
Plant Derived Natural Products against Pseudomonas aeruginosa and Staphylococcus aureus: Antibiofilm Activity and Molecular Mechanisms

 , , and
, , and 
Abstract
:1. Introduction
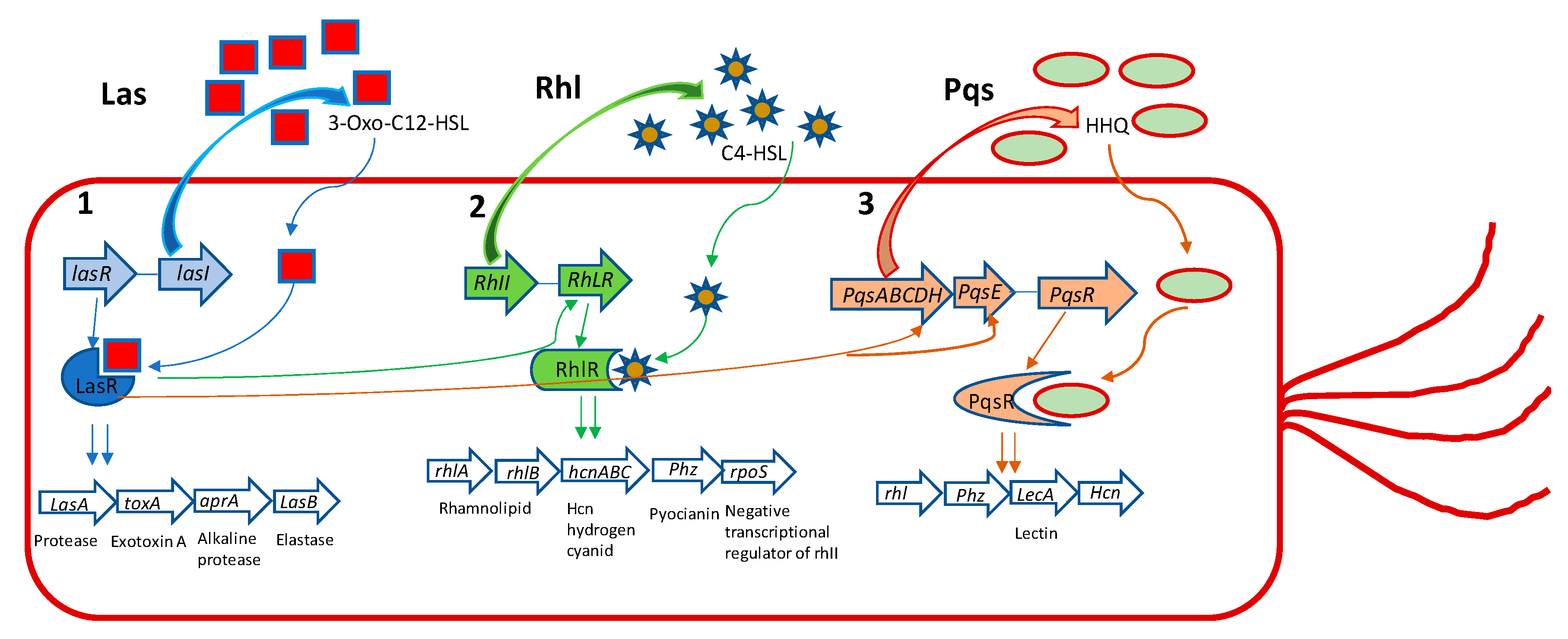
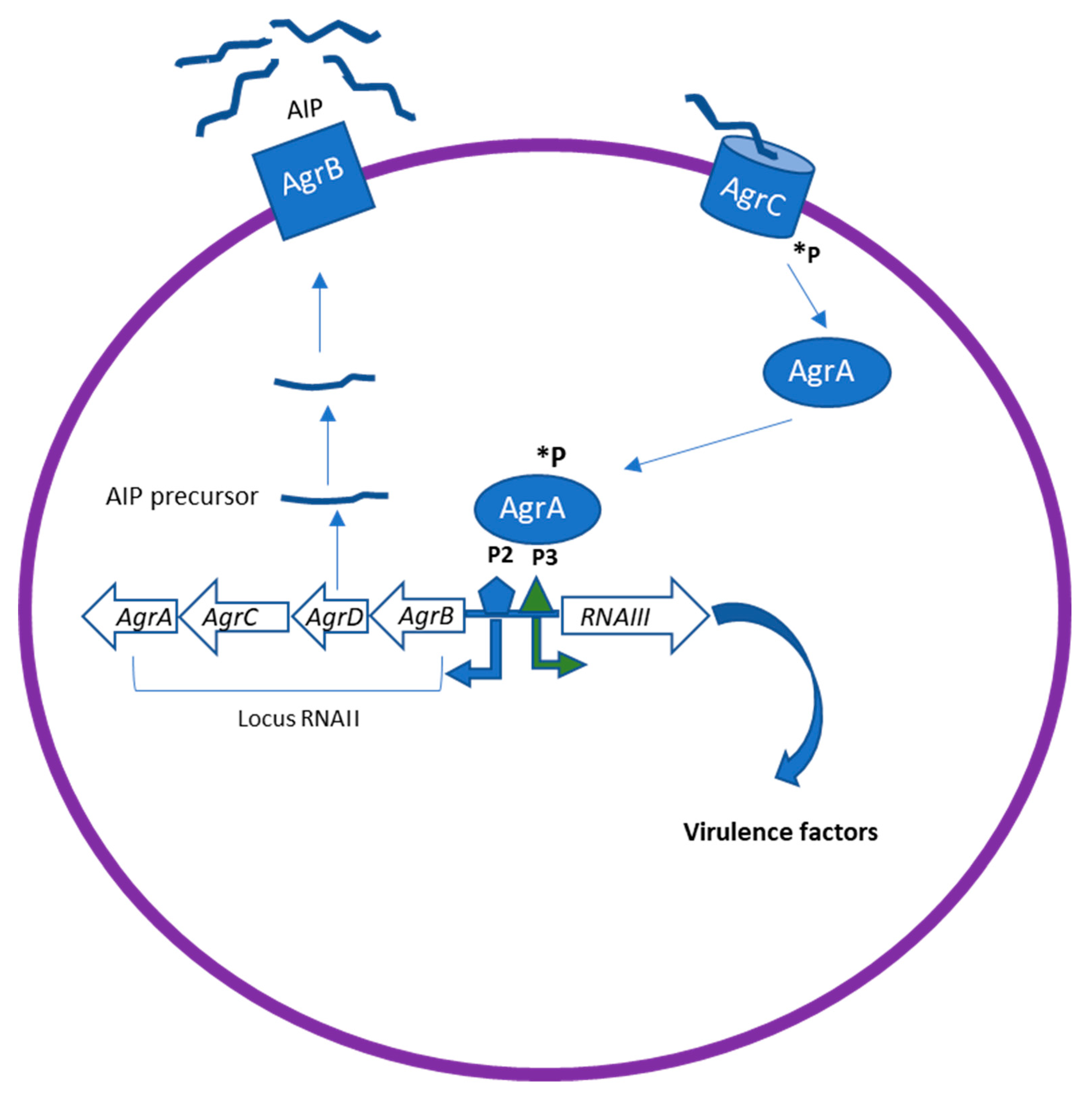
2. Quorum Sensing Mechanism
2.1. QS Molecular Signaling Network of Gram-Negative Bacteria
2.2. QS Molecular Signaling Network of Gram-Positive Bacteria
3. Anti-Biofilm Activity of Natural Compounds Against Pseudomonas aeruginosa
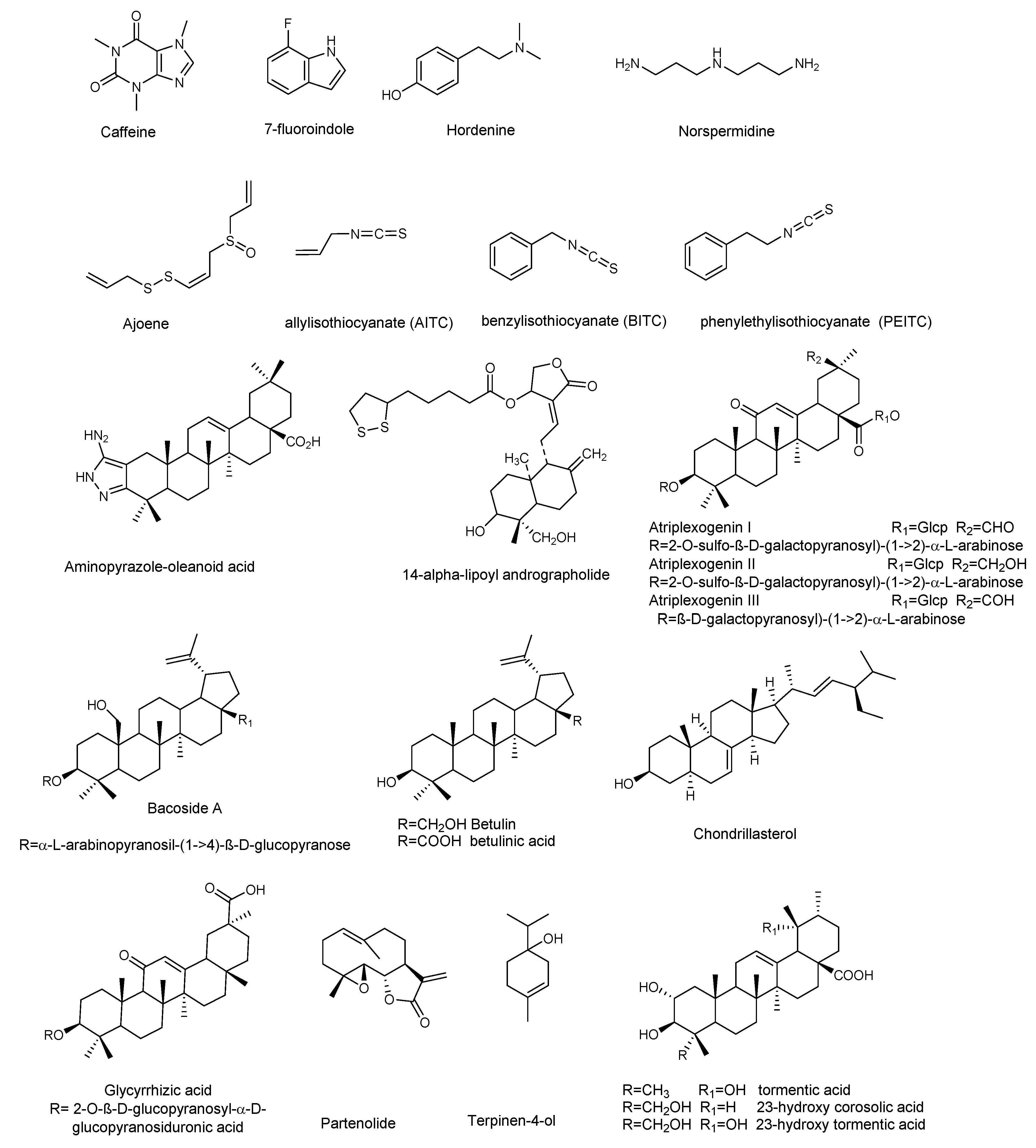
3.1. Alkaloids and Nitrogen-Containing Compounds
3.2. Terpenoids
3.3. Organosulfur Compounds
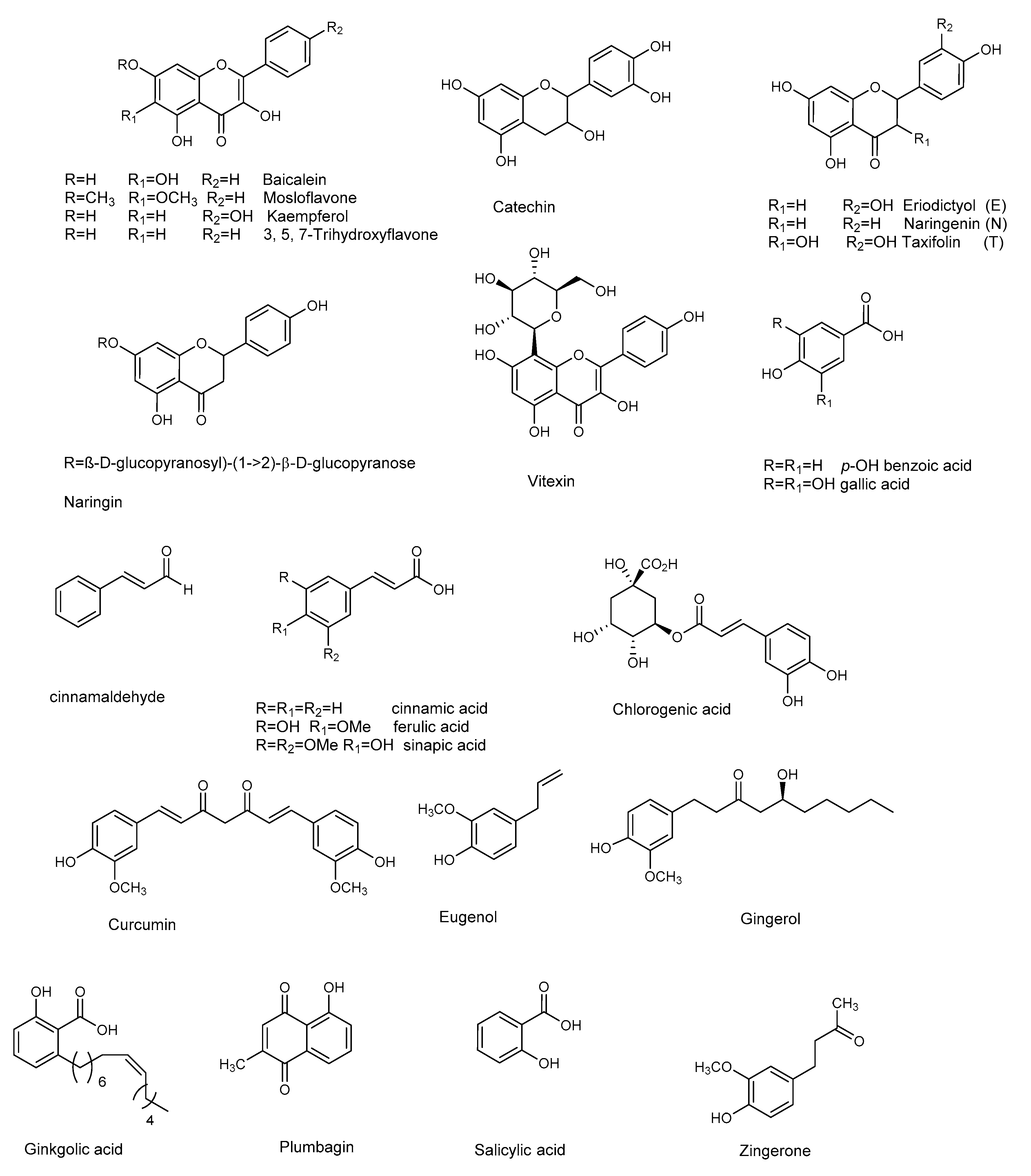
3.4. Flavonoids
3.5. Other Phenolic Compounds
4. Anti-Biofilm Properties of Natural Compounds against Staphylococcus aureus
4.1. Terpenes
4.2. Flavonoids
4.3. Other Phenolic Compounds
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Stacy, A.; McNally, L.; Darch, S.E.; Brown, S.P.; Whiteley, M. The biogeography of polymicrobial infection. Nat. Rev. Microbiol. 2016, 14, 93–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kostakioti, M.; Hadjifrangiskou, M.; Hultgren, S.J. Bacterial biofilms: Development, dispersal, and therapeutic strategies in the dawn of the postantibiotic era. Cold Spring Harb. Perspect. Med. 2013, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roilides, E.; Simitsopoulou, M.; Katragkou, A.; Walsh, T.J. How biofilms evade host defenses. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Trivedi, U.; Parameswaran, S.; Armstrong, A.; Burgueno-Vega, D.; Griswold, J.; Dissanaike, S.; Rumbaugh, K.P. Prevalence of multiple antibiotic resistant infections in diabetic versus nondiabetic Wounds. J. Pathog. 2014, 173053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, M.; Anju, C.P.; Biswas, L.; Kumar, V.A.; Mohan, C.G.; Biswas, R. Antibiotic resistance in Pseudomonas aeruginosa and alternative therapeutic options. Int. J. Med. Microbiol. 2016, 306, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, D.; Peters, B.M.; Li, L.; Li, B.; Xu, Z.; Shirliff, M.E. Staphylococcal chromosomal cassettes mec (SCCmec): A mobile genetic element in methicillin-resistant Staphylococcus aureus. Microb. Pathog. 2016, 101, 56–67. [Google Scholar] [CrossRef]
- Ibberson, C.B.; Whiteley, M. The social life of microbes in chronic infection. Curr. Opin. Microbiol. 2020, 53, 44–50. [Google Scholar] [CrossRef]
- Limoli, D.H.; Whitfield, G.B.; Kitao, T.; Ivey, M.L.; Davis, M.R., Jr.; Grahl, N.; Hogan, D.A.; Rahme, L.G.; Howell, P.L.; O’Toole, G.A.; et al. Pseudomonas aeruginosa alginate overproduction promotes coexistence with Staphylococcus aureus in a model of cystic fibrosis respiratory infection. MBio 2017, 8, e00186-17. [Google Scholar] [CrossRef] [Green Version]
- Limoli, D.H.; Hoffman, L.R. Help, hinder, hide and harm: What can we learn from the interactions between Pseudomonas aeruginosa and Staphylococcus aureus during respiratory infections? Thorax 2019, 74, 684–692. [Google Scholar] [CrossRef] [Green Version]
- Borges, A.; Abreu, A.C.; Dias, C.; Saavedra, M.J.; Borges, F.; Simões, M. New perspectives on the use of phytochemicals as an emergent strategy to Control Bacterial Infections Including Biofilms. Molecules 2016, 21, 877. [Google Scholar] [CrossRef]
- Shaw, E.; Wuest, W.M. Virulence attenuating combination therapy: A potential multi-target synergy approach to treat Pseudomonas aeruginosa infections in cystic fibrosis patients. RSC Med. Chem. 2020, 11, 358–369. [Google Scholar] [CrossRef]
- Asfour, H.Z. Anti-quorum sensing natural compounds. J. Microsc. Ultrastruct. 2018, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- D’Abrosca, B.; Buommino, E.; D’Angelo, G.; Coretti, L.; Scognamiglio, M.; Severino, V.; Pacifico, S.; Donnarumma, G.; Fiorentino, A. Spectroscopic identification and anti-biofilm properties of polar metabolites from the medicinal plant Helichrysum italicum against Pseudomonas aeruginosa. Bioorg. Med. Chem. 2013, 21, 7038–7046. [Google Scholar] [CrossRef]
- Lee, J.H.; Regmi, S.C.; Kim, J.A.; Cho, M.H.; Yun, H.; Lee, C.S.; Lee, J. Apple flavonoid phloretin inhibits Escherichia coli O157:H7 biofilm formation and ameliorates colon inflammation in rats. Infect. Immun. 2011, 79, 4819–4827. [Google Scholar] [CrossRef] [Green Version]
- Stowe, S.D.; Richards, J.J.; Tucker, A.T.; Thompson, R.; Melander, C.; Cavanagh, J. Anti-Biofilm Compounds Derived from Marine Sponges. Mar. Drugs 2011, 9, 2010–2035. [Google Scholar] [CrossRef] [Green Version]
- Masák, J.; Čejková, A.; Schreiberová, O.; Rezanka, T. Pseudomonas biofilms: Possibilities of their control. FEMS Microbiol. Ecol. 2014, 89, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Xia, Y.X.; He, Z.D.; Zhang, H.J. A Review of Natural Products with Anti-biofilm Activity. Curr. Org. Chem. 2018, 22, 789–817. [Google Scholar] [CrossRef]
- Waters, C.M.; Bassler, B. Quorum sensing: Cell-to-cell communication in bacteria. Annu. Rev. Cell Dev. Biol. 2005, 21, 319–346. [Google Scholar] [CrossRef] [Green Version]
- Flemming, H.; Wingende, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Lu, L.; Hu, W.; Tian, Z.; Yuan, D.; Yi, G.; Zhou, Y.; Cheng, Q.; Zhu, J.; Li, M. Developing natural products as potential anti-biofilm agents. Chin. Med. 2019, 14, 11. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Moser, C.; Wang, H.Z.; Høiby, N.; Song, Z.J. Strategies for combating bacterial biofilm infections. Int. J. Oral Sci. 2015, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal-response systems in gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef]
- García-Reyes, S.; Soberón-Chávez, G.; Cocotl-Yanez, M. The third quorum-sensing system of Pseudomonas aeruginosa: Pseudomonas quinolone signal and the enigmatic PqsE protein. J. Med. Microbiol. 2020, 69, 25–34. [Google Scholar] [CrossRef]
- Le, K.Y.; Otto, M. Quorum-sensing regulation in staphylococci-an overview. Front. Microbiol. 2015, 6, 1174. [Google Scholar] [CrossRef] [Green Version]
- Cheung, G.Y.; Wang, R.; Khan, B.A.; Sturdevant, D.E.; Otto, M. Role of the accessory gene regulator agr in community-associated methicillin-resistant Staphylococcus aureus pathogenesis. Infect. Immun. 2011, 79, 1927–1935. [Google Scholar] [CrossRef] [Green Version]
- Hapke, H.; Strathmann, W. Pharmacological effects of hordenine. Dtsch. Tierarztl. Wochenschr. 1995, 102, 228–232. [Google Scholar]
- Zhou, J.W.; Luo, H.Z.; Jiang, H.; Jian, T.K.; Chen, Z.Q.; Jia, A.Q. Hordenine: A Novel Quorum Sensing Inhibitor and Anti-biofilm Agent against Pseudomonas aeruginosa. J. Agric. Food Chem. 2018, 66, 1620–1628. [Google Scholar] [CrossRef]
- O’May, C.; Tufenkji, N. The swarming motility of Pseudomonas aeruginosa is blocked by cranberry proanthocyanidins and other tannin-containing materials. Appl. Environ. Microbiol. 2011, 77, 3061–3067. [Google Scholar] [CrossRef] [Green Version]
- Köhler, T.; Guanella, R.; Carlet, J.; van Delden, C. Quorum sensing-dependent virulence during Pseudomonas aeruginosa colonisation and pneumonia in mechanically ventilated patients. Thorax 2010, 65, 703–710. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, P.; Dastidar, D.G.; Paul, P.; Dutta, S.; Basu, D.; Sharma, S.R.; Basu, S.; Sarker, R.K.; Sen, A.; Sarkar, A.; et al. Inhibition of biofilm formation of Pseudomonas aeruginosa by caffeine: A potential approach for sustainable management of biofilm. Arch. Microbiol. 2020, 202, 623–635. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, Y.G.; Cho, M.H.; Kim, J.A.; Lee, J. 7-fluoroindole as an antivirulence compound against Pseudomonas aeruginosa. FEMS Microbiol. Lett. 2012, 329, 36–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabor, C.W.; Tabor, H. Polyamines. Ann. Rev. Biochem. 1984, 53, 749–790. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; She, P.; Wang, Y.; Liu, F.; Zhang, D.; Chen, L.; Luo, Z.; Xu, H.; Qi, Y.; Wu, Y. Effects of norspermidine on Pseudomonas aeruginosa biofilm formation and eradication. Microbiologyopen 2016, 5, 402–412. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, S.J.; Mutters, N.T.; Blessing, B.; Günther, F. Natural isothiocyanates express antimicrobial activity against developing and mature biofilms of Pseudomonas aeruginosa. Fitoterapia 2017, 119, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, T.H.; van Gennip, M.; Phipps, R.K.; Shanmugham, M.S.; Christensen, L.D.; Alhede, M.; Skindersoe, M.E.; Rasmussen, T.B.; Friedrich, K.; Uthe, F.; et al. Ajoene, a Sulfur-Rich Molecule from Garlic, Inhibits Genes Controlled by Quorum Sensing. Antimicrob. Agents Chemother. 2012, 56, 2314. [Google Scholar] [CrossRef] [Green Version]
- Bose, S.K.; Chauhan, M.; Dhingra, N.; Chhibber, S.; Harjai, K. Terpinen-4-ol attenuates quorum sensing regulated virulence factors and biofilm formation in Pseudomonas aeruginosa. Future Microbiol. 2020, 15, 127–142. [Google Scholar] [CrossRef]
- Pareek, A.; Suthar, M.; Rathore, G.S.; Bansal, V. Feverfew (Tanacetum parthenium L.): A systematic review. Pharmacogn. Rev. 2011, 5, 103–110. [Google Scholar] [CrossRef] [Green Version]
- Kalia, M.; Yadav, V.K.; Singh, P.K.; Sharma, D.; Narvi, S.S.; Agarwal, V. Exploring the impact of parthenolide as anti-quorum sensing and anti-biofilm agent against Pseudomonas aeruginosa. Life Sci. 2018, 199, 96–103. [Google Scholar] [CrossRef]
- Lawrence, J.A.; Huang, Z.; Rathinavelu, S.; Hua, J.F.; Garoa, E.; Ellis, M.; Normana, V.L.; Buckle, R.; Williams, R.B.; Starks, C.M.; et al. Optimized plant compound with potent anti-biofilm activity across gram-negative. Bioorg. Med. Chem. 2020, 28, 115–229. [Google Scholar] [CrossRef]
- Ghosh, C.; Bhowmik, J.; Ghosh, R.; Das, M.C.; Sandhu, P.; Kumari, M.; Acharjee, S.; Daware, A.V.; Akhter, Y.; Banerjee, B.; et al. The anti-biofilm potential of triterpenoids isolated from Sarcochlamys pulcherrima (Roxb.) Gaud. Microb. Pathog. 2020, 139, 103901. [Google Scholar] [CrossRef]
- Ghosh, R.; Das, M.C.; Sarkar, A.; Das, A.; Sandhu, P.; Dinda, B.; Akhter, Y.; Bhattacharjee, S.; De, U.C. Exploration of Phytoconstituents from Mussaenda roxburghii and Studies of their Antibiofilm Effect. Chem. Biodivers. 2017, 14, e1700165. [Google Scholar] [CrossRef] [PubMed]
- Rajkumari, J.; Borkotoky, S.; Murali, A.; Suchiang, K.; Mohanty, S.K.; Busi, S. Attenuation of quorum sensing controlled virulence factors and biofilm formation in Pseudomonas aeruginosa by pentacyclic triterpenes, betulin and betulinic acid. Microb. Pathog. 2018, 118, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Parai, D.; Islam, E.; Mitra, J.; Mukherjee, S.K. Effect of Bacoside A on growth and biofilm formation by Staphylococcus aureus and Pseudomonas aeruginosa. Can. J. Microbiol. 2017, 63, 169–178. [Google Scholar] [CrossRef] [Green Version]
- Stanković, J.; Gođevac, D.; Tešević, V.; Dajić-Stevanović, Z.; Ćirić, A.; Soković, M.; Novaković, M. Antibacterial and Antibiofilm Activity of Flavonoid and Saponin Derivatives from Atriplex tatarica against Pseudomonas aeruginosa. J. Nat. Prod. 2019, 82, 1487–1495. [Google Scholar] [CrossRef]
- Chakotiya, A.S.; Tanwar, A.; Narula, A.; Sharma, R.K. Alternative to antibiotics against Pseudomonas aeruginosa: Effects of Glycyrrhiza glabra on membrane permeability and inhibition of efflux activity and biofilm formation in Pseudomonas aeruginosa and its in vitro time-kill activity. Microb. Pathog. 2016, 98, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Mozirandi, W.; Tagwireyi, D.; Mukanganyama, S. Evaluation of antimicrobial activity of chondrillasterol isolated from Vernonia adoensis (Asteraceae). BMC Complement. Altern. Med. 2019, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, X.; Liu, X.; Bian, J.; Pei, G.; Dai, H.; Polyak, S.W.; Song, F.; Ma, L.; Wang, Y.; Zhang, L. Synergistic effect of 14-alpha-lipoyl andrographolide and various antibiotics on the formation of biofilms and production of exopolysaccharide and pyocyanin by Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2011, 55, 3015–3017. [Google Scholar] [CrossRef] [Green Version]
- Dietz, B.M.; Hajirahimkhan, A.; Dunlap, T.L.; Bolton, J.L. Botanicals and their bioactive phytochemicals for women’s health. Pharmacol. Rev. 2016, 68, 1026–1073. [Google Scholar] [CrossRef]
- Kuang, Z.; Hao, Y.; Walling, B.E.; Jeffries, J.L.; Ohman, D.E.; Gee, W.L. Pseudomonas aeruginosa elastase provides an escape from phagocytosis by degrading the pulmonary surfactant protein-A. PLoS ONE 2011, 6, e7091. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, A.; Harminder, A.K.; Singh, V. A review on the taxonomy, ethnobotany, chemistry and pharmacology of Oroxylum indicum Vent. Indian J. Pharm Sci. 2011, 73, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Chen, Q.; Zhou, W.; Gao, S.; Lin, H.; Ye, S.; Xu, S.; Cai, J. Chinese medicine injection Shuanghuanglian for treatment of acute upper respiratory tract infection: A systematic review of randomized controlled trials. J. Evid. Based Complement. Altern. Med. 2013, 2013, 987326. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Kong, J.L.; Dong, B.Y.; Huang, H.; Wang, K.; Wu, L.H.; Hou, C.C.; Liang, Y.; Li, B.; Chen, Y.Q. Baicalein attenuates the quorum sensing-controlled virulence factors of Pseudomonas aeruginosa and relieves the inflammatory response in P. aeruginosa-infected macrophages by downregulating the MAPK and NFκB signal-transduction pathways. Drug Des. Dev. Ther. 2016, 10, 183–203. [Google Scholar] [CrossRef] [Green Version]
- Vandeputte, O.; Kiendrebeogo, M.; Rasamiravaka, T.; Stévigny, C.; Duez, P.; Rajaonson, S.; Diallo, B.; Mol, A.; Baucher, M.; El Jaziri, M. The flavanone naringenin reduces the production of quorum sensing-controlled virulence factors in Pseudomonas aeruginosa PAO1. Microbiology 2011, 157, 2120–2132. [Google Scholar] [CrossRef] [Green Version]
- Vandeputte, O.M.; Kiendrebeogo, M.; Rajaonson, S.; Diallo, B.; Mol, A.; Jaziri, M.E.; Bacher, M. Identification of catechin as one of the flavonoids from Combretum albiflorum bark extract that reduces the production of quorum-sensing-controlled virulence factors in Pseudomonas aeruginosa PAO1. Appl. Environ. Microbiol. 2010, 76, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Dey, P.; Parai, D.; Banerjee, M.; Hossain, S.T.; Mukherjee, S.K. Naringin sensitizes the antibiofilm effect of ciprofloxacin and tetracycline against Pseudomonas aeruginosa biofilm. Int. J. Med. Microbiol. 2020, 310, 151410. [Google Scholar] [CrossRef]
- Abinaya, M.; Gayathri, M. Inhibition of biofilm formation, quorum sensing activity and molecular docking study of isolated 3, 5, 7-Trihydroxyflavone from Alstonia scholaris leaf against P. aeruginosa. Bioorg. Chem. 2019, 87, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Das, M.C.; Sandhu, P.; Gupta, P.; Rudrapaul, P.; De, U.C.; Tribedi, P.; Akhter, Y.; Bhattacharjee, S. Attenuation of Pseudomonas aeruginosa biofilm formation by Vitexin: A combinatorial study with azithromycin and gentamicin. Sci. Rep. 2016, 6, 23347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lalitha, K.; Sandeep, M.; Prasad, Y.S.; Sridharan, V.; Maheswari, C.U.; Srinandan, C.S.; Nagarajan, S. Intrinsic Hydrophobic Antibacterial Thin Film from renewable Resources: Application in the Development of Anti-Biofilm Urinary Catheters. Acs Sustain. Chem. Eng. 2017, 5, 436–449. [Google Scholar] [CrossRef]
- Deepika, M.S.; Thangam, R.; Sundarraj, S.; Sheena, T.S.; Sivasubramanian, S.; Kulandaivel, J.; Thirumurugan, R. Co-delivery of Diverse Therapeutic Compounds Using PEG-PLGA Nanoparticle Cargo against Drug-Resistant Bacteria: An Improved Anti-biofilm Strategy. ACS Appl. Bio Mater. 2020, 3, 385–399. [Google Scholar] [CrossRef] [Green Version]
- Hnamte, S.; Subhaswaraj, P.; Ranganathan, S.K.; Ampasala, D.R.; Muralitharan, G.; Siddhardha, B. Methanolic Extract of Plectranthus tenuiflorus Attenuates Quorum Sensing Mediated Virulence and Biofilm Formation in Pseudomonas aeruginosa PAO1. J. Pure Appl. Microbiol. 2018, 12, 1985–1996. [Google Scholar] [CrossRef] [Green Version]
- Vasavi, H.S.; Arun, A.B.; Rekha, P.D. Anti-quorum sensing activity of flavonoid-rich fraction from Centella asiatica L. against Pseudomonas aeruginosa PAO1. J. Microbiol. Immunol. Infect. 2016, 49, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Moghadamtousi, S.Z.; Kadir, H.A.; Hassandarvish, P.; Tajik, H.; Abubakar, S.; Zandi, K. A review on antibacterial, antiviral, and antifungal activity of curcumin. BioMed Res. Int. 2014, 2014, 186864. [Google Scholar] [CrossRef]
- Rudrappa, T.; Bais, H.P. Curcumin, a known phenolic from Curcuma longa, attenuates the virulence of Pseudomonas aeruginosa PAO1 in whole plant and animal pathogenicity models. J. Agric Food Chem. 2008, 56, 1955–1962. [Google Scholar] [CrossRef]
- Prateeksha Rao, C.V.; Das, A.K.; Barik, S.K.; Singh, B.N. ZnO/Curcumin Nanocomposites for Enhanced Inhibition of Pseudomonas aeruginosa Virulence via LasR-RhlR Quorum Sensing Systems. Mol. Pharm. 2019, 16, 3399–3413. [Google Scholar] [CrossRef]
- Gupta, P.; Sarkar, A.; Sandhu, P.; Daware, A.; Das, M.C.; Akhter, Y.; Bhattacharjee, S. Potentiation of antibiotic against Pseudomonas aeruginosa biofilm: A study with plumbagin and gentamicin. J. Appl. Microbiol. 2017, 123, 246–261. [Google Scholar] [CrossRef]
- Acharya, B.R.; Bhattacharyya, B.; Chakrabarti, G. The natural naphthoquinone plumbagin exhibits antiproliferative activity and disrupts the microtubule network through tubulin binding. Biochemistry 2008, 47, 7838–7845. [Google Scholar] [CrossRef]
- Rajkumari, J.; Borkotoky, S.; Murali, A.; Suchiang, K.; Mohanty, S.K.; Busi, S. Cinnamic acid attenuates quorum sensing associated virulence factors and biofilm formation in Pseudomonas aeruginosa PAO1. Biotechnol. Lett. 2018, 40, 1087–1100. [Google Scholar] [CrossRef]
- Tahrioui, A.; Ortiz, S.; Azuama, O.C.; Bouffartigues, E.; Benalia, N.; Tortuel, D.; Maillot, O.; Chemat, S.; Kritsanida, M.; Feuilloley, M.; et al. Membrane-interactive compounds from Pistacia lentiscus L. thwart Pseudomonas aeruginosa virulence. Front. Microbiol. 2020, 11, 1068. [Google Scholar] [CrossRef]
- Zhou, L.; Zheng, H.; Tang, Y.; Yu, W.; Gong, Q. Eugenol inhibits quorum sensing at sub-inhibitory concentrations. Biotechnol. Lett. 2013, 35, 631–637. [Google Scholar] [CrossRef]
- Plyuta, V.; Zaitseva, J.; Lobakova, E.; Zagoskina, N.; Kuznetsov, A.; Khmel, I. Effect of plant phenolic compounds on biofilm formation by Pseudomonas aeruginosa. APMIS 2013, 121, 1073–1081. [Google Scholar] [CrossRef]
- Borges, A.; Saavedra, M.J.; Simões, M. The activity of ferulic and gallic acids in biofilm prevention and control of pathogenic bacteria. Biofouling 2012, 28, 755–767. [Google Scholar] [CrossRef]
- Ahmed, S.A.; Rudden, M.; Smyth, T.J.; Dooley, J.S.; Marchant, R.; Banat, I.M. Natural quorum sensing inhibitors effectively down-regulate gene expression of Pseudomonas aeruginosa virulence factors. Appl. Microbiol. Biotechnol. 2019, 103, 3521–3535. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Lee, S.; Byun, Y.; Park, H.D. 6-Gingerol reduces Pseudomonas aeruginosa biofilm formation and virulence via quorum sensing inhibition. Sci. Rep. 2015, 5, 8656. [Google Scholar] [CrossRef]
- Kumar, L.; Chhibber, S.; Harjai, K. Zingerone inhibit biofilm formation and improve antibiofilm efficacy of ciprofloxacin against Pseudomonas aeruginosa PAO1. Fitoterapia 2013, 90, 73–78. [Google Scholar] [CrossRef]
- Kumar, L.; Chhibber, S.; Kumar, R.; Kumar, M.; Harjai, K. Zingerone silences quorum sensing and attenuates virulence of Pseudomonas aeruginosa. Fitoterapia 2015, 102, 84–95. [Google Scholar] [CrossRef]
- Ulrey, R.K.; Barksdale, S.M.; Zhou, W.; van Hoek, M.L. Cranberry proanthocyanidins have anti-biofilm properties against Pseudomonas aeruginosa. BMC Complement. Altern. Med. 2014, 14, 499. [Google Scholar] [CrossRef] [Green Version]
- Schilcher, K.; Horswill, A.R. Staphylococcal Biofilm Development: Structure, Regulation, and Treatment Strategies. Microbiol. Mol. Biol. 2020, 84, e00026-19. [Google Scholar] [CrossRef]
- Wu, S.C.; Liu, F.; Zhu, K.; Shen, J.Z. Natural Products That Target Virulence factors in Antibiotic-Resistant Staphylococcus aureus. J. Agric. Food Chem. 2019, 67, 13195–13211. [Google Scholar] [CrossRef]
- Merghni, A.; Noumi, E.; Hadded, O.; Dridi, N.; Panwar, H.; Ceylan, O.; Mastouri, M.; Snoussi, M. Assessment of the antibiofilm and antiquorum sensing activities of Eucalyptus globulus essential oil and its main component 1,8-cineole against methicillin-resistant Staphylococcus aureus strains. Microb. Pathog. 2018, 118, 74–80. [Google Scholar] [CrossRef]
- Burt, S.A.; Ojo-Fakunle, V.T.; Woertman, J.; Veldhuizen, E.J. The Natural Antimicrobial Carvacrol Inhibits Quorum Sensing in Chromobacterium violaceum and Reduces Bacterial Biofilm Formation at Sub-Lethal Concentrations. PLoS ONE 2014, 9, e93414. [Google Scholar] [CrossRef] [Green Version]
- Musthafa, K.S.; Voravuthikunchai, S.P. Anti-virulence potential of eugenyl acetate against pathogenic bacteria of medical importance. Antonie Leeuwenhoek. 2015, 107, 703–710. [Google Scholar] [CrossRef] [PubMed]
- Farha, A.K.; Yang, Q.Q.; Kim, G.; Zhang, D.; Mavumengwana, V.; Habimana, O.; Li, H.B.; Corke, H.; Gana, R.Y. Inhibition of multidrug-resistant foodborne Staphylococcus aureus biofilms by a natural terpenoid (+)-nootkatone and related molecular mechanism. Food Control. 2020, 107154. [Google Scholar] [CrossRef]
- Walencka, E.; Rozalska, S.; Wysokinska, H.; Rozalski, M.; Kuzma, L.; Rozalska, B. Salvipisone and aethiopinone from Salvia sclarea hairy roots modulate staphylococcal antibiotic resistance and express anti-biofilm activity. Planta Med. 2007, 73, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.G.; Lee, S.Y.; Lee, S.M.; Lim, K.H.; Ha, E.J.; Eom, Y.B. Activity of novel inhibitors of Staphylococcus aureus biofilms. Folia Microbiol. 2017, 62, 157–167. [Google Scholar] [CrossRef]
- Qin, N.; Tan, X.; Jiao, Y.; Liu, L.; Zhao, W.; Yang, S.; Jia, A. RNA-Seq-based transcriptome analysis of methicillin-resistant Staphylococcus aureus biofilm inhibition by ursolic acid and resveratrol. Sci. Rep. 2014, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Artini, M.; Papa, R.; Barbato, G.; Scoarughi, G.L.; Cellini, A.; Morazzoni, P.; Bombardelli, E.; Selan, L. Bacterial biofilm formation inhibitory activity revealed for plant derived natural compounds. Bioorg. Med. Chem. 2012, 20, 920–926. [Google Scholar] [CrossRef]
- Luo, M.; Qiu, J.; Zhang, Y.; Wang, J.; Dong, J.; Li, H.; Leng, B.; Zhang, Q.; Dai, X.; Niu, X.; et al. alpha-Cyperone Alleviates Lung Cell Injury Caused by Staphylococcus aureus via Attenuation of alpha-Hemolysin Expression. J. Microbiol. Biotechnol. 2012, 22, 1170–1176. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, T.; Wang, K.; Hou, C.; Cai, S.; Huang, Y.; Du, Z.; Huang, H.; Kong, J.; Chen, Y. Baicalein Inhibits Staphylococcus aureus Biofilm Formation and the Quorum Sensing System In Vitro. PLoS ONE 2016, 11, e0153468. [Google Scholar] [CrossRef] [Green Version]
- Silva, L.N.; Da Hora, G.C.; Soares, T.A.; Bojer, M.S.; Ingmer, H.; Macedo, A.J.; Trentin, D.S. Myricetin protects Galleria mellonella against Staphylococcus aureus infection and inhibits multiple virulence factors. Sci. Rep. 2017, 7, 2823. [Google Scholar] [CrossRef]
- Lopez, G.C.; Sanchez, C.A. Quercetin attenuates Staphylococcus aureus virulence by reducing alpha-toxin secretion. Rev. Argent. Microbiol. 2018, 50, 131–135. [Google Scholar] [CrossRef]
- Qiu, J.Z.; Xiang, H.; Hu, C.; Wang, Q.A.; Dong, J.; Li, H.E.; Luo, M.J.; Wang, J.F.; Deng, X.M. Subinhibitory concentrations of farrerol reduce alpha-toxin expression in Staphylococcus aureus. FEMS Microbiol. Lett. 2011, 315, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Li, H.; Wang, L.; Song, Z.; Shi, L.; Li, W.; Deng, X.; Wang, J. Isorhamnetin Attenuates Staphylococcus aureus-Induced Lung Cell Injury by Inhibiting Alpha-Hemolysin Expression. J. Microbiol. Biotechnol. 2016, 26, 596–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, Z.H.; Shi, D.X.; Liu, H.Y.; Shen, Z.Y.; Zha, Y.H.; Li, W.H.; Deng, X.M.; Wang, J.F. Lysionotin attenuates Staphylococcus aureus pathogenicity by inhibiting alpha-toxin expression. Appl. Microbiol. Biotechnol. 2017, 101, 6697–6703. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhou, X.; Li, W.H.; Zhang, H.; Zhang, B.; Li, G.; Liu, B.W.; Deng, X.M.; Peng, L.P. Diosmetin inhibits the expression of alpha-hemolysin in Staphylococcus aureus. Antonie Leeuwenhoek. 2015, 108, 383–389. [Google Scholar] [CrossRef]
- Dong, J.; Qiu, J.Z.; Wang, J.F.; Li, H.E.; Dai, X.H.; Zhang, Y.; Wang, X.; Tan, W.; Niu, X.D.; Deng, X.M.; et al. Apigenin alleviates the symptoms of Staphylococcus aureus pneumonia by inhibiting the production of alpha-hemolysin. FEMS Microbiol. Lett. 2013, 338, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.L.; Chen, C.Z.; Pan, J.; Deng, X.M.; Wang, J.F. Epigallocatechin gallate can attenuate human alveolar epithelial cell injury induced by alpha-haemolysin. Microb. Pathog. 2018, 115, 222–226. [Google Scholar] [CrossRef]
- Qiu, J.Z.; Niu, X.D.; Dong, J.; Wang, D.C.; Wang, J.F.; Li, H.G.; Luo, M.J.; Li, S.T.; Feng, H.H.; Deng, X.M. Baicalin Protects Mice From Staphylococcus aureus Pneumonia Via Inhibition of the Cytolytic Activity of alpha-Hemolysin. J. Infect. Dis. 2012, 206, 292–301. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Qiu, J.Z.; Zhang, Y.; Lu, C.J.; Dai, X.H.; Wang, J.F.; Li, H.G.; Wang, X.; Tan, W.; Luo, M.J.; et al. Oroxylin A Inhibits Hemolysis via Hindering the Self-Assembly of alpha-Hemolysin Heptameric Transmembrane Pore. PLoS Comput. Biol. 2013, 9, e1002869. [Google Scholar] [CrossRef]
- Liu, Y.M.; Shi, D.X.; Guo, Y.; Li, M.; Zha, Y.H.; Wang, Q.K.; Wang, J.F. Dracorhodin Perochlorate attenuates Staphylococcus aureus USA300 virulence by decreasing alpha-toxin expression. World J. Microbiol. Biotechnol. 2017, 33, 17. [Google Scholar] [CrossRef]
- Wang, X.; Dong, J.; Dai, X.H.; Zhang, Y.; Wang, J.F.; Li, H.G.; Lu, C.J.; Tan, W.; Gao, X.H.; Deng, X.M.; et al. Silibinin In Vitro Protects A549 Cells from Staphylococcus aureus-Mediated Injury and In Vivo Alleviates the Lung Injury of Staphylococcal Pneumonia. Planta Med. 2013, 79, 110–115. [Google Scholar] [CrossRef]
- Al-Shabib, N.A.; Husain, F.M.; Ahmad, I.; Khan, M.S.; Khan, R.A.; Khan, J.M. Rutin inhibits mono and multi-species biofilm formation by foodborne drug resistant Escherichia coli and Staphylococcus aureus. Food Control. 2017, 79, 325–332. [Google Scholar] [CrossRef]
- Elmasri, W.A.; Yang, T.; Tran, P.; Hegazy, M.E.; Hamood, A.N.; Mechref, Y.; Paré, P.W. Teucrium polium phenylethanol and iridoid glycoside characterization and flavonoid inhibition of biofilm-forming Staphylococcus aureus. J. Nat. Prod. 2015, 78, 2–9. [Google Scholar] [CrossRef]
- Ming, D.; Wang, D.; Cao, F.; Xiang, H.; Mu, D.; Cao, J.; Li, B.; Zhong, L.; Dong, X.; Zhong, X.; et al. Kaempferol Inhibits the Primary Attachment Phase of Biofilm Formation in Staphylococcus aureus. Front. Microbiol. 2017, 8, 2263. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Liu, S.; Li, W.H.; Zhang, B.; Liu, B.W.; Liu, Y.; Deng, X.M.; Peng, L.P. Phloretin derived from apple can reduce alpha-hemolysin expression in methicillin-resistant Staphylococcus aureus USA300. World J. Microbiol. Biotechnol. 2015, 31, 1259–1265. [Google Scholar] [CrossRef]
- Liu, S.; Liu, B.W.; Luo, Z.Q.; Qiu, J.M.; Zhou, X.A.; Li, G.; Zhang, B.; Deng, X.M.; Yang, Z.G.; Wang, J.F. The combination of osthole with baicalin protects mice from Staphylococcus aureus pneumonia. World J. Microbiol. Biotechnol. 2017. [Google Scholar] [CrossRef]
- Zhang, B.; Teng, Z.H.; Li, X.H.; Lu, G.J.; Deng, X.M.; Niu, X.D.; Wang, J.F. Chalcone Attenuates Staphylococcus aureus Virulence by Targeting Sortase A and Alpha-Hemolysin. Front. Microbiol. 2017, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Duan, J.; Li, M.; Hao, Z.; Shen, X.; Liu, L.; Jin, Y.; Wang, S.; Guo, Y.; Yang, L.; Wang, L.; et al. Subinhibitory concentrations of resveratrol reduce alpha-hemolysin production in Staphylococcus aureus isolates by down-regulating saeRS. Emerg. Microbes Infect. 2018, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.F.; Zhou, X.; Li, W.H.; Deng, X.M.; Deng, Y.H.; Niu, X.D. Curcumin protects mice from Staphylococcus aureus pneumonia by interfering with the self-assembly process of alpha-hemolysin. Sci. Rep. 2016, 6, 28254. [Google Scholar] [CrossRef] [Green Version]
- Peng, D.; Chen, A.L.; Shi, B.; Min, X.; Zhang, T.; Dong, Z.L.; Yang, H.; Chen, X.L.; Tian, Y.B.; Chen, Z.H. Preliminary study on the effect of brazilin on biofilms of Staphylococcus aureus. Exp. Ther. Med. 2018, 16, 2108–2118. [Google Scholar] [CrossRef]
- Vermote, A.; Brackman, G.; Risseeuw MD, P.; Vanhoutte, B.; Cos, P.; Van Hecke, K.; Breyne, K.; Meyer, E.; Coenye, T.; Van Calenbergh, S. Hamamelitannin Analogues that Modulate Quorum Sensing as Potentiators of Antibiotics against Staphylococcus aureus. Angew. Chem. Int. Ed. 2016, 55, 6551–6555. [Google Scholar] [CrossRef]
- Kiran, M.D.; Adikesavan, N.V.; Cirioni, O.; Giacometti, A.; Silvestri, C.; Scalise, G.; Ghiselli, R.; Saba, V.; Orlando, F.; Shoham, M.; et al. Discovery of a quorum-sensing inhibitor of drug resistant staphylococcal infections by structure-based virtual screening. Mol. Pharmacol. 2008, 73, 1578–1586. [Google Scholar] [CrossRef] [Green Version]
- Mun, S.H.; Kong, R.; Seo, Y.S.; Zhou, T.; Kang, O.H.; Shin, D.W.; Kwon, D.Y. Subinhibitory concentrations of punicalagin reduces expression of virulence-related exoproteins by Staphylococcus aureus. FEMS Microbiol. Lett. 2016, 363, fnw253. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Shi, C.; Wu, Q.; Zheng, Z.; Liu, P.; Li, G.; Peng, X.; Xia, X. Antimicrobial Activity of Punicalagin Against Staphylococcus aureus and Its Effect on Biofilm Formation. Foodborne Pathog. Dis. 2017, 14, 282–287. [Google Scholar] [CrossRef]
- Dong, G.; Liu, H.; Yu, X.; Zhang, X.; Lu, H.; Zhou, T.; Cao, J. Antimicrobial and anti-biofilm activity of tannic acid against Staphylococcus aureus. Nat. Prod. Res. 2018, 32, 2225–2228. [Google Scholar] [CrossRef]
- Lin, M.H.; Chang, F.R.; Hua, M.Y.; Wu, Y.C.; Liu, S.T. Inhibitory effects of 1, 2, 3, 4, 6-penta-O-galloyl-β-D-glucopyranose on biofilm formation by Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55, 1021–1027. [Google Scholar] [CrossRef] [Green Version]
- Vollaro, A.; Catania, M.R.; Iesce, M.R.; Sferruzza, R.; D’Abrosca, B.; Donnarumma, G.; De Filippis, A.; Cermola, F.; DellaGreca, M.; Buommino, E. Antimicrobial and anti-biofilm properties of novel synthetic lignan-like compounds. New Microbiol. 2019, 42, 21–28. [Google Scholar]
- Zhang, L.; Quan, C.; Zhang, X.; Xiong, W.; Fan, S. Proteoliposome-based model for screening inhibitors targeting histidine kinase AgrC. Chem. Biol. Drug Des. 2019, 93, 712. [Google Scholar] [CrossRef]
- Saising, J.; Ongsakul, M.; Voravuthikunchai, S.P. Rhodomyrtus tomentosa (Aiton) Hassk. ethanol extract and rhodomyrtone: A potential strategy for the treatment of biofilm-forming staphylococci. J. Med. Microbiol. 2011, 60, 1793–1800. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | BIOFILM ASSAY | MOTILITY ASSAY | REF. | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Total Protein Content | Crystal Violet (CV) or Safranin (S) Staining | Metabolic Activity of Biofilm | EPS Production | Pre-Formed Biofilms | Swarming | Swimming | Twitching | ||
| Hordenine | ↓31% (1 mg/mL) | ↓23% (1 mg/mL) | ↓54% (0.5–1.0 mg/mL) | ↓82% (0.5–1.0 mg/mL) | [27] | ||||
| 7-fluoroindole | ↓76% (1 mM) | abolished swarming (1.0 mM) | no effect | no effect | [31] | ||||
| Caffeine | ↓~30% (80 µg/mL) | ↓~50% (80 µg/mL) | ↓~50% (80 µg/mL) | ↓ (40–80 µg/mL) | [30] | ||||
| Norspermidine | ↓48–90% (10 mmol/L) | ↓43–85% (10 mmol/L) | ↓ 42.4% (4 mmol/L) | [33] | |||||
| Allylisothio-cynate | no effect (50 μg/mL) | ↓65–70% (200–800 μg/mL) | [34] | ||||||
| Benzyl-isothiocyanate | no effect (50 μg/mL) | ↓70% (250–1000 μg/mL) | [34] | ||||||
| Phenylethyl-isothiocyanate | ↓40% (500 μg/mL) | ↓30–60% (60–240 μg/mL) | [34] | ||||||
| Compounds | VIRULENCE FACTORS Regulated by QS | REF. | |||||||
|---|---|---|---|---|---|---|---|---|---|
| AHLs Levels | Alginate Assay | Elastase Assay | Hemolysin Assay | Protease Assay | Pyocyanin Assay | Pyoverdine Secretion Assay | Rhamno-Lipid | ||
| Hordenine | ↓79% C4-HSL, ↓66% 3-oxo-C12-HSL (1 mg/mL) | ↓60%, Res↓ 50%: (1 mg/mL) | ↓65%, Res ↓30%: (1 mg/mL) | ↓61%, Res ↓40% (1 mg/mL) | ↓80%, Res ↓40%: (1 mg/mL) | ↓65%, Res ↓40%: (1 mg/mL) | ↓53%, Res ↓40%: (1 mg/mL) | [27] | |
| 7-fluoroindole | ↓93% (1 mM) | ↓ marked reduction | ↓ marked reduction | ↓ marked reduction | [31] | ||||
| Caffeine | ↓70% (80 µg/mL) | ↓~60% (80 µg/mL) | [30] | ||||||
| Norspermidine | ↓59–69% (0, 2, and 4 mmol/L) | ↓53–66% (0, 2, and 4 mmol/L) | ↓45–54% (0, 2, and 4 mmol/L) | [33] | |||||
| Ajoene | ↓ 3-fold C4-HSL (80 µg/mL) | ↓3-fold (20 µg/mL) | [35] | ||||||
| Compounds | BIOFILM ASSAY | MOTILITY ASSAY | REF. | |||||
|---|---|---|---|---|---|---|---|---|
| Total Protein Content | Crystal Violet (CV) or Safranin (S) Staining | EPS Production | Pre-Formed Biofilms | Swarming | Swimming | Twitching | ||
| Terpinen-4-ol | ↓ (0.06% v/v) | ↓ young, peak mature biofilm | ↓33,3% (0.06% v/v) | ↓50% (0.06% v/v) | ↓25% (0.06% v/v) | [36] | ||
| Parthenolide | ↓56% (1 mM) | ↓(1 mM) | [38] | |||||
| Aminopyrazole- oleanoid acid | ↓>85% (1 µg/mL) | [39] | ||||||
| Tormentic acid | ↓25.4% (25 µg/mL) | (S)↓53.8% (25 µg/mL) | ↓39.2% (25 µg/mL) | ↓6.8 mm (25 µg/mL) | [40] | |||
| 23-OH corosolic acid | ↓28.7% (20 µg/mL) | ↓55.6% (20 µg/mL) | ↓41.9% (20 µg/mL) | ↓6.2 mm (20 µg/mL) | [40] | |||
| 23-OH tormentic acid | ↓37.1% (0.37 mM) | (S) ↓37.6% (0.37 mM) | ↓36.9% (0.37 mM) | ↓2 ± 0.4 (0.21 mM) | [41] | |||
| Betulin | ↓57.3% (125 µg/mL) | ↓31.3% (125 µg/mL) | ↓47.3% (125 µg/mL) | ↓ (125 µg/mL) | ↓ (125 µg/mL) | [42] | ||
| Betulinic acid | ↓33.0% (125 µg/mL) | ↓35.2% (125 µg/mL) | ↓ 51.4% (125 µg/mL) | ↓ (125 µg/mL) | ↓ (125 µg/mL) | [42] | ||
| Bacoside | ↓90% (200 µg/mL) | 20% cell viability (200 µg/mL) | [43] | |||||
| Atriplexogenin I | ↓7.2–56.2% (125–500 µM) | [44] | ||||||
| Atriplexogenin II | ↓12.5–26.5% (62.5–250 µM) | [44] | ||||||
| Atriplexogenin III | ↓39.4–53.4% (345–690 µM) | [44] | ||||||
| Glycyrrhizic acid | ↓65.1–83.3% (50–200 mg/mL) | [45] | ||||||
| Chondrillasterol | ↓>90% (100 µg/mL) | ↓>60% (1.6–100 µg/mL) | [46] | |||||
| 14-alpha-lipoyl andrographolide | ↓ (0.5 mM) | ↓ (0.5 mM) | [47] | |||||
| Compounds | VIRULENCE FACTORS Regulated by QS | REF. | ||||||
|---|---|---|---|---|---|---|---|---|
| Alginate Assay | Elastase Assay | Hemolysin Assay | Protease Assay | Pyocyanin Assay | Pyoverdine Secretion Assay | Rhamno-Lipid | ||
| Terpinen-4-ol | ↓56% (0.06% v/v) | ↓50% (0.06% v/v) | ↓60% (0.06% v/v) | ↓5% (0.06% v/v) | ↓33% (0.06% v/v) | [36] | ||
| Parthenolide | ↓45% (1 mM) | ↓35.1% (1 mM) | [38] | |||||
| Tormentic acid | ↓ 53.0% (25 µg/mL) | ↓(20 µg/mL) | [40] | |||||
| 23-OH corosolic acid | ↓46.6% (20 µg/mL) | ↓(20 µg/mL) | [40] | |||||
| 23-OH tormentic acid | ↓37.8% (0.37 mM) | [41] | ||||||
| Betulin | ↓88.3% (125 µg/mL) | ↓74.5% (125 µg/mL) | ↓19.0% (125 µg/mL) | [42] | ||||
| Betulinic acid | ↓ 54.7% (125 µg/mL) | ↓54.7% (125 µg/mL) | ↓21.6% (125 µg/mL) | [42] | ||||
| 14-alpha-lipoyl andrographolide | ↓ (0.5 mM) | [47] | ||||||
| Compounds | BIOFILM ASSAY | MOTILITY ASSAY | REF. | |||||
|---|---|---|---|---|---|---|---|---|
| Total Protein Content | Crystal Violet (CV) or Safranin (S) Staining | Metabolic Activity of Biofilm | EPS Production | Swarming | Swimming | Twitching | ||
| Baicalein | ↓35.7% and ↓53% (1 and 5 days) at 128 µg/mL | ↓ (128 µg /mL) | no effect | ↓ (128 µg /mL) | [52] | |||
| Catechin | ↓ 30% (4mM) | [54] | ||||||
| Naringin | ↓49.5% (410 µg/mL), *Cpr, Tet | ↓49.5% (410 µg/mL), *Cpr, Tet | ↓ 40% (410 µg/mL), *Cpr, Tet | ↓ 42% (410 µg/mL) | ↓14% (410 µg/mL) | [55] | ||
| 3, 5, 7-Trihydroxyflavone | ↓76% (100) | ↓74,5% (100 µg/mL) | ↓(25–100 µg /mL) | [56] | ||||
| Vitexin | (S)↓56% (110 µg/mL) | ↓40% (110 μg/mL) | ↓100 µg /mL | [57] | ||||
| Compounds | VIRULENCE FACTORS Regulated by QS | REF. | ||||||
|---|---|---|---|---|---|---|---|---|
| AHLs Levels | Crystal Violet (CV) or Safranin (S) Staining | Elastase Assay | Protease Assay | Pyocyanin Assay | Pyoverdine Secretion Assay | Rhamno-Lipid | ||
| Baicalein | ↓ | (32–128 µg/mL) | ↓94.2%, LasB (128 µg/mL) | ↓74.56% LasA (128 µg/mL) | ↓69.9% (128 µg/mL) | ↓74.1% (128 µg/mL) | [52] | |
| Eriodictyol (E), Naringenin (N), Taxifolin (T) | ↓3-oxo-C12-HSL and C4-HSL N (4 mM) | ↓46% N, 62% E, 47% T (4 mM) | ↓87% N, 73% E, ¯56% T (4 mM) | [53] | ||||
| Catechin | ↓30% (4 mM) | ↓(0.125 and 16 mM) | [54] | |||||
| 3, 5, 7-Trihydroxyflavone | ↓52% (0.1 µg/mL) | no effect | [56] | |||||
| Vitexin | LasB ↓37.5% (110 µg/mL) | ↓39.04% inhibition of LasA | ↓ moderate (100 µg/mL) | ↓ moderate (100 µg /mL) | [57] | |||
| Compounds | BIOFILM ASSAY | MOTILITY ASSAY | REF. | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Total Protein Content | Crystal Violet (CV) or Safranin (S) Staining | Metabolic Activity of Biofilm | EPS Production | Pre-Formed Biofilms | Swarming | Swimming | Twitching | ||
| Plumbagin | ↓76% (150 µg/mL) | (S)↓60% (150 µg/mL) | ↓52% (150 µg/mL) | ↓41% (150 µg/mL) | ↓55.5% (250 µg/mL) | [65] | |||
| Cinnamic acid | ↓50.1 (250 µg/mL) | [67] | |||||||
| Curcumin | ↓1.5–3 µg/mL | [63] | |||||||
| Eugenol | ↓43% at 400 µM | No inhibition (200 μM) | [69] | ||||||
| Gallic acid | ↓2- to 2.5-fold | ↓20–50%, (400–800 μg/mL) | ↓0–15% (400–800 μg/mL) | [70] | |||||
| 4-OH benzoic acid | ↓12–30% (400–800 μg/mL) | ↓0–15% (400–800 μg/mL) | [70] | ||||||
| Ferulic (FA) cinnamic (CA) acids | ↓2- to 2.5-fold CA, FA | [70] | |||||||
| Chlorogenic acid | ↓2- to 2.5-fold | [70] | |||||||
| Gallic acid (GA) Ferulic acid (FA) | ↓84% GA ↓ 81% FA | ¯100% GA, 0% FA | ↓42% GA, FA (1 mg/mL) | ↓42% GA, 47% FA (1 mg/mL) | ↓42% GA, 8% FA (1 mg/mL) | [71] | |||
| Salicylic acid (SA) cinnamaldehyde (CIN) | ¯26% CIN ¯54% SA | [72] | |||||||
| 6-gingerol | ↓19–53% (0.1–100 µM) | [73] | |||||||
| Zingerone | ↓ | ↓ | ↓51.3% (10 mg/mL) | ↓53% (10 mg/mL) | 53% (10 mg/mL) | [74] | |||
| Zingerone | ↓ 50% reduction of biofilm | ↓55% (10 µg/mL) | ↓68% (10 µg/mL) | ↓67% (10 µg/mL) | [75] | ||||
| Proanthocyanidin monomer A-type | ↓40.9% (1 μg/mL) | ↓36.9% (10 μg/mL) | ↓54.1% at 10 μg/mL | ↓100 μg/mL | [76] | ||||
| Compounds | VIRULENCE FACTORS Regulated by QS | REF. | |||||||
|---|---|---|---|---|---|---|---|---|---|
| AHLs Levels | Alginate Assay | Elastase Assay | Hemolysin Assay | Protease Assay | Pyocyanin Assay | Pyoverdine Secretion Assay | Rhamno-Lipid | ||
| Plumbagin | ↓40% (150 µg/mL) | ↓>80% (150 µg/mL) | ↓ (150 µg/mL) | [65] | |||||
| Cinnamic acid | ↓21.8 (250 µg/mL) | ↓49.9 (250 µg/mL) | ↓ 80.9 (250 µg/mL) | ↓71.4 CA (250 µg/mL) | ↓16.5 (250 µg/mL) | [67] | |||
| Ginkgolic Acid | ↓90% | [68] | |||||||
| Curcumin | ↓25% in 3-oxoC12-HSL, ↓>2% C4-HSL (1 µg /mL) | ↓2-fold (3–5 µg/mL) | ↓2-fold vs ctr (3–5 µg/mL) | ↓60–80% (1.5–3 µg/mL) | [63] | ||||
| Eugenol | ↓32 and 46% (200and 400 µM) | ↓56% at 50 µM | [69] | ||||||
| Salicylic acid (SA) cinnamaldehyde (CIN) | ↓22% CIN, ↓28% SA | ↓65% CIN, ↓31% SA | ↓32% CIN ¯70% SA | significant ↓ | [72] | ||||
| 6-gingerol | ↓ (0.1–1 mM) | ↓21–43% (1, 10, and 100 mM) | ↓36–60% (1, 10, and 100 mM) | ↓36–60% (1, 10, and 100 mM) | [73] | ||||
| Zingerone | ↓ C4-HSL, OdDHL, | Marked ↓ | Marked ↓ | ↓Significant | ↓ Significant | Marked ↓ | [75] | ||
| Proanthocyanidin monomer A-type | [76] | ||||||||
| Compounds | BIOFILM ASSAY | VIRULENCE FACTORS Regulated by QS | Ref. | |||
|---|---|---|---|---|---|---|
| Cristal Violet (CV) or Safrin (S) Staining | Metabolic Activity of Biofilm-Forming by MTT Assay | EPS Production | Pre-Formed Biofilms | Hemolysin | ||
| Chelerythrine (CH) Sanguinarine (SA) | ↓SA (24.5 μM) ↓CH (15.2 μM) | [86] | ||||
| Celastrol | ↓25.5–85.07% ATCC 29213 ↓27–89.3% MRSA (40 μmol/L) | ↓40.8–76.0% ATCC 29213 ↓42.0–51.1% MRSA (1.25 μmol/L) | ↓40.5–80.2% ATCC 29213 ↓49.5–82.8% MRSA (40 μmol/L) | [84] | ||
| Carvacrol | ↓(0.50 to 1 mM) | ↓(8 mM) | [80] | |||
| 1,8-Cineole | ↓(0.095 mg/mL) | ↓77.46 ± 1.91%–90.81 ± 4.05% (0.048, 0.096, 0.192 mg/mL) | [79] | |||
| alpha-Cyperone | ↓6.3% ATCC 29213; 4.4% BAA-1717; 12.6% Wood 46; 6.1% 83254 (16 μg/mL). | [87] | ||||
| Ursolic acid | ↓66.3% (30 µg/mL) | [85] | ||||
| Bacoside A | ↓90% (200 µg/mL) | ↓10% (200 µg/mL) | ↓ | [43] | ||
| Compounds | BIOFILM ASSAY | VIRULENCE FACTORS Regulated by QS | Ref. | ||
|---|---|---|---|---|---|
| Total Protein Content | Metabolic Activity of Biofilm-Forming by MTT Assay | Hemolysin | Protease | ||
| Myricetin | ↓(200 µg/mL) | ↓(200 µg/mL) | [89] | ||
| Farrerol | ↓(0.5 μg/µg/mL) | ↓(0.5 μg/µg/mL) | [91] | ||
| Isorhamnetin | ↓(16 µg/mL) | [92] | |||
| Lysionotin | ↓(8 μg/mL) | [93] | |||
| Diosmetin | ↓(32 µg/mL) | [94] | |||
| 5,6,7,3′,4′-Pentahydroxyflavone | ↓(80 μg/disc) | [102] | |||
| 3′,4′,5-trihydroxy-6,7-dimethoxy-flavone | ↓(80 μg/disc) | [102] | |||
| Phloretin | ↓(16 µg/mL) | [104] | |||
| Apigenin | ↓(8 µg/mL) | [95] | |||
| Epigallocatechin gallate | ↓96.6% (32 µg/mL) | [96] | |||
| Baicalin | ↓(16 µg/mL) | [97] | |||
| Oroxylin A | ↓(8 µg/mL) | [98] | |||
| Quercetin | ↓(16 µg/mL) | [90] | |||
| Dracorhodin Perochlorate | ↓(16 µg/mL) | [99] | |||
| Silibinin | ↓(32 µg/mL) | [100] | |||
| Baicalein | ↓(32 µg/mL, 64 µg/mL) | ↓(32 µg/mL, 64 µg/mL) | [88] | ||
| Baicalin | ↓(16 μg/mL Baicalin with Osthole) | [105] | |||
| Rutin | ↓19–88% MRSA ↓24–58%, 19–77%, 63–88% NSA-02,-06,-08 (75–600 μg/mL) | ↓18–65%, 39–90%, 58–92% NSA-02,-06,-08 (75–600 µg/mL) | [101] | ||
| Chalcone | ↓(76 μg/mL) | ↓(38 μg/mL) | [106] | ||
| Compounds | BIOFILM ASSAY | VIRULENCE FACTORS Regulated by QS | Ref. | |||
|---|---|---|---|---|---|---|
| Total Protein Content | Cristal Violet (CV) Staining | Metabolic Activity of Biofilm-Forming by MTT Assay | EPS Production | Hemolysin | ||
| Gallic acid (GA) Ferulic acid (FA) | ↓90% GA ↓7% FA | ↓70% GA ↓6% FA | [71] | |||
| Eugenyl acetate | ↓(150 µg/mL) | [81] | ||||
| Resveratrol (Res) | ↓39.8% (100 µg/mL) | ↓55.4% Res + Van ↓23.4% Res | ↓ (64 µg/mL) | [85] | ||
| Dihydroxybenzofurane (DHBF);Pro-antocyanidin A2 (proAc) | ↓DHBF (8.2 μM) ↓proAc (6.9 μM) | [86] | ||||
| Dihydroxybenzofurane (DHBF);Pro-antocyanidin A2 (proAc) | ↓DHBF (8.2 μM) ↓proAc (6.9 μM) | [86] | ||||
| Curcumin | ↓(16 μg/mL) | [108] | ||||
| Osthole | ↓(at 16 μg/mL, alone and with Baicalin) | [105] | ||||
| Brazilin | ↓(32 μg/mL) | [109] | ||||
| Punicagalin | ↓47% (3.9 μg/mL); ↓90% (7.8 μg/mL). | ↓(0.125 mg/mL) | [112,113] | |||
| 1,2,3,4,6-Penta-O-galloyl- -D-glucopyranose (PGG) | ↓93%, 96%, 97% (6.25, 12.5, 25 μM); ↓83%, 97% (50 μM) | ↓7%, 58%, 87% (3.13, 12.5, 50 μM) | [115] | |||
| Rhein (Rhe) and Aloeemodin (Alo) | ↓20.0% Rhe; ↓33.3% Alo | [117] | ||||
| Hamamelitannin | ↓(50 µg/mL) | [47,56] | ||||
| Rhodomyrtone | ↓(0.125–1 μg/mL) | ↓(4–16 μg/mL) | [118] | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guzzo, F.; Scognamiglio, M.; Fiorentino, A.; Buommino, E.; D’Abrosca, B. Plant Derived Natural Products against Pseudomonas aeruginosa and Staphylococcus aureus: Antibiofilm Activity and Molecular Mechanisms. Molecules 2020, 25, 5024. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25215024
Guzzo F, Scognamiglio M, Fiorentino A, Buommino E, D’Abrosca B. Plant Derived Natural Products against Pseudomonas aeruginosa and Staphylococcus aureus: Antibiofilm Activity and Molecular Mechanisms. Molecules. 2020; 25(21):5024. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25215024
Chicago/Turabian StyleGuzzo, Francesca, Monica Scognamiglio, Antonio Fiorentino, Elisabetta Buommino, and Brigida D’Abrosca. 2020. "Plant Derived Natural Products against Pseudomonas aeruginosa and Staphylococcus aureus: Antibiofilm Activity and Molecular Mechanisms" Molecules 25, no. 21: 5024. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25215024