
Identification of Phytochemical-Based β-Catenin Nuclear Localization Inhibitor in NSCLC: Differential Targeting Population from Member of Isothiocyanates


Abstract
:1. Introduction
2. Results
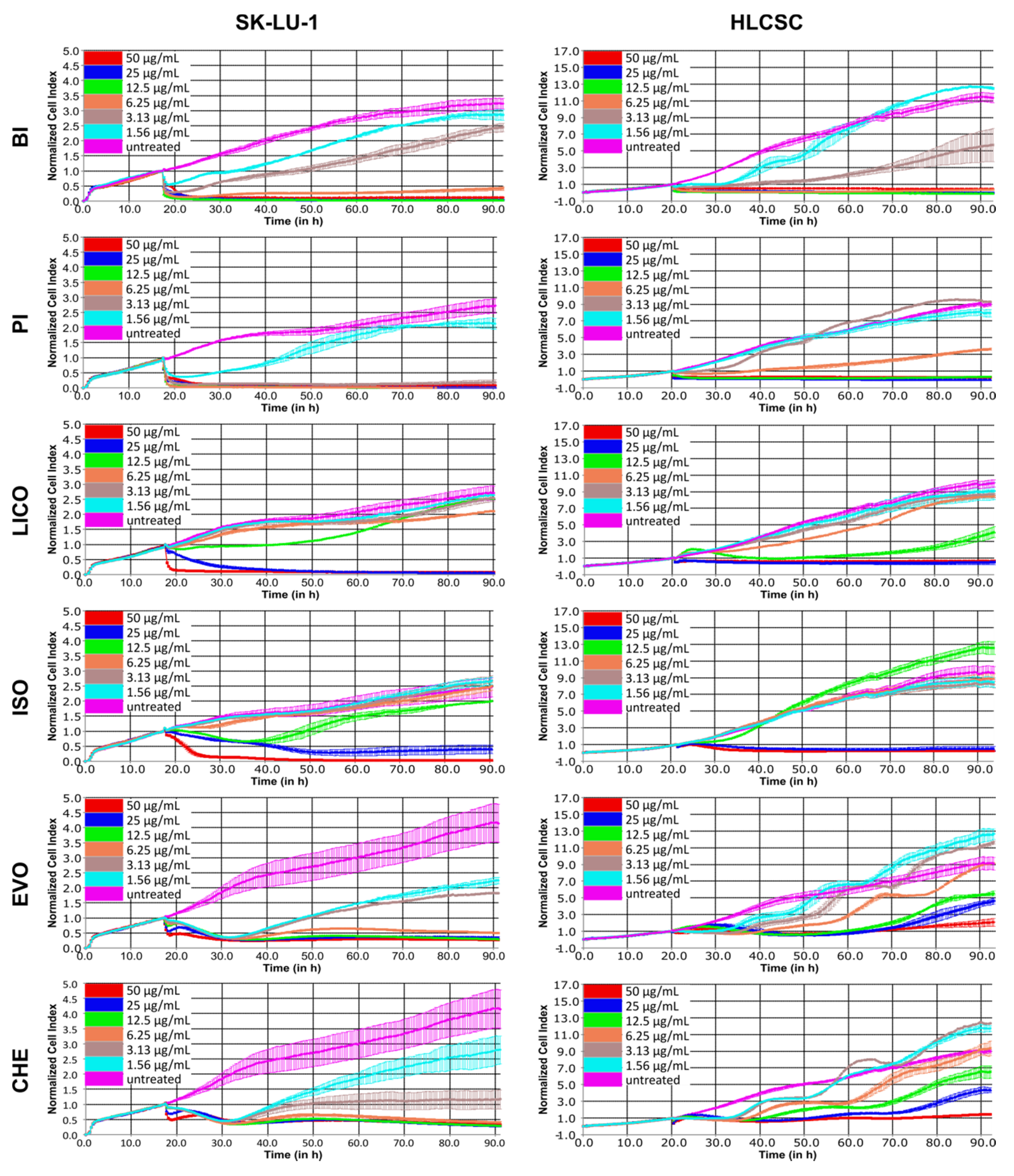
2.1. The Growth Inhibitory Profiles and IC50 Values of Alkaloids, Chalcones and Isothiocyanates
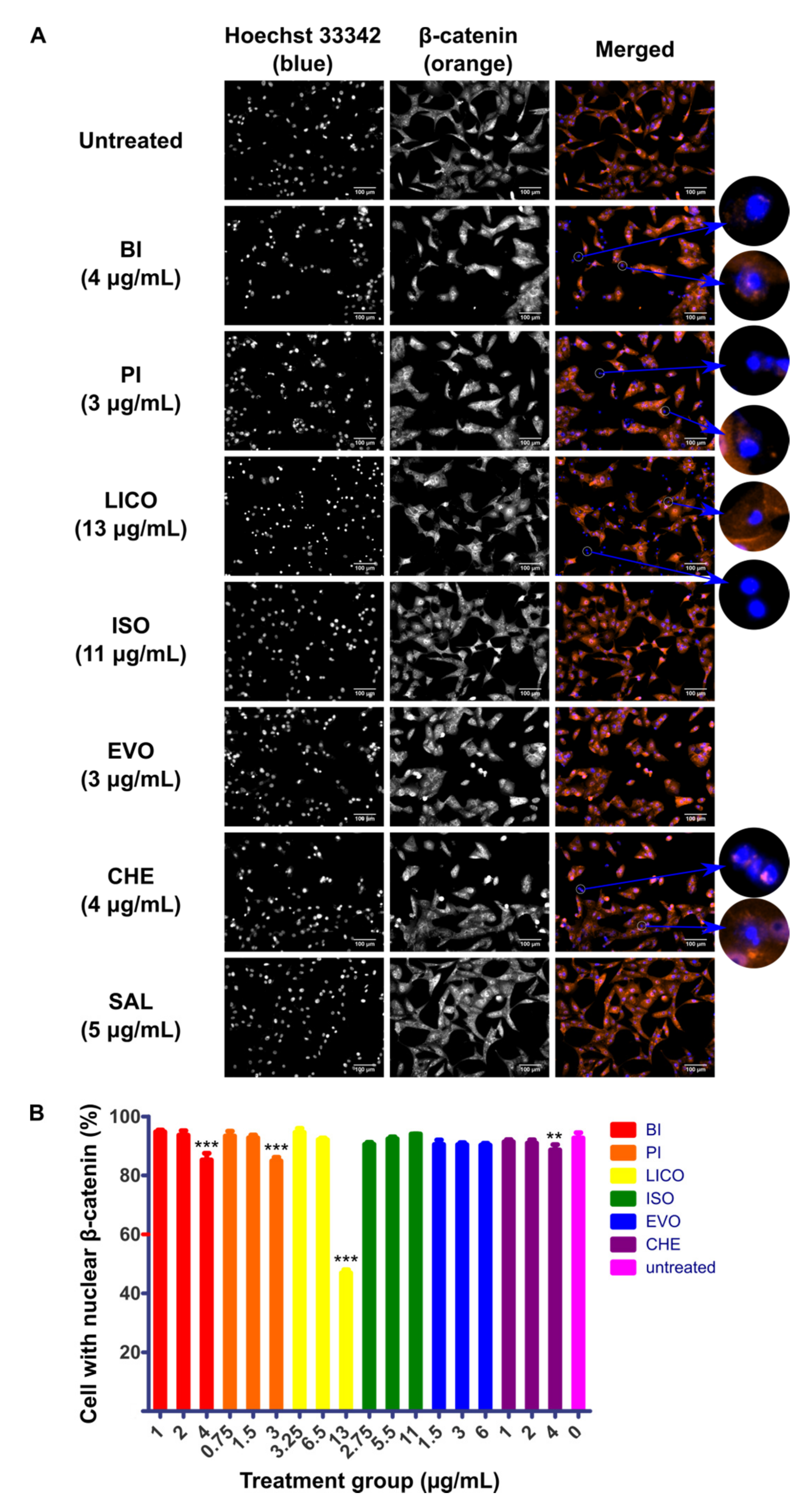
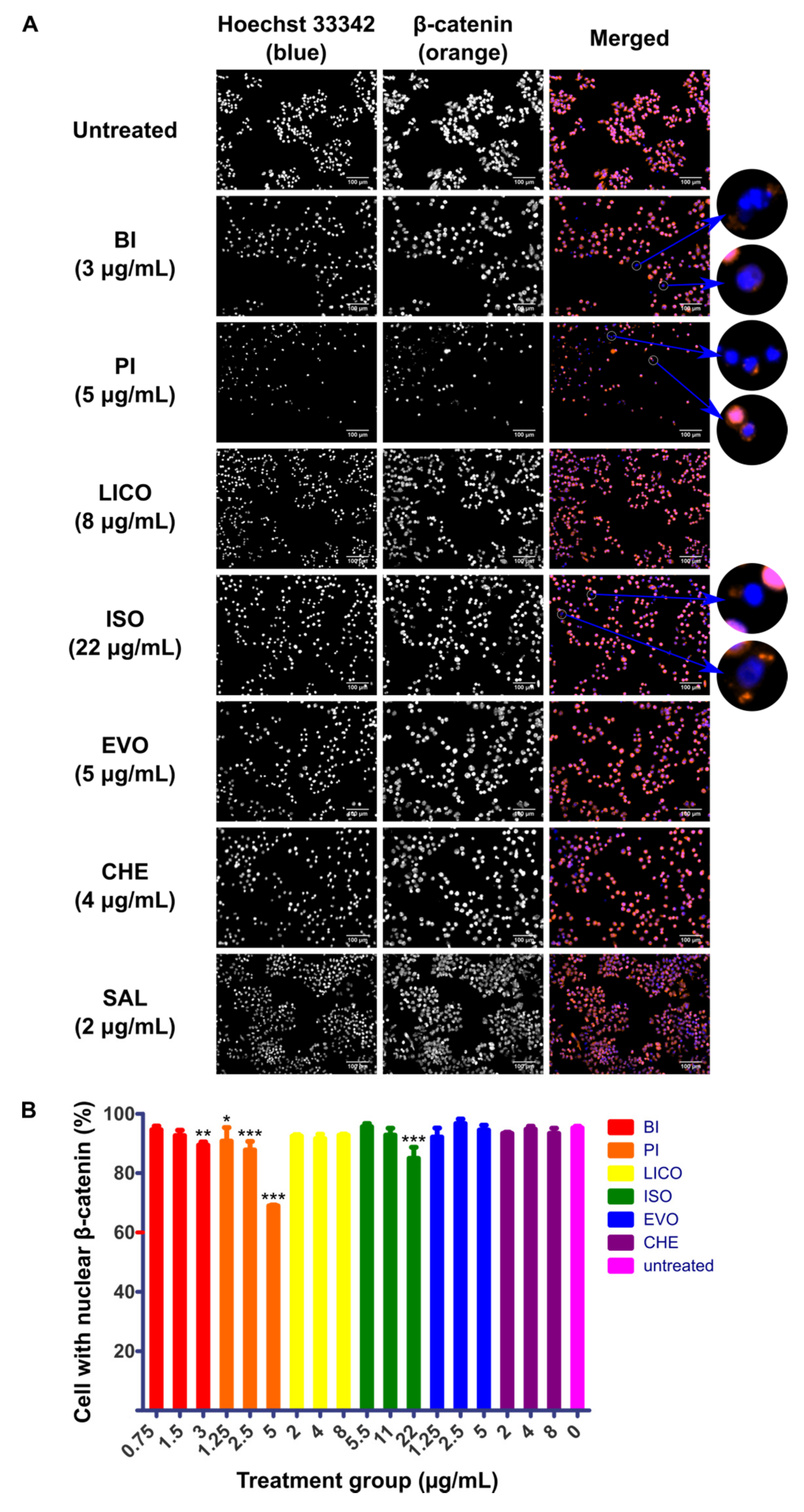
2.2. Chalcones and Isothiocyanates Modulated Reduction of β-Catenin Nuclear Localization
2.3. Both Isothiocyanate Members were Equally Potent to Inhibit the Growth of Wnt-Dependent Cell Line NCI-H1703 Although with Differing Sensitivity in Inhibiting Undifferentiated and Differentiated Lung Cancer Cells
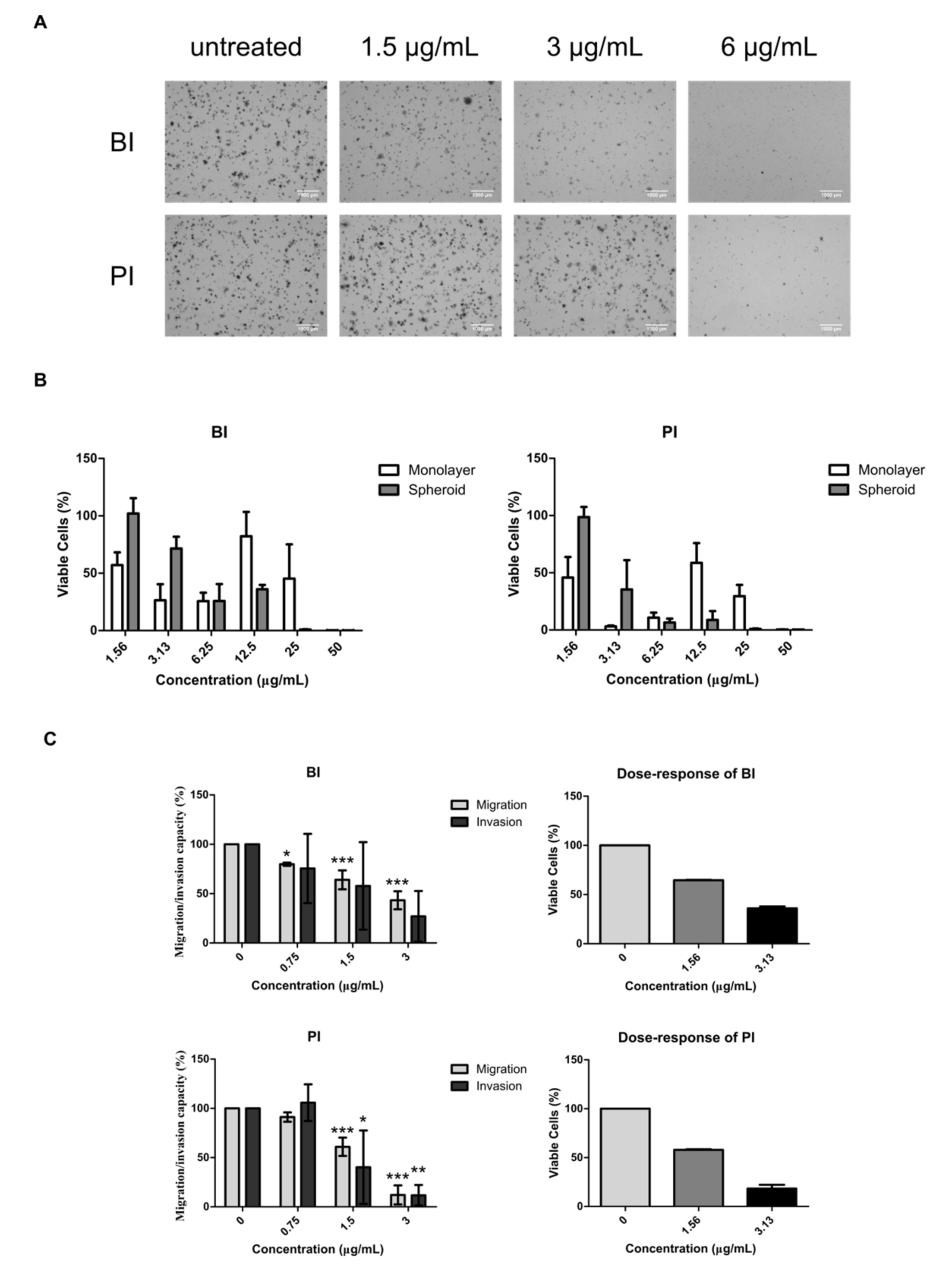
2.4. BI and PI Inhibited the Growth of NCI-H1703’s Soft Agar Colony and Spheroid, But not Migration and Invasion
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Chemicals
4.3. Cell Viability, Cytotoxicity and Anti-Proliferative Assay
4.4. Immunocytochemistry
4.5. Soft Agar Colony Formation Assay
4.6. Multicellular Tumor Spheroid Generation Assay
4.7. Migration and Invasion Assay
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katiyar, C.; Gupta, A.; Kanjilal, S.; Katiyar, S. Drug discovery from plant sources: An integrated approach. Ayu 2012, 33, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zappa, C.; Mousa, S.A. Non-small cell lung cancer: Current treatment and future advances. Transl Lung Cancer Res. 2016, 5, 288–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duma, N.; Santana-Davila, R.; Molina, J.R. Non–Small Cell Lung Cancer: Epidemiology, Screening, Diagnosis, and Treatment. Mayo Clin. Proc. 2019, 94, 1623–1640. [Google Scholar] [CrossRef] [PubMed]
- Shields, B.D.; Mahmoud, F.; Taylor, E.M.; Byrum, S.D.; Sengupta, D.; Koss, B.; Baldini, G.; Ransom, S.; Cline, K.; Mackintosh, S.G.; et al. Indicators of responsiveness to immune checkpoint inhibitors. Sci. Rep. 2017, 7, 807. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Leighl, N.B.; Wu, Y.L.; Zhong, W.Z. Emerging therapies for non-small cell lung cancer. J. Hematol. Oncol. 2019, 12, 45. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.B.; Sullivan, R.J.; Menzies, A.M. Immune checkpoint inhibitors in challenging populations. Cancer 2017, 123, 1904–1911. [Google Scholar] [CrossRef] [Green Version]
- Heng, W.S.; Gosens, R.; Kruyt, F.A.E. Lung cancer stem cells: Origin, features, maintenance mechanisms and therapeutic targeting. Biochem. Pharmacol. 2019, 160, 121–133. [Google Scholar] [CrossRef]
- Rapp, J.; Jaromi, L.; Kvell, K.; Miskei, G.; Pongracz, J.E. WNT signaling–lung cancer is no exception. Respir. Res. 2017, 18. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Wang, H.; Zhang, S. Negative regulators of Wnt signaling in non-small cell lung cancer: Theoretical basis and therapeutic potency. Biomed. Pharmacother. 2019, 118, 109336. [Google Scholar] [CrossRef] [PubMed]
- He, B.; You, L.; Uematsu, K.; Xu, Z.; Lee, A.Y.; Matsangou, M.; McCormick, F.; Jablons, D.M. A Monoclonal Antibody against Wnt-1 Induces Apoptosis in Human Cancer Cells. Neoplasia 2004, 6, 7–14. [Google Scholar] [CrossRef] [Green Version]
- You, L.; He, B.; Xu, Z.; Uematsu, K.; Mazieres, J.; Mikami, I.; Reguart, N.; Moody, T.W.; Kitajewski, J.; McCormick, F.; et al. Inhibition of Wnt-2-mediated signaling induces programmed cell death in non-small-cell lung cancer cells. Oncogene 2004, 23, 6170–6174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nabhan, A.; Brownfield, D.G.; Harbury, P.B.; Krasnow, M.A.; Desai, T.J. Single-cell Wnt signaling niches maintain stemness of alveolar type 2 cells. Science 2018, 359, 1118–1123. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Tammela, T.; Hofree, M.; Choi, J.; Marjanovic, N.D.; Han, S.; Canner, D.; Wu, K.; Paschini, M.; Bhang, D.H.; et al. Anatomically and Functionally Distinct Lung Mesenchymal Populations Marked by Lgr5 and Lgr6. Cell 2017, 170, 1149–1163. [Google Scholar] [CrossRef] [Green Version]
- Stewart, D.J. Wnt Signaling Pathway in Non–Small Cell Lung Cancer. JNCI J. Natl. Cancer Inst. 2014, 106, djt356. [Google Scholar] [CrossRef]
- Sunaga, N.; Kohno, T.; Kolligs, F.T.; Fearon, E.R.; Saito, R.; Yokota, J. Constitutive activation of the Wnt signaling pathway by CTNNB1 (β-catenin) mutations in a subset of human lung adenocarcinoma. Genes Chromosomes Cancer 2001, 30, 316–321. [Google Scholar] [CrossRef]
- Li, X.Q.; Yang, X.L.; Zhang, G.; Wu, S.P.; Deng, X.B.; Xiao, S.J.; Liu, Q.Z.; Yao, K.T.; Xiao, G.H. Nuclear β-catenin accumulation is associated with increased expression of Nanog protein and predicts poor prognosis of non-small cell lung cancer. J. Transl. Med. 2013, 11, 114. [Google Scholar] [CrossRef] [Green Version]
- Shrivastava, S.; Jeengar, M.K.; Thummuri, D.; Koval, A.; Katanaev, V.L.; Marepally, S.; Naidu, V.G.M. Cardamonin, a chalcone, inhibits human triple negative breast cancer cell invasiveness by downregulation of Wnt/β-catenin signaling cascades and reversal of epithelial–mesenchymal transition. Bio. Factors. [CrossRef] [Green Version]
- Fonseca, B.F.; Predes, D.; Cerqueira, D.M.; Reis, A.H.; Amado, N.G.; Cayres, M.C.L.; Kuster, R.M.; Oliveira, F.L.; Mendes, F.A.; Abreu, J.G. Derricin and Derricidin Inhibit Wnt/β-Catenin Signaling and Suppress Colon Cancer Cell Growth In Vitro. PLoS ONE 2015, 10, e0120919. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, T.; Korkaya, H.; Liu, S.; Lee, H.F.; Newman, B.; Yu, Y.; Clouthier, S.G.; Schwartz, S.J.; Wicha, M.S.; et al. Sulforaphane, a Dietary Component of Broccoli/Broccoli Sprouts, Inhibits Breast Cancer Stem Cells. Clin. Cancer Res. 2010, 16, 2580–2590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernkopf, D.B.; Daum, G.; Brückner, M.; Behrens, J. Sulforaphane inhibits growth and blocks Wnt/β-catenin signaling of colorectal cancer cells. Oncotarget 2018, 9, 33982–33994. [Google Scholar] [CrossRef] [Green Version]
- Heng, W.S.; Cheah, S.C. Chelerythrine Chloride Downregulates β-Catenin and Inhibits Stem Cell Properties of Non-Small Cell Lung Carcinoma. Molecules 2020, 25, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borowicz, S.; van Scoyk, M.; Avasarala, S.; Karuppusamy Rathinam, M.K.; Tauler, J.; Bikkavilli, R.K.; Winn, R.A. The Soft Agar Colony Formation Assay. J. Vis. Exp. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivascu, A. Manfred Kubbies Rapid Generation of Single-Tumor Spheroids for High-Throughput Cell Function and Toxicity Analysis. J. Biomol. Screen 2006, 11, 922–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiswald, L.B.; Bellet, D.; Dangles-Marie, V. Spherical Cancer Models in Tumor Biology. Neoplasia 2015, 17, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minchinton, A.I.; Tannock, I.F. Drug penetration in solid tumours. Nat. Rev. Cancer 2006, 6, 583–592. [Google Scholar] [CrossRef]
- Omar, H.R.; Komarova, I.; El-Ghonemi, M.; Fathy, A.; Rashad, R.; Abdelmalak, H.D.; Yerramadha, M.R.; Ali, Y.; Helal, E.; Camporesi, E.M. Licorice abuse: Time to send a warning message. Adv. Endocrinol. Metab. 2012, 3, 125–138. [Google Scholar] [CrossRef]
- Tang, L.; Paonessa, J.D.; Zhang, Y.; Ambrosone, C.B.; McCann, S.E. Total isothiocyanate yield from raw cruciferous vegetables commonly consumed in the United States. J. Funct. Foods 2013, 5, 1996–2001. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Hu, C. Evodiamine: A Novel Anti-Cancer Alkaloid from Evodia rutaecarpa. Molecules 2009, 14, 1852–1859. [Google Scholar] [CrossRef]
- Zielińska, S.; Jezierska-Domaradzka, A.; Wójciak-Kosior, M.; Sowa, I.; Junka, A.; Matkowski, A.M. Greater Celandine’s Ups and Downs−21 Centuries of Medicinal Uses of Chelidonium majus From the Viewpoint of Today’s Pharmacology. Front. Pharm. 2018, 9, 299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Meth. 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Baranwal, S.; Wang, Y.; Rathinam, R.; Lee, J.; Jin, L.; McGoey, R.; Pylayeva, Y.; Giancotti, F.; Blobe, G.C.; Alahari, S.K. Molecular Characterization of the Tumor-Suppressive Function of Nischarin in Breast Cancer. JNCI J. Natl. Cancer Inst. 2011, 103, 1513–1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytochemical Category | Phytochemical Compound | Cell Line | Independent Variables | Results |
|---|---|---|---|---|
| Isothiocyanate | BI | SK-LU-1 | Time | F (184, 1288) = 992.6, p < 0.0001, R2 = 0.08 |
| Treatment | F (6, 1288) = 1095, p < 0.0001, R2 = 0.79 | |||
| Interaction | F (1104, 1288) = 250.8, p < 0.0001, R2 = 0.13 | |||
| HLCSC | Time | F (184, 1288) = 352.5, p < 0.0001, R2 = 0.12 | ||
| Treatment | F (6, 1288) = 301.2, p < 0.0001, R2 = 0.65 | |||
| Interaction | F (1104, 1288) = 105.6, p < 0.0001, R2 = 0.22 | |||
| PI | SK-LU-1 | Time | F (184, 1288) = 195.1, p < 0.0001, R2 = 0.03 | |
| Treatment | F (6, 1288) = 1584, p < 0.0001, R2 = 0.85 | |||
| Interaction | F (1104, 1288) = 116.9, p < 0.0001, R2 = 0.12 | |||
| HLCSC | Time | F (184, 1288) = 4931, p < 0.0001, R2 = 0.16 | ||
| Treatment | F (6, 1288) = 777.7, p < 0.0001, R2 = 0.67 | |||
| Interaction | F (1104, 1288) = 851.8, p < 0.0001, R2 = 0.17 | |||
| Chalcone | LICO | SK-LU-1 | Time | F (184, 1288) = 362.4, p < 0.0001, R2 = 0.11 |
| Treatment | F (6, 1288) = 180.9, p < 0.0001, R2 = 0.78 | |||
| Interaction | F (1104, 1288) = 56.6, p < 0.0001, R2 = 0.10 | |||
| HLCSC | Time | F (184, 1288) = 1362, p < 0.0001, R2 = 0.27 | ||
| Treatment | F (6, 1288) = 201, p < 0.0001, R2 = 0.55 | |||
| Interaction | F (1104, 1288) = 149.5, p < 0.0001, R2 = 0.18 | |||
| ISO | SK-LU-1 | Time | F (184, 1288) = 365.3, p < 0.0001, R2 = 0.12 | |
| Treatment | F (6, 1288) = 307.5, p < 0.0001, R2 = 0.75 | |||
| Interaction | F (1104, 1288) = 62, p < 0.0001, R2 = 0.13 | |||
| HLCSC | Time | F (184, 1288) = 1985, p < 0.0001, R2 = 0.31 | ||
| Treatment | F (6, 1288) = 362.8, p < 0.0001, R2 = 0.52 | |||
| Interaction | F (1104, 1288) = 175.1, p < 0.0001, R2 = 0.17 | |||
| Alkaloid | EVO | SK-LU-1 | Time | F (184, 1288) = 151.7, p < 0.0001, R2 = 0.08 |
| Treatment | F (6, 1288) = 52.7, p < 0.0001, R2 = 0.76 | |||
| Interaction | F (1104, 1288) = 47.9, p < 0.0001, R2 = 0.15 | |||
| HLCSC | Time | F (184, 1288) = 1929, p < 0.0001, R2 = 0.48 | ||
| Treatment | F (6, 1288) = 206.3, p < 0.0001, R2 = 0.33 | |||
| Interaction | F (1104, 1288) = 130.5, p < 0.0001, R2 = 0.19 | |||
| CHE | SK-LU-1 | Time | F (184, 1288) = 59.4, p < 0.0001, R2 = 0.06 | |
| Treatment | F (6, 1288) = 38.9, p < 0.0001, R2 = 0.74 | |||
| Interaction | F (1104, 1288) = 31.4, p < 0.0001, R2 = 0.18 | |||
| HLCSC | Time | F (184, 1288) = 328.3, p < 0.0001, R2 = 0.49 | ||
| Treatment | F (6, 1288) = 121.8, p < 0.0001, R2 = 0.34 | |||
| Interaction | F (1104, 1288) = 17.2, p < 0.0001, R2 = 0.16 | |||
| Positive control | SAL | SK-LU-1 | Time | F (184, 1288) = 71.5, p < 0.0001, R2 = 0.04 |
| Treatment | F (6, 1288) = 80.9, p < 0.0001, R2 = 0.79 | |||
| Interaction | F (1104, 1288) = 47.1, p < 0.0001, R2 = 0.16 | |||
| HLCSC | Time | F (184, 1288) = 933.5, p < 0.0001, R2 = 0.15 | ||
| Treatment | F (6, 1288) = 591.4, p < 0.0001, R2 = 0.65 | |||
| Interaction | F (1104, 1288) = 205.4, p < 0.0001, R2 = 0.20 |
| IC50 Values (mean ± SD) | |||||||
|---|---|---|---|---|---|---|---|
| Phytochemical Category | Phytochemical Compound | SK-LU-1 | |||||
| 24 h | 48 h | 72 h | |||||
| μg/mL | μM | μg/mL | μM | μg/mL | μM | ||
| Isothiocyanate | BI | 3.96 ± 0.27 | 26.54 ± 1.80 | 4.54 ± 1.35 | 30.39 ± 9.05 | 4.99 ± 1.04 | 33.41 ± 6.97 |
| PI | 2.68 ± 0.01 | 16.39 ± 0.04 | 2.69 ± 0.00 | 16.48 ± 0.00 | 2.81 ± 0.04 | 17.18 ± 0.22 | |
| Chalcone | LICO | 12.75 ± 0.38 | 37.68 ± 1.13 | 13.96 ± 1.41 | 41.24 ± 4.16 | 16.28 ± 1.79 | 48.09 ± 5.29 |
| ISO | 10.93 ± 2.28 | 42.63 ± 8.91 | 16.73 ± 0.40 | 65.29 ± 1.55 | 17.11 ± 0.79 | 66.75 ± 3.06 | |
| Alkaloid | EVO | 5.71 ± 0.69 | 18.81 ± 2.26 | 5.41 ± 1.06 | 17.83 ± 3.50 | 4.33 ± 0.36 | 14.26 ± 1.19 |
| CHE | 4.32 ± 0.21 | 12.21 ± 0.58 | 2.86 ± 0.55 | 8.09 ± 1.56 | 2.74 ± 0.47 | 7.74 ± 1.34 | |
| Positive control | SAL | 10.40 ± 2.33 | 13.84 ± 3.10 | 9.96 ± 1.10 | 13.26 ± 1.46 | 10.18 ± 0.33 | 13.56 ± 0.43 |
| Phytochemical Category | Phytochemical Compound | HLCSC | |||||
| 24 h | 48 h | 72 h | |||||
| μg/mL | μM | μg/mL | μM | μg/mL | μM | ||
| Isothiocyanate | BI | 2.88 ± 0.24 | 19.30 ± 1.61 | 2.80 ± 0.11 | 18.73 ± 0.71 | 3.14 ± 0.23 | 21.04 ± 1.52 |
| PI | 5.14 ± 0.20 | 31.49 ± 1.21 | 6.10 ± 0.07 | 37.37 ± 0.43 | 6.27 ± 0.16 | 38.38 ± 1.00 | |
| Chalcone | LICO | 8.27 ± 1.49 | 24.42 ± 4.41 | 8.65 ± 0.63 | 25.55 ± 1.86 | 12.01 ± 0.59 | 35.49 ± 1.76 |
| ISO | 22.25 ± 0.67 | 86.81 ± 2.62 | 23.01 ± 0.12 | 89.76 ± 0.47 | 23.52 ± 0.45 | 91.77 ± 1.74 | |
| Alkaloid | EVO | 4.71 ± 1.49 | 15.53 ± 4.90 | 7.72 ± 0.72 | 25.45 ± 2.38 | 9.13 ± 0.54 | 30.10 ± 1.77 |
| CHE | 8.2 ± 0.44 | 23.21 ± 1.24 | 7.86 ± 0.04 | 22.23 ± 0.10 | 12.57 ± 2.72 | 35.57 ± 7.68 | |
| Positive control | SAL | 3.46 ± 0.35 | 4.61 ± 0.47 | 4.42 ± 1.90 | 5.89 ± 2.52 | 2.57 ± 0.38 | 3.42 ± 0.50 |
| Cell Lines | IC20 (μg/mL) 1 | ||
|---|---|---|---|
| 24 h | 48 h | 72 h | |
| NCI-H1703 | 1.49 ± 0.08 | 1.72 ± 0.07 | 1.83 ± 0.07 |
| SK-LU-1 | 1.87 ± 0.13 | 2.91 ± 0.87 | 3.46 ± 0.72 |
| HLCSC | 1.88 ± 0.16 | 2.65 ± 0.10 | 2.91 ± 0.21 |
| Cell Lines | IC50 (μg/mL) 2 | ||
| 24 h | 48 h | 72 h | |
| NCI-H1703 | 2.90 ± 0.16 | 2.93 ± 0.13 | 2.95 ± 0.11 |
| SK-LU-1 | 3.96 ± 0.27 | 4.54 ± 1.35 | 4.99 ± 1.04 |
| HLCSC | 2.88 ± 0.24 | 2.80 ± 0.11 | 3.14 ± 0.23 |
| Cell Lines | IC80 (μg/mL) 1 | ||
| 24 h | 48 h | 72 h | |
| NCI-H1703 | 5.65 ± 0.30 | 5.01 ± 0.22 | 4.73 ± 0.17 |
| SK-LU-1 | 8.41 ± 0.57 | 7.08 ± 2.11 | 7.19 ± 1.50 |
| HLCSC | 4.41 ± 0.37 | 2.95 ± 0.11 | 3.38 ± 0.24 |
| Cell Lines | IC20 (μg/mL) 1 | ||
|---|---|---|---|
| 24 h | 48 h | 72 h | |
| NCI-H1703 | 1.74 ± 0.14 | 1.81 ± 0.14 | 1.99 ± 0.01 |
| SK-LU-1 | 1.81 ± 0.00 | 2.14 ± 0.00 | 2.03 ± 0.03 |
| HLCSC | 3.81 ± 0.15 | 5.78 ± 0.07 | 5.89 ± 0.15 |
| Cell Lines | IC50 (μg/mL) 2 | ||
| 24 h | 48 h | 72 h | |
| NCI-H1703 | 2.82 ± 0.22 | 2.78 ± 0.21 | 2.92 ± 0.01 |
| SK-LU-1 | 2.68 ± 0.01 | 2.69 ± 0.00 | 2.81 ± 0.04 |
| HLCSC | 5.14 ± 0.20 | 6.10 ± 0.07 | 6.27 ± 0.16 |
| Cell Lines | IC80 (μg/mL) 1 | ||
| 24 h | 48 h | 72 h | |
| NCI-H1703 | 4.55 ± 0.35 | 4.26 ± 0.32 | 4.29 ± 0.02 |
| SK-LU-1 | 3.96 ± 0.01 | 3.38 ± 0.00 | 3.87 ± 0.05 |
| HLCSC | 6.94 ± 0.27 | 6.44 ± 0.07 | 6.66 ± 0.17 |
| Cell Lines | IC20 (μg/mL) 1 | ||
|---|---|---|---|
| 24 h | 48 h | 72 h | |
| NCI-H1703 | 0.85 ± 0.31 | 0.60 ± 0.03 | 0.54 ± 0.05 |
| SK-LU-1 | 2.50 ± 0.56 | 1.22 ± 0.13 | 1.13 ± 0.04 |
| HLCSC | 1.02 ± 0.10 | 0.71 ± 0.30 | 0.26 ± 0.04 |
| Cell Lines | IC50 (μg/mL) 2 | ||
| 24 h | 48 h | 72 h | |
| NCI-H1703 | 11.03 ± 4.03 | 6.00 ± 0.28 | 4.96 ± 0.42 |
| SK-LU-1 | 10.40 ± 2.33 | 9.96 ± 1.10 | 10.18 ± 0.33 |
| HLCSC | 3.46 ± 0.35 | 4.42 ± 1.90 | 2.57 ± 0.37 |
| Cell Lines | IC80 (μg/mL) 1 | ||
| 24 h | 48 h | 72 h | |
| NCI-H1703 | 143.92 ± 52.59 | 60.36 ± 2.78 | 45.37 ± 3.82 |
| SK-LU-1 | 43.17 ± 9.66 | 80.94 ± 8.91 | 91.89 ± 2.94 |
| HLCSC | 11.79 ± 1.20 | 27.64 ± 11.85 | 25.32 ± 3.70 |
Sample Availability: Not available. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heng, W.S.; Cheah, S.-C. Identification of Phytochemical-Based β-Catenin Nuclear Localization Inhibitor in NSCLC: Differential Targeting Population from Member of Isothiocyanates. Molecules 2021, 26, 399. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26020399
Heng WS, Cheah S-C. Identification of Phytochemical-Based β-Catenin Nuclear Localization Inhibitor in NSCLC: Differential Targeting Population from Member of Isothiocyanates. Molecules. 2021; 26(2):399. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26020399
Chicago/Turabian StyleHeng, Win Sen, and Shiau-Chuen Cheah. 2021. "Identification of Phytochemical-Based β-Catenin Nuclear Localization Inhibitor in NSCLC: Differential Targeting Population from Member of Isothiocyanates" Molecules 26, no. 2: 399. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26020399