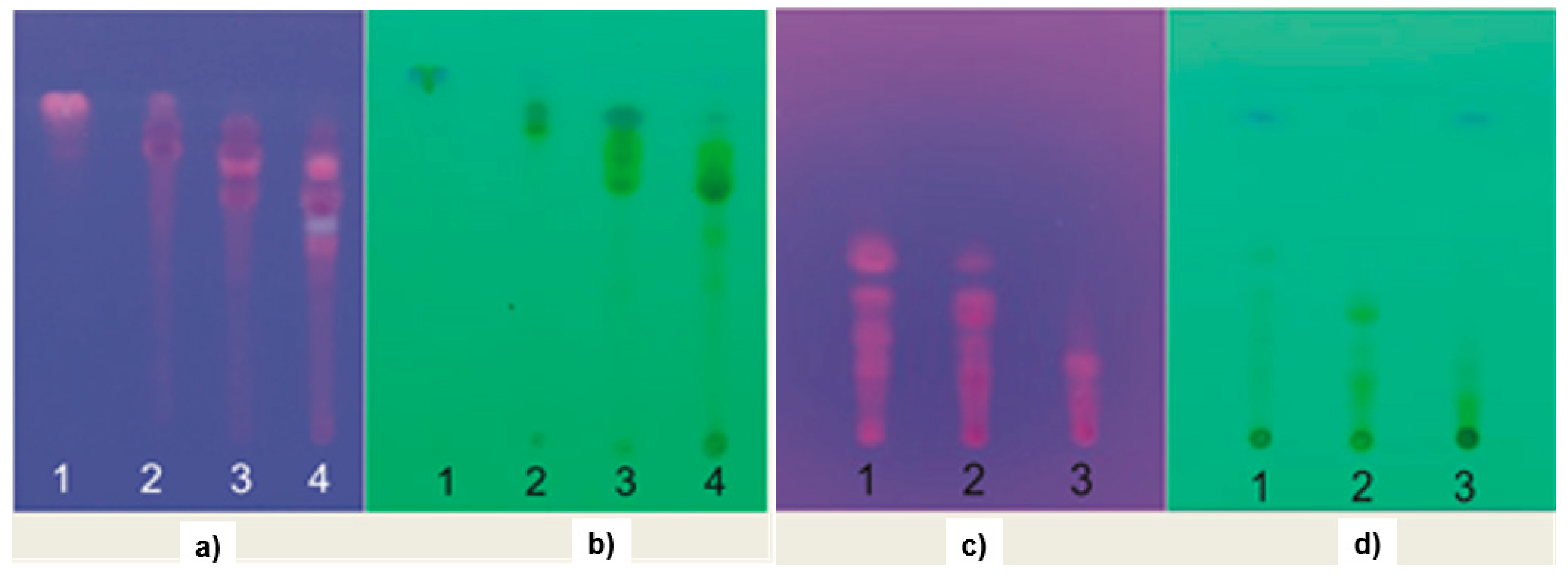
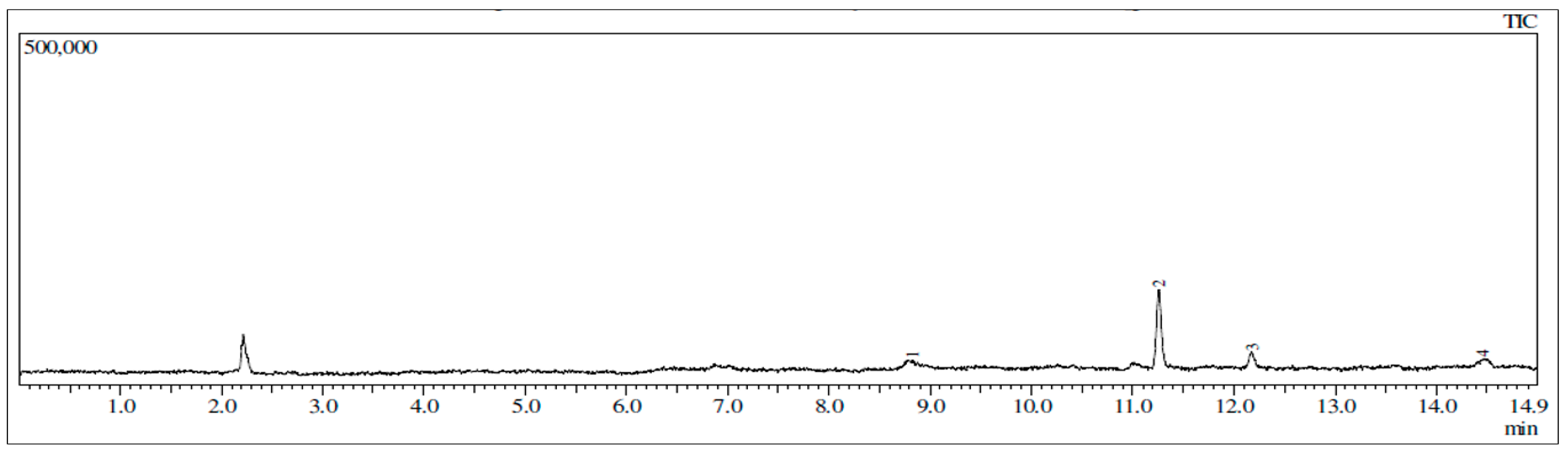
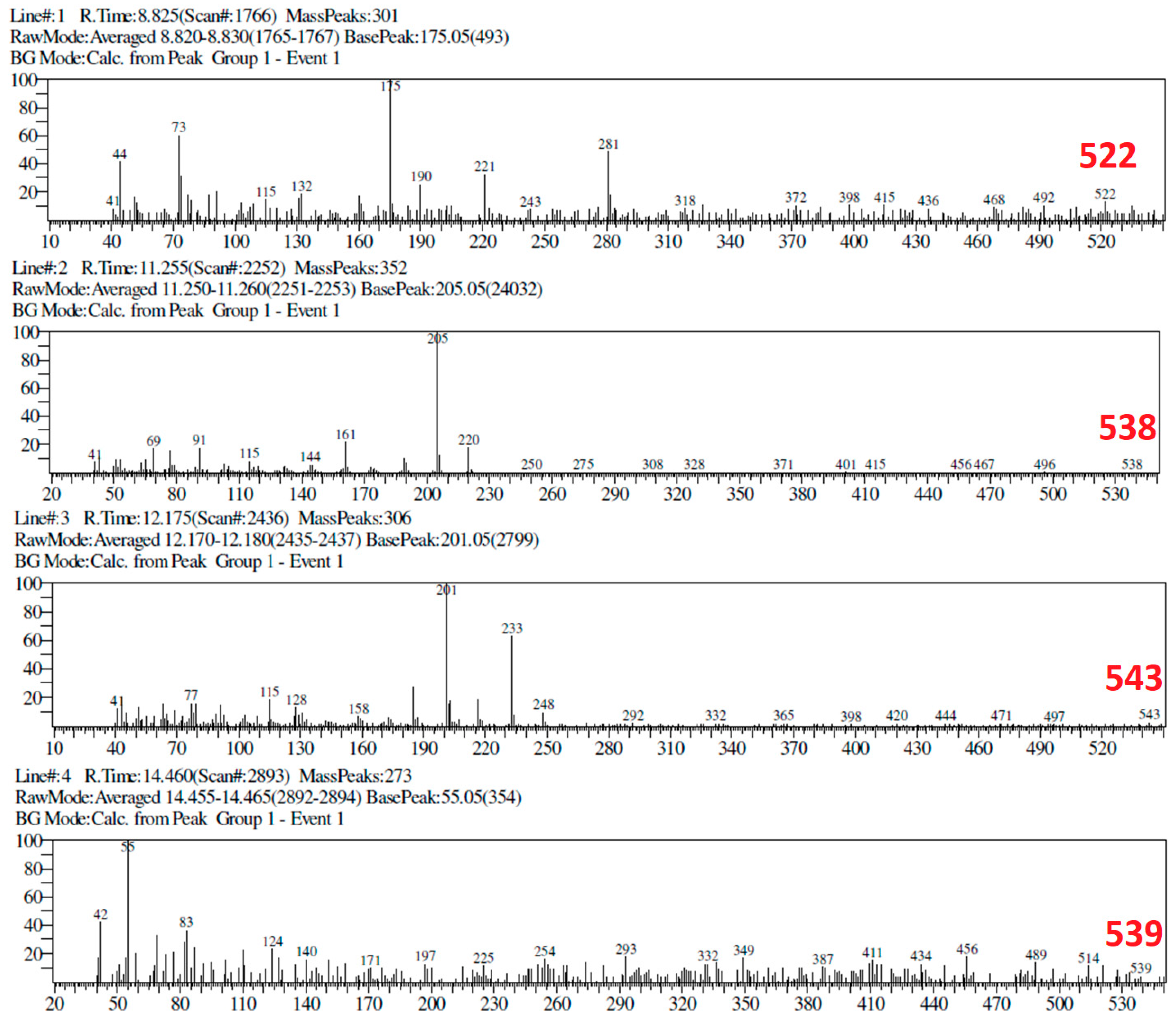
2.6. TLC and GC-MS Profiles
3. Discussion
4. Materials and Methods
4.1. Software and Hardware
4.2. Chemicals
4.3. Control Docking and External Validation
4.4. In Silico Screening
4.5. Plant Collections and Extractions
4.6. Liquid-Liquid Partitions and Fractionation
4.7. In Vitro MMP9 Inhibition Assay
4.8. In Vitro Cytotoxicity Assay
4.9. Gas Chromatography-Mass Spectroscopy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Kim, Y.H. Cancer prevention from the perspective of global cancer burden patterns. J. Thorac. Dis. 2017, 9, 448–451. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Koo, J.S.; Kim, M.S.; Park, H.S.; Lee, J.S.; Lee, J.S.; Kim, S.I.; Park, B.W. Characteristics and outcomes according to molecular subtypes of breast cancer as classified by a panel of four biomarkers using immunohistochemistry. Breast 2012, 21, 50–57. [Google Scholar] [CrossRef]
- Serrano, D.; Lazzeroni, M.; Bonanni, B. Cancer chemoprevention: Much has been done, but there is still much to do. State of the art and possible new approaches. Mol. Oncol. 2015, 9, 1008–1017. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Feng, Y.; Lin, S.; Chen, J.; Lin, H.; Liang, X.; Zheng, H.; Cai, X. Mechanisms involved in breast cancer liver metastasis. J. Transl. Med. 2015, 13, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Rollando, R.; Hariono, M. Antimicrobial, Antioxidant and T47D Cytotoxic Activities of Trichaptum sp., A Fungal Endophyte from Phyllantus niruri Linn.: In vitro and in silico Studies. Asian J. Cell Biol. 2016, 12, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Mehner, C.; Hockla, A.; Miller, E.; Ran, S.; Radisky, D.C.; Radisky, E.S. Tumor cell-produced matrix metalloproteinase 9 (MMP-9) drives malignant progression and metastasis of basal-like triple negative breast cancer. Oncotarget 2014, 5, 2736–2749. [Google Scholar] [CrossRef] [Green Version]
- Lebert, J.M.; Lester, R.; Powell, E.; Seal, M.; McCarthy, J. Advances in the systemic treatment of triple-negative breast cancer. Curr. Oncol. 2018, 25, S142–S150. [Google Scholar] [CrossRef] [Green Version]
- Hariono, M.; Yuliani, S.H.; Istyastono, E.P.; Riswanto, F.D.O.; Adhipandito, C.F. Matrix metalloproteinase 9 (MMP9) in wound healing of diabetic foot ulcer: Molecular target and structure-based drug design. Wound Med. 2018, 22, 1–13. [Google Scholar] [CrossRef]
- Tallant, C.; Marrero, A.; Gomis-Rüth, F.X. Matrix metalloproteinases: Fold and function of their catalytic domains. Biochim. Biophys. Acta Mol. Cell Res. 2010, 1803, 20–28. [Google Scholar] [CrossRef]
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousef, E.M.; Tahir, M.R.; St-Pierre, Y.; Gaboury, L.A. MMP-9 expression varies according to molecular subtypes of breast cancer. BMC Cancer 2014, 14, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Coussens, L.M.; Fingleton, B.; Matrisian, L.M. Matrix metalloproteinase inhibitors and cancer: Trials and tribulations. Science 2002, 295, 2387–2392. [Google Scholar] [CrossRef] [PubMed]
- Scannevin, R.H.; Alexander, R.; Haarlander, T.M.; Burke, S.L.; Singer, M.; Huo, C.; Zhang, Y.M.; Maguire, D.; Spurlino, J.; Deckman, I.; et al. Discovery of a highly selective chemical inhibitor of matrix metalloproteinase-9 (MMP-9) that allosterically inhibits zymogen activation. J. Biol. Chem. 2017, 292, 17963–17974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalva, S.; Azhagiya Singam, E.R.; Rajapandian, V.; Saleena, L.M.; Subramanian, V. Discovery of potent inhibitor for matrix metalloproteinase-9 by pharmacophore based modeling and dynamics simulation studies. J. Mol. Graph. Model. 2014, 49, 25–37. [Google Scholar] [CrossRef]
- Pirard, B. Insight into the structural determinants for selective inhibition of matrix metalloproteinases. Drug Discov. Today 2007, 12, 640–646. [Google Scholar] [CrossRef]
- Adhipandito, C.F.; Ludji, D.P.K.S.; Aprilianto, E.; Jenie, R.I.; Al-Najjar, B.; Hariono, M. Matrix metalloproteinase9 as the protein target in anti-breast cancer drug discovery: An approach by targeting hemopexin domain. Future J. Pharm. Sci. 2019, 5, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Glasheen, B.M.; Kabra, A.T.; Page-McCaw, A. Distinct functions for the catalytic and hemopexin domains of a Drosophila matrix metalloproteinase. Proc. Natl. Acad. Sci. USA 2009, 106, 2659–2664. [Google Scholar] [CrossRef] [Green Version]
- Mazurek, S.G.N.; Li, J.; Nabozny, G.H.; Reinhart, G.A.; Muthukumarana, A.C.; Harrison, P.C.; Fryer, R.M. Functional biomarkers of musculoskeletal syndrome (MSS) for early in vivo screening of selective MMP-13 inhibitors. J. Pharmacol. Toxicol. Methods 2011, 64, 89–96. [Google Scholar] [CrossRef]
- Overall, C.M.; Kleifeld, O. Towards third generation matrix metalloproteinase inhibitors for cancer therapy. Br. J. Cancer 2006, 94, 941–946. [Google Scholar] [CrossRef] [Green Version]
- Sela-Passwell, N.; Rosenblum, G.; Shoham, T.; Sagi, I. Structural and functional bases for allosteric control of MMP activities: Can it pave the path for selective inhibition? Biochim. Biophys. Acta Mol. Cell Res. 2010, 1803, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Ma, Q.; Ye, L.; Piao, G. The traditional medicine and modern medicine from natural products. Molecules 2016, 21, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palethorpe, H.M.; Smith, E.; Tomita, Y.; Nakhjavani, M.; Yool, A.J.; Price, T.J.; Young, J.P.; Townsend, A.R.; Hardingham, J.E. Bacopasides I and II act in synergy to inhibit the growth, migration and invasion of breast cancer cell lines. Molecules 2019, 24, 3539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jabeen, A.; Sharma, A.; Gupta, I.; Kheraldine, H.; Vranic, S.; Al Moustafa, A.E.; Al Farsi, H.F. Elaeagnus angustifolia plant extract inhibits epithelial-mesenchymal transition and induces apoptosis via HER2 inactivation and JNK pathway in HER2-positive breast cancer cells. Molecules 2020, 25, 4240. [Google Scholar] [CrossRef]
- Paramanantham, A.; Kim, M.J.; Jung, E.J.; Kim, H.J.; Chang, S.H.; Jung, J.M.; Hong, S.C.; Shin, S.C.; Kim, G.S.; Lee, W.S. Anthocyanins isolated from vitis coignetiae pulliat enhances cisplatin sensitivity in MCF-7 human breast cancer cells through inhibition of Akt and NF-κB activation. Molecules 2020, 25, 3623. [Google Scholar] [CrossRef] [PubMed]
- Mori, K. Bioactive natural products and chirality. Chirality 2011, 23, 449–462. [Google Scholar] [CrossRef]
- Hevener, K.E.; Zhao, W.; Ball, D.M.; Babaoglu, K.; Qi, J.; White, S.W.; Lee, R.E. Validation of molecular docking programs for virtual screening against dihydropteroate synthase. J. Chem. Inf. Model. 2009, 49, 444–460. [Google Scholar] [CrossRef]
- Alford, V.M.; Kamath, A.; Ren, X.; Kumar, K.; Gan, Q.; Awwa, M.; Tong, M.; Seeliger, M.A.; Cao, J.; Ojima, I.; et al. Targeting the Hemopexin-like Domain of Latent Matrix Metalloproteinase-9 (proMMP-9) with a Small Molecule Inhibitor Prevents the Formation of Focal Adhesion Junctions. ACS Chem. Biol. 2017, 12, 2788–2803. [Google Scholar] [CrossRef]
- Dufour, A.; Sampson, N.S.; Li, J.; Kuscu, C.; Rizzo, R.C.; DeLeon, J.L.; Zhi, J.; Jaber, N.; Liu, E.; Zucker, S.; et al. Small-Molecule Anticancer Compounds Selectively Target the Hemopexin Domain of Matrix Metalloproteinase-9. Cancer Res. 2011, 71, 4977–4988. [Google Scholar] [CrossRef] [Green Version]
- Varbanov, H.P.; Kuttler, F.; Banfi, D.; Turcatti, G.; Dyson, P.J. Repositioning approved drugs for the treatment of problematic cancers using a screening approach. PLoS ONE 2017, 12, e0171052. [Google Scholar] [CrossRef] [Green Version]
- Braga, R.C.; Andrade, C.H. Assessing the Performance of 3D Pharmacophore Models in Virtual Screening: How Good are They? Curr. Top. Med. Chem. 2013, 13, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Ikram, N.K.K.; Durrant, J.D.; Muchtaridi, M.; Zalaludin, A.S.; Purwitasari, N.; Mohamed, N.; Rahim, A.S.A.; Lam, C.K.; Normi, Y.M.; Rahman, N.A.; et al. A Virtual Screening Approach for Identifying Plants with Anti H5N1 Neuraminidase Activity. J. Chem. Inf. Model. 2015, 55, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Hariono, M.; Choi, S.B.; Roslim, R.F.; Nawi, M.S.; Tan, M.L.; Kamarulzaman, E.E.; Mohamed, N.; Yusof, R.; Othman, S.; Abd Rahman, N.; et al. Thioguanine-based DENV-2 NS2B/NS3 protease inhibitors: Virtual screening, synthesis, biological evaluation and molecular modelling. PLoS ONE 2019, 14, e0210869. [Google Scholar] [CrossRef] [PubMed]
- Sakkiah, S.; Thangapandian, S.; John, S.; Kwon, Y.J.; Lee, K.W. 3D QSAR pharmacophore based virtual screening and molecular docking for identification of potential HSP90 inhibitors. Eur. J. Med. Chem. 2010, 45, 2132–2140. [Google Scholar] [CrossRef] [PubMed]
- Rowsell, S.; Hawtin, P.; Minshull, C.A.; Jepson, H.; Brockbank, S.M.V.; Barratt, D.G.; Slater, A.M.; McPheat, W.L.; Waterson, D.; Henney, A.M.; et al. Crystal structure of human MMP9 in complex with a reverse hydroxamate inhibitor. J. Mol. Biol. 2002, 319, 173–181. [Google Scholar] [CrossRef]
- González, M.L.; Joray, M.B.; Laiolo, J.; Crespo, M.I.; Palacios, S.M.; Ruiz, G.M.; Carpinella, M.C. Cytotoxic Activity of Extracts from Plants of Central Argentina on Sensitive and Multidrug-Resistant Leukemia Cells: Isolation of an Active Principle from Gaillardia megapotamica. Evid.-Based Complement. Altern. Med. 2018, 2018, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adebayo, A.H.; Ji, C.-J.; Zhang, Y.-M.; He, W.-J.; Zeng, G.-Z.; Han, H.-J.; Xu, J.-J.; Akindahunsi, A.A.; Tan, N.-H. A New Chromene Isolated from Ageratum conyzoides. Nat. Prod. Commun. 2011, 6. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, A.; Rijhwani, S. A comprehensive review on phytochemistry of Ageratum conyzoides Linn. (Goat weed). Int. J. Eng. Technol. Manag. Appl. Sci. 2015, 3, 348–358. [Google Scholar]
- Kamboj, A.; Saluja, A.K. Isolation of stigmasterol and β-sitosterol from petroleum ether extract of aerial parts of Ageratum conyzoides (Asteraceae). Int. J. Pharm. Pharm. Sci. 2011, 3, 94–96. [Google Scholar]
- Duke, S.; B Powles, S. Glyphosate: A once-in-a-century herbicide. Pest. Manag. Sci. 2008, 63, 1100–1106. [Google Scholar] [CrossRef]
- Singh, S.B.; Devi, W.R.; Marina, A.; Devi, W.I.; Swapana, N.; Singh, C.B. Ethnobotany, phytochemistry and pharmacology of Ageratum conyzoides Linn (Asteraceae). J. Med. Plants Res. 2013, 7, 371–385. [Google Scholar] [CrossRef]
- Lee, C.-L.; Liao, Y.-C.; Hwang, T.-L.; Wu, C.-C.; Chang, F.-R.; Wu, Y.-C. Ixorapeptide I and ixorapeptide II, bioactive peptides isolated from Ixora coccinea. Bioorg. Med. Chem. Lett. 2010, 20, 7354–7357. [Google Scholar] [CrossRef] [PubMed]
- Baskar, G.; Devi, A.G.; Nishanthini, T.; Aiswarya, R. Anticancer activity of Ixora coccinea linn flower extracts against Human Breast Adenocarcinoma cells. Int. J. Mod. Sci. Technol. 2017, 2, 288–291. [Google Scholar]
- Urolagin, D.K.; Jayakumari, S. In vitro Anti-Cancer Study of Vitis viniferae, Ixora coccinea and Piper longum Extract on Human Breast Carcinoma Cells. Res. J. Pharm. Technol. 2018, 11, 5345. [Google Scholar] [CrossRef]
- Prabhu, V.; Devi, K.V.; Priya, M.K. Gc-Ms Analysis of Bioactive Compounds Present in the Petroleum Ether, Chloroform and Methanol Extract of Ixora Coccinea’ S Flower and in-Vitro Cytotoxic Activity of. Int. J. Res. Anal. Rev. 2018, 5, 801–807. [Google Scholar]
- Ali, A.; Siddiqui, F.; Fazil, P.; Farooq, A.D.; Siddiqui, A.J.; Musharraf, S.G. Cytotoxic activity of extracts of Ixora species and their GC-MS analysis. J. Chem. Soc. Pak. 2018, 40, 941. [Google Scholar]
- Adebayo, A.H.; Tan, N.H.; Akindahunsi, A.A.; Zeng, G.Z.; Zhang, Y.M. Anticancer and antiradical scavenging activity of Ageratum conyzoides L. (Asteraceae). Pharmacogn. Mag. 2010, 6, 62–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acheampong, F.; Larbie, C.; Appiah-Opong, R.; Arthur, F.; Tuffour, I. In vitro Antioxidant and Anticancer Properties of Hydroethanolic Extracts and Fractions of Ageratum conyzoides. Eur. J. Med. Plants 2015, 7, 205–214. [Google Scholar] [CrossRef]
- Cha, H.; Kopetzki, E.; Huber, R.; Lanzendörfer, M.; Brandstetter, H. Structural Basis of the Adaptive Molecular Recognition by MMP9. J. Mol. Biol. 2002, 320, 1065–1079. [Google Scholar] [CrossRef]
- Zeng, X.; Zhang, P.; He, W.; Qin, C.; Chen, S.; Tao, L.; Wang, Y.; Tan, Y.; Gao, D.; Wang, B.; et al. NPASS: Natural product activity and species source database for natural product research, discovery and tool development. Nucleic Acids Res. 2018, 46, D1217–D1222. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, H.K.A.; Backer, C.A.; van den Brink, R.C.B.J. Flora of Java. Kew Bull. 1967, 21, 160. [Google Scholar] [CrossRef]
- Silva, V.L.; Ferreira, D.; Nobrega, F.L.; Martins, I.M.; Kluskens, L.D.; Rodrigues, L.R. Selection of Novel Peptides Homing the 4T1 CELL Line: Exploring Alternative Targets for Triple Negative Breast Cancer. PLoS ONE 2016, 11, e0161290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hariono, M.; Nuwarda, R.F.; Yusuf, M.; Rollando, R.; Jenie, R.I.; Al-Najjar, B.; Julianus, J.; Putra, K.C.; Nugroho, E.S.; Wisnumurti, Y.K.; et al. Arylamide as Potential Selective Inhibitor for Matrix Metalloproteinase 9 (MMP9): Design, Synthesis, Biological Evaluation, and Molecular Modeling. J. Chem. Inf. Model. 2020, 60, 349–359. [Google Scholar] [CrossRef]
- Yu, S.; Kim, T.; Yoo, K.H.; Kang, K. The T47D cell line is an ideal experimental model to elucidate the progesterone-specific effects of a luminal A subtype of breast cancer. Biochem. Biophys. Res. Commun. 2017, 486, 752–758. [Google Scholar] [CrossRef]
- Sakuma, C.; Sekizuka, T.; Kuroda, M.; Kasai, F.; Saito, K.; Ikeda, M.; Yamaji, T.; Osada, N.; Hanada, K. Novel endogenous simian retroviral integrations in Vero cells: Implications for quality control of a human vaccine cell substrate. Sci. Rep. 2018, 8, 644. [Google Scholar] [CrossRef]
- Li, K.; Yang, X.; Hu, X.; Han, C.; Lei, Z.; Zhang, Z. In vitro antioxidant, immunomodulatory and anticancer activities of two fractions of aqueous extract from Helicteres angustifolia L. root. J. Taiwan Inst. Chem. Eng. 2016, 61, 75–82. [Google Scholar] [CrossRef]
Sample Availability: Samples of the crude extracts of Cordyline fruticosa leaves, Amaranthus spinosus aerial part, Turnera diffusa leaves, Hibiscus rosa-sinensis leaves, Ageratum conyzoides aerial part, Ixora coccinea leaves, Plumeria alba leaves, and Melaleuca leucadendron leaves are available from the authors. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PubChem ID | Ligand | Binding Affinity (kcal/mol) | Plant |
|---|---|---|---|
| 441207 | digitoxin | −11.2 | Cordyline fruticosa |
| 2724385 | digoxin | −10.4 | Cordyline fruticosa |
| 91540 | gitoxin | −10.1 | Cordyline fruticosa |
| 10328286 | thonningianin A | −9.1 | Piper betle |
| 6325284 | amaranthin | −8.7 | Amaranthus spinosus |
| 4483248 | ophiopogonin C | −8.6 | Parkia javanica |
| 92825 | gypsogenin | −8.6 | Turnera diffusa |
| 101277 | pachyrrizin | −8.5 | Pachyrhizus erosus |
| 5484010 | sequoiaflavone | −8.4 | Elateriospermum tapos |
| 11467 | γ-terpineol | −8.4 | Melaleuca leucadendron |
| 15411208 | 2,3-dihydrowithaferin A | −8.4 | Withania somnifera |
| 72307 | sesamin | −8.3 | Ageratum conyzoides |
| 100257 | Thalrugosin | −8.3 | Cyclea barbata |
| 15484640 | ixorapeptide I | −8.3 | Ixora coccinea |
| 23265223 | quercetin−3-O-(3′6″-O-di-p-coumaroyl)-glucoside | −8.2 | Hibiscus rosa-sinensis |
| 131750919 | Cryptochrome | −8.2 | Averrhoa carambola |
| 92097 | Taraxerol | −8.2 | Plumeria alba |
| 99620 | Homoaromoline | −8.2 | Arcangelisia flava |
| 162807 | (−)-glyceollin I | −8.1 | Glycine soja |
| 490367 | 19-α-hydroxyasiatic acid | −8.1 | Cordyline fruticosa |
| Crude Extract | Yields (%) | % Inhibition ± SEM | IC50 ± SEM (µg/mL) | R2 |
|---|---|---|---|---|
| Cordyline fruticosa leaves | 43 | −6 ± 12 | ND | ND |
| Amaranthus spinosus aerial part | 62 | 81 ± 4 | 783 ± 40 | 0.7501 |
| Turnera diffusa leaves | 65 | 55 ± 4 | 495 ± 20 | 0.9898 |
| Ageratum conyzoides aerial part | 65 | 75 ± 3 | 64 ± 14 | 0.8071 |
| Ixora coccinea leaves | 81 | 86 ± 1 | 82 ± 3 | 0.9879 |
| Hibiscus rosa-sinensis leaves | 72 | 55 ± 2 | 822 ± 20 | 0.9723 |
| Plumeria alba leaves | 77 | 85 ± 9 | 24 ± 8 | 0.8268 |
| Melaleuca leucadendron leaves | 41 | −15 ± 14 | ND | ND |
| No | Sample | IC50 ± SEM (µg/mL) | SI | |||
|---|---|---|---|---|---|---|
| 4T1 (R2) | T47D (R2) | Vero (R2) | 4T1 | T47D | ||
| 1 | Turnera diffusa leaves | 104 ± 5 (0.9192) | 1295 ± 9 (0.9680) | 613 ± 6 (0.9298) | 5.90 | 0.47 |
| 2 | Ageratum conyzoides aerial part | 22 ± 14 (0.9388) | 163 ± 3 (0.9387) | 307 ± 5 (0.9708) | 14.27 | 1.88 |
| 3 | Ixora coccinea leaves | 270 ± 3 (0.9887) | 2200 ± 42 (0.8831) | 653 ± 10 (0.9058) | 2.42 | 0.29 |
| 4 | Plumeria alba leaves | 151 ± 4 (0.9566) | 229 ± 4 (0.9741) | 225 ± 4 (0.9741) | 1.49 | 0.98 |
| 5 | Doxorubicin | 37 ± 6 (0.9483) | 9 ± 4 (0.8740) | 211 ± 7 (0.9549) | 5.71 | 24.59 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hariono, M.; Rollando, R.; Karamoy, J.; Hariyono, P.; Atmono, M.; Djohan, M.; Wiwy, W.; Nuwarda, R.; Kurniawan, C.; Salin, N.; et al. Bioguided Fractionation of Local Plants against Matrix Metalloproteinase9 and Its Cytotoxicity against Breast Cancer Cell Models: In Silico and In Vitro Study. Molecules 2020, 25, 4691. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25204691
Hariono M, Rollando R, Karamoy J, Hariyono P, Atmono M, Djohan M, Wiwy W, Nuwarda R, Kurniawan C, Salin N, et al. Bioguided Fractionation of Local Plants against Matrix Metalloproteinase9 and Its Cytotoxicity against Breast Cancer Cell Models: In Silico and In Vitro Study. Molecules. 2020; 25(20):4691. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25204691
Chicago/Turabian StyleHariono, Maywan, Rollando Rollando, Jasson Karamoy, Pandu Hariyono, M. Atmono, Maria Djohan, Wiwy Wiwy, Rina Nuwarda, Christopher Kurniawan, Nurul Salin, and et al. 2020. "Bioguided Fractionation of Local Plants against Matrix Metalloproteinase9 and Its Cytotoxicity against Breast Cancer Cell Models: In Silico and In Vitro Study" Molecules 25, no. 20: 4691. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25204691