
SUMO and Transcriptional Regulation: The Lessons of Large-Scale Proteomic, Modifomic and Genomic Studies

 , and
, and 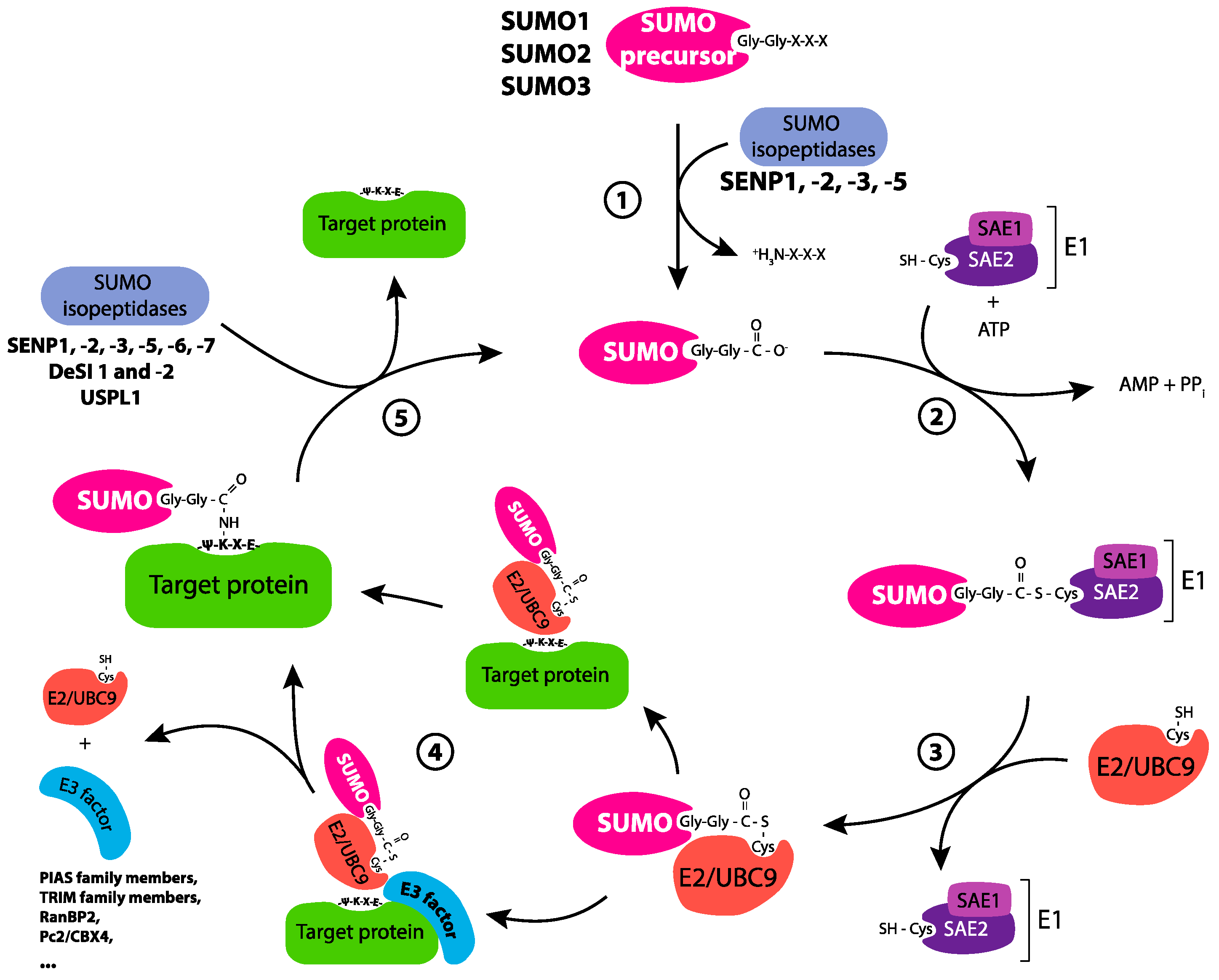{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. SUMOylation: A Principally Nuclear Post-Translational Modification
2.1. SUMO Isoforms and the General Effects of SUMOylation on Proteins
2.2. The SUMOylation Cycle
2.3. SUMO Substrates and Nuclear Functions of SUMOylation
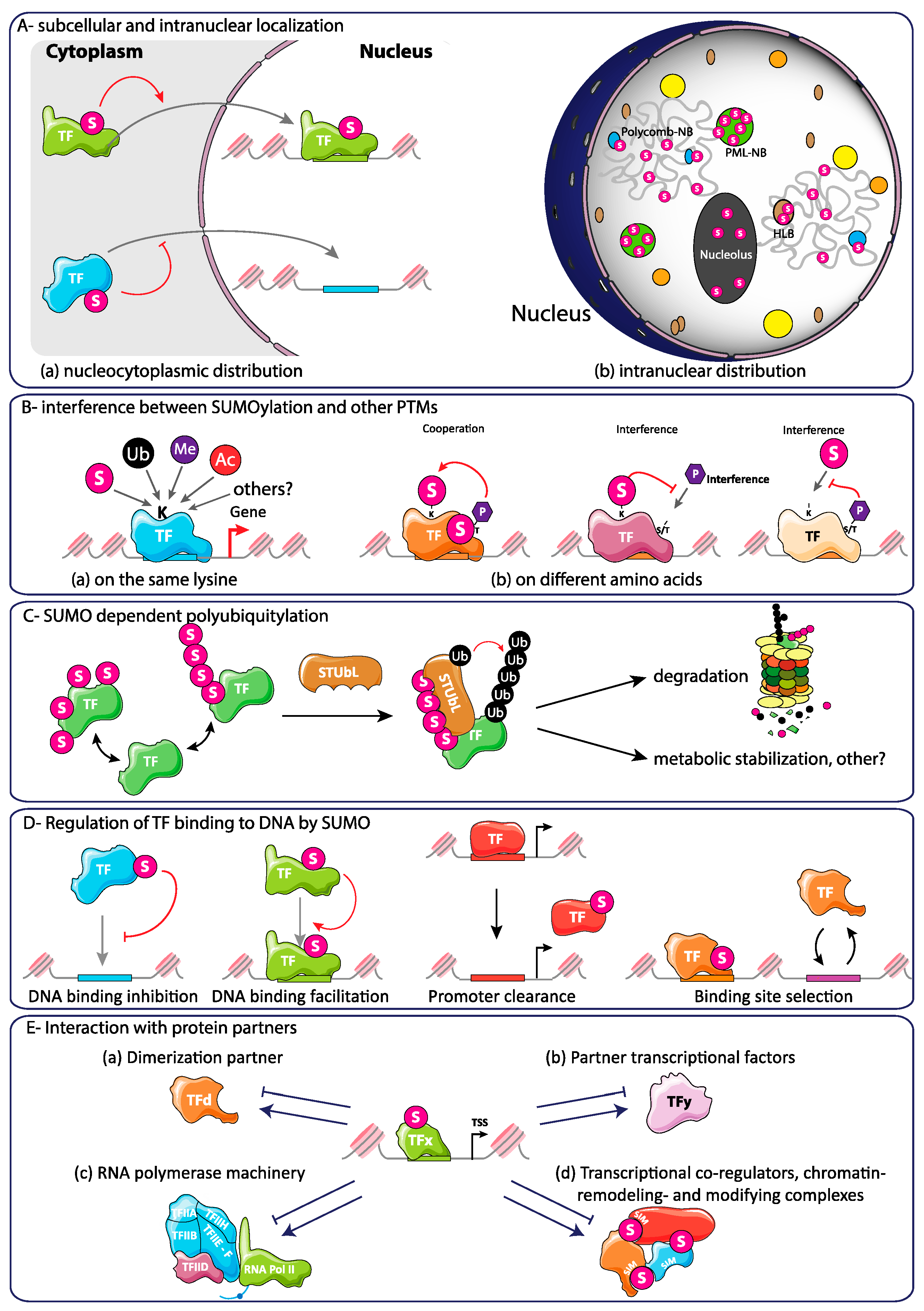
2.4. Cross-Talks and Competition of SUMOylation with Other Post-Translational Modifications: Possible Consequences for Transcription
2.5. The Specific Case of SUMO Crosstalk with Ubiquitin: when SUMO Chains Come into Play
2.6. Can Transcription Involve Group SUMOylation?
2.7. Can SUMO-Dependent Phase Separation Contribute to Transcriptional Regulation?
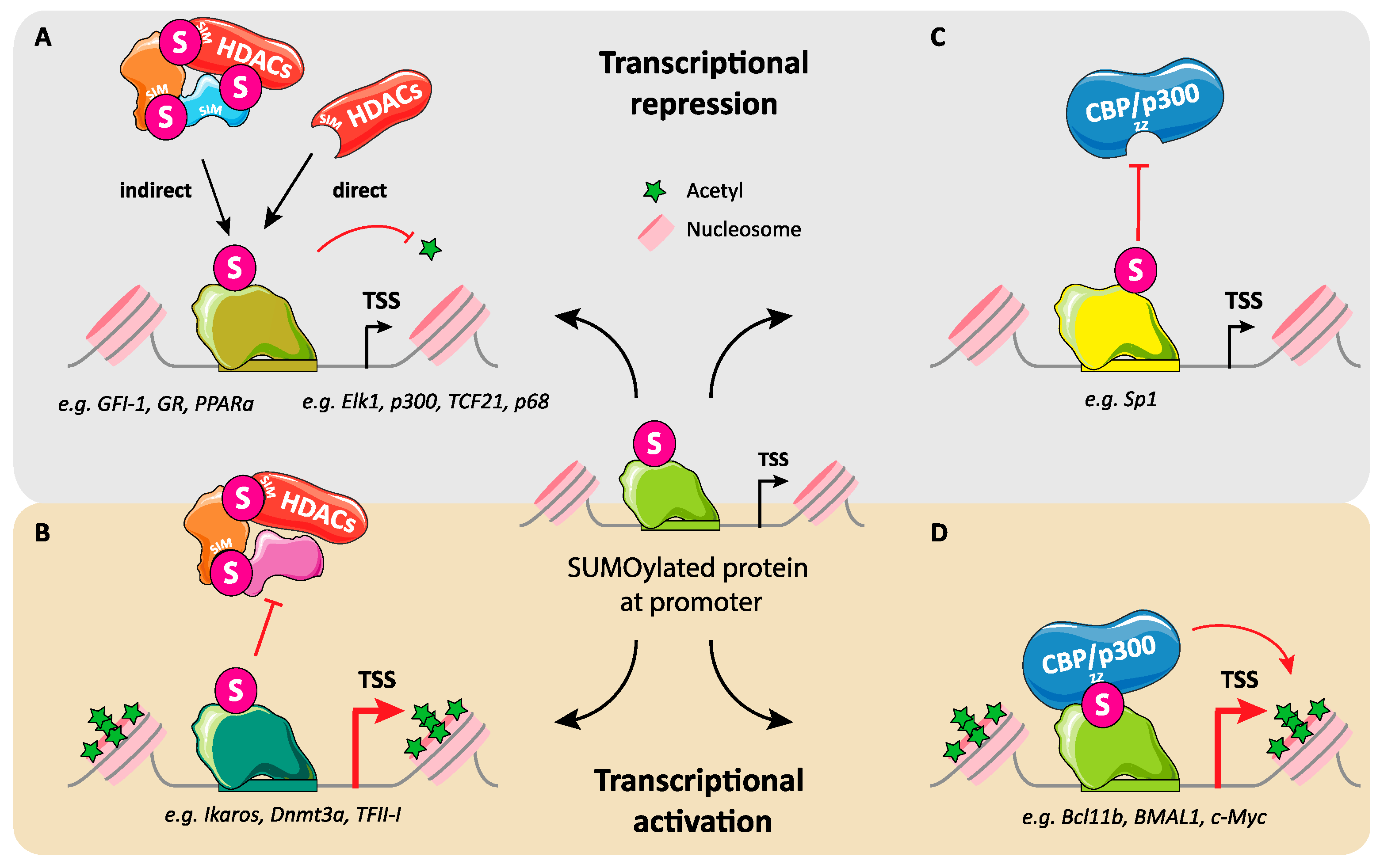
3. Regulation of Transcription by SUMO: Not Just Gene Repression
4. Transcriptional Regulation by SUMO: Pol II-, but Also Pol I- and Pol III-Dependent Transcriptions
5. The General Lessons of Genome-Wide Studies of Chromatin SUMOylation
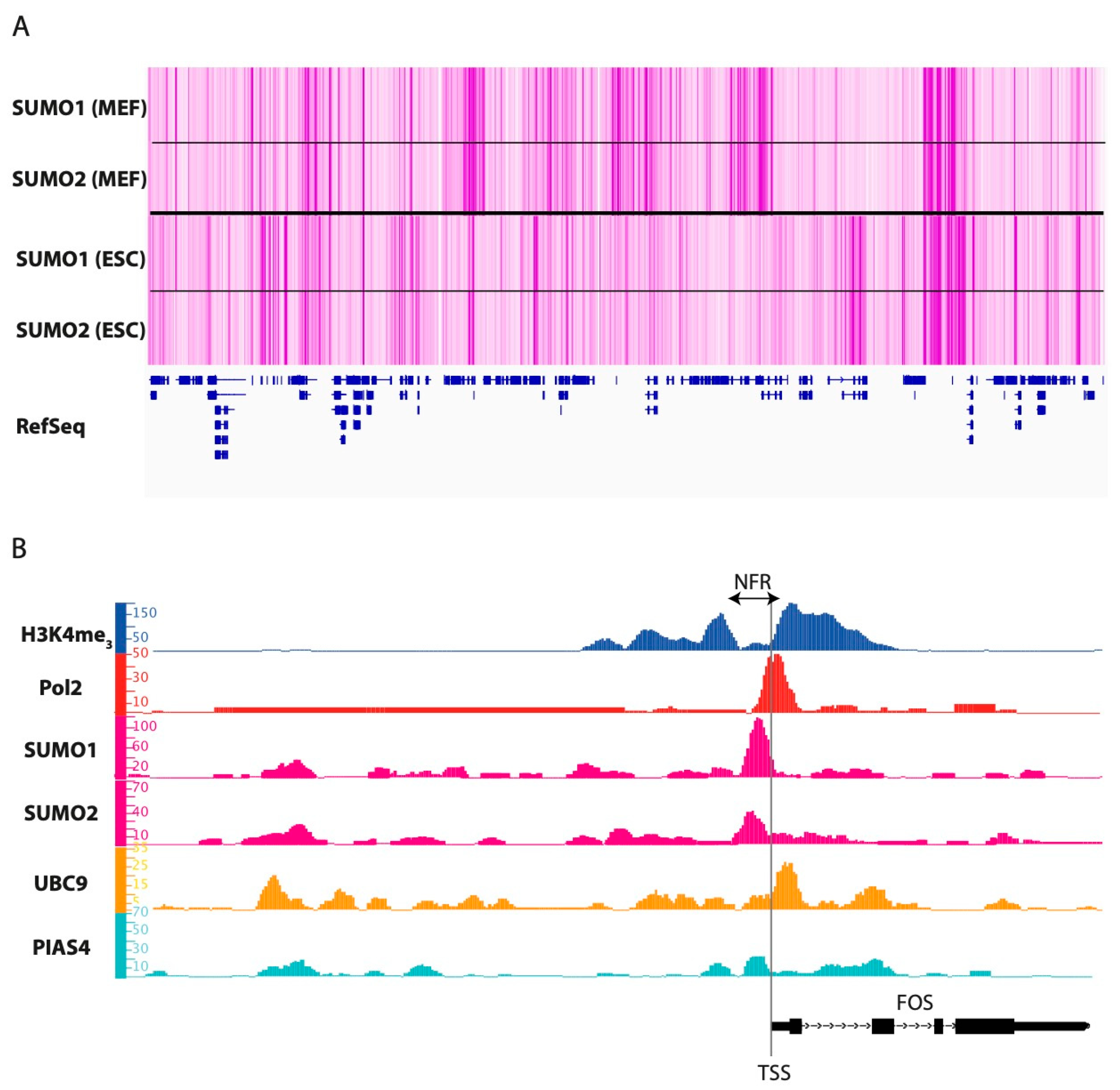
5.1. Where, When and What?
5.2. A Common and Conserved Role for TF SUMOylation in TFBS Selection?
6. Molecular Mechanisms Whereby SUMOylation Can Regulate Pol II-Dependent Gene Expression
6.1. Technical and Biological Limitations when Studying the Regulation of Transcription by SUMO
6.2. SUMOylation-Dependent Alteration of the Intrinsic Properties of TFs
6.3. Regulation of Interactions between Transcription Factors and Transcription Co-Regulators by SUMO
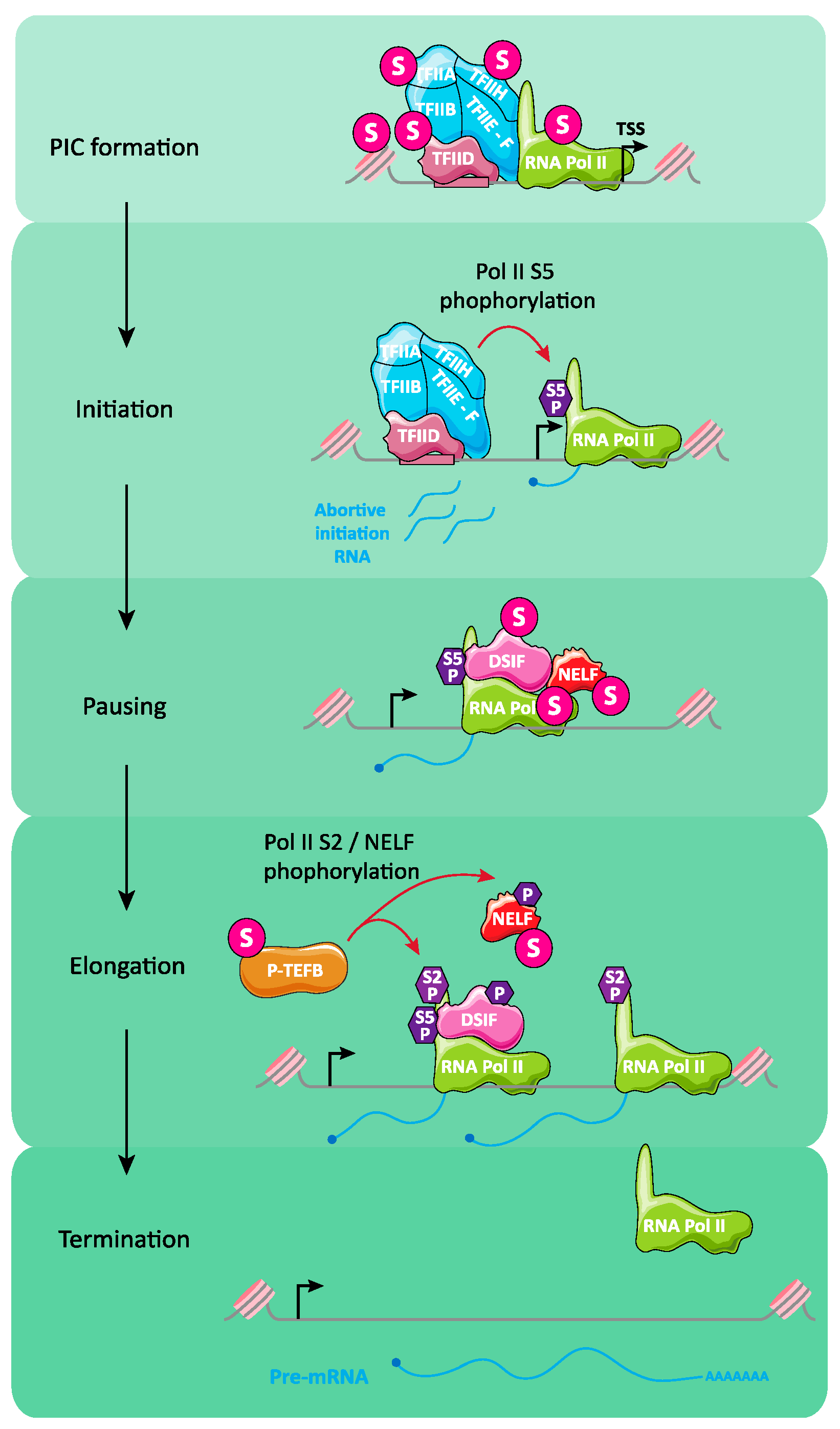
6.4. Regulation of Transcription Initiation and Elongation by SUMO
6.5. SUMO-Dependent Configuration of Nuclear Substructures
6.6. Histone SUMOylation and Transcription
6.7. SUMOylation and Organization of Topologically-Associating Domains
6.8. SUMO in the Establishment of Heterochromatin
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ouyang, J.; Gill, G. SUMO Engages Multiple Corepressors to Regulate Chromatin Structure and Transcription. Epigenetics 2009, 4, 440–444. [Google Scholar] [CrossRef] [Green Version]
- Raman, N.; Nayak, A.; Muller, S. The SUMO System: A Master Organizer of Nuclear Protein Assemblies. Chromosoma 2013, 122, 475–485. [Google Scholar] [CrossRef]
- Chymkowitch, P.; Nguéa, P.A.; Enserink, J.M. SUMO-Regulated Transcription: Challenging the Dogma. BioEssays 2015, 37, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- Rosonina, E.; Akhter, A.; Dou, Y.; Babu, J.; Sri Theivakadadcham, V.S. Regulation of Transcription Factors by Sumoylation. Transcription 2017, 8, 220–231. [Google Scholar] [CrossRef] [Green Version]
- Wotton, D.; Pemberton, L.F.; Merrill-Schools, J. SUMO and Chromatin Remodeling. Adv. Exp. Med. Biol. 2017, 963, 35–50. [Google Scholar] [CrossRef]
- Geiss-Friedlander, R.; Melchior, F. Concepts in Sumoylation: A Decade On. Nat. Rev. Mol. Cell Biol. 2007, 8, 947–956. [Google Scholar] [CrossRef] [PubMed]
- Gareau, J.R.; Lima, C.D. The SUMO Pathway: Emerging Mechanisms That Shape Specificity, Conjugation and Recognition. Nat. Rev. Mol. Cell Biol. 2010, 11, 861–871. [Google Scholar] [CrossRef] [Green Version]
- Flotho, A.; Melchior, F. Sumoylation: A Regulatory Protein Modification in Health and Disease. Annu. Rev. Biochem. 2013, 82, 357–385. [Google Scholar] [CrossRef]
- Matunis, M.J.; Rodriguez, M.S. Concepts and Methodologies to Study Protein SUMOylation: An Overview. Methods Mol. Biol. Clifton NJ 2016, 1475, 3–22. [Google Scholar] [CrossRef]
- Celen, A.B.; Sahin, U. Sumoylation on Its 25th Anniversary: Mechanisms, Pathology, and Emerging Concepts. FEBS J. 2020, 287, 3110–3140. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-M.; Yeh, E.T.H. SUMO: From Bench to Bedside. Physiol. Rev. 2020, 100, 1599–1619. [Google Scholar] [CrossRef]
- Bhat, K.P.; Greer, S.F. Proteolytic and Non-Proteolytic Roles of Ubiquitin and the Ubiquitin Proteasome System in Transcriptional Regulation. Biochim. Biophys. Acta 2011, 1809, 150–155. [Google Scholar] [CrossRef]
- Dwane, L.; Gallagher, W.M.; Ní Chonghaile, T.; O’Connor, D.P. The Emerging Role of Non-Traditional Ubiquitination in Oncogenic Pathways. J. Biol. Chem. 2017, 292, 3543–3551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weissman, A.M.; Shabek, N.; Ciechanover, A. The Predator Becomes the Prey: Regulating the Ubiquitin System by Ubiquitylation and Degradation. Nat. Rev. Mol. Cell Biol. 2011, 12, 605–620. [Google Scholar] [CrossRef] [PubMed]
- Nagaraj, N.; Wisniewski, J.R.; Geiger, T.; Cox, J.; Kircher, M.; Kelso, J.; Pääbo, S.; Mann, M. Deep Proteome and Transcriptome Mapping of a Human Cancer Cell Line. Mol. Syst. Biol. 2011, 7, 548. [Google Scholar] [CrossRef]
- Graña-Montes, R.; Marinelli, P.; Reverter, D.; Ventura, S. N-Terminal Protein Tails Act as Aggregation Protective Entropic Bristles: The SUMO Case. Biomacromolecules 2014, 15, 1194–1203. [Google Scholar] [CrossRef]
- Liebelt, F.; Sebastian, R.M.; Moore, C.L.; Mulder, M.P.C.; Ovaa, H.; Shoulders, M.D.; Vertegaal, A.C.O. SUMOylation and the HSF1-Regulated Chaperone Network Converge to Promote Proteostasis in Response to Heat Shock. Cell Rep. 2019, 26, 236–249.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Droescher, M.; Begitt, A.; Marg, A.; Zacharias, M.; Vinkemeier, U. Cytokine-Induced Paracrystals Prolong the Activity of Signal Transducers and Activators of Transcription (STAT) and Provide a Model for the Regulation of Protein Solubility by Small Ubiquitin-like Modifier (SUMO). J. Biol. Chem. 2011, 286, 18731–18746. [Google Scholar] [CrossRef] [Green Version]
- Castro, P.H.; Tavares, R.M.; Bejarano, E.R.; Azevedo, H. SUMO, a Heavyweight Player in Plant Abiotic Stress Responses. Cell. Mol. Life Sci. CMLS 2012, 69, 3269–3283. [Google Scholar] [CrossRef]
- Ureña, E.; Pirone, L.; Chafino, S.; Pérez, C.; Sutherland, J.D.; Lang, V.; Rodriguez, M.S.; Lopitz-Otsoa, F.; Blanco, F.J.; Barrio, R.; et al. Evolution of SUMO Function and Chain Formation in Insects. Mol. Biol. Evol. 2016, 33, 568–584. [Google Scholar] [CrossRef]
- Vertegaal, A.C.O.; Andersen, J.S.; Ogg, S.C.; Hay, R.T.; Mann, M.; Lamond, A.I. Distinct and Overlapping Sets of SUMO-1 and SUMO-2 Target Proteins Revealed by Quantitative Proteomics. Mol. Cell. Proteom. MCP 2006, 5, 2298–2310. [Google Scholar] [CrossRef] [Green Version]
- Seifert, A.; Schofield, P.; Barton, G.J.; Hay, R.T. Proteotoxic Stress Reprograms the Chromatin Landscape of SUMO Modification. Sci. Signal. 2015, 8, rs7. [Google Scholar] [CrossRef] [Green Version]
- Kunz, K.; Piller, T.; Müller, S. SUMO-Specific Proteases and Isopeptidases of the SENP Family at a Glance. J. Cell Sci. 2018, 131. [Google Scholar] [CrossRef] [Green Version]
- Talamillo, A.; Barroso-Gomila, O.; Giordano, I.; Ajuria, L.; Grillo, M.; Mayor, U.; Barrio, R. The Role of SUMOylation during Development. Biochem. Soc. Trans. 2020, 48, 463–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Sternsdorf, T.; Bolger, T.A.; Evans, R.M.; Yao, T.-P. Regulation of MEF2 by Histone Deacetylase 4- and SIRT1 Deacetylase-Mediated Lysine Modifications. Mol. Cell. Biol. 2005, 25, 8456–8464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, C.; Ho, C.-C.; Reineke, E.; Lam, M.; Cheng, X.; Stanya, K.J.; Liu, Y.; Chakraborty, S.; Shih, H.-M.; Kao, H.-Y. Histone Deacetylase 7 Promotes PML Sumoylation and Is Essential for PML Nuclear Body Formation. Mol. Cell. Biol. 2008, 28, 5658–5667. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Tse, A.K.-W.; Li, P.; Ma, Q.; Xiang, S.; Nicosia, S.V.; Seto, E.; Zhang, X.; Bai, W. Inhibition of Androgen Receptor Activity by Histone Deacetylase 4 through Receptor SUMOylation. Oncogene 2011, 30, 2207–2218. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, D.; Moriuchi, T.; Osumi, T.; Hirose, F. Transcription Factor HDREF Is a Novel SUMO E3 Ligase of Mi2α. J. Biol. Chem. 2016, 291, 11619–11634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenhardt, N.; Chaugule, V.K.; Koidl, S.; Droescher, M.; Dogan, E.; Rettich, J.; Sutinen, P.; Imanishi, S.Y.; Hofmann, K.; Palvimo, J.J.; et al. A New Vertebrate SUMO Enzyme Family Reveals Insights into SUMO-Chain Assembly. Nat. Struct. Mol. Biol. 2015, 22, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Jansen, N.S.; Vertegaal, A.C.O. A Chain of Events: Regulating Target Proteins by SUMO Polymers. Trends Biochem. Sci. 2020. [Google Scholar] [CrossRef]
- Keiten-Schmitz, J.; Schunck, K.; Müller, S. SUMO Chains Rule on Chromatin Occupancy. Front. Cell Dev. Biol. 2019, 7, 343. [Google Scholar] [CrossRef]
- Aichem, A.; Sailer, C.; Ryu, S.; Catone, N.; Stankovic-Valentin, N.; Schmidtke, G.; Melchior, F.; Stengel, F.; Groettrup, M. The Ubiquitin-like Modifier FAT10 Interferes with SUMO Activation. Nat. Commun. 2019, 10, 4452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herhaus, L.; Dikic, I. Expanding the Ubiquitin Code through Post-Translational Modification. EMBO Rep. 2015, 16, 1071–1083. [Google Scholar] [CrossRef] [Green Version]
- Pérez Berrocal, D.A.; Witting, K.F.; Ovaa, H.; Mulder, M.P.C. Hybrid Chains: A Collaboration of Ubiquitin and Ubiquitin-Like Modifiers Introducing Cross-Functionality to the Ubiquitin Code. Front. Chem. 2019, 7, 931. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Luo, Z.-Q. Post-Translational Regulation of Ubiquitin Signaling. J. Cell Biol. 2019, 218, 1776–1786. [Google Scholar] [CrossRef] [PubMed]
- Yau, R.; Rape, M. The Increasing Complexity of the Ubiquitin Code. Nat. Cell Biol. 2016, 18, 579–586. [Google Scholar] [CrossRef]
- Hendriks, I.A.; Vertegaal, A.C.O. A Comprehensive Compilation of SUMO Proteomics. Nat. Rev. Mol. Cell Biol. 2016, 17, 581–595. [Google Scholar] [CrossRef]
- Hendriks, I.A.; Lyon, D.; Young, C.; Jensen, L.J.; Vertegaal, A.C.O.; Nielsen, M.L. Site-Specific Mapping of the Human SUMO Proteome Reveals Co-Modification with Phosphorylation. Nat. Struct. Mol. Biol. 2017, 24, 325–336. [Google Scholar] [CrossRef]
- Ptak, C.; Wozniak, R.W. SUMO and Nucleocytoplasmic Transport. Adv. Exp. Med. Biol. 2017, 963, 111–126. [Google Scholar] [CrossRef]
- Dantuma, N.P.; van Attikum, H. Spatiotemporal Regulation of Posttranslational Modifications in the DNA Damage Response. EMBO J. 2016, 35, 6–23. [Google Scholar] [CrossRef] [Green Version]
- Dhingra, N.; Zhao, X. Intricate SUMO-Based Control of the Homologous Recombination Machinery. Genes Dev. 2019, 33, 1346–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, P.; Vethantham, V.; Manley, J.L. Roles of Sumoylation in MRNA Processing and Metabolism. Adv. Exp. Med. Biol. 2017, 963, 15–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, N.R.; Hochstrasser, M. The Regulation of Chromatin by Dynamic SUMO Modifications. Methods Mol. Biol. Clifton NJ 2016, 1475, 23–38. [Google Scholar] [CrossRef]
- Augustine, R.C.; Vierstra, R.D. SUMOylation: Re-Wiring the Plant Nucleus during Stress and Development. Curr. Opin. Plant. Biol. 2018, 45, 143–154. [Google Scholar] [CrossRef]
- Liebelt, F.; Schimmel, J.; Verlaan-de Vries, M.; Klemann, E.; van Royen, M.E.; van der Weegen, Y.; Luijsterburg, M.S.; Mullenders, L.H.; Pines, A.; Vermeulen, W.; et al. Transcription-Coupled Nucleotide Excision Repair Is Coordinated by Ubiquitin and SUMO in Response to Ultraviolet Irradiation. Nucleic Acids Res. 2020, 48, 231–248. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Xu, X.; Chang, C.-W.; Zheng, L.; Shen, B.; Liu, Y. SUMO2 Conjugation of PCNA Facilitates Chromatin Remodeling to Resolve Transcription-Replication Conflicts. Nat. Commun. 2018, 9, 2706. [Google Scholar] [CrossRef] [Green Version]
- Texari, L.; Dieppois, G.; Vinciguerra, P.; Contreras, M.P.; Groner, A.; Letourneau, A.; Stutz, F. The Nuclear Pore Regulates GAL1 Gene Transcription by Controlling the Localization of the SUMO Protease Ulp1. Mol. Cell 2013, 51, 807–818. [Google Scholar] [CrossRef] [Green Version]
- Finkbeiner, E.; Haindl, M.; Muller, S. The SUMO System Controls Nucleolar Partitioning of a Novel Mammalian Ribosome Biogenesis Complex. EMBO J. 2011, 30, 1067–1078. [Google Scholar] [CrossRef]
- Del Prete, S.; Mikulski, P.; Schubert, D.; Gaudin, V. One, Two, Three: Polycomb Proteins Hit All Dimensions of Gene Regulation. Genes 2015, 6, 520–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuettengruber, B.; Bourbon, H.-M.; Di Croce, L.; Cavalli, G. Genome Regulation by Polycomb and Trithorax: 70 Years and Counting. Cell 2017, 171, 34–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duronio, R.J.; Marzluff, W.F. Coordinating Cell Cycle-Regulated Histone Gene Expression through Assembly and Function of the Histone Locus Body. RNA Biol. 2017, 14, 726–738. [Google Scholar] [CrossRef] [PubMed]
- Lallemand-Breitenbach, V.; de Thé, H. PML Nuclear Bodies: From Architecture to Function. Curr. Opin. Cell Biol. 2018, 52, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Golebiowski, F.; Matic, I.; Tatham, M.H.; Cole, C.; Yin, Y.; Nakamura, A.; Cox, J.; Barton, G.J.; Mann, M.; Hay, R.T. System-Wide Changes to SUMO Modifications in Response to Heat Shock. Sci. Signal. 2009, 2, ra24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendriks, I.A.; D’Souza, R.C.J.; Yang, B.; Verlaan-de Vries, M.; Mann, M.; Vertegaal, A.C.O. Uncovering Global SUMOylation Signaling Networks in a Site-Specific Manner. Nat. Struct. Mol. Biol. 2014, 21, 927–936. [Google Scholar] [CrossRef] [Green Version]
- Schimmel, J.; Eifler, K.; Sigurðsson, J.O.; Cuijpers, S.A.G.; Hendriks, I.A.; Verlaan-de Vries, M.; Kelstrup, C.D.; Francavilla, C.; Medema, R.H.; Olsen, J.V.; et al. Uncovering SUMOylation Dynamics during Cell-Cycle Progression Reveals FoxM1 as a Key Mitotic SUMO Target Protein. Mol. Cell 2014, 53, 1053–1066. [Google Scholar] [CrossRef] [Green Version]
- Becker, J.; Barysch, S.V.; Karaca, S.; Dittner, C.; Hsiao, H.-H.; Berriel Diaz, M.; Herzig, S.; Urlaub, H.; Melchior, F. Detecting Endogenous SUMO Targets in Mammalian Cells and Tissues. Nat. Struct. Mol. Biol. 2013, 20, 525–531. [Google Scholar] [CrossRef]
- Tammsalu, T.; Matic, I.; Jaffray, E.G.; Ibrahim, A.F.M.; Tatham, M.H.; Hay, R.T. Proteome-Wide Identification of SUMO Modification Sites by Mass Spectrometry. Nat. Protoc. 2015, 10, 1374–1388. [Google Scholar] [CrossRef]
- Xiao, Z.; Chang, J.-G.; Hendriks, I.A.; Sigurðsson, J.O.; Olsen, J.V.; Vertegaal, A.C.O. System-Wide Analysis of SUMOylation Dynamics in Response to Replication Stress Reveals Novel Small Ubiquitin-like Modified Target Proteins and Acceptor Lysines Relevant for Genome Stability. Mol. Cell. Proteom. MCP 2015, 14, 1419–1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendriks, I.A.; Treffers, L.W.; Verlaan-de Vries, M.; Olsen, J.V.; Vertegaal, A.C.O. SUMO-2 Orchestrates Chromatin Modifiers in Response to DNA Damage. Cell Rep. 2015, 10, 1778–1791. [Google Scholar] [CrossRef] [Green Version]
- Impens, F.; Radoshevich, L.; Cossart, P.; Ribet, D. Mapping of SUMO Sites and Analysis of SUMOylation Changes Induced by External Stimuli. Proc. Natl. Acad. Sci. USA 2014, 111, 12432–12437. [Google Scholar] [CrossRef] [Green Version]
- Lamoliatte, F.; Caron, D.; Durette, C.; Mahrouche, L.; Maroui, M.A.; Caron-Lizotte, O.; Bonneil, E.; Chelbi-Alix, M.K.; Thibault, P. Large-Scale Analysis of Lysine SUMOylation by SUMO Remnant Immunoaffinity Profiling. Nat. Commun. 2014, 5, 5409. [Google Scholar] [CrossRef] [Green Version]
- Hendriks, I.A.; D’Souza, R.C.; Chang, J.-G.; Mann, M.; Vertegaal, A.C.O. System-Wide Identification of Wild-Type SUMO-2 Conjugation Sites. Nat. Commun. 2015, 6, 7289. [Google Scholar] [CrossRef] [PubMed]
- Schimmel, J.; Larsen, K.M.; Matic, I.; van Hagen, M.; Cox, J.; Mann, M.; Andersen, J.S.; Vertegaal, A.C.O. The Ubiquitin-Proteasome System Is a Key Component of the SUMO-2/3 Cycle. Mol. Cell. Proteom. MCP 2008, 7, 2107–2122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bursomanno, S.; Beli, P.; Khan, A.M.; Minocherhomji, S.; Wagner, S.A.; Bekker-Jensen, S.; Mailand, N.; Choudhary, C.; Hickson, I.D.; Liu, Y. Proteome-Wide Analysis of SUMO2 Targets in Response to Pathological DNA Replication Stress in Human Cells. DNA Repair 2015, 25, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Schou, J.; Kelstrup, C.D.; Hayward, D.G.; Olsen, J.V.; Nilsson, J. Comprehensive Identification of SUMO2/3 Targets and Their Dynamics during Mitosis. PLoS ONE 2014, 9, e100692. [Google Scholar] [CrossRef] [Green Version]
- Matic, I.; Schimmel, J.; Hendriks, I.A.; van Santen, M.A.; van de Rijke, F.; van Dam, H.; Gnad, F.; Mann, M.; Vertegaal, A.C.O. Site-Specific Identification of SUMO-2 Targets in Cells Reveals an Inverted SUMOylation Motif and a Hydrophobic Cluster SUMOylation Motif. Mol. Cell 2010, 39, 641–652. [Google Scholar] [CrossRef]
- Matafora, V.; D’Amato, A.; Mori, S.; Blasi, F.; Bachi, A. Proteomics Analysis of Nucleolar SUMO-1 Target Proteins upon Proteasome Inhibition. Mol. Cell. Proteom. MCP 2009, 8, 2243–2255. [Google Scholar] [CrossRef] [Green Version]
- Bruderer, R.; Tatham, M.H.; Plechanovova, A.; Matic, I.; Garg, A.K.; Hay, R.T. Purification and Identification of Endogenous PolySUMO Conjugates. EMBO Rep. 2011, 12, 142–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatham, M.H.; Matic, I.; Mann, M.; Hay, R.T. Comparative Proteomic Analysis Identifies a Role for SUMO in Protein Quality Control. Sci. Signal. 2011, 4, rs4. [Google Scholar] [CrossRef] [Green Version]
- Sohn, S.-Y.; Bridges, R.G.; Hearing, P. Proteomic Analysis of Ubiquitin-like Posttranslational Modifications Induced by the Adenovirus E4-ORF3 Protein. J. Virol. 2015, 89, 1744–1755. [Google Scholar] [CrossRef] [Green Version]
- Tu, J.; Chen, Y.; Cai, L.; Xu, C.; Zhang, Y.; Chen, Y.; Zhang, C.; Zhao, J.; Cheng, J.; Xie, H.; et al. Functional Proteomics Study Reveals SUMOylation of TFII-I Is Involved in Liver Cancer Cell Proliferation. J. Proteome Res. 2015, 14, 2385–2397. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; McManus, F.P.; Plutoni, C.; Pascariu, C.M.; Nelson, T.; Alberici Delsin, L.E.; Emery, G.; Thibault, P. Quantitative SUMO Proteomics Identifies PIAS1 Substrates Involved in Cell Migration and Motility. Nat. Commun. 2020, 11, 834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gärtner, A.; Wagner, K.; Hölper, S.; Kunz, K.; Rodriguez, M.S.; Müller, S. Acetylation of SUMO2 at Lysine 11 Favors the Formation of Non-Canonical SUMO Chains. EMBO Rep. 2018, 19, e46117. [Google Scholar] [CrossRef] [PubMed]
- McManus, F.P.; Bourdeau, V.; Acevedo, M.; Lopes-Paciencia, S.; Mignacca, L.; Lamoliatte, F.; Rojas Pino, J.W.; Ferbeyre, G.; Thibault, P. Quantitative SUMO Proteomics Reveals the Modulation of Several PML Nuclear Body Associated Proteins and an Anti-Senescence Function of UBC9. Sci. Rep. 2018, 8, 7754. [Google Scholar] [CrossRef]
- Rytz, T.C.; Miller, M.J.; McLoughlin, F.; Augustine, R.C.; Marshall, R.S.; Juan, Y.-T.; Charng, Y.-Y.; Scalf, M.; Smith, L.M.; Vierstra, R.D. SUMOylome Profiling Reveals a Diverse Array of Nuclear Targets Modified by the SUMO Ligase SIZ1 during Heat Stress. Plant. Cell 2018, 30, 1077–1099. [Google Scholar] [CrossRef]
- Maroui, M.A.; Maarifi, G.; McManus, F.P.; Lamoliatte, F.; Thibault, P.; Chelbi-Alix, M.K. Promyelocytic Leukemia Protein (PML) Requirement for Interferon-Induced Global Cellular SUMOylation. Mol. Cell. Proteom. MCP 2018, 17, 1196–1208. [Google Scholar] [CrossRef] [Green Version]
- Uzoma, I.; Hu, J.; Cox, E.; Xia, S.; Zhou, J.; Rho, H.-S.; Guzzo, C.; Paul, C.; Ajala, O.; Goodwin, C.R.; et al. Global Identification of Small Ubiquitin-Related Modifier (SUMO) Substrates Reveals Crosstalk between SUMOylation and Phosphorylation Promotes Cell Migration. Mol. Cell. Proteom. MCP 2018, 17, 871–888. [Google Scholar] [CrossRef] [Green Version]
- Drabikowski, K.; Ferralli, J.; Kistowski, M.; Oledzki, J.; Dadlez, M.; Chiquet-Ehrismann, R. Comprehensive List of SUMO Targets in Caenorhabditis Elegans and Its Implication for Evolutionary Conservation of SUMO Signaling. Sci. Rep. 2018, 8, 1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McManus, F.P.; Lamoliatte, F.; Thibault, P. Identification of Cross Talk between SUMOylation and Ubiquitylation Using a Sequential Peptide Immunopurification Approach. Nat. Protoc. 2017, 12, 2342–2358. [Google Scholar] [CrossRef]
- Munk, S.; Sigurðsson, J.O.; Xiao, Z.; Batth, T.S.; Franciosa, G.; von Stechow, L.; Lopez-Contreras, A.J.; Vertegaal, A.C.O.; Olsen, J.V. Proteomics Reveals Global Regulation of Protein SUMOylation by ATM and ATR Kinases during Replication Stress. Cell Rep. 2017, 21, 546–558. [Google Scholar] [CrossRef] [Green Version]
- Namuduri, A.V.; Heras, G.; Mi, J.; Cacciani, N.; Hörnaeus, K.; Konzer, A.; Lind, S.B.; Larsson, L.; Gastaldello, S. A Proteomic Approach to Identify Alterations in the Small Ubiquitin-like Modifier (SUMO) Network during Controlled Mechanical Ventilation in Rat Diaphragm Muscle. Mol. Cell. Proteom. MCP 2017, 16, 1081–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, L.; Tu, J.; Song, L.; Gao, Z.; Li, K.; Wang, Y.; Liu, Y.; Zhong, F.; Ge, R.; Qin, J.; et al. Proteome-Wide Mapping of Endogenous SUMOylation Sites in Mouse Testis. Mol. Cell. Proteom. MCP 2017, 16, 717–727. [Google Scholar] [CrossRef] [Green Version]
- Cox, E.; Hwang, W.; Uzoma, I.; Hu, J.; Guzzo, C.M.; Jeong, J.; Matunis, M.J.; Qian, J.; Zhu, H.; Blackshaw, S. Global Analysis of SUMO-Binding Proteins Identifies SUMOylation as a Key Regulator of the INO80 Chromatin Remodeling Complex. Mol. Cell. Proteom. MCP 2017, 16, 812–823. [Google Scholar] [CrossRef] [Green Version]
- Lamoliatte, F.; McManus, F.P.; Maarifi, G.; Chelbi-Alix, M.K.; Thibault, P. Uncovering the SUMOylation and Ubiquitylation Crosstalk in Human Cells Using Sequential Peptide Immunopurification. Nat. Commun. 2017, 8, 14109. [Google Scholar] [CrossRef] [Green Version]
- Domingues, P.; Golebiowski, F.; Tatham, M.H.; Lopes, A.M.; Taggart, A.; Hay, R.T.; Hale, B.G. Global Reprogramming of Host SUMOylation during Influenza Virus Infection. Cell Rep. 2015, 13, 1467–1480. [Google Scholar] [CrossRef] [Green Version]
- Galisson, F.; Mahrouche, L.; Courcelles, M.; Bonneil, E.; Meloche, S.; Chelbi-Alix, M.K.; Thibault, P. A Novel Proteomics Approach to Identify SUMOylated Proteins and Their Modification Sites in Human Cells. Mol. Cell. Proteom. MCP 2011, 10, M110.004796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theurillat, I.; Hendriks, I.A.; Cossec, J.-C.; Andrieux, A.; Nielsen, M.L.; Dejean, A. Extensive SUMO Modification of Repressive Chromatin Factors Distinguishes Pluripotent from Somatic Cells. Cell Rep. 2020, 33, 108251. [Google Scholar] [CrossRef]
- Pirone, L.; Xolalpa, W.; Mayor, U.; Barrio, R.; Sutherland, J.D. Analysis of SUMOylated Proteins in Cells and In Vivo Using the BioSUMO Strategy. Methods Mol. Biol. Clifton NJ 2016, 1475, 161–169. [Google Scholar] [CrossRef]
- Handu, M.; Kaduskar, B.; Ravindranathan, R.; Soory, A.; Giri, R.; Elango, V.B.; Gowda, H.; Ratnaparkhi, G.S. SUMO-Enriched Proteome for Drosophila Innate Immune Response. G3 Bethesda Md 2015, 5, 2137–2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosonina, E. A Conserved Role for Transcription Factor Sumoylation in Binding-Site Selection. Curr. Genet. 2019, 65, 1307–1312. [Google Scholar] [CrossRef]
- Lis, J.T. A 50 Year History of Technologies That Drove Discovery in Eukaryotic Transcription Regulation. Nat. Struct. Mol. Biol. 2019, 26, 777–782. [Google Scholar] [CrossRef] [PubMed]
- Roeder, R.G. 50+ Years of Eukaryotic Transcription: An Expanding Universe of Factors and Mechanisms. Nat. Struct. Mol. Biol. 2019, 26, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Desterro, J.M.P.; Rodriguez, M.S.; Hay, R.T. SUMO-1 Modification of IκBα Inhibits NF-ΚB Activation. Mol. Cell 1998, 2, 233–239. [Google Scholar] [CrossRef]
- Wei, F.; Schöler, H.R.; Atchison, M.L. Sumoylation of Oct4 Enhances Its Stability, DNA Binding, and Transactivation. J. Biol. Chem. 2007, 282, 21551–21560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.-B.; Ruan, C.-C.; Chen, D.-R.; Zhang, K.; Yan, C.; Gao, P.-J. Activating Transcription Factor 3 SUMOylation Is Involved in Angiotensin II-Induced Endothelial Cell Inflammation and Dysfunction. J. Mol. Cell. Cardiol. 2016, 92, 149–157. [Google Scholar] [CrossRef]
- Van Rechem, C.; Boulay, G.; Pinte, S.; Stankovic-Valentin, N.; Guérardel, C.; Leprince, D. Differential Regulation of HIC1 Target Genes by CtBP and NuRD, via an Acetylation/SUMOylation Switch, in Quiescent versus Proliferating Cells. Mol. Cell. Biol. 2010, 30, 4045–4059. [Google Scholar] [CrossRef] [Green Version]
- Escobar-Ramirez, A.; Vercoutter-Edouart, A.-S.; Mortuaire, M.; Huvent, I.; Hardivillé, S.; Hoedt, E.; Lefebvre, T.; Pierce, A. Modification by SUMOylation Controls Both the Transcriptional Activity and the Stability of Delta-Lactoferrin. PLoS ONE 2015, 10, e0129965. [Google Scholar] [CrossRef] [PubMed]
- Urvalek, A.M.; Lu, H.; Wang, X.; Li, T.; Yu, L.; Zhu, J.; Lin, Q.; Zhao, J. Regulation of the Oncoprotein KLF8 by a Switch between Acetylation and Sumoylation. Am. J. Transl. Res. 2011, 3, 121–132. [Google Scholar]
- Van Nguyen, T.; Angkasekwinai, P.; Dou, H.; Lin, F.-M.; Lu, L.-S.; Cheng, J.; Chin, Y.E.; Dong, C.; Yeh, E.T.H. SUMO-Specific Protease 1 Is Critical for Early Lymphoid Development through Regulation of STAT5 Activation. Mol. Cell 2012, 45, 210–221. [Google Scholar] [CrossRef]
- Grégoire, S.; Yang, X.-J. Association with Class IIa Histone Deacetylases Upregulates the Sumoylation of MEF2 Transcription Factors. Mol. Cell. Biol. 2005, 25, 2273–2287. [Google Scholar] [CrossRef] [Green Version]
- Ullmann, R.; Chien, C.D.; Avantaggiati, M.L.; Muller, S. An Acetylation Switch Regulates SUMO-Dependent Protein Interaction Networks. Mol. Cell 2012, 46, 759–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, R.; Sandelin, A. Determinants of Enhancer and Promoter Activities of Regulatory Elements. Nat. Rev. Genet. 2020, 21, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Benton, M.L.; Talipineni, S.C.; Kostka, D.; Capra, J.A. Genome-Wide Enhancer Annotations Differ Significantly in Genomic Distribution, Evolution, and Function. BMC Genom. 2019, 20, 511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasperini, M.; Tome, J.M.; Shendure, J. Towards a Comprehensive Catalogue of Validated and Target-Linked Human Enhancers. Nat. Rev. Genet. 2020, 21, 292–310. [Google Scholar] [CrossRef] [PubMed]
- Ho, E.Y.-K.; Cao, Q.; Gu, M.; Chan, R.W.-L.; Wu, Q.; Gerstein, M.; Yip, K.Y. Shaping the Nebulous Enhancer in the Era of High-Throughput Assays and Genome Editing. Brief. Bioinform. 2020, 21, 836–850. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, C.; Saiardi, A. Why Always Lysine? The Ongoing Tale of One of the Most Modified Amino Acids. Adv. Biol. Regul. 2016, 60, 144–150. [Google Scholar] [CrossRef]
- Sabari, B.R.; Zhang, D.; Allis, C.D.; Zhao, Y. Metabolic Regulation of Gene Expression through Histone Acylations. Nat. Rev. Mol. Cell Biol. 2017, 18, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Bossis, G.; Malnou, C.E.; Farras, R.; Andermarcher, E.; Hipskind, R.; Rodriguez, M.; Schmidt, D.; Muller, S.; Jariel-Encontre, I.; Piechaczyk, M. Down-Regulation of c-Fos/c-Jun AP-1 Dimer Activity by Sumoylation. Mol. Cell. Biol. 2005, 25, 6964–6979. [Google Scholar] [CrossRef] [Green Version]
- Muller, S.; Berger, M.; Lehembre, F.; Seeler, J.S.; Haupt, Y.; Dejean, A. C-Jun and P53 Activity Is Modulated by SUMO-1 Modification. J. Biol. Chem. 2000, 275, 13321–13329. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.-H.; Xiao, Z.; Kwon, S.; Sun, X.; Ryerson, D.; Tkac, D.; Ma, P.; Wu, S.-Y.; Chiang, C.-M.; Zhou, E.; et al. A Dysregulated Acetyl/SUMO Switch of FXR Promotes Hepatic Inflammation in Obesity. EMBO J. 2015, 34, 184–199. [Google Scholar] [CrossRef]
- Cui, W.; Sun, M.; Zhang, S.; Shen, X.; Galeva, N.; Williams, T.D.; Staudinger, J.L. A SUMO-Acetyl Switch in PXR Biology. Biochim. Biophys. Acta 2016, 1859, 1170–1182. [Google Scholar] [CrossRef] [Green Version]
- Hietakangas, V.; Anckar, J.; Blomster, H.A.; Fujimoto, M.; Palvimo, J.J.; Nakai, A.; Sistonen, L. PDSM, a Motif for Phosphorylation-Dependent SUMO Modification. Proc. Natl. Acad. Sci. USA 2006, 103, 45–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Vogel, W.K.; Liu, X.; Topark-Ngarm, A.; Arbogast, B.L.; Maier, C.S.; Filtz, T.M.; Leid, M. Coordinated Regulation of Transcription Factor Bcl11b Activity in Thymocytes by the Mitogen-Activated Protein Kinase (MAPK) Pathways and Protein Sumoylation. J. Biol. Chem. 2012, 287, 26971–26988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, H.; Liu, B.; Zhang, F.-J.; Zhang, J.; Dong, R.; Chen, L.; Qu, D.-M.; Lu, Y.; Yu, B.-W. The E3 Ligase APC/C-Cdh1 Regulates MEF2A-Dependent Transcription by Targeting SUMO-Specific Protease 2 for Ubiquitination and Degradation. Cell Cycle Georget. Tex. 2014, 13, 3892–3902. [Google Scholar] [CrossRef] [Green Version]
- Pilla, E.; Möller, U.; Sauer, G.; Mattiroli, F.; Melchior, F.; Geiss-Friedlander, R. A Novel SUMO1-Specific Interacting Motif in Dipeptidyl Peptidase 9 (DPP9) That Is Important for Enzymatic Regulation. J. Biol. Chem. 2012, 287, 44320–44329. [Google Scholar] [CrossRef] [Green Version]
- Diehl, C.; Akke, M.; Bekker-Jensen, S.; Mailand, N.; Streicher, W.; Wikström, M. Structural Analysis of a Complex between Small Ubiquitin-like Modifier 1 (SUMO1) and the ZZ Domain of CREB-Binding Protein (CBP/P300) Reveals a New Interaction Surface on SUMO. J. Biol. Chem. 2016, 291, 12658–12672. [Google Scholar] [CrossRef] [Green Version]
- Pichler, A.; Fatouros, C.; Lee, H.; Eisenhardt, N. SUMO Conjugation—A Mechanistic View. Biomol. Concepts 2017, 8, 13–36. [Google Scholar] [CrossRef] [PubMed]
- Liebelt, F.; Jansen, N.S.; Kumar, S.; Gracheva, E.; Claessens, L.A.; Verlaan-de Vries, M.; Willemstein, E.; Vertegaal, A.C.O. The Poly-SUMO2/3 Protease SENP6 Enables Assembly of the Constitutive Centromere-Associated Network by Group DeSUMOylation. Nat. Commun. 2019, 10, 3987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lallemand-Breitenbach, V.; Jeanne, M.; Benhenda, S.; Nasr, R.; Lei, M.; Peres, L.; Zhou, J.; Zhu, J.; Raught, B.; de Thé, H. Arsenic Degrades PML or PML-RARalpha through a SUMO-Triggered RNF4/Ubiquitin-Mediated Pathway. Nat. Cell Biol. 2008, 10, 547–555. [Google Scholar] [CrossRef]
- Tatham, M.H.; Geoffroy, M.-C.; Shen, L.; Plechanovova, A.; Hattersley, N.; Jaffray, E.G.; Palvimo, J.J.; Hay, R.T. RNF4 Is a Poly-SUMO-Specific E3 Ubiquitin Ligase Required for Arsenic-Induced PML Degradation. Nat. Cell Biol. 2008, 10, 538–546. [Google Scholar] [CrossRef]
- Kumar, R.; González-Prieto, R.; Xiao, Z.; Verlaan-de Vries, M.; Vertegaal, A.C.O. The STUbL RNF4 Regulates Protein Group SUMOylation by Targeting the SUMO Conjugation Machinery. Nat. Commun. 2017, 8, 1809. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Sabapathy, K. RNF4-A Paradigm for SUMOylation-Mediated Ubiquitination. Proteomics 2019, 19, e1900185. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Sang, J.; Ren, Y.; Liu, K.; Liu, X.; Zhang, J.; Wang, H.; Wang, J.; Orian, A.; Yang, J.; et al. SENP3 Regulates the Global Protein Turnover and the Sp1 Level via Antagonizing SUMO2/3-Targeted Ubiquitination and Degradation. Protein Cell 2016, 7, 63–77. [Google Scholar] [CrossRef] [Green Version]
- González-Prieto, R.; Cuijpers, S.A.; Kumar, R.; Hendriks, I.A.; Vertegaal, A.C. C-Myc Is Targeted to the Proteasome for Degradation in a SUMOylation-Dependent Manner, Regulated by PIAS1, SENP7 and RNF4. Cell Cycle Georget. Tex. 2015, 14, 1859–1872. [Google Scholar] [CrossRef] [Green Version]
- Rabellino, A.; Melegari, M.; Tompkins, V.S.; Chen, W.; Van Ness, B.G.; Teruya-Feldstein, J.; Conacci-Sorrell, M.; Janz, S.; Scaglioni, P.P. PIAS1 Promotes Lymphomagenesis through MYC Upregulation. Cell Rep. 2016, 15, 2266–2278. [Google Scholar] [CrossRef] [Green Version]
- Sriramachandran, A.M.; Meyer-Teschendorf, K.; Pabst, S.; Ulrich, H.D.; Gehring, N.H.; Hofmann, K.; Praefcke, G.J.K.; Dohmen, R.J. Arkadia/RNF111 Is a SUMO-Targeted Ubiquitin Ligase with Preference for Substrates Marked with SUMO1-Capped SUMO2/3 Chain. Nat. Commun. 2019, 10, 3678. [Google Scholar] [CrossRef]
- Sriramachandran, A.M.; Dohmen, R.J. SUMO-Targeted Ubiquitin Ligases. Biochim. Biophys. Acta 2014, 1843, 75–85. [Google Scholar] [CrossRef] [Green Version]
- Tsubakihara, Y.; Hikita, A.; Yamamoto, S.; Matsushita, S.; Matsushita, N.; Oshima, Y.; Miyazawa, K.; Imamura, T. Arkadia Enhances BMP Signalling through Ubiquitylation and Degradation of Smad6. J. Biochem. 2015, 158, 61–71. [Google Scholar] [CrossRef]
- Koinuma, D.; Shinozaki, M.; Komuro, A.; Goto, K.; Saitoh, M.; Hanyu, A.; Ebina, M.; Nukiwa, T.; Miyazawa, K.; Imamura, T.; et al. Arkadia Amplifies TGF-Beta Superfamily Signalling through Degradation of Smad7. EMBO J. 2003, 22, 6458–6470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McIntosh, D.J.; Walters, T.S.; Arinze, I.J.; Davis, J. Arkadia (RING Finger Protein 111) Mediates Sumoylation-Dependent Stabilization of Nrf2 Through K48-Linked Ubiquitination. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 46, 418–430. [Google Scholar] [CrossRef] [PubMed]
- Erker, Y.; Neyret-Kahn, H.; Seeler, J.S.; Dejean, A.; Atfi, A.; Levy, L. Arkadia, a Novel SUMO-Targeted Ubiquitin Ligase Involved in PML Degradation. Mol. Cell. Biol. 2013, 33, 2163–2177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, K.-S.; Kao, H.-Y. PML: Regulation and Multifaceted Function beyond Tumor Suppression. Cell Biosci. 2018, 8, 5. [Google Scholar] [CrossRef] [Green Version]
- González-Prieto, R.; Cuijpers, S.A.G.; Luijsterburg, M.S.; van Attikum, H.; Vertegaal, A.C.O. SUMOylation and PARylation Cooperate to Recruit and Stabilize SLX4 at DNA Damage Sites. EMBO Rep. 2015, 16, 512–519. [Google Scholar] [CrossRef]
- Jentsch, S.; Psakhye, I. Control of Nuclear Activities by Substrate-Selective and Protein-Group SUMOylation. Annu. Rev. Genet. 2013, 47, 167–186. [Google Scholar] [CrossRef] [PubMed]
- Psakhye, I.; Jentsch, S. Protein Group Modification and Synergy in the SUMO Pathway as Exemplified in DNA Repair. Cell 2012, 151, 807–820. [Google Scholar] [CrossRef] [Green Version]
- Sahin, U.; Ferhi, O.; Jeanne, M.; Benhenda, S.; Berthier, C.; Jollivet, F.; Niwa-Kawakita, M.; Faklaris, O.; Setterblad, N.; de Thé, H.; et al. Oxidative Stress-Induced Assembly of PML Nuclear Bodies Controls Sumoylation of Partner Proteins. J. Cell Biol. 2014, 204, 931–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cossec, J.-C.; Theurillat, I.; Chica, C.; Búa Aguín, S.; Gaume, X.; Andrieux, A.; Iturbide, A.; Jouvion, G.; Li, H.; Bossis, G.; et al. SUMO Safeguards Somatic and Pluripotent Cell Identities by Enforcing Distinct Chromatin States. Cell Stem Cell 2018, 23, 742–757.e8. [Google Scholar] [CrossRef] [Green Version]
- Hyman, A.A.; Weber, C.A.; Jülicher, F. Liquid-Liquid Phase Separation in Biology. Annu. Rev. Cell Dev. Biol. 2014, 30, 39–58. [Google Scholar] [CrossRef] [Green Version]
- Hnisz, D.; Shrinivas, K.; Young, R.A.; Chakraborty, A.K.; Sharp, P.A. A Phase Separation Model for Transcriptional Control. Cell 2017, 169, 13–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, L.; Li, E.-M.; Xu, L.-Y. From Start to End: Phase Separation and Transcriptional Regulation. Biochim. Biophys. Acta Gene Regul. Mech. 2020, 1863, 194641. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.R.; Ostwal, Y.B. Post-Translational Modification, Phase Separation, and Robust Gene Transcription. Trends Genet. TIG 2019, 35, 89–92. [Google Scholar] [CrossRef]
- Banani, S.F.; Rice, A.M.; Peeples, W.B.; Lin, Y.; Jain, S.; Parker, R.; Rosen, M.K. Compositional Control of Phase-Separated Cellular Bodies. Cell 2016, 166, 651–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ford, L.; Ling, E.; Kandel, E.R.; Fioriti, L. CPEB3 Inhibits Translation of MRNA Targets by Localizing Them to P Bodies. Proc. Natl. Acad. Sci. USA 2019, 116, 18078–18087. [Google Scholar] [CrossRef] [Green Version]
- Min, J.; Wright, W.E.; Shay, J.W. Clustered Telomeres in Phase-Separated Nuclear Condensates Engage Mitotic DNA Synthesis through BLM and RAD52. Genes Dev. 2019, 33, 814–827. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhao, R.; Tones, J.; Liu, M.; Dilley, R.L.; Chenoweth, D.M.; Greenberg, R.A.; Lampson, M.A. Nuclear Body Phase Separation Drives Telomere Clustering in ALT Cancer Cells. Mol. Biol. Cell 2020, 31, 2048–2056. [Google Scholar] [CrossRef] [PubMed]
- Cubeñas-Potts, C.; Matunis, M.J. SUMO: A Multifaceted Modifier of Chromatin Structure and Function. Dev. Cell 2013, 24, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.-H.; Sharrocks, A.D. SUMO Promotes HDAC-Mediated Transcriptional Repression. Mol. Cell 2004, 13, 611–617. [Google Scholar] [CrossRef]
- Holmstrom, S.; Van Antwerp, M.E.; Iñiguez-Lluhi, J.A. Direct and Distinguishable Inhibitory Roles for SUMO Isoforms in the Control of Transcriptional Synergy. Proc. Natl. Acad. Sci. USA 2003, 100, 15758–15763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chymkowitch, P.; Nguéa, A.P.; Aanes, H.; Koehler, C.J.; Thiede, B.; Lorenz, S.; Meza-Zepeda, L.A.; Klungland, A.; Enserink, J.M. Sumoylation of Rap1 Mediates the Recruitment of TFIID to Promote Transcription of Ribosomal Protein Genes. Genome Res. 2015, 25, 897–906. [Google Scholar] [CrossRef] [Green Version]
- Gomez-del Arco, P.; Koipally, J.; Georgopoulos, K. Ikaros SUMOylation: Switching Out of Repression. Mol. Cell. Biol. 2005, 25, 2688–2697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledsaak, M.; Bengtsen, M.; Molværsmyr, A.-K.; Fuglerud, B.M.; Matre, V.; Eskeland, R.; Gabrielsen, O.S. PIAS1 Binds P300 and Behaves as a Coactivator or Corepressor of the Transcription Factor C-Myb Dependent on SUMO-Status. Biochim. Biophys. Acta BBA Gene Regul. Mech. 2016, 1859, 705–718. [Google Scholar] [CrossRef]
- Rosonina, E.; Duncan, S.M.; Manley, J.L. SUMO Functions in Constitutive Transcription and during Activation of Inducible Genes in Yeast. Genes Dev. 2010, 24, 1242–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.; Chun, S.K.; Kim, K. Sumoylation Controls CLOCK-BMAL1-Mediated Clock Resetting via CBP Recruitment in Nuclear Transcriptional Foci. Biochim. Biophys. Acta 2015, 1853, 2697–2708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Q.; Gong, L.; Deng, M.; Zhang, L.; Sun, S.; Liu, J.; Ma, H.; Yuan, D.; Chen, P.-C.; Hu, X.; et al. Sumoylation Activates the Transcriptional Activity of Pax-6, an Important Transcription Factor for Eye and Brain Development. Proc. Natl. Acad. Sci. USA 2010, 107, 21034–21039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shyu, Y.-C.; Lee, T.-L.; Ting, C.-Y.; Wen, S.-C.; Hsieh, L.-J.; Li, Y.-C.; Hwang, J.-L.; Lin, C.-C.; Shen, C.-K.J. Sumoylation of P45/NF-E2: Nuclear Positioning and Transcriptional Activation of the Mammalian Beta-like Globin Gene Locus. Mol. Cell. Biol. 2005, 25, 10365–10378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.-Y.; Johnson, K.D.; Fujiwara, T.; Boyer, M.E.; Kim, S.-I.; Bresnick, E.H. Controlling Hematopoiesis through Sumoylation-Dependent Regulation of a GATA Factor. Mol. Cell 2009, 36, 984–995. [Google Scholar] [CrossRef] [Green Version]
- Rytinki, M.; Kaikkonen, S.; Sutinen, P.; Paakinaho, V.; Rahkama, V.; Palvimo, J.J. Dynamic SUMOylation Is Linked to the Activity Cycles of Androgen Receptor in the Cell Nucleus. Mol. Cell. Biol. 2012, 32, 4195–4205. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Feng, X.-H.; Schwartz, R.J. SUMO-1 Modification Activated GATA4-Dependent Cardiogenic Gene Activity. J. Biol. Chem. 2004, 279, 49091–49098. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Liang, M.; Liang, Y.-Y.; Brunicardi, F.C.; Melchior, F.; Feng, X.-H. Activation of Transforming Growth Factor-Beta Signaling by SUMO-1 Modification of Tumor Suppressor Smad4/DPC4. J. Biol. Chem. 2003, 278, 18714–18719. [Google Scholar] [CrossRef] [Green Version]
- Terui, Y.; Saad, N.; Jia, S.; McKeon, F.; Yuan, J. Dual Role of Sumoylation in the Nuclear Localization and Transcriptional Activation of NFAT1. J. Biol. Chem. 2004, 279, 28257–28265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, B.; Sharrocks, A.D. Extracellular Signal-Regulated Kinase Mitogen-Activated Protein Kinase Signaling Initiates a Dynamic Interplay between Sumoylation and Ubiquitination to Regulate the Activity of the Transcriptional Activator PEA3. Mol. Cell. Biol. 2009, 29, 3204–3218. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.; Rogers, R.; Matunis, M.J.; Mayhew, C.N.; Goodson, M.L.; Park-Sarge, O.K.; Sarge, K.D.; Goodson, M. Regulation of Heat Shock Transcription Factor 1 by Stress-Induced SUMO-1 Modification. J. Biol. Chem. 2001, 276, 40263–40267. [Google Scholar] [CrossRef] [Green Version]
- Goodson, M.L.; Hong, Y.; Rogers, R.; Matunis, M.J.; Park-Sarge, O.K.; Sarge, K.D. Sumo-1 Modification Regulates the DNA Binding Activity of Heat Shock Transcription Factor 2, a Promyelocytic Leukemia Nuclear Body Associated Transcription Factor. J. Biol. Chem. 2001, 276, 18513–18518. [Google Scholar] [CrossRef] [Green Version]
- Hilgarth, R.S.; Murphy, L.A.; O’Connor, C.M.; Clark, J.A.; Park-Sarge, O.-K.; Sarge, K.D. Identification of Xenopus Heat Shock Transcription Factor-2: Conserved Role of Sumoylation in Regulating Deoxyribonucleic Acid-Binding Activity of Heat Shock Transcription Factor-2 Proteins. Cell Stress Chaperones 2004, 9, 214–220. [Google Scholar] [CrossRef] [Green Version]
- Garaude, J.; Farrás, R.; Bossis, G.; Charni, S.; Piechaczyk, M.; Hipskind, R.A.; Villalba, M. SUMOylation Regulates the Transcriptional Activity of JunB in T Lymphocytes. J. Immunol. 2008, 180, 5983–5990. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, M.S.; Desterro, J.M.; Lain, S.; Midgley, C.A.; Lane, D.P.; Hay, R.T. SUMO-1 Modification Activates the Transcriptional Response of P53. EMBO J. 1999, 18, 6455–6461. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, H.; Ihara, M.; Matsuura, Y.; Kikuchi, A. Sumoylation Is Involved in Beta-Catenin-Dependent Activation of Tcf-4. EMBO J. 2003, 22, 2047–2059. [Google Scholar] [CrossRef] [Green Version]
- Gostissa, M.; Hengstermann, A.; Fogal, V.; Sandy, P.; Schwarz, S.E.; Scheffner, M.; Del Sal, G. Activation of P53 by Conjugation to the Ubiquitin-like Protein SUMO-1. EMBO J. 1999, 18, 6462–6471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.-Y.; Chiang, C.-M. Crosstalk between Sumoylation and Acetylation Regulates P53-Dependent Chromatin Transcription and DNA Binding. EMBO J. 2009, 28, 1246–1259. [Google Scholar] [CrossRef] [Green Version]
- Brunet Simioni, M.; De Thonel, A.; Hammann, A.; Joly, A.L.; Bossis, G.; Fourmaux, E.; Bouchot, A.; Landry, J.; Piechaczyk, M.; Garrido, C. Heat Shock Protein 27 Is Involved in SUMO-2/3 Modification of Heat Shock Factor 1 and Thereby Modulates the Transcription Factor Activity. Oncogene 2009, 28, 3332–3344. [Google Scholar] [CrossRef] [Green Version]
- Hietakangas, V.; Ahlskog, J.K.; Jakobsson, A.M.; Hellesuo, M.; Sahlberg, N.M.; Holmberg, C.I.; Mikhailov, A.; Palvimo, J.J.; Pirkkala, L.; Sistonen, L. Phosphorylation of Serine 303 Is a Prerequisite for the Stress-Inducible SUMO Modification of Heat Shock Factor 1. Mol. Cell. Biol. 2003, 23, 2953–2968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhter, A.; Rosonina, E. Chromatin Association of Gcn4 Is Limited by Post-Translational Modifications Triggered by Its DNA-Binding in Saccharomyces Cerevisiae. Genetics 2016, 204, 1433–1445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosonina, E.; Duncan, S.M.; Manley, J.L. Sumoylation of Transcription Factor Gcn4 Facilitates Its Srb10-Mediated Clearance from Promoters in Yeast. Genes Dev. 2012, 26, 350–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tempé, D.; Vives, E.; Brockly, F.; Brooks, H.; De Rossi, S.; Piechaczyk, M.; Bossis, G. SUMOylation of the Inducible (c-Fos:C-Jun)/AP-1 Transcription Complex Occurs on Target Promoters to Limit Transcriptional Activation. Oncogene 2014, 33, 921–927. [Google Scholar] [CrossRef] [Green Version]
- Neyret-Kahn, H.; Benhamed, M.; Ye, T.; Le Gras, S.; Cossec, J.-C.; Lapaquette, P.; Bischof, O.; Ouspenskaia, M.; Dasso, M.; Seeler, J.; et al. Sumoylation at Chromatin Governs Coordinated Repression of a Transcriptional Program Essential for Cell Growth and Proliferation. Genome Res. 2013, 23, 1563–1579. [Google Scholar] [CrossRef] [Green Version]
- Niskanen, E.A.; Palvimo, J.J. Chromatin SUMOylation in Heat Stress: To Protect, Pause and Organise?: SUMO Stress Response on Chromatin. BioEssays News Rev. Mol. Cell. Dev. Biol. 2017, 39, 1600263. [Google Scholar] [CrossRef]
- Niskanen, E.A.; Malinen, M.; Sutinen, P.; Toropainen, S.; Paakinaho, V.; Vihervaara, A.; Joutsen, J.; Kaikkonen, M.U.; Sistonen, L.; Palvimo, J.J. Global SUMOylation on Active Chromatin Is an Acute Heat Stress Response Restricting Transcription. Genome Biol. 2015, 16, 153. [Google Scholar] [CrossRef] [Green Version]
- Salinas, S.; Briançon-Marjollet, A.; Bossis, G.; Lopez, M.-A.; Piechaczyk, M.; Jariel-Encontre, I.; Debant, A.; Hipskind, R.A. SUMOylation Regulates Nucleo-Cytoplasmic Shuttling of Elk-1. J. Cell Biol. 2004, 165, 767–773. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.-H.; Sharrocks, A.D. PIASx Acts as an Elk-1 Coactivator by Facilitating Derepression. EMBO J. 2005, 24, 2161–2171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Wang, Z.; Wang, Z.; Yu, F.; Li, J.; Wong, J. SUMOylation Down-Regulates RDNA Transcription by Repressing Expression of Upstream-Binding Factor and Proto-Oncogene c-Myc. J. Biol. Chem. 2019, 294, 19155–19166. [Google Scholar] [CrossRef]
- Wohlschlegel, J.A.; Johnson, E.S.; Reed, S.I.; Yates, J.R. Global Analysis of Protein Sumoylation in Saccharomyces Cerevisiae. J. Biol. Chem. 2004, 279, 45662–45668. [Google Scholar] [CrossRef] [Green Version]
- Das, A.; Banday, M.; Bellofatto, V. RNA Polymerase Transcription Machinery in Trypanosomes. Eukaryot. Cell 2008, 7, 429–434. [Google Scholar] [CrossRef] [Green Version]
- López-Farfán, D.; Bart, J.-M.; Rojas-Barros, D.I.; Navarro, M. SUMOylation by the E3 Ligase TbSIZ1/PIAS1 Positively Regulates VSG Expression in Trypanosoma Brucei. PLoS Pathog. 2014, 10, e1004545. [Google Scholar] [CrossRef]
- Saura, A.; Iribarren, P.A.; Rojas-Barros, D.; Bart, J.M.; López-Farfán, D.; Andrés-León, E.; Vidal-Cobo, I.; Boehm, C.; Alvarez, V.E.; Field, M.C.; et al. SUMOylated SNF2PH Promotes Variant Surface Glycoprotein Expression in Bloodstream Trypanosomes. EMBO Rep. 2019, 20, e48029. [Google Scholar] [CrossRef]
- Chymkowitch, P.; Nguéa, P.A.; Aanes, H.; Robertson, J.; Klungland, A.; Enserink, J.M. TORC1-Dependent Sumoylation of Rpc82 Promotes RNA Polymerase III Assembly and Activity. Proc. Natl. Acad. Sci. USA 2017, 114, 1039–1044. [Google Scholar] [CrossRef] [Green Version]
- Nguéa, P.A.; Robertson, J.; Herrera, M.C.; Chymkowitch, P.; Enserink, J.M. Desumoylation of RNA Polymerase III Lies at the Core of the Sumo Stress Response in Yeast. J. Biol. Chem. 2019, 294, 18784–18795. [Google Scholar] [CrossRef]
- Wang, Z.; Wu, C.; Aslanian, A.; Yates, J.R.; Hunter, T. Defective RNA Polymerase III Is Negatively Regulated by the SUMO-Ubiquitin-Cdc48 Pathway. eLife 2018, 7, e35447. [Google Scholar] [CrossRef]
- Zhang, S.; Li, X.; Wang, H.-Y.; Steven Zheng, X.F. Beyond Regulation of Pol III: Role of MAF1 in Growth, Metabolism, Aging and Cancer. Biochim. Biophys. Acta Gene Regul. Mech. 2018, 1861, 338–343. [Google Scholar] [CrossRef]
- Rohira, A.D.; Chen, C.-Y.; Allen, J.R.; Johnson, D.L. Covalent Small Ubiquitin-like Modifier (SUMO) Modification of Maf1 Protein Controls RNA Polymerase III-Dependent Transcription Repression. J. Biol. Chem. 2013, 288, 19288–19295. [Google Scholar] [CrossRef] [Green Version]
- Chang, P.-C.; Cheng, C.-Y.; Campbell, M.; Yang, Y.-C.; Hsu, H.-W.; Chang, T.-Y.; Chu, C.-H.; Lee, Y.-W.; Hung, C.-L.; Lai, S.-M.; et al. The Chromatin Modification by SUMO-2/3 but Not SUMO-1 Prevents the Epigenetic Activation of Key Immune-Related Genes during Kaposi’s Sarcoma Associated Herpesvirus Reactivation. BMC Genom. 2013, 14, 824. [Google Scholar] [CrossRef] [Green Version]
- Decque, A.; Joffre, O.; Magalhaes, J.G.; Cossec, J.-C.; Blecher-Gonen, R.; Lapaquette, P.; Silvin, A.; Manel, N.; Joubert, P.-E.; Seeler, J.-S.; et al. Sumoylation Coordinates the Repression of Inflammatory and Anti-Viral Gene-Expression Programs during Innate Sensing. Nat. Immunol. 2016, 17, 140–149. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, J.; Heine, G.F.; Arora, M.; Gulcin Ozer, H.; Onti-Srinivasan, R.; Huang, K.; Parvin, J.D. Chromatin Modification by SUMO-1 Stimulates the Promoters of Translation Machinery Genes. Nucleic Acids Res. 2012, 40, 10172–10186. [Google Scholar] [CrossRef] [Green Version]
- Paakinaho, V.; Kaikkonen, S.; Makkonen, H.; Benes, V.; Palvimo, J.J. SUMOylation Regulates the Chromatin Occupancy and Anti-Proliferative Gene Programs of Glucocorticoid Receptor. Nucleic Acids Res. 2014, 42, 1575–1592. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Castañeda, F.; Lemma, R.B.; Cuervo, I.; Bengtsen, M.; Moen, L.M.; Ledsaak, M.; Eskeland, R.; Gabrielsen, O.S. The SUMO Protease SENP1 and the Chromatin Remodeler CHD3 Interact and Jointly Affect Chromatin Accessibility and Gene Expression. J. Biol. Chem. 2018, 293, 15439–15454. [Google Scholar] [CrossRef] [Green Version]
- Sri Theivakadadcham, V.S.; Bergey, B.G.; Rosonina, E. Sumoylation of DNA-Bound Transcription Factor Sko1 Prevents Its Association with Nontarget Promoters. PLoS Genet. 2019, 15, e1007991. [Google Scholar] [CrossRef] [Green Version]
- Toropainen, S.; Malinen, M.; Kaikkonen, S.; Rytinki, M.; Jääskeläinen, T.; Sahu, B.; Jänne, O.A.; Palvimo, J.J. SUMO Ligase PIAS1 Functions as a Target Gene Selective Androgen Receptor Coregulator on Prostate Cancer Cell Chromatin. Nucleic Acids Res. 2015, 43, 848–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.-S.; Hsu, H.-W.; Campbell, M.; Cheng, C.-Y.; Chang, P.-C. K-BZIP Mediated SUMO-2/3 Specific Modification on the KSHV Genome Negatively Regulates Lytic Gene Expression and Viral Reactivation. PLoS Pathog. 2015, 11, e1005051. [Google Scholar] [CrossRef] [PubMed]
- MacPherson, M.J.; Beatty, L.G.; Zhou, W.; Du, M.; Sadowski, P.D. The CTCF Insulator Protein Is Posttranslationally Modified by SUMO. Mol. Cell. Biol. 2009, 29, 714–725. [Google Scholar] [CrossRef] [Green Version]
- Zilio, N.; Eifler-Olivi, K.; Ulrich, H.D. Functions of SUMO in the Maintenance of Genome Stability. Adv. Exp. Med. Biol. 2017, 963, 51–87. [Google Scholar] [CrossRef] [PubMed]
- Ninova, M.; Fejes Tóth, K.; Aravin, A.A. The Control of Gene Expression and Cell Identity by H3K9 Trimethylation. Dev. Camb. Engl. 2019, 146, dev181180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yalçin, Z.; Selenz, C.; Jacobs, J.J.L. Ubiquitination and SUMOylation in Telomere Maintenance and Dysfunction. Front. Genet. 2017, 8, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuijpers, S.A.G.; Vertegaal, A.C.O. Guiding Mitotic Progression by Crosstalk between Post-Translational Modifications. Trends Biochem. Sci. 2018, 43, 251–268. [Google Scholar] [CrossRef]
- Nottke, A.C.; Kim, H.-M.; Colaiácovo, M.P. Wrestling with Chromosomes: The Roles of SUMO During Meiosis. Adv. Exp. Med. Biol. 2017, 963, 185–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lickwar, C.R.; Mueller, F.; Hanlon, S.E.; McNally, J.G.; Lieb, J.D. Genome-Wide Protein-DNA Binding Dynamics Suggest a Molecular Clutch for Transcription Factor Function. Nature 2012, 484, 251–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rytinki, M.M.; Kaikkonen, S.; Pehkonen, P.; Jääskeläinen, T.; Palvimo, J.J. PIAS Proteins: Pleiotropic Interactors Associated with SUMO. Cell. Mol. Life Sci. CMLS 2009, 66, 3029–3041. [Google Scholar] [CrossRef]
- Bertolotto, C.; Lesueur, F.; Giuliano, S.; Strub, T.; de Lichy, M.; Bille, K.; Dessen, P.; d’Hayer, B.; Mohamdi, H.; Remenieras, A.; et al. A SUMOylation-Defective MITF Germline Mutation Predisposes to Melanoma and Renal Carcinoma. Nature 2011, 480, 94–98. [Google Scholar] [CrossRef]
- Sutinen, P.; Rahkama, V.; Rytinki, M.; Palvimo, J.J. Nuclear Mobility and Activity of FOXA1 with Androgen Receptor Are Regulated by SUMOylation. Mol. Endocrinol. Baltim. Md 2014, 28, 1719–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Yuan, C.; Wu, J.; Elsayed, Z.; Fu, Z. Polo-like Kinase 1-Mediated Phosphorylation of Forkhead Box Protein M1b Antagonizes Its SUMOylation and Facilitates Its Mitotic Function. J. Biol. Chem. 2015, 290, 3708–3719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Ma, Y.; Qian, L.; Wang, J. Sumoylation Regulates Nuclear Localization and Function of Zinc Finger Transcription Factor ZIC3. Biochim. Biophys. Acta 2013, 1833, 2725–2733. [Google Scholar] [CrossRef] [Green Version]
- Sahin, U.; de Thé, H.; Lallemand-Breitenbach, V. PML Nuclear Bodies: Assembly and Oxidative Stress-Sensitive Sumoylation. Nucl. Austin Tex. 2014, 5, 499–507. [Google Scholar] [CrossRef] [Green Version]
- Nayak, A.; Glöckner-Pagel, J.; Vaeth, M.; Schumann, J.E.; Buttmann, M.; Bopp, T.; Schmitt, E.; Serfling, E.; Berberich-Siebelt, F. Sumoylation of the Transcription Factor NFATc1 Leads to Its Subnuclear Relocalization and Interleukin-2 Repression by Histone Deacetylase. J. Biol. Chem. 2009, 284, 10935–10946. [Google Scholar] [CrossRef] [Green Version]
- Gong, L.; Ji, W.-K.; Hu, X.-H.; Hu, W.-F.; Tang, X.-C.; Huang, Z.-X.; Li, L.; Liu, M.; Xiang, S.-H.; Wu, E.; et al. Sumoylation Differentially Regulates Sp1 to Control Cell Differentiation. Proc. Natl. Acad. Sci. USA 2014, 111, 5574–5579. [Google Scholar] [CrossRef] [Green Version]
- Pan, M.-R.; Chang, T.-M.; Chang, H.-C.; Su, J.-L.; Wang, H.-W.; Hung, W.-C. Sumoylation of Prox1 Controls Its Ability to Induce VEGFR3 Expression and Lymphatic Phenotypes in Endothelial Cells. J. Cell Sci. 2009, 122, 3358–3364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myatt, S.S.; Kongsema, M.; Man, C.W.-Y.; Kelly, D.J.; Gomes, A.R.; Khongkow, P.; Karunarathna, U.; Zona, S.; Langer, J.K.; Dunsby, C.W.; et al. SUMOylation Inhibits FOXM1 Activity and Delays Mitotic Transition. Oncogene 2014, 33, 4316–4329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamard, P.-J.; Boyer-Guittaut, M.; Camuzeaux, B.; Dujardin, D.; Hauss, C.; Oelgeschläger, T.; Vigneron, M.; Kedinger, C.; Chatton, B. Sumoylation Delays the ATF7 Transcription Factor Subcellular Localization and Inhibits Its Transcriptional Activity. Nucleic Acids Res. 2007, 35, 1134–1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, C.H.; Akhter, A.; Yurko, N.; Burgener, J.M.; Rosonina, E.; Manley, J.L. Sumoylation Controls the Timing of Tup1-Mediated Transcriptional Deactivation. Nat. Commun. 2015, 6, 6610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voss, A.K.; Thomas, T. Histone Lysine and Genomic Targets of Histone Acetyltransferases in Mammals. BioEssays News Rev. Mol. Cell. Dev. Biol. 2018, 40, e1800078. [Google Scholar] [CrossRef]
- Gill, G. Something about SUMO Inhibits Transcription. Curr. Opin. Genet. Dev. 2005, 15, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, J.; Shi, Y.; Valin, A.; Xuan, Y.; Gill, G. Direct Binding of CoREST1 to SUMO-2/3 Contributes to Gene-Specific Repression by the LSD1/CoREST1/HDAC Complex. Mol. Cell 2009, 34, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Andrade, D.; Velinder, M.; Singer, J.; Maese, L.; Bareyan, D.; Nguyen, H.; Chandrasekharan, M.B.; Lucente, H.; McClellan, D.; Jones, D.; et al. SUMOylation Regulates Growth Factor Independence 1 in Transcriptional Control and Hematopoiesis. Mol. Cell. Biol. 2016, 36, 1438–1450. [Google Scholar] [CrossRef] [Green Version]
- Hua, G.; Ganti, K.P.; Chambon, P. Glucocorticoid-Induced Tethered Transrepression Requires SUMOylation of GR and Formation of a SUMO-SMRT/NCoR1-HDAC3 Repressing Complex. Proc. Natl. Acad. Sci. USA 2016, 113, E635–E643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, G.; Paulen, L.; Chambon, P. GR SUMOylation and Formation of an SUMO-SMRT/NCoR1-HDAC3 Repressing Complex Is Mandatory for GC-Induced IR NGRE-Mediated Transrepression. Proc. Natl. Acad. Sci. USA 2016, 113, E626–E634. [Google Scholar] [CrossRef] [Green Version]
- Ahn, J.-W.; Lee, Y.-A.; Ahn, J.-H.; Choi, C.Y. Covalent Conjugation of Groucho with SUMO-1 Modulates Its Corepressor Activity. Biochem. Biophys. Res. Commun. 2009, 379, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Ao, X.; Li, S.; Xu, Z.; Yang, Y.; Chen, M.; Jiang, X.; Wu, H. Sumoylation of TCF21 Downregulates the Transcriptional Activity of Estrogen Receptor-Alpha. Oncotarget 2016, 7, 26220–26234. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, A.-M.F.; Nicol, S.M.; Hislop, R.G.; Jaffray, E.G.; Hay, R.T.; Fuller-Pace, F.V. SUMO Modification of the DEAD Box Protein P68 Modulates Its Transcriptional Activity and Promotes Its Interaction with HDAC1. Oncogene 2007, 26, 5866–5876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Choi, H.J.; Kim, B.; Kim, M.H.; Lee, J.M.; Kim, I.S.; Lee, M.H.; Choi, S.J.; Kim, K.I.; Kim, S.-I.; et al. Roles of Sumoylation of a Reptin Chromatin-Remodelling Complex in Cancer Metastasis. Nat. Cell Biol. 2006, 8, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Pascual, G.; Fong, A.L.; Ogawa, S.; Gamliel, A.; Li, A.C.; Perissi, V.; Rose, D.W.; Willson, T.M.; Rosenfeld, M.G.; Glass, C.K. A SUMOylation-Dependent Pathway Mediates Transrepression of Inflammatory Response Genes by PPAR-Gamma. Nature 2005, 437, 759–763. [Google Scholar] [CrossRef] [PubMed]
- Pourcet, B.; Pineda-Torra, I.; Derudas, B.; Staels, B.; Glineur, C. SUMOylation of Human Peroxisome Proliferator-Activated Receptor Alpha Inhibits Its Trans-Activity through the Recruitment of the Nuclear Corepressor NCoR. J. Biol. Chem. 2010, 285, 5983–5992. [Google Scholar] [CrossRef] [Green Version]
- Ceballos-Chávez, M.; Rivero, S.; García-Gutiérrez, P.; Rodríguez-Paredes, M.; García-Domínguez, M.; Bhattacharya, S.; Reyes, J.C. Control of Neuronal Differentiation by Sumoylation of BRAF35, a Subunit of the LSD1-CoREST Histone Demethylase Complex. Proc. Natl. Acad. Sci. USA 2012, 109, 8085–8090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, T.; Kiweler, N.; Wolff, K.; Knauer, S.K.; Brandl, A.; Hemmerich, P.; Dannenberg, J.-H.; Heinzel, T.; Schneider, G.; Krämer, O.H. Sumoylation of HDAC2 Promotes NF-ΚB-Dependent Gene Expression. Oncotarget 2015, 6, 7123–7135. [Google Scholar] [CrossRef] [Green Version]
- Brandl, A.; Wagner, T.; Uhlig, K.M.; Knauer, S.K.; Stauber, R.H.; Melchior, F.; Schneider, G.; Heinzel, T.; Krämer, O.H. Dynamically Regulated Sumoylation of HDAC2 Controls P53 Deacetylation and Restricts Apoptosis Following Genotoxic Stress. J. Mol. Cell Biol. 2012, 4, 284–293. [Google Scholar] [CrossRef] [Green Version]
- Citro, S.; Jaffray, E.; Hay, R.T.; Seiser, C.; Chiocca, S. A Role for Paralog-Specific Sumoylation in Histone Deacetylase 1 Stability. J. Mol. Cell Biol. 2013, 5, 416–427. [Google Scholar] [CrossRef]
- Joung, H.; Kwon, S.; Kim, K.-H.; Lee, Y.-G.; Shin, S.; Kwon, D.-H.; Lee, Y.-U.; Kook, T.; Choe, N.; Kim, J.C.; et al. Sumoylation of Histone Deacetylase 1 Regulates MyoD Signaling during Myogenesis. Exp. Mol. Med. 2018, 50, e427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyun, K.; Jeon, J.; Park, K.; Kim, J. Writing, Erasing and Reading Histone Lysine Methylations. Exp. Mol. Med. 2017, 49, e324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, D. Lysine Methylation Signaling of Non-Histone Proteins in the Nucleus. Cell. Mol. Life Sci. CMLS 2019, 76, 2873–2883. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shankar, S.R.; Kher, D.; Ling, B.M.T.; Taneja, R. Sumoylation of the Basic Helix-Loop-Helix Transcription Factor Sharp-1 Regulates Recruitment of the Histone Methyltransferase G9a and Function in Myogenesis. J. Biol. Chem. 2013, 288, 17654–17662. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, S.; Shankar, S.R.; Wang, Y.; Taneja, R. SUMOylation of G9a Regulates Its Function as an Activator of Myoblast Proliferation. Cell Death Dis. 2019, 10, 250. [Google Scholar] [CrossRef]
- Zahnow, C.A.; Topper, M.; Stone, M.; Murray-Stewart, T.; Li, H.; Baylin, S.B.; Casero, R.A. Inhibitors of DNA Methylation, Histone Deacetylation, and Histone Demethylation: A Perfect Combination for Cancer Therapy. Adv. Cancer Res. 2016, 130, 55–111. [Google Scholar] [CrossRef] [PubMed]
- Mottis, A.; Mouchiroud, L.; Auwerx, J. Emerging Roles of the Corepressors NCoR1 and SMRT in Homeostasis. Genes Dev. 2013, 27, 819–835. [Google Scholar] [CrossRef] [Green Version]
- Tiefenbach, J.; Novac, N.; Ducasse, M.; Eck, M.; Melchior, F.; Heinzel, T. SUMOylation of the Corepressor N-CoR Modulates Its Capacity to Repress Transcription. Mol. Biol. Cell 2006, 17, 1643–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muraoka, A.; Maeda, A.; Nakahara, N.; Yokota, M.; Nishida, T.; Maruyama, T.; Ohshima, T. Sumoylation of CoREST Modulates Its Function as a Transcriptional Repressor. Biochem. Biophys. Res. Commun. 2008, 377, 1031–1035. [Google Scholar] [CrossRef]
- Sáez, J.E.; Arredondo, C.; Rivera, C.; Andrés, M.E. PIASγ Controls Stability and Facilitates SUMO-2 Conjugation to CoREST Family of Transcriptional Co-Repressors. Biochem. J. 2018, 475, 1441–1454. [Google Scholar] [CrossRef]
- He, Z.; Zhang, J.; Huang, Z.; Du, Q.; Li, N.; Zhang, Q.; Chen, Y.; Sun, Z. Sumoylation of RORγt Regulates TH17 Differentiation and Thymocyte Development. Nat. Commun. 2018, 9, 4870. [Google Scholar] [CrossRef] [Green Version]
- Boyer-Guittaut, M.; Birsoy, K.; Potel, C.; Elliott, G.; Jaffray, E.; Desterro, J.M.; Hay, R.T.; Oelgeschläger, T. SUMO-1 Modification of Human Transcription Factor (TF) IID Complex Subunits: Inhibition of TFIID Promoter-Binding Activity through SUMO-1 Modification of HsTAF5. J. Biol. Chem. 2005, 280, 9937–9945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosas-Acosta, G.; Russell, W.K.; Deyrieux, A.; Russell, D.H.; Wilson, V.G. A Universal Strategy for Proteomic Studies of SUMO and Other Ubiquitin-like Modifiers. Mol. Cell. Proteom. MCP 2005, 4, 56–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Chen, Z.; Shim, M.S.; Lee, M.S.; Kim, J.E.; Kwon, Y.E.; Yoo, T.J.; Kim, J.Y.; Bang, J.Y.; Carlson, B.A.; et al. SUMO Modification of NZFP Mediates Transcriptional Repression through TBP Binding. Mol. Cells 2013, 35, 70–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, F.; Shi, G.; Cheng, S.; Chen, J.; Wu, S.-Y.; Wang, Z.; Xia, N.; Zhai, Y.; Wang, Z.; Peng, Y.; et al. SUMO Suppresses and MYC Amplifies Transcription Globally by Regulating CDK9 Sumoylation. Cell Res. 2018, 28, 670–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Yang, T.; Luo, Y.; Wu, L.; Jiang, Y.; Song, Z.; Pan, T.; Liu, B.; Liu, G.; Liu, J.; et al. TRIM28 Promotes HIV-1 Latency by SUMOylating CDK9 and Inhibiting P-TEFb. eLife 2019, 8, e42426. [Google Scholar] [CrossRef]
- Cavalli, G.; Heard, E. Advances in Epigenetics Link Genetics to the Environment and Disease. Nature 2019, 571, 489–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jerković, I.; Szabo, Q.; Bantignies, F.; Cavalli, G. Higher-Order Chromosomal Structures Mediate Genome Function. J. Mol. Biol. 2020, 432, 676–681. [Google Scholar] [CrossRef]
- Nacerddine, K.; Lehembre, F.; Bhaumik, M.; Artus, J.; Cohen-Tannoudji, M.; Babinet, C.; Pandolfi, P.P.; Dejean, A. The SUMO Pathway Is Essential for Nuclear Integrity and Chromosome Segregation in Mice. Dev. Cell 2005, 9, 769–779. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X. SUMO-Mediated Regulation of Nuclear Functions and Signaling Processes. Mol. Cell 2018, 71, 409–418. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, I.; Mateos-Langerak, J.; Thomas, A.; Cheutin, T.; Cavalli, G. Identification of Regulators of the Three-Dimensional Polycomb Organization by a Microscopy-Based Genome-Wide RNAi Screen. Mol. Cell 2014, 54, 485–499. [Google Scholar] [CrossRef] [Green Version]
- Salomoni, P. The PML-Interacting Protein DAXX: Histone Loading Gets into the Picture. Front. Oncol. 2013, 3, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kagey, M.H.; Melhuish, T.A.; Wotton, D. The Polycomb Protein Pc2 Is a SUMO E3. Cell 2003, 113, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Kagey, M.H.; Melhuish, T.A.; Powers, S.E.; Wotton, D. Multiple Activities Contribute to Pc2 E3 Function. EMBO J. 2005, 24, 108–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Sharrocks, A.D. The SUMO E3 Ligase Activity of Pc2 Is Coordinated through a SUMO Interaction Motif. Mol. Cell. Biol. 2010, 30, 2193–2205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merrill, J.C.; Melhuish, T.A.; Kagey, M.H.; Yang, S.-H.; Sharrocks, A.D.; Wotton, D. A Role for Non-Covalent SUMO Interaction Motifs in Pc2/CBX4 E3 Activity. PLoS ONE 2010, 5, e8794. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Zhou, J.; Liu, P.; Hu, J.; Jin, H.; Shimono, Y.; Takahashi, M.; Xu, G. Polycomb Protein Cbx4 Promotes SUMO Modification of de Novo DNA Methyltransferase Dnmt3a. Biochem. J. 2007, 405, 369–378. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Xu, Y.; Long, X.-D.; Wang, W.; Jiao, H.-K.; Mei, Z.; Yin, Q.-Q.; Ma, L.-N.; Zhou, A.-W.; Wang, L.-S.; et al. Cbx4 Governs HIF-1α to Potentiate Angiogenesis of Hepatocellular Carcinoma by Its SUMO E3 Ligase Activity. Cancer Cell 2014, 25, 118–131. [Google Scholar] [CrossRef] [Green Version]
- Kang, X.; Qi, Y.; Zuo, Y.; Wang, Q.; Zou, Y.; Schwartz, R.J.; Cheng, J.; Yeh, E.T.H. SUMO-Specific Protease 2 Is Essential for Suppression of Polycomb Group Protein-Mediated Gene Silencing during Embryonic Development. Mol. Cell 2010, 38, 191–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Tardat, M.; Gill, M.E.; Royo, H.; Thierry, R.; Ozonov, E.A.; Peters, A.H. SUMOylated PRC1 Controls Histone H3.3 Deposition and Genome Integrity of Embryonic Heterochromatin. EMBO J. 2020, 39, e103697. [Google Scholar] [CrossRef]
- Martin, N.; Schwamborn, K.; Schreiber, V.; Werner, A.; Guillier, C.; Zhang, X.-D.; Bischof, O.; Seeler, J.-S.; Dejean, A. PARP-1 Transcriptional Activity Is Regulated by Sumoylation upon Heat Shock. EMBO J. 2009, 28, 3534–3548. [Google Scholar] [CrossRef] [PubMed]
- Shiio, Y.; Eisenman, R.N. Histone Sumoylation Is Associated with Transcriptional Repression. Proc. Natl. Acad. Sci. USA 2003, 100, 13225–13230. [Google Scholar] [CrossRef] [Green Version]
- Nathan, D.; Ingvarsdottir, K.; Sterner, D.E.; Bylebyl, G.R.; Dokmanovic, M.; Dorsey, J.A.; Whelan, K.A.; Krsmanovic, M.; Lane, W.S.; Meluh, P.B.; et al. Histone Sumoylation Is a Negative Regulator in Saccharomyces Cerevisiae and Shows Dynamic Interplay with Positive-Acting Histone Modifications. Genes Dev. 2006, 20, 966–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.-T.; Alpert, A.; Leiter, C.; Gong, F.; Jackson, S.P.; Miller, K.M. Systematic Identification of Functional Residues in Mammalian Histone H2AX. Mol. Cell. Biol. 2013, 33, 111–126. [Google Scholar] [CrossRef] [Green Version]
- Kalocsay, M.; Hiller, N.J.; Jentsch, S. Chromosome-Wide Rad51 Spreading and SUMO-H2A.Z-Dependent Chromosome Fixation in Response to a Persistent DNA Double-Strand Break. Mol. Cell 2009, 33, 335–343. [Google Scholar] [CrossRef]
- Fukuto, A.; Ikura, M.; Ikura, T.; Sun, J.; Horikoshi, Y.; Shima, H.; Igarashi, K.; Kusakabe, M.; Harata, M.; Horikoshi, N.; et al. SUMO Modification System Facilitates the Exchange of Histone Variant H2A.Z-2 at DNA Damage Sites. Nucl. Austin Tex. 2018, 9, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Dhall, A.; Wei, S.; Fierz, B.; Woodcock, C.L.; Lee, T.-H.; Chatterjee, C. Sumoylated Human Histone H4 Prevents Chromatin Compaction by Inhibiting Long-Range Internucleosomal Interactions. J. Biol. Chem. 2014, 289, 33827–33837. [Google Scholar] [CrossRef] [Green Version]
- Dhall, A.; Weller, C.E.; Chu, A.; Shelton, P.M.M.; Chatterjee, C. Chemically Sumoylated Histone H4 Stimulates Intranucleosomal Demethylation by the LSD1-CoREST Complex. ACS Chem. Biol. 2017, 12, 2275–2280. [Google Scholar] [CrossRef]
- Ryu, H.-Y.; Su, D.; Wilson-Eisele, N.R.; Zhao, D.; López-Giráldez, F.; Hochstrasser, M. The Ulp2 SUMO Protease Promotes Transcription Elongation through Regulation of Histone Sumoylation. EMBO J. 2019, 38, e102003. [Google Scholar] [CrossRef]
- Jain, N.; Tamborrini, D.; Evans, B.; Chaudhry, S.; Wilkins, B.J.; Neumann, H. Interaction of RSC Chromatin Remodeling Complex with Nucleosomes Is Modulated by H3 K14 Acetylation and H2B SUMOylation In Vivo. iScience 2020, 23, 101292. [Google Scholar] [CrossRef]
- Debaugny, R.E.; Skok, J.A. CTCF and CTCFL in Cancer. Curr Opin. Genet. Dev. 2020, 61, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Kitchen, N.S.; Schoenherr, C.J. Sumoylation Modulates a Domain in CTCF That Activates Transcription and Decondenses Chromatin. J. Cell. Biochem. 2010, 111, 665–675. [Google Scholar] [CrossRef]
- Capelson, M.; Corces, V.G. SUMO Conjugation Attenuates the Activity of the Gypsy Chromatin Insulator. EMBO J. 2006, 25, 1906–1914. [Google Scholar] [CrossRef]
- Golovnin, A.; Volkov, I.; Georgiev, P. SUMO Conjugation Is Required for the Assembly of Drosophila Su(Hw) and Mod(Mdg4) into Insulator Bodies That Facilitate Insulator Complex Formation. J. Cell Sci. 2012, 125, 2064–2074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jox, T.; Buxa, M.K.; Bohla, D.; Ullah, I.; Mačinković, I.; Brehm, A.; Bartkuhn, M.; Renkawitz, R. Drosophila CP190- and DCTCF-Mediated Enhancer Blocking Is Augmented by SUMOylation. Epigenetics Chromatin 2017, 10, 32. [Google Scholar] [CrossRef] [PubMed]
- Dobreva, G.; Dambacher, J.; Grosschedl, R. SUMO Modification of a Novel MAR-Binding Protein, SATB2, Modulates Immunoglobulin Mu Gene Expression. Genes Dev. 2003, 17, 3048–3061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Banerjee, T.; Guan, X.; Freitas, M.A.; Parvin, J.D. The Chromatin Scaffold Protein SAFB1 Localizes SUMO-1 to the Promoters of Ribosomal Protein Genes to Facilitate Transcription Initiation and Splicing. Nucleic Acids Res. 2015, 43, 3605–3613. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.-A.T.; Sun, Y.; Song, J.; Chen, Y.; Krontiris, T.G.; Durrin, L.K. SUMO Conjugation to the Matrix Attachment Region-Binding Protein, Special AT-Rich Sequence-Binding Protein-1 (SATB1), Targets SATB1 to Promyelocytic Nuclear Bodies Where It Undergoes Caspase Cleavage. J. Biol. Chem. 2008, 283, 18124–18134. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.P.; Bischof, O.; Purbey, P.K.; Notani, D.; Urlaub, H.; Dejean, A.; Galande, S. Functional Interaction between PML and SATB1 Regulates Chromatin-Loop Architecture and Transcription of the MHC Class I Locus. Nat. Cell Biol. 2007, 9, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Mahmud, I.; Liao, D. DAXX in Cancer: Phenomena, Processes, Mechanisms and Regulation. Nucleic Acids Res. 2019, 47, 7734–7752. [Google Scholar] [CrossRef] [Green Version]
- Delbarre, E.; Ivanauskiene, K.; Spirkoski, J.; Shah, A.; Vekterud, K.; Moskaug, J.Ø.; Bøe, S.O.; Wong, L.H.; Küntziger, T.; Collas, P. PML Protein Organizes Heterochromatin Domains Where It Regulates Histone H3.3 Deposition by ATRX/DAXX. Genome Res. 2017, 27, 913–921. [Google Scholar] [CrossRef] [Green Version]
- Penagos-Puig, A.; Furlan-Magaril, M. Heterochromatin as an Important Driver of Genome Organization. Front. Cell Dev. Biol. 2020, 8, 579137. [Google Scholar] [CrossRef]
- Stielow, B.; Sapetschnig, A.; Wink, C.; Krüger, I.; Suske, G. SUMO-Modified Sp3 Represses Transcription by Provoking Local Heterochromatic Gene Silencing. EMBO Rep. 2008, 9, 899–906. [Google Scholar] [CrossRef] [Green Version]
- Stielow, B.; Krüger, I.; Diezko, R.; Finkernagel, F.; Gillemans, N.; Kong-a-San, J.; Philipsen, S.; Suske, G. Epigenetic Silencing of Spermatocyte-Specific and Neuronal Genes by SUMO Modification of the Transcription Factor Sp3. PLoS Genet. 2010, 6, e1001203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.A.; Choi, E.S.; Kim, H.S.; Ho, J.C.Y.; Watts, F.Z.; Park, S.D.; Jang, Y.K. SUMO Modification Is Involved in the Maintenance of Heterochromatin Stability in Fission Yeast. Mol. Cell 2005, 19, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Maison, C.; Bailly, D.; Roche, D.; Montes de Oca, R.; Probst, A.V.; Vassias, I.; Dingli, F.; Lombard, B.; Loew, D.; Quivy, J.-P.; et al. SUMOylation Promotes de Novo Targeting of HP1α to Pericentric Heterochromatin. Nat. Genet. 2011, 43, 220–227. [Google Scholar] [CrossRef] [Green Version]
- Maison, C.; Bailly, D.; Quivy, J.-P.; Almouzni, G. The Methyltransferase Suv39h1 Links the SUMO Pathway to HP1α Marking at Pericentric Heterochromatin. Nat. Commun. 2016, 7, 12224. [Google Scholar] [CrossRef] [PubMed]
- Maison, C.; Romeo, K.; Bailly, D.; Dubarry, M.; Quivy, J.-P.; Almouzni, G. The SUMO Protease SENP7 Is a Critical Component to Ensure HP1 Enrichment at Pericentric Heterochromatin. Nat. Struct. Mol. Biol. 2012, 19, 458–460. [Google Scholar] [CrossRef] [PubMed]
- Romeo, K.; Louault, Y.; Cantaloube, S.; Loiodice, I.; Almouzni, G.; Quivy, J.-P. The SENP7 SUMO-Protease Presents a Module of Two HP1 Interaction Motifs That Locks HP1 Protein at Pericentric Heterochromatin. Cell Rep. 2015, 10, 771–782. [Google Scholar] [CrossRef] [PubMed]
- Garvin, A.J.; Densham, R.M.; Blair-Reid, S.A.; Pratt, K.M.; Stone, H.R.; Weekes, D.; Lawrence, K.J.; Morris, J.R. The DeSUMOylase SENP7 Promotes Chromatin Relaxation for Homologous Recombination DNA Repair. EMBO Rep. 2013, 14, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, K.; Shinkai, Y. SETDB1-Mediated Silencing of Retroelements. Viruses 2020, 12, 596. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.X.; El Farran, C.A.; Guo, H.C.; Yu, T.; Fang, H.T.; Wang, H.F.; Schlesinger, S.; Seah, Y.F.S.; Goh, G.Y.L.; Neo, S.P.; et al. Systematic Identification of Factors for Provirus Silencing in Embryonic Stem Cells. Cell 2015, 163, 230–245. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, K.; Okuda, A.; Yusa, K.; Shinkai, Y. A CRISPR Knockout Screen Identifies SETDB1-Target Retroelement Silencing Factors in Embryonic Stem Cells. Genome Res. 2018, 28, 846–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, A.V.; Peng, H.; Yurchenko, V.; Yap, K.L.; Negorev, D.G.; Schultz, D.C.; Psulkowski, E.; Fredericks, W.J.; White, D.E.; Maul, G.G.; et al. PHD Domain-Mediated E3 Ligase Activity Directs Intramolecular Sumoylation of an Adjacent Bromodomain Required for Gene Silencing. Mol. Cell 2007, 28, 823–837. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boulanger, M.; Chakraborty, M.; Tempé, D.; Piechaczyk, M.; Bossis, G. SUMO and Transcriptional Regulation: The Lessons of Large-Scale Proteomic, Modifomic and Genomic Studies. Molecules 2021, 26, 828. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26040828
Boulanger M, Chakraborty M, Tempé D, Piechaczyk M, Bossis G. SUMO and Transcriptional Regulation: The Lessons of Large-Scale Proteomic, Modifomic and Genomic Studies. Molecules. 2021; 26(4):828. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26040828
Chicago/Turabian StyleBoulanger, Mathias, Mehuli Chakraborty, Denis Tempé, Marc Piechaczyk, and Guillaume Bossis. 2021. "SUMO and Transcriptional Regulation: The Lessons of Large-Scale Proteomic, Modifomic and Genomic Studies" Molecules 26, no. 4: 828. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26040828