
Regulation of Selenium/Sulfur Interactions to Enhance Chemopreventive Effects: Lessons to Learn from Brassicaceae



Abstract
:1. Introduction
2. The Distribution of Selenium and Sulfur Compounds in Brassicaceae and Their Chemopreventive Potency
3. Selenium-to-Sulfur Ratios and Their Accumulation in Brassica Vegetables
4. Reported Studies about the Impact of Se or Se/S Biofortification on Chemopreventive Inhibitory Effects in Brassica Vegetables
5. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- World Health Organization. Global Health Observatory; World Health Organization: Geneva, Switzerland, 2018; Available online: who.int/gho/database/en/ (accessed on 18 July 2020).
- Shamberger, R.J.; Tytko, S.A.; Willis, C.E. Antioxidants and cancer. VI. Selenium and age-adjusted human cancer mortality. Arch. Environ. Health 1976, 31, 231–235. [Google Scholar] [CrossRef]
- Clark, L.C.; Cantor, K.; Allaway, W.H. Selenium in forage crops and cancer mortality in US counties. Arch. Environ. Health 1991, 46, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Salonen, J.T.; Salonen, R.; Lappeteläinen, R.; Mäenpää, P.H.; Alfthan, G.; Puska, P. Risk of cancer in relation to serum concentrations of selenium and vitamins A and E: Matched case-control analysis of prospective data. Br. Med. J. 1985, 290, 417–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, L.C.; Hixson, L.J.; Combs, G.F., Jr.; Reid, M.E.; Turnbull, B.W.; Sampliner, R.E. Plasma selenium concentration predicts the prevalence of colorectal adenomatous polyps. Cancer Epidemiol. Biomark. Prev. 1993, 2, 41–46. [Google Scholar]
- Luo, C.; Sun, J.; Liu, D.; Sun, B.; Miao, L.; Musetti, S.; Li, J.; Han, X.; Du, Y.; Huang, L.; et al. Self-assembled redox dual-responsive prodrug-nanosystem formed by single thioether-bridged paclitaxel-fatty acid conjugate for cancer chemotherapy. Nano Lett. 2016, 16, 5401–5408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Sun, X.; Mao, W.; Sun, W.; Tang, J.; Sui, M.; Shen, Y.; Gu, Z. Tumor redox heterogeneity-responsive prodrug nanocapsules for cancer chemotherapy. Adv. Mater. 2013, 25, 3670–3676. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Sun, J.; Sun, B.; Liu, D.; Miao, L.; Goodwin, T.J.; Huang, L.; He, Z. Facile fabrication of tumor redox-sensitive nanoassemblies of small-molecule oleate prodrug as potent chemotherapeutic nanomedicine. Small 2016, 12, 6353–6362. [Google Scholar] [CrossRef]
- Sun, B.; Luo, C.; Yu, H.; Zhang, X.; Chen, Q.; Yang, W.; Wang, M.; Kan, Q.; Zhang, H.; Wang, Y.; et al. Disulfide bond-driven oxidation- and reduction-responsive prodrug nanoassemblies for cancer therapy. Nano Lett. 2018, 18, 3643–3650. [Google Scholar] [CrossRef]
- Kieliszek, M.; Lipinski, B.; Blazejak, S. Application of sodium selenite in the prevention and treatment of cancers. Cells 2017, 6, 39. [Google Scholar] [CrossRef] [Green Version]
- Cao, W.; Wang, L.; Xu, H. Selenium/tellurium containing polymer materials in nanobiotechnology. Nano Today 2015, 10, 717–736. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Cao, W.; Zhang, X. Selenium-containing polymers: Promising biomaterials for controlled release and enzyme mimics. Acc. Chem. Res. 2013, 46, 1647–1658. [Google Scholar] [CrossRef] [PubMed]
- Sors, T.G.; Ellis, D.R.; Salt, D.E. Selenium uptake, translocation, assimilation and metabolic fate in plants. Photosynth. Res. 2005, 86, 373–389. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, M.A.; Meschede, C.A.C.; Mühling, K.H. Selenium foliar application alters patterns of glucosinolate hydrolysis products of pak choi Brassica rapa L. var. Chinensis. Sci. Hortic. 2020, 273, 109614. [Google Scholar] [CrossRef]
- Anderson, J.W.; Scarf, A.R. Selenium and Plant Metabolism. In Metals and Micronutrients: Uptake and Utilization by Plants; Robb, D.A., Pierpoint, W.S., Eds.; Academic Press: New York, NY, USA, 1983; pp. 241–275. [Google Scholar]
- De Souza, M.P.; Pilon-Smits, E.A.; Lytle, C.M.; Hwang, S.; Tai, J.; Honma, T.S.; Yeh, L.; Terry, N. Rate-limiting steps in selenium assimilation and volatilization by indian mustard. Plant Physiol. 1998, 117, 1487–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdez-Barilla, J.R.; Quinn, C.F.; Pilon-Smits, E.A.H. Selenium accumulation in plants—Phytotechnological applications and ecological implications. Int. J. Phytoremediat. 2011, 13, 166–178. [Google Scholar] [CrossRef] [PubMed]
- Trippe, R.C.; Pilon-Smits, E.A.H. Selenium transport and metabolism in plants: Phytoremediation and biofortification implications. J. Hazard Mater. 2020, 404, 124178. [Google Scholar] [CrossRef] [PubMed]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular pathways and physiological roles. Physiol. Rev. 2014, 94, 739–777. [Google Scholar] [CrossRef] [Green Version]
- Brown, K.M.; Arthur, J.R. Selenium, selenoproteins and human health: A review. Public Health Nutr. 2001, 4, 593–599. [Google Scholar] [CrossRef] [Green Version]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Roman, M.; Jitaru, P.; Barbante, C. Selenium biochemistry and its role for human health. Metallomics 2014, 6, 25–54. [Google Scholar] [CrossRef]
- Fairweather-Tait, S.J.; Bao, Y.; Broadley, M.R.; Collings, R.; Ford, D.; Hesketh, J.E.; Hurst, R. Selenium in human health and disease. Antioxid Redox Sign 2011, 14, 1337–1383. [Google Scholar] [CrossRef] [PubMed]
- Mistry, H.D.; Pipkin, F.P.; Redman, C.W.G.; Poston, L. Selenium in reproductive health. Am. J. Obstet. Gynecol. 2012, 206, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvino, A.; Barbieri, G. Vegetables of Temperate Climates: Leafy Vegetables. In The Encyclopedia of Food and Health; Caballero, B., Finglas, P., Toldrá, F., Eds.; Academic Press: Oxford, UK, 2016; Volume 5, pp. 393–400. [Google Scholar]
- Ramirez, D.; Abellán-Victorio, A.; Beretta, V.; Alejandra Camargo, A.; Moreno, D.A. Functional ingredients from Brassicaceae species: Overview and perspective. Int. J. Mol. Sci. 2020, 21, 1998. [Google Scholar] [CrossRef] [Green Version]
- Abdalla, M.A.; Mühling, K.H. Plant-derived sulfur containing natural products produced as a response to biotic and abiotic stresses: A review of their structural diversity and medicinal importance. J. Appl. Bot. Food Qual. 2019, 92, 204–215. [Google Scholar]
- Bednarek, P. Sulfur-containing secondary metabolites from Arabidopsis thaliana and other Brassicaceae with function in plant immunity. Chembiochem 2012, 13, 1846–1859. [Google Scholar] [CrossRef]
- Meschede, C.A.C.; Abdalla, M.A.; Mühling, K.H. Sulfur but not nitrogen supply increases the ITC/Nitrile ratio in Pak Choi (Brassica rapa subsp. Chinensis (L.) Hanelt). J. Appl. Bot. Food Qual. 2020, 93, 95–104. [Google Scholar]
- Mitsiogianni, M.; Amery, T.; Franco, R.; Zoumpourlis, V.; Pappa, A.; Panayiotidis, M.I. From chemo-prevention to epigenetic regulation: The role of isothiocyanates in skin cancer prevention. Pharmacol. Ther. 2018, 190, 187–201. [Google Scholar] [CrossRef]
- Blažević, I.; Montaut, S.; Burčul, F.; Olsen, C.E.; Burow, M.; Rollin, P.; Agerbirk, N. Glucosinolate structural diversity, identification, chemical synthesis and metabolism in plants. Phytochemistry 2020, 169, 112100. [Google Scholar] [CrossRef]
- Veeranki, O.L.; Bhattacharya, A.; Tang, L.; Marshall, J.R.; Zhang, Y. Cruciferous vegetables, isothiocyanates, and prevention of bladder cancer. Curr. Pharmacol. Rep. 2015, 1, 272–282. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Tang, L. Discovery and development of sulforaphane as a cancer chemopreventive phytochemical. Acta Pharmacol. Sin. 2007, 28, 1343–1354. [Google Scholar] [CrossRef]
- Mandrich, L.; Caputo, E. Brassicaceae-derived anticancer agents: Towards a green approach to beat cancer. Nutrients 2020, 12, 868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Talalay, P.; Cho, C.G.; Posner, G.H. A major inducer of anticarcinogenic protective enzymes from broccoli: Isolation and elucidation of structure. Proc. Natl. Acad. Sci. USA 1992, 89, 2399–2403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Sabol, M.; Kutschy, P.; Siegfried, L.; Miroššay, A.; Suchý, M.; Hrbková, H.; Dzurilla, M.; Marušková, R.; Starková, J.; Paulíková, E. Cytotoxic effect of cruciferous phytoalexins against murine L1210 leukemia and B16 melanoma. Biologia 2000, 55, 701–707. [Google Scholar]
- Pilátová, M.; Sarisský, M.; Kutschy, P.; Mirossay, A.; Mezencev, R.; Curillová, Z.; Suchý, M.; Monde, K.; Mirossay, L.; Mojzis, J. Cruciferous phytoalexins: Antiproliferative effects in T-Jurkat leukemic cells. Leuk. Res. 2005, 29, 415–421. [Google Scholar]
- Pedras, M.S.C.; Zheng, Q.A.; Strelkov, S. Metabolic changes in roots of the oilseed canola infected with the biotroph Plasmodiophora brassicae: Phytoalexins and phytoanticipins. J. Agric. Food Chem. 2008, 56, 9949–9961. [Google Scholar] [CrossRef]
- Monde, K.; Takasugi, M.; Shirata, A. Three sulphur-containing stress metabolites from Japanese radish. Phytochemistry 1995, 39, 581–586. [Google Scholar] [CrossRef]
- Monde, K.; Taniguchi, T.; Miura, N.; Kutschy, P.; Curillova, Z.; Pilatova, M.; Mojzis, J. Chiral cruciferous phytoalexins: Preparation, absolute configuration, and biological activity. Bioorg. Med. Chem. 2005, 13, 5206–5212. [Google Scholar] [CrossRef]
- Combs, G.F., Jr. Current evidence and research needs to support a health claim for selenium and cancer prevention. J. Nutr. 2005, 135, 343–347. [Google Scholar] [CrossRef] [Green Version]
- Jackson, M.I.; Combs, G.F., Jr. Selenium and anticarcinogenesis: Underlying mechanisms. Curr. Opin. Clin. Nutr. Metab. Care 2008, 11, 718–726. [Google Scholar] [CrossRef]
- Zeng, H.; Combs, G.F., Jr. Selenium as an anticancer nutrient: Roles in cell proliferation and tumor cell invasion. J. Nutr. Biochem. 2008, 19, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.P.; Gandin, V. Selenium compounds as therapeutic agents in cancer. Biochim. Biophys. Acta 2015, 1850, 1642–1660. [Google Scholar] [CrossRef] [PubMed]
- El-Bayoumy, K.; Sinha, R. Mechanisms of mammary cancer chemoprevention by organoselenium compounds. Mutat. Res. 2004, 551, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Ip, C.; Thompson, H.J.; Zhu, Z.; Ganther, H.E. In vitro and in vivo studies of methylseleninic acid: Evidence that a monomethylated selenium metabolite is critical for cancer chemoprevention. Cancer Res. 2000, 60, 2882–2886. [Google Scholar]
- Sinha, R.; Unni, E.; Ganther, H.E.; Medina, D. Methylseleninic acid, a potent growth inhibitor of synchronized mouse mammary epithelial tumor cells in vitro. Biochem. Pharmacol. 2000, 61, 311–317. [Google Scholar] [CrossRef]
- Medina, D.; Thompson, H.; Ganther, H.; Ip, C. Se-Methylselenocysteine: A new compound for chemoprevention of breast cancer. Nutr. Cancer 2001, 40, 12–17. [Google Scholar] [CrossRef]
- Babaer, D.; Zheng, M.; Ivy, M.T.; Zent, R.; Tiriveedhi1, V. Methylselenol producing selenocompounds enhance the efficiency of mammaglobin-A peptide vaccination against breast cancer cells. Oncol. Lett. 2019, 18, 6891–6898. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Lisk, D.; Block, E.; Ip, C. Characterization of the biological activity of γ-glutamyl-Se- methylselenocysteine: A novel, naturally occurring anticancer agent from garlic. Cancer Res. 2001, 61, 2923–2928. [Google Scholar]
- Ávila, F.W.; Yang, Y.; Faquin, V.; Ramos, S.J.; Guilherme, L.R.G.; Thannhauser, T.W.; Li, L. Impact of selenium supply on Se-methylselenocysteine and glucosinolate accumulation in selenium-biofortified Brassica sprouts. Food Chem. 2014, 165, 578–586. [Google Scholar] [CrossRef]
- Cai, X.J.; Block, E.; Uden, P.C.; Zhang, X.; Quimby, B.D.; Sullivan, J.J. Allium chemistry: Identification of selenoamino acids in ordinary and selenium-enriched garlic, onion, and broccoli using gas chromatography with atomic emission detection. J. Agric. Food Chem. 1995, 43, 1754–1757. [Google Scholar] [CrossRef]
- Kotrebai, M.; Birringer, M.; Tyson, J.F.; Block, E.; Uden, P.C. Selenium speciation in enriched and natural samples by HPLC-ICP-MS and HPLC-ESI-MS with perfluorinated carboxylic acid ionpairing agents. Analyst 2000, 125, 71–78. [Google Scholar] [CrossRef]
- Reich, H.J.; Hondal, R.J. Why nature chose selenium. ACS Chem. Biol. 2016, 11, 821–841. [Google Scholar] [CrossRef] [PubMed]
- Iwasawa, S.; Yamano, Y.; Takiguchi, Y.; Tanzawa, H.; Tatsumi, K.; Uzawa, K. Upregulation of thioredoxin reductase 1 in human oral squamous cell carcinoma. Oncol. Rep. 2011, 25, 637–644. [Google Scholar]
- Drake, E.N. Cancer chemoprevention: Selenium as a prooxidant, not an antioxidant. Med. Hypotheses 2006, 67, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Chaudière, J.; Courtin, O.; Leclaire, J. Glutathione oxidase activity of selenocystamine: A mechanistic study. Arch. Biochem. Biophys. 1992, 296, 328–336. [Google Scholar] [CrossRef]
- Misra, S.; Boylan, M.; Selvam, A.; Spallholz, J.E.; Björnstedt, M. Redox-active selenium compounds–from toxicity and cell death to cancer treatment. Nutrients 2015, 7, 3536–3556. [Google Scholar] [CrossRef] [Green Version]
- Brodin, O.; Eksborg, S.; Wallenberg, M.; Asker-Hagelberg, C.; Larsen, E.H.; Mohlkert, D.; Lenneby-Helleday, C.; Jacobsson, H.; Linder, S.; Misra, S.; et al. Pharmacokinetics and toxicity of sodium selenite in the treatment of patients with carcinoma in a phase I clinical trial: The SECAR study. Nutrients 2015, 7, 4978–4994. [Google Scholar] [CrossRef] [Green Version]
- Schiavon, M.; Lima, L.W.; Jiang, Y.; Hawkesford, M.J. Effects of selenium on plant metabolism and implications for crops and consumers. In Selenium in Plants; Pilon-Smits, E.A.H., Winkel, L.H.E., Lin, Z.-Q., Eds.; Springer International Publishing AG: Cham, Switzerland, 2017; Volume 11, pp. 257–275. [Google Scholar]
- Schiavon, M.; Berto, C.; Malagoli, M.; Trentin, A.; Sambo, P.; Dall’Acqua, S.; Pilon-Smits, E.A.H. Selenium biofortification in radish enhances nutritional quality via accumulation of methyl-selenocysteine and promotion of transcripts and metabolites related to glucosinolates, phenolics amino acids. Front. Plant Sci. 2016, 7, 1371. [Google Scholar] [CrossRef] [Green Version]
- Schiavon, M.; Pittarello, M.; Pilon-Smits, E.A.H.; Wirtz, M.; Hell, R.; Malagoli, M. Selenate and molybdate alter sulfate transport and assimilation in Brassica juncea L. Czern.: Implications for phytoremediation. Environ. Exp. Bot. 2012, 75, 41–51. [Google Scholar] [CrossRef]
- El Mehdawi, A.F.; Pilon-Smits, E.A.H. Ecological aspects of plant selenium hyperaccumulation. Plant Biol. 2012, 14, 1–10. [Google Scholar] [CrossRef]
- Pilon-Smits, E.A.H.; Quinn, C.F.; Tapken, W.; Malagoli, M.; Schiavon, M. Physiological functions of beneficial elements. Curr. Opin. Plant Biol. 2009, 12, 267–274. [Google Scholar] [CrossRef]
- Terry, N.; Zayed, A.M.; de Souza, M.P.; Tarun, A.S. Selenium in higher plants. Annu. Rev. Plant Physiol. Plan. Mol. Biol. 2000, 51, 401–432. [Google Scholar] [CrossRef] [Green Version]
- Pilon-Smits, E.A.H. On the ecology of selenium accumulation in plants. Plants 2019, 8, 197. [Google Scholar] [CrossRef] [Green Version]
- Pilon-Smits, E.A.H.; LeDuc, D.L. Phytoremediation of selenium using transgenic plants. Curr. Opin. Biotechnol. 2009, 20, 207–212. [Google Scholar] [CrossRef]
- White, P.J.; Bowen, H.C.; Parmaguru, P.; Fritz, M.; Spracklen, W.P.; Spiby, R.E.; Meacham, M.C.; Mead, A.; Harriman, M.; Trueman, L.J.; et al. Interactions between selenium and sulphur nutrition in Arabidopsis thaliana. J. Exp. Bot. 2004, 55, 1927–1937. [Google Scholar] [CrossRef] [Green Version]
- Galeas, M.L.; Zhang, L.H.; John, L.; Freeman, J.L.; Wegner, M.; Pilon-Smits, E.A.H. Seasonal fluctuations of selenium and sulfur accumulation in selenium hyperaccumulators and related nonaccumulators. New Phytol. 2007, 173, 517–525. [Google Scholar] [CrossRef]
- El Mehdawi, A.F.; Jiang, Y.; Guignardi, Z.S.; Esmat, A.; Pilon, M.; Pilon-Smits, E.A.H.; Schiavon, M. Influence of sulfate supply on selenium uptake dynamics and expression of sulfate/selenate transporters in selenium hyperaccumulator and nonhyperaccumulator Brassicaceae. New Phytol. 2018, 217, 194–205. [Google Scholar] [CrossRef] [Green Version]
- Schiavon, M.; Pilon, M.; Malagoli, M.; Pilon-Smits, E.A. Exploring the importance of sulfate transporters and ATP sulphurylases for selenium hyperaccumulation-a comparison of Stanleya pinnata and Brassica juncea (Brassicaceae). Front. Plant Sci. 2015, 6, 2. [Google Scholar] [CrossRef] [Green Version]
- Tian, M.; Hui, M.; Thannhauser, T.W.; Pan, S.; Li, L. Selenium-induced toxicity is counteracted by sulfur in broccoli (Brassica oleracea L. var. italica). Front. Plant Sci. 2017, 8, 1425. [Google Scholar] [CrossRef] [Green Version]
- Guignardi, Z.; Schiavon, M. Biochemistry of plant selenium uptake and metabolism. In Selenium in Plants. Plant Ecophysiology; Pilon-Smits, E., Winkel, L., Lin, Z.Q., Eds.; Springer International Publishing AG: Cham, Switzerland, 2017; Volume 11, pp. 21–34. [Google Scholar]
- Bellof, S.; Mühling, K.H.; Bender, L. Detection of putative selenoproteins in Chinese cabbage (Brassica pekinensis L.). J. Soil Sci. Plant Nutr. 2007, 170, 657–659. [Google Scholar] [CrossRef]
- Hawkesford, M.J.; Zhao, F.J. Strategies for increasing the selenium content of wheat. J. Cereal Sci. 2007, 46, 282–292. [Google Scholar] [CrossRef]
- MacFarquhar, J.K.; Broussard, D.L.; Melstrom, P.; Hutchinson, R.; Wolkin, A.; Martin, C.; Burk, R.F.; Dunn, J.R.; Green, A.L.; Hammond, R.; et al. Acute selenium toxicity associated with a dietary supplement. Arch. Intern. Med. 2010, 170, 256–261. [Google Scholar] [CrossRef] [Green Version]
- Toler, H.D.; Charron, C.S.; Sams, C.E.; Randle, W.R. Selenium increases sulfur uptake and regulates glucosinolate metabolism in rapid-cycling Brassica oleracea. J. Am. Soc. Hortic. Sci. 2007, 132, 14–19. [Google Scholar] [CrossRef] [Green Version]
- Barickman, T.C.; Kopsell, D.A.; Sams, C.E. Selenium influences glucosinolate and isothiocyanates and increases sulfur uptake in Arabidopsis thaliana and rapid-cycling Brassica oleracea. J. Agric. Food Chem. 2013, 61, 202–209. [Google Scholar] [CrossRef]
- Bachiega, P.; Salgado, J.M.; de Carvalho, J.E.; Ruiz, A.L.T.G.; Schwarz, K.; Tezotto, T.; Morzelle, M.G. Antioxidant and antiproliferative activities in different maturation stages of broccoli (Brassica oleracea Italica) biofortified with selenium. Food Chem. 2016, 190, 771–776. [Google Scholar] [CrossRef] [Green Version]
- Dall’Acqua, S.; Ertani, A.; Pilon-Smits, E.A.H.; Fabrega-Prats, M.; Schiavon, M. Selenium biofortification differentially affects sulfur metabolism and Accumulation of phytochemicals in two rocket species (Eruca Sativa Mill. and Diplotaxis Tenuifolia) Grown in Hydroponics. Plants 2019, 8, 68. [Google Scholar] [CrossRef] [Green Version]
- Charron, C.S.; Kopsell, D.A.; Randle, W.M.; Sams, C.E. Sodium selenate fertilisation increases Se accumulation and decreases glucosinolate concentration in rapid-cycling Brassica oleracea. J. Sci. Food Agric. 2001, 81, 962–966. [Google Scholar] [CrossRef]
- Freeman, J.L.; Zhang, L.H.; Marcus, M.A.; Fakra, S.; McGrath, S.P.; Pilon-Smits, E.A.H. Spatial imaging, speciation, and quantification of selenium in the hyperaccumulator plants Astragalus bisulcatus and Stanleya pinnata. Plant Physiol. 2006, 142, 124–134. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Mao, S.; Xu, H.; Wu, Q.; Liang, M.; Yuan, Y.; Liu, M.; Huang, K.; Wu, Q. Effects of sulfur and selenium on glucosinolate biosynthesis in cabbage. Plant Mol. Biol. Rep. 2020, 38, 62–74. [Google Scholar] [CrossRef]
- Stroud, J.; Zhao, F.J.; Buchner, P.; Shinmachi, F.; McGrath, S.P.; Abecassis, J.; Hawkesford, M.J.; Shewry, P.R. Impacts on sulphur nutrition on selenium and molybdenum concentrations in wheat grain. J. Cereal Sci. 2010, 52, 111–113. [Google Scholar] [CrossRef]
- Shinmachi, F.; Buchner, P.; Stroud, J.L.; Parmar, S.; Zhao, F.J.; McGrath, S.P.; Hawkesford, M.J. Influence of sulfur deficiency on the expression of specific sulfate transporters and the distribution of sulfur, selenium and molybdenum in wheat. Plant Physiol. 2010, 153, 327–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mechora, Š.; Germ, M.; Stibilj, V. Selenium compounds in selenium-enriched cabbage. Pure Appl. Chem. 2012, 84, 259–268. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Jenkins, S.N.; Fahey, J.W.; Ye, L.; Wehage, S.L.; Liby, K.T.; Stephenson, K.K.; Wade, K.L.; Talalay, P. Protection against UV light-induced skin carcinogenesis in SKH-1 high-risk mice by sulforaphane-containing broccoli sprout extracts. Cancer Lett. 2006, 240, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Munday, R.; Mhawech-Fauceglia, P.; Munday, C.M.; Paonessa, J.D.; Tang, L.; Munday, J.S.; Lister, C.; Wilson, P.; Fahey, J.W.; Davis, W.; et al. Inhibition of urinary bladder carcinogenesis by broccoli sprouts. Cancer Res. 2008, 68, 1593–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, J.D.; Paton, V.G.; Vidanes, G. Potent induction of phase 2 enzymes in human prostate cells by sulforaphane. Cancer Epidemiol. Biomark. Prev. 2001, 10, 949–954. [Google Scholar]
- Tang, L.; Zhang, Y.; Jobson, H.E.; Li, J.; Stephenson, K.K.; Wade, K.L.; Fahey, J.W. Potent activation of mitochondria-mediated apoptosis and arrest in S and M phases of cancer cells by a broccoli sprout extract. Mol. Cancer Ther. 2006, 5, 935–944. [Google Scholar] [CrossRef] [Green Version]
- Abdulah, R.; Koyama, H.; Faried, A.; Kobayashi, K.; Yamazaki, C.; Suradji, E.W.; Ito, K.; Suzuki, K.; Murakami, M.; Kuwano, H.; et al. Selenium enrichment of broccoli sprout extract increases chemosensitivity and apoptosis of LNCaP prostate cancer cells. BMC Cancer 2009, 9, 414. [Google Scholar] [CrossRef] [Green Version]
- Ellis, D.R.; Salt, D.E. Plants, selenium and human health. Curr. Opin. Plant Biol. 2003, 6, 273–279. [Google Scholar] [CrossRef]
- Matich, A.J.; McKenzie, M.J.; Lill, R.E.; Brummell, D.A.; McGhie, T.K.; Chen, R.K.-Y.; Rowan, D.D. Selenoglucosinolates and their metabolites produced in Brassica spp. fertilised with sodium selenate. Phytochemistry 2012, 75, 140–152. [Google Scholar] [CrossRef]
- Emmert, S.W.; Desai, D.; Amin, S.; Richie, J.P., Jr. Enhanced Nrf2-dependent induction of glutathione in mouse embryonic fibroblasts by isoselenocyanate analog of sulforaphane. Bioorg. Med. Chem. Lett. 2010, 20, 2675–2679. [Google Scholar] [CrossRef] [Green Version]
- Palomo-Siguero, M.; Lopez-Heras, M.I.; Camara, C.; Madrid, Y. Accumulation and biotransformation of chitosan-modified selenium nanoparticles in exposed radish (Raphanus sativus). J. Anal. Atom. Spectrom. 2015, 30, 1237–1244. [Google Scholar] [CrossRef]
- Chan, Q.L.; Afton, S.E.; Caruso, J.A. Investigation of selenium metabolites in se-enriched kale, Brassica oleracea a, via HPLC-ICPMS and nanoESI-ITMS. J. Anal. Atom. Spectrom. 2010, 25, 186–192. [Google Scholar] [CrossRef]
- De Vos, M.; Kriksunov, K.L.; Jander, G. Indole-3-acetonitrile production from indole glucosinolates deters oviposition by Pieris rapae. Plant Physiol. 2008, 146, 916–926. [Google Scholar] [CrossRef] [Green Version]
- Peñas, E.; Martinez-Villaluenga, C.; Frias, J.; Sánchez-Martínez, M.J.; Pérez-Corona, M.T.; Madrid, Y.; Cámara, C.; Vidal-Valverde, C. Se improves indole glucosinolate hydrolysis products content, se-methylselenocysteine content, antioxidant capacity and potential anti-inflammatory properties of sauerkraut. Food Chem. 2012, 132, 907–914. [Google Scholar] [CrossRef] [Green Version]
- Ouerdane, L.; Aureli, F.; Flis, P.; Bierla, K.; Preud’homme, H.; Cubadda, F.; Szpunar, J. Comprehensive speciation of low-molecular weight selenium metabolites in mustard seeds using HPLC—Electrospray linear trap/orbitrap tandem mass spectrometry. Metallomics 2013, 5, 1294–1304. [Google Scholar] [CrossRef]
- Maneetong, S.; Chookhampaeng, S.; Chantiratikul, A.; Chinrasri, O.; Thosaikham, W.; Sittipout, R.; Chantiratikul, P. Hydroponic cultivation of selenium-enriched kale (Brassica oleracea var. alboglabra L.) seedling and speciation of selenium with HPLC-ICPMS. Microchem. J. 2013, 108, 87–91. [Google Scholar] [CrossRef]
- Ávila, F.W.; Faquin, V.; Yang, Y.; Ramos, S.J.; Guilherme, L.R.G.; Thannhauser, T.W.; Li, L. Assessment of the anticancer compounds Se-methylselenocysteine and glucosinolates in Se-biofortified broccoli (Brassica oleracea L. var. italica) sprouts and florets. J. Agric. Food Chem. 2013, 61, 6216–6223. [Google Scholar] [CrossRef]
- Bañuelos, G.S.; Arroyo, I.; Pickering, I.J.; Yang, S.I.; Freeman, J.L. Selenium biofortification of broccoli and carrots grown in soil amended with Se-enriched hyperaccumulator Stanleya pinnata. Food Chem. 2015, 166, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Bañuelos, G.S.; Arroyo, I.S.; Dangi, S.R.; Zambrano, M.C. Continued selenium biofortification of carrots and broccoli grown in soils once amended with se-enriched S. pinnata. Front. Plant Sci. 2016, 7, 1251. [Google Scholar] [CrossRef] [Green Version]
- Kahakachchi, C.; Boakye, H.T.; Uden, P.C.; Tyson, J.F. Chromatographic speciation of anionic and neutral selenium compounds in accumulating Brassica juncea (Indian mustard) and in selenized yeast. J. Chromatogr. A 2004, 1054, 303–312. [Google Scholar] [CrossRef]
- Jones, G.D.; Droz, B.; Greve, P.; Gottschalk, P.; Poffet, D.; McGrath, S.P.; Seneviratne, S.I.; Smith, P.; Winkel, L.H.E. Selenium deficiency risk predicted to increase under future climate change. Proc. Natl. Acad. Sci. USA 2017, 114, 2848–2853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellingray, L.; Tapp, H.S.; Saha, S.; Doleman, J.F.; Narbad, A.; Mithen, R.F. Consumption of a diet rich in Brassica vegetables is associated with a reduced abundance of sulphate-reducing bacteria: A randomised crossover study. Mol. Nutr. Food Res. 2017, 61, 1600992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Traw, M.B.; Dawson, T.E. Differential induction of trichomes by three herbivores of black mustard. Oecologia 2002, 131, 526–532. [Google Scholar] [CrossRef]
- Wiesner-Reinhold, M.; Schreiner, M.; Baldermann, S.; Schwarz, D.; Hanschen, F.S.; Kipp, A.P.; Rowan, D.D.; Bentley-Hewitt, K.L.; McKenzie, M.J. Mechanisms of selenium enrichment and measurement in Brassicaceous vegetables, and their application to human health. Front. Plant Sci. 2017, 8, 1365. [Google Scholar] [CrossRef] [Green Version]
- Wilschefski, S.C.; Baxter, M.R. Inductively coupled plasma mass spectrometry: Introduction to analytical aspects. Clin. Biochem. Rev. 2019, 40, 115–133. [Google Scholar] [CrossRef]
- Campillo, N.; Penalver, R.; López-García, I.; Hernández-Córdoba, M. Headspace solid-phase microextraction for the determination of volatile organic sulphur and selenium compounds in beers, wines and spirits using gas chromatography and atomic emission detection. J. Chromatogr. A 2009, 1216, 6735–6740. [Google Scholar] [CrossRef]



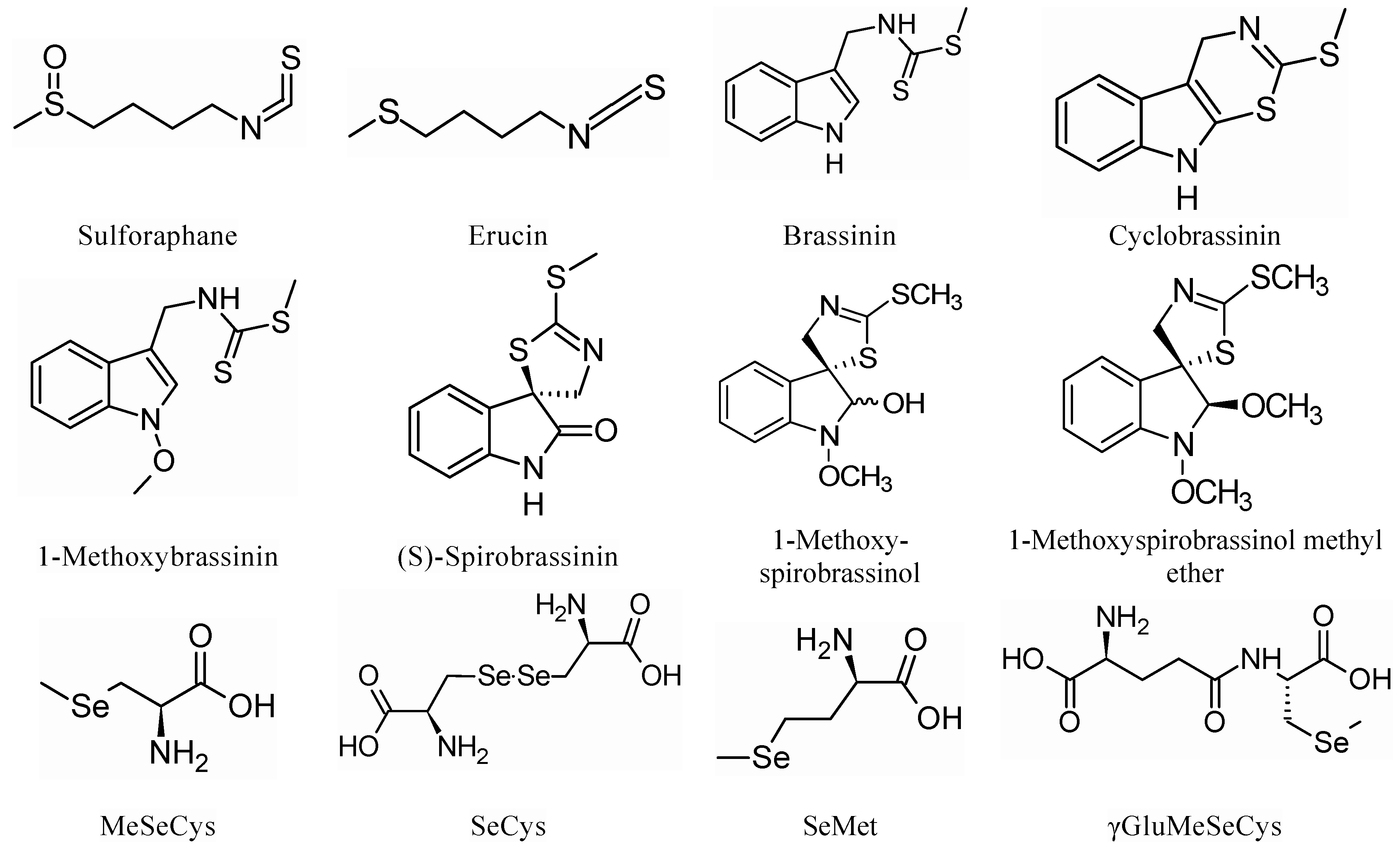{kind=link}
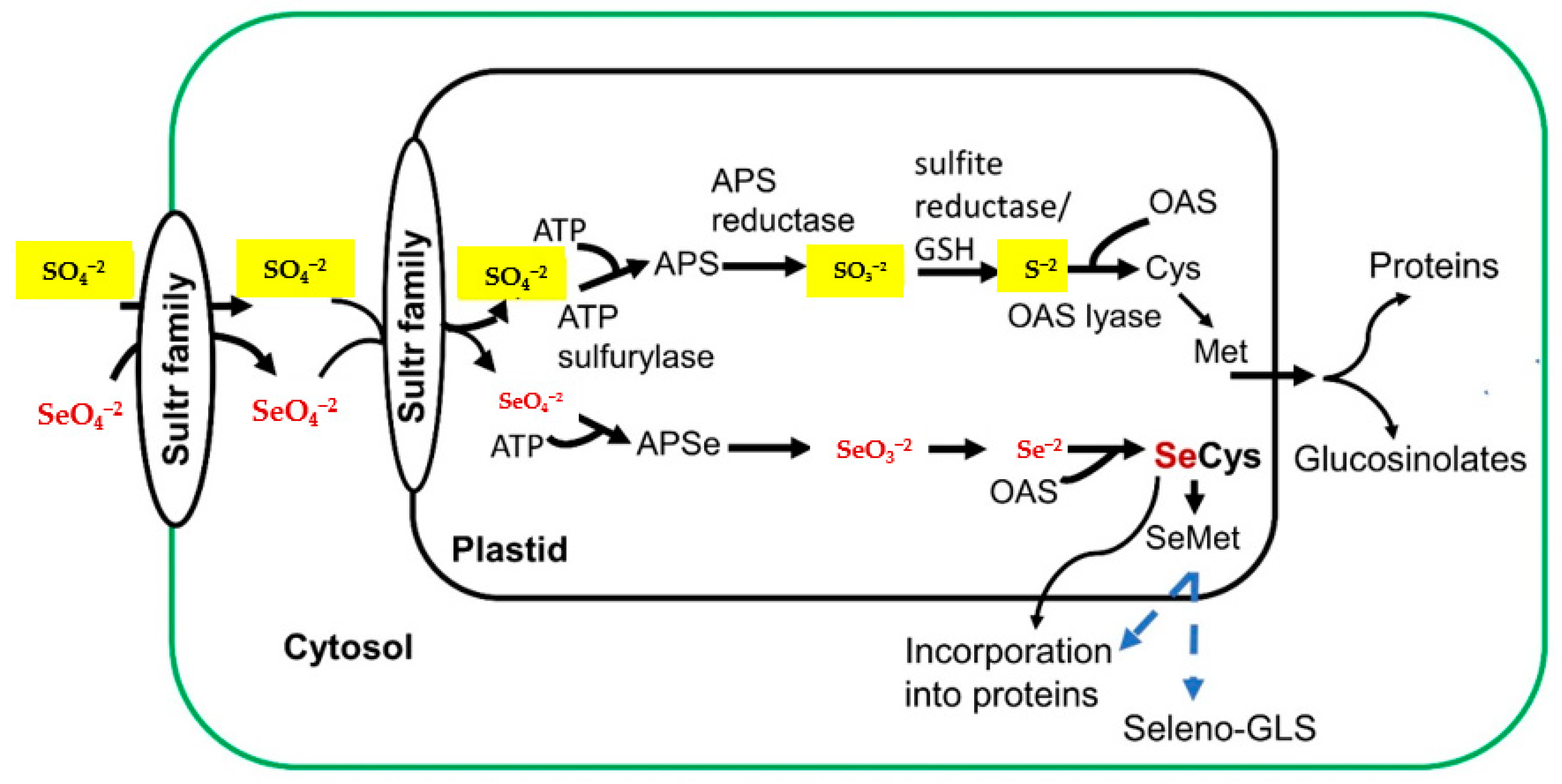{kind=link}
| Studied Plant/Plants | Se or Se/S Application Methods | Observed Findings | References |
|---|---|---|---|
| Arabidopsis thaliana cv. Columbia and rapid cycling B. oleracea | Plants grown in hydroponic system. Four Na2SeO4 treatments (0.4, 0.8, 1.6, and 3.2 mg Se L−1) in addition to control treatment (no Se). One Se/S combination (0.8 mg Se L−1 and high S level 37 mg S L−1). All plants were harvested in 28−31 days | In both B. oleracea and A. thaliana the authors detected glucoraphanin, glucoiberin, glucoerucin, sinigrin, and indole-3-ylmethyl glucosinolates in response to all Se and higher sulfate doses. This might afford dietary Se and anticarcinogenic ITCs compounds at promising concentrations required to obtain chemopreventive properties | [79] |
| Broccoli (B. oleracea Italica) | Treatments of seeds and seedlings consisted of distilled water and 50 µM Na2SeO4, and inflorescences treated with distilled water and 1.5 mM Na2SeO4 | The non-polar extracts of all samples exhibited antiproliferative effects. Se biofortified extract of broccoli seedlings with selenium showed cytocidal activity for a glioma cell line (U251, GI50 28.5 mg L−1). | [80] |
| Salad rocket (E. sativa Mill.) and wild rocket (Diplotaxis tenuifolia) | Hydroponically grown plants under basal nutrient solution included MgSO4 at 1.5 µM in addition to other elements. At 30 days sodium selenate (Na2SeO4) was applied to the nutrient solution at 5, 10, 20 or 40 µM. | Se fertilization reduced the production of total GLS in E. sativa, consisted of aliphatic GLS, which synthesized from methionine. Additionally, higher amounts of phenolic compounds were decreased in D. tenuifolia in response to all Se levels, whereas in E. sativa the depletetion of phenolic compounds occurred only at higher Se concentrations. | [81] |
| Rapid-cycling B. oleracea | Plants grown in hydroponic system under different Se concentrations: 0, 1.0, 2.0, 3.0, 6.0, 7.2 or 9.0 mg L−1 Na2SeO4, S concentration was 246.48 mg L−1 MgSO4·7H2O. | The detection of 3-indolylmethyl glucosinolate and 2-propenyl glucosinolate in shoots under varied Se concentrations might offer chemopreventive effect. | [82] |
| A. bisulcatus, S. pinnata and B. juncea | Plants were cultivated for 8 or 6 months and irrigated 3 times a week with 40 μM SeO4−2. B. juncea were grown for one month and watered 3 times a week with 20 μM SeO4−2 for an extra month | Se was accumulated in the form of MeSeCys and γ-glutamyl-MeSeCys in the trichomes, which might show their capability of sequestration or biosynthesis of organo Se comounds in trichomes | [83] |
| Cabbage (B. oleracea var. capitata L. cv Jingfeng) | Sodium selenite (Na2SeO3) via foliage leaf spray (50, 100, or 150 μmol L−1) and MgSO4 via root irrigation (1 mM and 4 mM) and Na2SO4 treatments (3 mM), which were added with Hoagland’s nutrient solution for ten weeks. | GLS level was enhanced by low selenite (50 μmol L−1) and S (1 mmol L−1) concentrations or elevated selenite (100 μmol L−1) and S (4 mmol L−1) treatments. | [84] |
| cabbage (B.oleracea var. capitata L., cv. Pandion) and red cabbage (B. oleracea var. capitata L. f. rubra, cv. Erfurtsko rano) | Se was foliarly sprayed in the form of Na2SeO4 twice with 20 mg Se L−1 for cabbage and twice with 0.5 mg Se L−1 for red cabbage. | SeMet was the most abundant compound detected in all parts of cabbage and red cabbage; it constituted 94 and 55%, respectively, of the soluble Se concentration in the roots of both cabbages. In the stems and leaves of cabbage, SeMet was only 23% of the soluble Se content. In red cabbage SeMet was 80% and 41% of the soluble Se content in stems and leaves, respectively. | [87] |
| 16 different brassica seeds cultivars including three broccoli cultivars, three cauliflower cultivars, three green cabbage cultivars, three Chinese cabbage cultivars, three kale cultivars and one Brussels sprouts | Seeds were soaked with 50 μM Na2SeO4, sprouts were treated with the same Se concentration every 24 h and harvested after 7 days | All sprouts treated with Se accumulate substantial amounts of MeSeCys. The lowest MeSeCys amounts were detected in sprouts of both green cabbage cultivars and brussels sprouts. Chinese cabbage had accommodated relatively high MeSeCys levels. | [52] |
| Broccoli sprouts B. oleracea var. italica | Seeds were grown in 10 ppm of Na2SeO3 solution to produce Se enriched sprouts or in deionized H2O to produce control broccoli sprouts | Selenium-enriched broccoli sprouts showed better cell proliferation inhibition than control broccoli sprouts including reducing prostate-specific antigen secretion and enhancing apoptosis of prostate cancer cells. Selenium-enriched sprouts but not normal sprouts, stimulate a downregulation of survival protein kinase B (Akt)/mammalian target of rapamycin (mTOR) pathway, which regulates various cellular processes | [92] |
| Broccoli (B. oleracea L. var. italica cv. Triathlon), cauliflower (B. oleracea L. var. botrytis cv. Liberty), and forage rape (B. napus cv. Maxima) | Plants were grown in soil and treated with 20 mL of 5.0 mM Na2SeO4 twice weekly for 4 weeks. | Three selenoglucosinolates were obtained including 3-(methylseleno)propylglucosinolate (glucoselenoiberverin), 4-(methylseleno) butylglucosinolate (glucoselenoerucin), and 5-(methylseleno)pentylglucosinolate (glucoselenoberteroin). Additionally, six organoselenium compounds were detected. Selenoglucosinolates levels and their aglycones including ITCs in forage rape, were up to 10% and 70%, respectively, of their S counterparts. Moreover, in broccoli, selenoglucosinolates concentrations and their aglycones including nitriles occurred up to 60% and 1300%, of their S counterparts. | [94] |
| Radish Raphanus sativus | Plants were grown under hydroponic conditions with addition of selenite or Se nanoparticles (1 mg L−1) for 40 days | More than 95% of the accommodated Se was incorporated into MeSeCys and SeMet | [96] |
| Kale B. oleracea A. | Plants were grown in cells contained Promix BX. Seven weeks after seeding, the cells were fertilized with Na2SeO3 or Na2SeO4 at a concentration of 10 µg g−1 Se for two weeks | MeSeCys was found to be the prominent Seleno compound found in kale and indicated to be responsible for its chemopreventive effect | [97] |
| White cabbage (B. oleracea L. var. capitata cv. Megaton) | Cabbages were shredded, and 0.5% NaCl and 0.3 mg of Na2SeO3/kg of fresh cabbage were added. Sauerkrauts without Se addition was prepared (control). Fermentation was performed by using indigenous microbiota of raw cabbage at room temperature (22–25 °C) for 7 days. | MeSeCys was found to be the major Se metabolite in Se-enriched sauerkraut | [99] |
| Black mustard (B. nigra) | Seeds cultivated in the seleniferous belt in the Punjab region (India) and the Se soil level is 2 to 7 mg kg−1 | Selenoglucosinolates were identified as the main important group among Se compounds, which constituted 15% of the total existing Se and up to 50% of all present Se species. Other characterized Se metabolites were selenoamino acids, selenosugars, selenosinapine and selenourea derivatives | [100] |
| Kale B. oleracea var. sabellica L. | Seedlings were grown under hydroponic conditions with Hoagland’s solution and treated with 5, 10, 15, 30 and 45 μg mL−1 Na2SeO3 for 15 days and harvested every 5 days. | SeMet, and MeSeCys are found to be predominant seleno compunds | [101] |
| Broccoli B. oleracea L. var. Italica Group | Seeds soaked in Magenta boxes and Se treatments were 10, 25, 50, 75, and 100 μM as Na2SeO4 or Na2SeO3. S treatments were 0.1, 1, and 10 mM. Sprouts harvested in 7 days. In the subsequent two experiments 5 cultivars were used. One experiment with sprouts using the same procedure as above. The other one with mature broccoli plants in which seeds were planted in soil, additionally 100 mL of 1.5 mM Na2SeO4 applied to give a final concentration of 25 μM Se. 6 applications were performed 2× per week for 3 weeks | MeSeCys level in sprouts simultaneously enhanced with increasing Se treatments. Increasing S treatments diminished the total Se and MeSeCys levels in sprouts biofortified with selenate, but not in sprouts treated with selenite. Importantly, sprouts had a 6-fold higher glucoraphanin level than florets | [102] |
| Broccoli B. oleracea L.var. Marathon | Soil fertilization with increasing amounts of dried Se-enriched S. pinnata (~700 μg Se g−1 DW). Broccoli was then cultivated in triplicate as 14 day old transplants | Soluble Se metabolites in broccoli florets were 58% seleno-methionine, 15% selenocystine, 7.4% MeSeCys, 6% selenate and 3.1% selenite | [103] |
| Broccoli B. oleracea L.var. Marathon | Se-amended soil by using S. pinnata plant material as discriebed in Bañuelos et al. (2015) [103] | The identified Se species constituted an average of 50, 11, and 5% as SeMet, SeCys, and MeSeCys, respectively, in addition to unknown compounds | [104] |
| Indian mustard (B. juncea) | Plants grown in hydroponic conditions and after 30 days Na2SeO4 or Na2SeO3 were used to obtain a concentration of 5 µg Se mL−1 and the S concentration was 2 mM MgSO4 | The identified Se species were MeSeCys (3.1%) in shoot biofortified with selenate, (10.7%) and S-(methylseleno)cysteine was found in both shoot and root supplemented with selenate or selenite. The percentage of SeMet was 27.8 and 43.1% in both selenate and selenite treated roots, respectively. Additionally, SeMet found in shoot biofortified with selenate and selenite were 10.7% and 34.2%, respectively. | [105] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdalla, M.A.; Sulieman, S.; Mühling, K.H. Regulation of Selenium/Sulfur Interactions to Enhance Chemopreventive Effects: Lessons to Learn from Brassicaceae. Molecules 2020, 25, 5846. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25245846
Abdalla MA, Sulieman S, Mühling KH. Regulation of Selenium/Sulfur Interactions to Enhance Chemopreventive Effects: Lessons to Learn from Brassicaceae. Molecules. 2020; 25(24):5846. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25245846
Chicago/Turabian StyleAbdalla, Muna Ali, Saad Sulieman, and Karl H. Mühling. 2020. "Regulation of Selenium/Sulfur Interactions to Enhance Chemopreventive Effects: Lessons to Learn from Brassicaceae" Molecules 25, no. 24: 5846. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25245846