
Health-Promoting Compounds in Stevia: The Effect of Mycorrhizal Symbiosis, Phosphorus Supply and Harvest Time


Abstract
:1. Introduction
2. Results
2.1. Secondary Metabolites and Antioxidant Activities
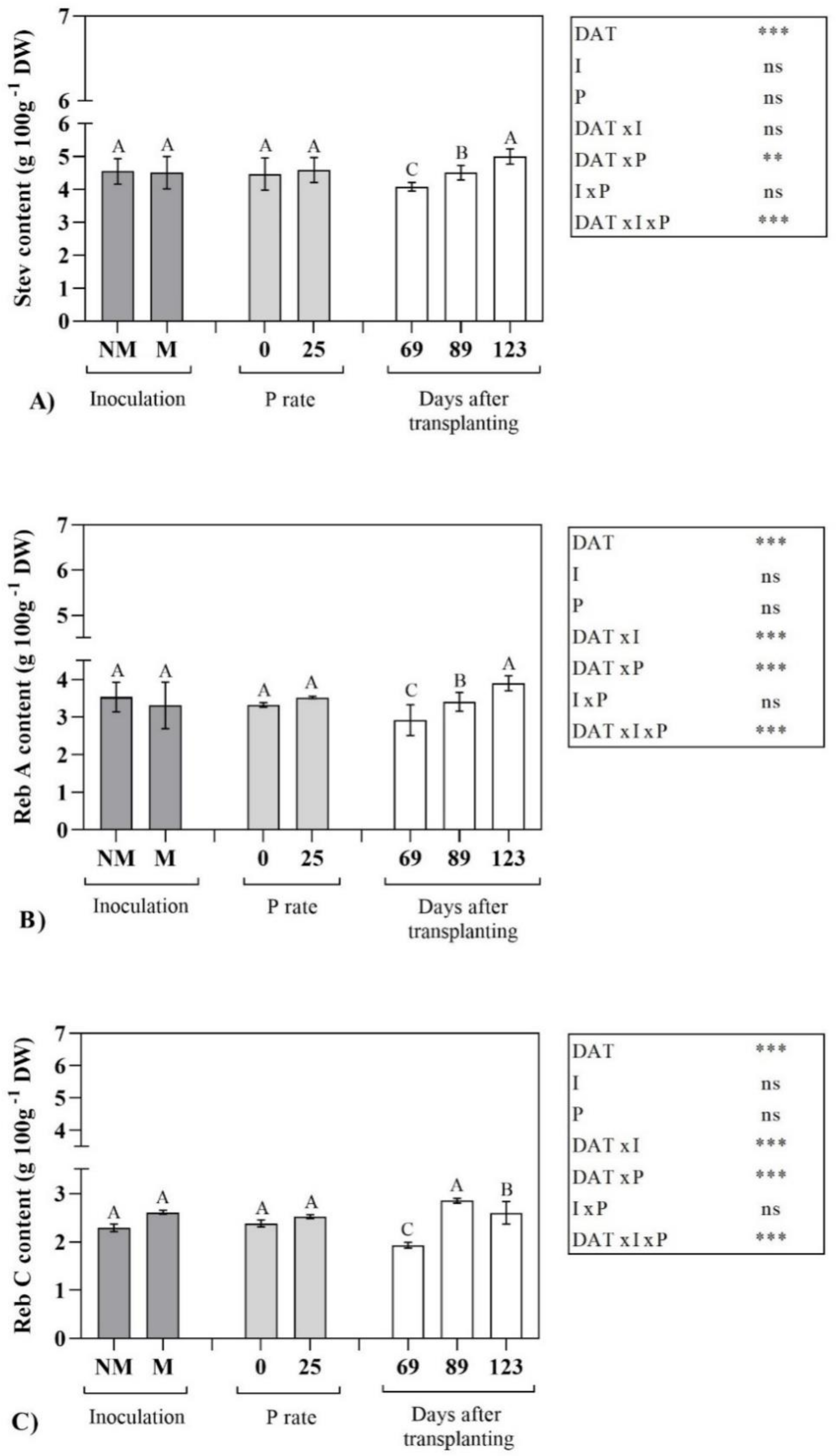
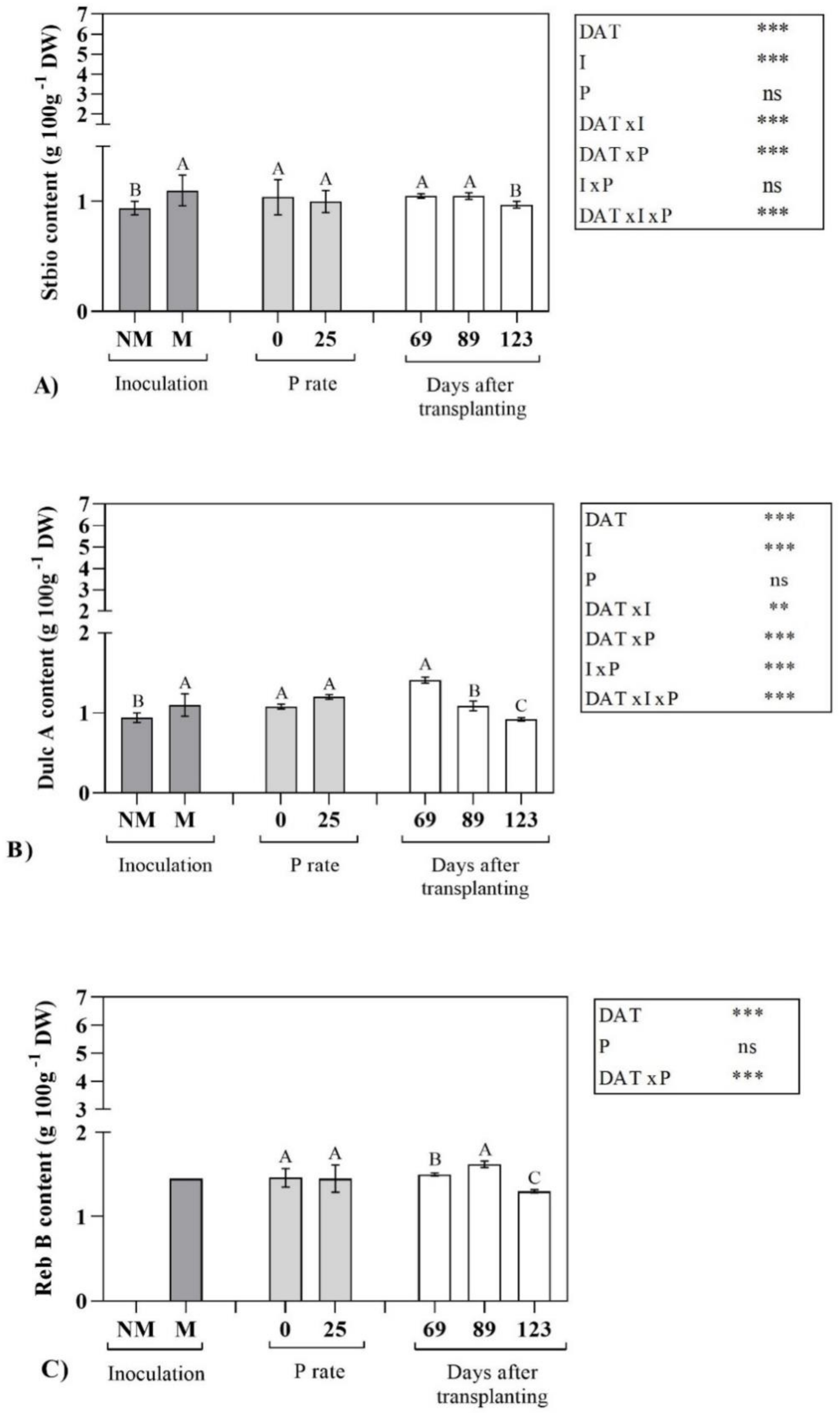
2.2. Steviol Glycoside Profile
2.3. Effects of Bioactive Compounds on Antioxidant Activities
3. Discussion
3.1. The role of Variability Factors on Stevia Bioactive Compounds
3.2. The Role of Variability Factors on Antioxidant Activities
3.3. Antioxidant Activities and Secondary Metabolites Correlation
4. Materials and Methods
4.1. Chemicals
4.2. Plant Material and Experimental Conditions
4.3. AMF Inoculum
4.4. Steviol glycosides determination
4.5. Preparation of Extract
4.6. Analysis of Total Phenols and Flavonoids
4.7. Ferric Reducing Antioxidant Power (FRAP) Assay
4.8. Oxygen Radical Absorbance Capacity (ORAC) Assay
4.9. Cellular Antioxidant Activity (CAA) Assay
4.10. Free Radical-Scavenging Assay
4.11. Specific Antioxidant Capacity (SAC)
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lemus-Mondaca, R.; Vega-Gálvez, A.; Zura-Bravo, L.; Ah-Hen, K. Stevia rebaudiana Bertoni, Source of a High-Potency Natural Sweetener: A Comprehensive Review on the Biochemical, Nutritional and Functional Aspects. Food Chem. 2012, 132, 1121–1132. [Google Scholar] [CrossRef] [PubMed]
- Wölwer-Rieck, U. The Leaves of Stevia rebaudiana (Bertoni), Their Constituents and the Analyses Thereof: A Review. J. Agric. Food Chem. 2012, 60, 886–895. [Google Scholar] [CrossRef]
- Prakash, I.; Markosyan, A.; Bunders, C. Development of Next Generation Stevia Sweetener: Rebaudioside, M. Foods 2014, 3, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Gaweł-Bęben, K.; Bujak, T.; Nizioł-Łukaszewska, Z.; Antosiewicz, B.; Jakubczyk, A.; Karaś, M.; Rybczyńska-Tkaczyk, K. Stevia rebaudiana Bert. Leaf Extracts as a Multifunctional Source of Natural Antioxidants. Molecules 2015, 20, 5468–5486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Ruiz, J.C.; Moguel-Ordoñez, Y.B.; Segura-Campos, M.R. Biological Activity of Stevia rebaudiana Bertoni and Their Relationship to Health. Crit. Rev. Food Sci. Nutr. 2017, 57, 2680–2690. [Google Scholar] [CrossRef]
- Taleie, N.; Hamidoghli, Y.; Rabiei, B.; Hamidoghli, S. Effects of Plant Density and Transplanting Date on Herbage, Stevioside, Phenol and Flavonoid Yield of Stevia rebaudiana Bertoni. IJACS 2012, 4, 298–302. [Google Scholar]
- Tavarini, S.; Angelini, L.G. Stevia rebaudiana Bertoni As a Source of Bioactive Compounds: The Effect of Harvest Time, Experimental Site and Crop Age on Steviol Glycoside Content and Antioxidant Properties. J. Sci. Food Agric. 2013, 93, 2121–2129. [Google Scholar] [CrossRef]
- Sharma, S.; Walia, S.; Singh, B.; Kumar, R. Comprehensive Review on Agro Technologies of Low-Calorie Natural Sweetener Stevia (Stevia rebaudiana Bertoni): A Boon to Diabetic Patients. J. Sci. Food Agric. 2015, 96, 1867–1879. [Google Scholar] [CrossRef]
- Tavarini, S.; Sgherri, C.; Ranieri, A.M.; Angelini, L.G. Effect of Nitrogen Fertilization and Harvest Time on Steviol Glycosides, Flavonoid Composition, and Antioxidant Properties in Stevia rebaudiana Bertoni. J. Agric. Food Chem. 2015, 63, 7041–7050. [Google Scholar] [CrossRef]
- Angelini, L.G.; Martini, A.; Passera, B.; Tavarini, S. Cultivation of Stevia rebaudiana Bertoni and Associated Challenges. In Reference Series in Phytochemistry; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2018; pp. 35–85. [Google Scholar] [CrossRef]
- Demain, A.L.; Fang, A. The Natural Functions of Secondary Metabolites. In Process Integration in Biochemical Engineering; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2000; Volume 69, pp. 1–39. [Google Scholar] [CrossRef]
- Avio, L.; Turrini, A.; Giovannetti, M.; Sbrana, C. Designing the Ideotype Mycorrhizal Symbionts for the Production of Health-Promoting Foods. Front. Plant Sci. 2018, 9, 1–19. [Google Scholar] [CrossRef]
- Schüβler, A.; Schwarzott, D.; Walker, C. A New Fungal Phylum, the Glomeromycota: Phylogeny and Evolution. Mycol. Res. 2001, 105, 1413–1421. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.E.; Jakobsen, I.; Grønlund, M.; Smith, F.A. Roles of Arbuscular Mycorrhizas in Plant Phosphorus Nutrition: Interactions Between Pathways of Phosphorus Uptake in Arbuscular Mycorrhizal Roots Have Important Implications for Understanding and Manipulating Plant Phosphorus Acquisition. Plant Physiol. 2011, 156, 1050–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sbrana, C.; Avio, L.; Giovannetti, M. Beneficial Mycorrhizal Symbionts Affecting the Production of Health-Promoting Phytochemicals. Electrophoresis 2014, 35, 1535–1546. [Google Scholar] [CrossRef] [PubMed]
- Tavarini, S.; Passera, B.; Martini, A.; Avio, L.; Sbrana, C.; Giovannetti, M.; Angelini, L.G. Plant Growth, Steviol Glycosides and Nutrient Uptake As Affected by Arbuscular Mycorrhizal Fungi and Phosphorous Fertilization in Stevia rebaudiana Bert. Ind. Crop. Prod. 2018, 111, 899–907. [Google Scholar] [CrossRef]
- Mandal, S.; Evelin, H.; Giri, B.; Singh, V.P.; Kapoor, R. Arbuscular Mycorrhiza Enhances the Production of Stevioside and Rebaudioside A in Stevia rebaudiana via Nutritional and Non-Nutritional Mechanisms. Appl. Soil Ecol. 2013, 72, 187–194. [Google Scholar] [CrossRef]
- Mandal, S.; Upadhyay, S.; Singh, V.P.; Kapoor, R. Enhanced Production of Steviol Glycosides in Mycorrhizal Plants: A Concerted Effect of Arbuscular Mycorrhizal Symbiosis on Transcription of Biosynthetic Genes. Plant Physiol. Biochem. 2015, 89, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants; Elsevier BV: Amsterdam, The Netherlands, 2002. [Google Scholar] [CrossRef]
- Nell, M.; Vötsch, M.; Vierheilig, H.; Steinkellner, S.; Zitterl-Eglseer, K.; Franz, C.; Novak, J. Effect of Phosphorus Uptake on Growth and Secondary Metabolites of Garden Sage (Salvia officinalis, L.). J. Sci. Food Agric. 2009, 89, 1090–1096. [Google Scholar] [CrossRef]
- Ceunen, S.; Geuns, J.M.C. Steviol Glycosides: Chemical Diversity, Metabolism, and Function. J. Nat. Prod. 2013, 76, 1201–1228. [Google Scholar] [CrossRef]
- Singh, G.; Singh, G.; Singh, P.; Parmar, R.; Paul, N.; Vashist, R.; Swarnkar, M.K.; Kumar, A.; Singh, S.; Singh, A.K.; et al. Molecular Dissection of Transcriptional Reprogramming of Steviol Glycosides Synthesis in Leaf Tissue During Developmental Phase Transitions in Stevia rebaudiana Bert. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Bondarev, N.; Sukhanova, M.; Reshetnyak, O.; Nosov, A. Steviol Glycoside Content in Different Organs of Stevia rebaudiana and Its Dynamics During Ontogeny. Biol. Plant. 2003, 46, 261–264. [Google Scholar] [CrossRef]
- Christaki, E.; Bonos, E.; Giannenas, I.; Florou-Paneri, P. Aromatic Plants as a Source of Bioactive Compounds. Agriculture 2012, 2, 228–243. [Google Scholar] [CrossRef] [Green Version]
- Rivera, T.; Oesterling, J. Naturally Sweetened Juice Beverage Products. US Patent US9788562B2, 17 October 2017. [Google Scholar]
- Letenneur, L.; Proust-Lima, C.; Le Gouge, A.; Dartigues, J.F.; Barberger-Gateau, P. Flavonoid Intake and Cognitive Decline over a 10-Year Period. Am. J. Epidemiol. 2007, 165, 1364–1371. [Google Scholar] [CrossRef] [PubMed]
- Carocho, M.; Ferreira, I.C. A Review on Antioxidants, Prooxidants and Related Controversy: Natural and Synthetic Compounds, Screening and Analysis Methodologies and Future Perspectives. Food Chem. Toxicol. 2013, 51, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Tungmunnithum, D.; Thongboonyou, A.; Pholboon, A.; Yangsabai, A. Flavonoids and Other Phenolic Compounds from Medicinal Plants for Pharmaceutical and Medical Aspects: An Overview. Medicines 2018, 5, 93. [Google Scholar] [CrossRef]
- Bender, C.; Graziano, S.; Zimmermann, B.F. Study of Stevia rebaudiana Bertoni Antioxidant Activities and Cellular Properties. Int. J. Food Sci. Nutr. 2015, 66, 553–558. [Google Scholar] [CrossRef]
- Carlsen, M.H.; Halvorsen, B.L.; Holte, K.; Bøhn, S.K.; Dragland, S.; Sampson, L.; Willey, C.; Senoo, H.; Umezono, Y.; Sanada, C.; et al. The Total Antioxidant Content of More Than 3100 Foods, Beverages, Spices, Herbs and Supplements Used Worldwide. Nutr. J. 2010, 9, 3. [Google Scholar] [CrossRef]
- Haytowitz, D.B.; Bhagwat, S.A. USDA Database for the Oxygen Radical Capacity (ORAC) of Selected Foods, Release 2. USDA National Nutrient Database for Standard Reference. 2010. Available online: http://www.ars.usda.gov/nutrientdata (accessed on 9 November 2020).
- Yesil-Celiktas, O.; Girgin, G.; Orhan, H.; Wichers, H.J.; Bedir, E.; Vardar-Sukan, F. Screening of Free Radical Scavenging Capacity and Antioxidant Activities of Rosmarinus officinalis Extracts with Focus on Location and Harvesting Times. Eur. Food Res. Technol. 2006, 224, 443–451. [Google Scholar] [CrossRef]
- Duda, S.C.; Mărghitaş, L.A.; Dezmirean, D.S.; Duda, M.; Mărgăoan, R.; Bobiş, O. Changes in Major Bioactive Compounds with Antioxidant Activity of Agastache foeniculum, Lavandula angustifolia, Melissa officinalis and Nepeta cataria: Effect of Harvest Time and Plant Species. Ind. Crop. Prod. 2015, 77, 499–507. [Google Scholar] [CrossRef]
- Liu, S.-C.; Lin, J.-T.; Hu, C.-C.; Shen, B.-Y.; Chen, T.-Y.; Chang, Y.-L.; Shih, C.-H.; Yang, D.-J. Phenolic Compositions and Antioxidant Attributes of Leaves and Stems from Three Inbred Varieties of Lycium chinense Miller Harvested at Various Times. Food Chem. 2017, 215, 284–291. [Google Scholar] [CrossRef]
- Bender, C. Evaluation of Stevia rebaudiana Bertoni Antioxidant Properties at Cellular Level, Cytotoxicity and Proliferative Effects. In Stevia Growth in Knowledge and Taste; B-3001; Euprint: Heverlee, Belgium, 2015; pp. 117–122. ISBN 978-90-742-53291. [Google Scholar]
- Wolfe, K.L.; Kang, X.; He, X.; Dong, M.; Zhang, Q.; Liu, R.H. Cellular Antioxidant Activity of Common Fruits. J. Agric. Food Chem. 2008, 56, 8418–8426. [Google Scholar] [CrossRef]
- Song, W.; DeRito, C.M.; Liu, M.K.; He, X.; Dong, M.; Liu, R.H. Cellular Antioxidant Activity of Common Vegetables. J. Agric. Food Chem. 2010, 58, 6621–6629. [Google Scholar] [CrossRef] [PubMed]
- Bender, C. Stevia Rebaudiana’s Antioxidant Properties; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2018; pp. 349–375. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J.; Paganga, G. Structure-Antioxidant Activity Relationships of Flavonoids and Phenolic Acids. Free. Radic. Biol. Med. 1996, 20, 933–956. [Google Scholar] [CrossRef]
- Reyes, L.F.; Miller, J.C.; Cisneros-Zevallos, L. Antioxidant Capacity, Anthocyanins and Total Phenolics in Purple-and Red-Fleshed Potato (Solanum tuberosum, L.) Genotypes. Am. J. Potato Res. 2005, 82, 271–277. [Google Scholar] [CrossRef]
- Jacobo-Velázquez, D.; Cisneros-Zevallos, L. Correlations of Antioxidant Activity Against Phenolic Content Revisited: A New Approach in Data Analysis for Food and Medicinal Plants. J. Food Sci. 2009, 74, 107–113. [Google Scholar] [CrossRef]
- Reyes, L.F.; Cisneros-Zevallos, L. Wounding Stress Increases the Phenolic Content and Antioxidant Capacity of Purple-Flesh Potatoes (Solanum tuberosum L.). J. Agric. Food Chem. 2003, 51, 5296–5300. [Google Scholar] [CrossRef]
- Heredia, J.B.; Cisneros-Zevallos, L. The Effect of Exogenous Ethylene and Methyl Jasmonate on Pal Activity, Phenolic Profiles and Antioxidant Capacity of Carrots (Daucus carota) under Different Wounding Intensities. Postharvest Biol. Technol. 2009, 51, 242–249. [Google Scholar] [CrossRef]
- Vizzotto, M.; Cisneros-Zevallos, L.; Byrne, D.H.; Ramming, D.W.; Okie, W. Large Variation Found in the Phytochemical and Antioxidant Activity of Peach and Plum Germplasm. J. Am. Soc. Hortic. Sci. 2007, 132, 334–340. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, K.L.; Liu, R.H. Structure−Activity Relationships of Flavonoids in the Cellular Antioxidant Activity Assay. J. Agric. Food Chem. 2008, 56, 8404–8411. [Google Scholar] [CrossRef]
- Wolfe, K.L.; Liu, R.H. Cellular Antioxidant Activity (CAA) Assay for Assessing Antioxidants, Foods, and Dietary Supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef]
- Eberhardt, M.V.; Kobira, K.; Sigrid-Keck, A.; Juvik, J.A.; Jeffery, E. Correlation Analyses of Phytochemical Composition, Chemical, and Cellular Measures of Antioxidant Activity of Broccoli (Brassica oleracea L. Var. Italica). J. Agric. Food Chem. 2005, 53, 7421–7431. [Google Scholar] [CrossRef]
- Bender, C.; Graziano, S.; Zimmermann, B.F.; Weidlich, H.H. Antioxidant Potential of Aqueous Plant Extracts Assessed by the Cellular Antioxidant Activity Assay. Am. J. Biol. Life Sci. 2014, 2, 72–79. [Google Scholar]
- Le Bihan, Z.; Cosson, P.; Rolin, D.; Schurdi-Levraud, V. Phenological Growth Stages of Stevia (Stevia rebaudiana Bertoni) According to the Biologische Bundesanstalt Bundessortenamt and Chemical Industry (BBCH) Scale. Ann. Appl. Biol. 2020, 1–13. [Google Scholar] [CrossRef]
- Wölwer-Rieck, U.; Tomberg, W.; Wawrzun, A. Investigations on the Stability of Stevioside and Rebaudioside A in Soft Drinks. J. Agric. Food Chem. 2010, 58, 12216–12220. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Jia, Z.; Mengcheng, T.; Jianming, W. The Determination of Flavonoid Contents in Mulberry and Their Scavenging Effects on Superoxide Radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J. Ferric reducing/Antioxidant Power Assay: Direct Measure of Total Antioxidant Activity of Biological Fluids and Modified Version for Simultaneous Measurement of Total Antioxidant Power and Ascorbic Acid Concentration. Methods Enzymol. 1999, 299, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Alessio, H.M.; Cutler, R.G. Oxygen-Radical Absorbance Capacity Assay for Antioxidants. Free. Radic. Biol. Med. 1993, 14, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized Methods for the Determination of Antioxidant Capacity and Phenolics in Foods and Dietary Supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef] [PubMed]
- Brand-Williams, W.; Cuvelier, M.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT 1995, 28, 25–30. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Factor | Factor Level | TPC (mg GAE/g DW) | TFC (mg CE/g DW) | FRAP (mmol TE/g DW) | ORAC (µmol TE/g DW) | CAA (µmol QE/g DW) | DPPH (mmol TE/g DW) | Total SVglys (g/100g DW) |
|---|---|---|---|---|---|---|---|---|
| Main Effects | ||||||||
| Mychorizzal inoculation (I) | NMP | 81.91 ± 3.00 a | 71.13 ± 2.39 a | 0.560 ± 0.02 a | 2231.19 ± 106.2 a | 11.28 ± 0.77 a | 0.161 ± 0.002 a | 12.95 ± 0.30 b |
| MP | 74.00 ± 3.77 b | 68.92 ± 0.99 b | 0.553 ± 0.03 a | 1417.01 ± 90.30 b | 9.14 ± 0.73 b | 0.163 ± 0.002 a | 14.37 ± 0.57 a | |
| Phosphorus fertilization (P) | 0P | 74.88 ± 3.20 b | 68.29 ± 2.10 b | 0.532 ± 0.03 b | 1593.51 ± 113.5 b | 10.67 ± 0.77 a | 0.161 ± 0.001 a | 13.79 ± 1.66 a |
| 25P | 81.04 ± 3.72 a | 71.76 ± 1.44 a | 0.581 ± 0.02 a | 2054.67 ± 139.9 a | 9.71 ± 0.78 b | 0.162 ± 0.002 a | 13.52 ± 0.16 a | |
| Days after transplanting (DAT) | 69 DAT | 87.06 ± 1.30 a | 63.95 ± 1.68 c | 0.591 ± 0.02 a | 1636.41 ± 183.2 c | 10.37 ± 1.03 b | 0.155 ± 0.001 c | 12.18 ± 0.19 c |
| 89 DAT | 87.64 ± 1.38 a | 69.76 ± 0.90 b | 0.609 ± 0.03 a | 1953.47 ± 75.21 a | 12.36 ± 0.33 a | 0.168 ± 0.002 a | 14.80 ± 0.80 a | |
| 123 DAT | 59.17 ± 1.89 b | 76.37 ± 1.51 a | 0.470 ± 0.01 b | 1882.41 ± 129.4 b | 7.66 ± 0.26 c | 0.162 ± 0.002 b | 14.00 ± 0.24 b | |
| Significance | ||||||||
| I | *** | * | ns | *** | *** | ns | *** | |
| P | *** | *** | *** | *** | * | ns | ns | |
| DAT | *** | *** | *** | *** | *** | *** | *** | |
| I x P | * | *** | *** | *** | ns | ns | ** | |
| I x DAT | *** | *** | *** | *** | *** | * | *** | |
| P x DAT | ns | *** | *** | *** | *** | ns | *** | |
| I x P x DAT | ** | ns | *** | *** | ns | ns | *** |
| Assay | TPC | TFC | FRAP | ORAC | CAA | DPPH | Total SVglys |
|---|---|---|---|---|---|---|---|
| TPC | 1 | 0.110 * y= −0.156x + 81.53 | 0.268 ** y= 0.004x + 0.260 | 0.050 n.s. | 0.187 * y= 0.097x + 2.638 | 0.038 n.s. | 0.035 n.s. |
| TFC | 0.127 * y= −0.706x + 127.8 | 1 | 0.003 n.s. | 0.169 * y = 32.51x − 449.9 | 0.244 ** y = −0.228x + 26.08 | 0.014 n.s. | 0.226 * y = 0.134x + 4.331 |
| FRAP | 0.268 ** y = 74.24x + 37.71 | 0.004 n.s. | 1 | 0.230 ** y = 2695x + 362.8 | 0.202 * y = 14.21x + 2.460 | 0,141 * y = −5.805x + 1,483 | 0.212 * y = 8.847x + 8.868 |
| ORAC | 0.044 n.s. | 0.169 * y = 0.005x + 60.23 | 0.230 * y = 8.55 x 10-4 + 0.386 | 1 | 0.065 n.s. | 0,1767 * y = −36485x + 7739 | 0.013 n.s. |
| CAA | 0.187 * y = 1.951x + 58.47 | 0.244 ** y = −1.063x + 80.26 | 0.202 * y = 0.014x + 0.399 | 0.065 n.s. | 1 | 0,181 * y = −207.7x + 43,84 | 0.008 n.s. |
| DPPH | 0.038 n.s. | 0.014 n.s. | 0,141 * y = -5.805x + 1.483 | 0,1767 * y = −36485x + 7739 | 0,181 * y = −207.7x + 43.84 | 1 | 0.001 n.s. |
| Total SVglys | 0.035 n.s. | 0.226 * y = 1.687x + 46.69 | 0.212 * y = 0.024x + 0.214 | 0.013 n.s. | 0.007 n.s. | 0.001 n.s. | 1 |
| Specific Antioxidant Capacity | |
|---|---|
| FRAP/TPC (µmol TE/mg GAE) | 7.13 |
| CAA/TPC (µmol QE/mg GAE) | 0.130 |
| ORAC/TFC (µmol TE/mg CE) | 26.04 |
| FRAP/SVglys (µmol TE/mg DW) | 4.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tavarini, S.; Clemente, C.; Bender, C.; Angelini, L.G. Health-Promoting Compounds in Stevia: The Effect of Mycorrhizal Symbiosis, Phosphorus Supply and Harvest Time. Molecules 2020, 25, 5399. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25225399
Tavarini S, Clemente C, Bender C, Angelini LG. Health-Promoting Compounds in Stevia: The Effect of Mycorrhizal Symbiosis, Phosphorus Supply and Harvest Time. Molecules. 2020; 25(22):5399. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25225399
Chicago/Turabian StyleTavarini, Silvia, Clarissa Clemente, Cecilia Bender, and Luciana G. Angelini. 2020. "Health-Promoting Compounds in Stevia: The Effect of Mycorrhizal Symbiosis, Phosphorus Supply and Harvest Time" Molecules 25, no. 22: 5399. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25225399