
Spatio-Temporal Variation of the Bacterial Communities along a Salinity Gradient within a Thalassohaline Environment (Saline di Tarquinia Salterns, Italy)
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
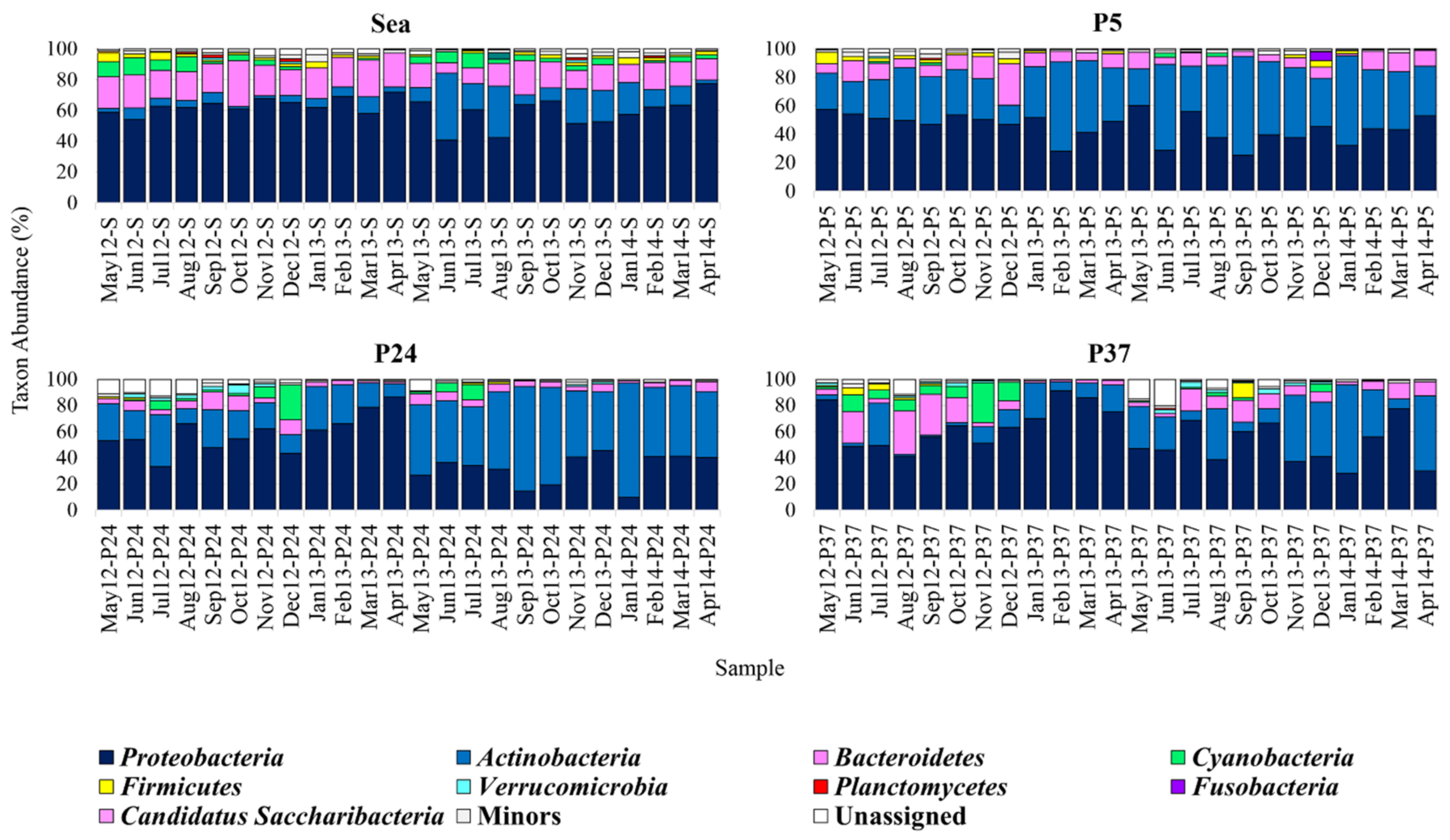
2.1. Composition of the Bacterial Communities
2.2. Alpha- and Beta-Diversity of the Bacterial Communities
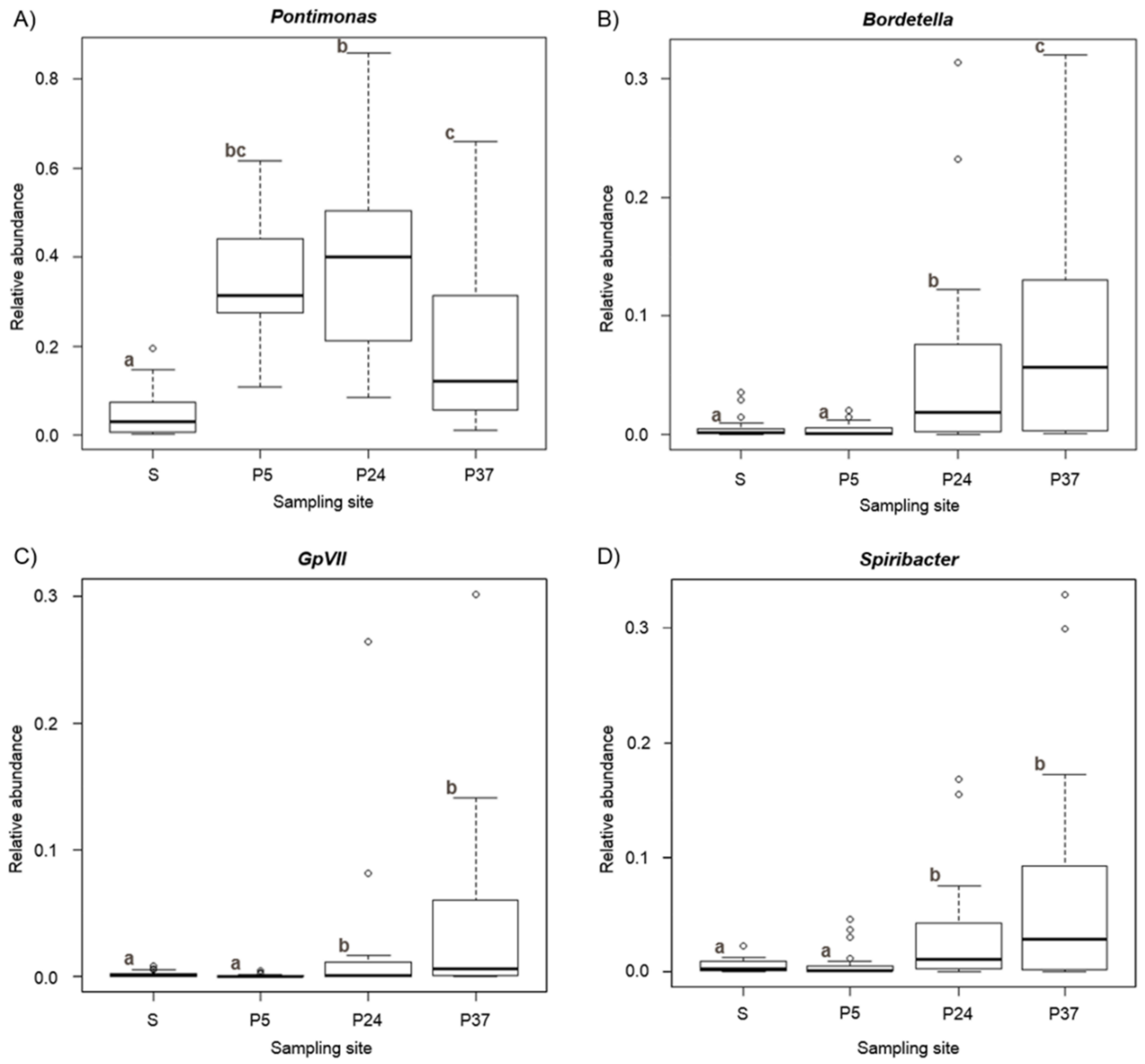
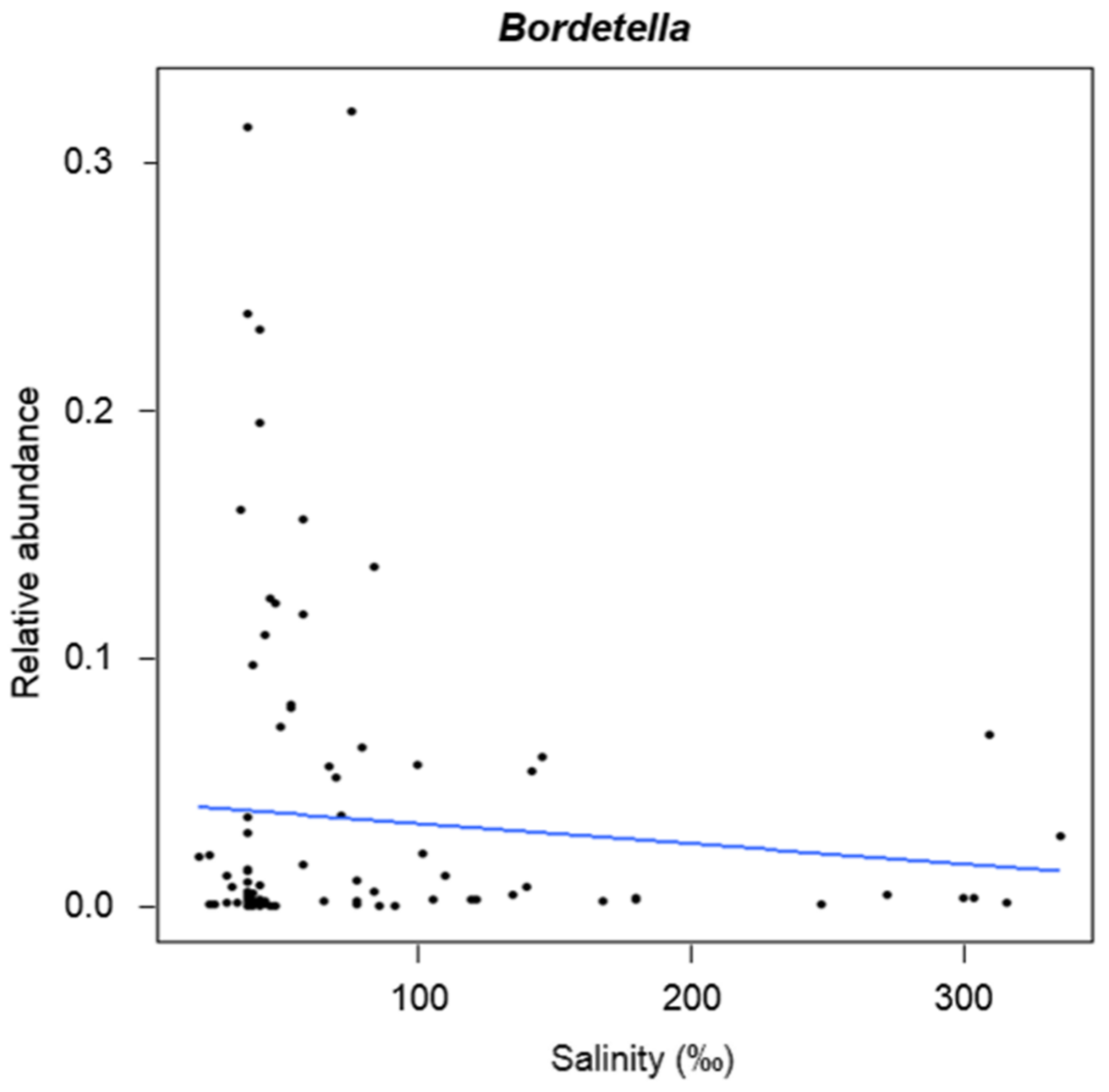
2.3. Variation of α-Diversity Indices and General Abundance in Relation to the Parameters Structuring the Bacterial Communities
3. Discussion
4. Materials and Methods
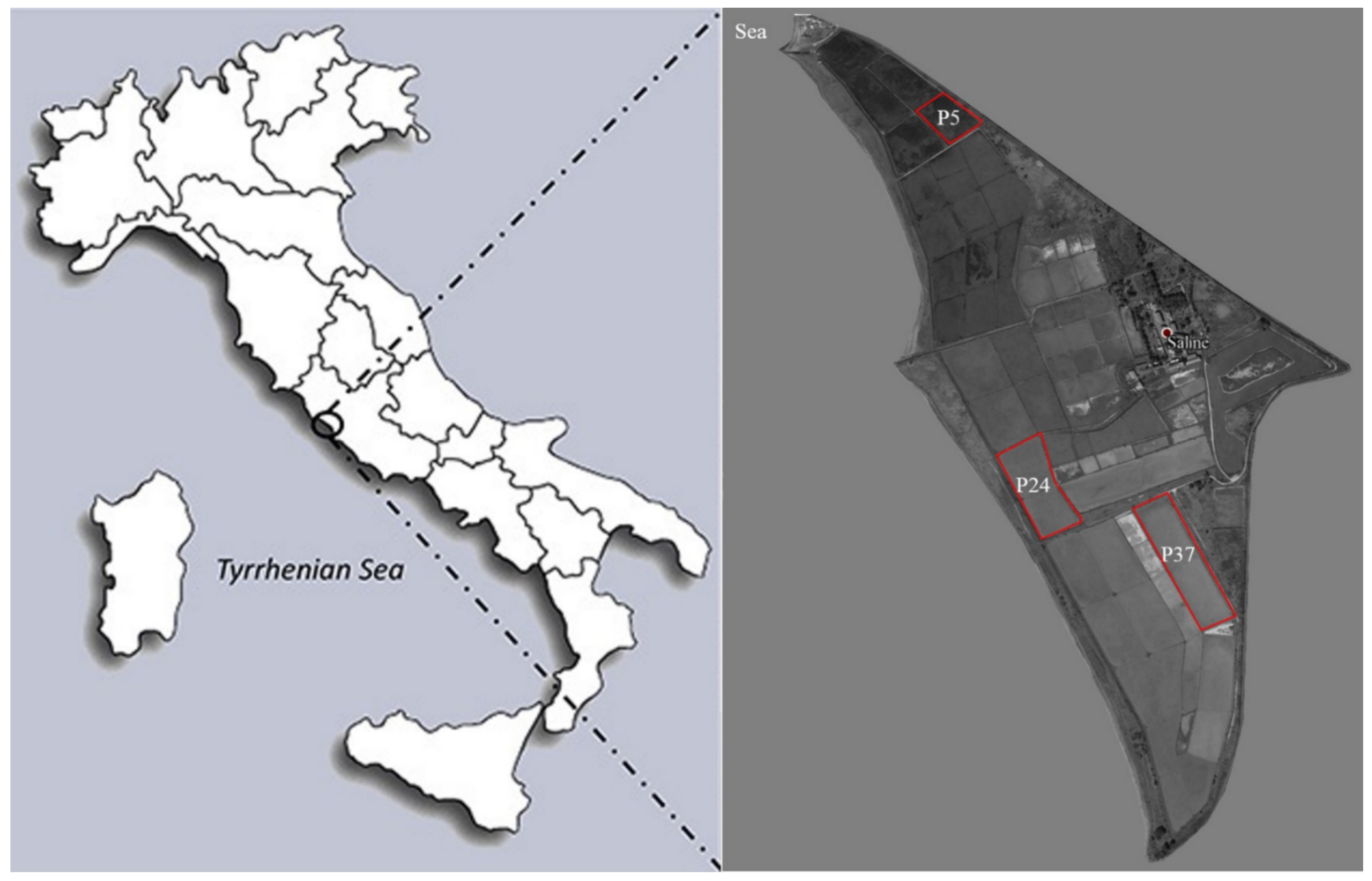
4.1. Sample Collection and Characterisation
4.2. DNA Extraction, 16S rDNA Amplicon Libraries and Sequencing
4.3. Sequence Processing and Data Analysis
4.4. Statistical Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Reboleiro-Rivas, P.; Juarez-Jimenez, B.; Martinez-Toledo, M.V.; Rodelas, M.; Andrade, L.; Gonzalez-Lopez, J.; Fenice, M. Bacterial Communities Structure in a High Mountain Lake During the Ice-free Season: Cultural and PCR-TGGE Investigations. Int. J. Environ. Res. 2013, 7, 685–696. [Google Scholar]
- Andrade, L.; González-López, J.; Fenice, M.; Martínez-Toledo, M.V.; Pesciaroli, C.; Maza-Márquez, P.; Juárez-Jiménez, B. Application of Response Surface Methodology (RSM) for Culture Conditions and Biomass Production of Psychrophilic Microalgae Isolated from High Mountains Lake During the Ice-free Season. Int. J. Environ. Res. 2014, 8, 799–812. [Google Scholar] [CrossRef]
- Barghini, P.; Pasqualetti, M.; Gorrasi, S.; Fenice, M. Bacteria from the “Saline di Tarquinia” Marine Salterns Reveal Very Atypical Growth Profiles with Regards to Salinity and Temperature. Mediterr. Mar. Sci. 2018, 19, 513–525. [Google Scholar] [CrossRef] [Green Version]
- Ventosa, A.; Fernández, A.B.; León, M.J.; Sánchez-Porro, C.; Rodriguez-Valera, F. The Santa Pola Saltern as a Model for Studying the Microbiota of Hypersaline Environments. Extremophiles 2014, 18, 811–824. [Google Scholar] [CrossRef] [PubMed]
- Halse, S.A.; Cale, D.; Jasinska, E.J.; Shiel, R.J. Monitoring Change in Aquatic Invertebrate Biodiversity: Sample Size, Faunal elements and Analytical Methods. Aquat. Ecol. 2002, 36, 395–410. [Google Scholar] [CrossRef]
- Cimmaruta, R.; Blasi, S.; Angeletti, D.; Nascetti, G. The Recent History of the Tarquinia Salterns Offers the Opportunity to Investigate Parallel Changes at the Habitat and Biodiversity Levels. Transit. Water. Bull. 2010, 4, 53–59. [Google Scholar]
- Barghini, P.; Silvi, S.; Aquilanti, A.; Marcelli, M.; Fenice, M. Bacteria from Marine Salterns as a Model of Microorganisms Adapted to High Environmental Variations. J. Environ. Prot. Ecol. 2014, 15, 897. [Google Scholar]
- Barghini, P.; Giovannini, V.; Fenice, M.; Gorrasi, S.; Pasqualetti, M. High Lutein Production by a Halo-tolerant Strain of Dunaliella sp. (Chlorophyceae) Isolated from Solar Salterns in Central Italy. J. Environ. Prot. Ecol. 2018, 19, 704–712. [Google Scholar]
- Pedrós-Alió, C. Trophic Ecology of Solar Salterns. In Halophilic Microorganisms; Ventosa, A., Ed.; Springer: Heidelberg/Berlin, Germany, 2004; pp. 33–48. ISBN 978-3-642-05664-2. [Google Scholar]
- Barghini, P.; Aquilanti, A.; Silvi, S.; Gorrasi, S.; Juarez-Jimenez, B.; Cordova-Salas, K.; Fenice, M. Study on the Bacterial Diversity of the Productive Marine Salterns ‘Saline di Trapani-Paceco’, Trapani, Italy. J. Environ. Prot. Ecol. 2015, 16, 1389–1396. [Google Scholar]
- Forster, D.; Behnke, A.; Stoeck, T. Meta-analyses of Environmental Sequence Data Identify Anoxia and Salinity as Parameters Shaping Ciliate Communities. Syst. Biodivers. 2012, 10, 277–288. [Google Scholar] [CrossRef]
- Filker, S.; Forster, D.; Weinisch, L.; Mora-Ruiz, M.; González, B.; Farías, M.E.; Rosselló-Móra, R.; Stoeck, T. Transition Boundaries for Protistan Species Turnover in Hypersaline Waters of Different Biogeographic Regions. Environ. Microbiol. 2017, 19, 3186–3200. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Knight, R. Global Patterns in Bacterial Diversity. Proc. Natl. Acad. Sci. USA 2007, 104, 11436–11440. [Google Scholar] [CrossRef] [Green Version]
- Oren, A. Microscopic Examination of Microbial Communities Along a Salinity Gradient in Saltern Evaporation Ponds: A ‘Halophilic Safari’. In Adaptation to Life at High Salt Concentrations in Archaea, Bacteria, and Eukarya, 1st ed.; Gunde-Cimerman, N., Oren, A., Plemenitaš, A., Eds.; Springer: Dordrecht, The Netherlands, 2005; Volume 9, pp. 41–57. ISBN 978-1-4020-3632-3. [Google Scholar]
- Salgaonkar, B.B.; Mani, K.; Bragança, J.M. Accumulation of Polyhydroxyalkanoates by Halophilic Archaea Isolated from Traditional Solar Salterns of India. Extremophiles 2013, 17, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Barghini, P.; Pasqualetti, M.; Gorrasi, S.; Fenice, M. Study of Bacterial Diversity of a Saltern Crystallisation Pond (‘Saline di Tarquinia’, Italy) and Its Correlation with Salinity Variations. J. Environ. Prot. Ecol. 2018, 19, 139–145. [Google Scholar]
- Benlloch, S.; López-López, A.; Casamayor, E.O.; Øvreås, L.; Goddard, V.; Daae, F.L.; Smerdon, G.; Massana, R.; Joint, I.; Thingstad, F.; et al. Prokaryotic Genetic Diversity Throughout the Salinity Gradient of a Coastal Solar Saltern. Environ. Microbiol. 2002, 4, 349–360. [Google Scholar] [CrossRef]
- Park, S.J.; Kang, C.H.; Rhee, S.K. Characterization of the Microbial Diversity in a Korean Solar Saltern by 16S rRNA Gene Analysis. J. Microbiol. Biotechnol. 2006, 16, 1640–1645. [Google Scholar]
- Baati, H.; Guermazi, S.; Amdouni, R.; Gharsallah, N.; Sghir, A.; Ammar, E. Prokaryotic Diversity of a Tunisian Multipond Solar Saltern. Extremophiles 2008, 12, 505–518. [Google Scholar] [CrossRef]
- Tsiamis, G.; Katsaveli, K.; Ntougias, S.; Kyrpides, N.; Andersen, G.; Piceno, Y.; Bourtzis, K. Prokaryotic Community Profiles at Different Operational Stages of a Greek Solar Saltern. Res. Microbiol. 2008, 159, 609–627. [Google Scholar] [CrossRef] [PubMed]
- Trigui, H.; Masmoudi, S.; Brochier-Armanet, C.; Barani, A.; Grégori, G.; Denis, M.; Dukan, S.; Maalej, S. Characterization of Heterotrophic Prokaryote Subgroups in the Sfax Coastal Solar Salterns by Combining Flow Cytometry Cell Sorting and Phylogenetic Analysis. Extremophiles 2011, 15, 347–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boujelben, I.; Gomariz, M.; Martínez-García, M.; Santos, F.; Peña, A.; López, C.; Antón, J.; Maalej, S. Spatial and Seasonal Prokaryotic Community Dynamics in Ponds of Increasing Salinity of Sfax Solar Saltern in Tunisia. Antonie Van Leeuwenhoek 2012, 101, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Dillon, J.G.; Carlin, M.; Gutierrez, A.; Nguyen, V.; McLain, N. Patterns of Microbial Diversity Along a Salinity Gradient in the Guerrero Negro Solar Saltern, Baja CA Sur, Mexico. Front. Microbiol. 2013, 4, 399. [Google Scholar] [CrossRef] [Green Version]
- Gomariz, M.; Martínez-García, M.; Santos, F.; Rodriguez, F.; Capella-Gutiérrez, S.; Gabaldon, T.; Rosselló-Móra, R.; Meseguer, I.; Antón, J. From Community Approaches to Single-Cell Genomics: The Discovery of Ubiquitous Hyperhalophilic Bacteroidetes Generalists. ISME J. 2015, 9, 16–31. [Google Scholar] [CrossRef]
- Mutlu, M.B.; Güven, K. Bacterial Diversity in Çamaltı Saltern, Turkey. Pol. J. Microbiol. 2015, 64, 37–45. [Google Scholar] [CrossRef]
- Zhang, J.; Ma, G.; Deng, Y.; Dong, J.; Van Stappen, G.; Sui, L. Bacterial Diversity in Bohai Bay Solar Saltworks, China. Curr. Microbiol. 2016, 72, 55–63. [Google Scholar] [CrossRef]
- Kambourova, M.; Tomova, I.; Boyadzhieva, I.; Radchenkova, N.; Vasileva-Tonkova, E. Phylogenetic Analysis of the Bacterial Community in a Crystallizer Pond, Pomorie Salterns, Bulgaria. Biotechnol. Biotechnol. Equip. 2017, 31, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Ghai, R.; Pašić, L.; Fernández, A.B.; Martin-Cuadrado, A.B.; Mizuno, C.M.; McMahon, K.D.; Papke, R.T.; Stepanauskas, R.; Rodriguez-Brito, B.; Rohwer, F.; et al. New Abundant Microbial Groups in Aquatic Hypersaline Environments. Sci. Rep. 2011, 1, 135. [Google Scholar] [CrossRef]
- Fernández, A.B.; Ghai, R.; Martin-Cuadrado, A.B.; Sanchez-Porro, C.; Rodriguez-Valera, F.; Ventosa, A. Metagenome Sequencing of Prokaryotic Microbiota from Two Hypersaline Ponds of a Marine Saltern in Santa Pola, Spain. Genome. Announc. 2013, 1. [Google Scholar] [CrossRef] [Green Version]
- Fernández, A.B.; León, M.J.; Vera, B.; Sánchez-Porro, C.; Ventosa, A. Metagenomic Sequence of Prokaryotic Microbiota from an Intermediate-salinity Pond of a Saltern in Isla Cristina, Spain. Genome. Announc. 2014, 2, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Fernández, A.B.; Ghai, R.; Martin-Cuadrado, A.B.; Sanchez-Porro, C.; Rodriguez-Valera, F.; Ventosa, A. Prokaryotic Taxonomic and Metabolic Diversity of an Intermediate Salinity Hypersaline Habitat Assessed by Metagenomics. FEMS Microbiol. Ecol. 2014, 88, 623–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plominsky, A.M.; Delherbe, N.; Ugalde, J.A.; Allen, E.E.; Blanchet, M.; Ikeda, P.; Santibanez, F.; Hanselmann, K.; Ulloa, O.; De la Iglesia, R.; et al. Metagenome Sequencing of the Microbial Community of a Solar Saltern Crystallizer Pond at Cáhuil Lagoon, Chile. Genome. Announc. 2014, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plominsky, M.A.; Henríquez, C.A.; Delherbe, N.; Podell, S.; Ramirez, S.; Ugalde, J.A.; Santibañez, F.; van den Engh, G.; Hanselmann, K.; Ulloa, O.; et al. Distinctive Archaeal Composition of an Artisanal Crystallizer Pond and Functional Insights into Salt-saturated Hypersaline Environment Adaptation. Front. Microbiol. 2018, 9, 1800. [Google Scholar] [CrossRef] [Green Version]
- Leoni, C.; Volpicella, M.; Fosso, B.; Manzari, C.; Piancone, E.; Dileo, M.C.; Arcadi, E.; Yakimov, M.; Pesol, G.; Ceci, L.R. A Differential Metabarcoding Approach to Describe Taxonomy Profiles of Bacteria and Archaea in the Saltern of Margherita di Savoia (Italy). Microorganisms 2020, 8, 936. [Google Scholar] [CrossRef] [PubMed]
- John, J.; Siva, V.; Kumari, R.; Arya, A.; Kumar, A. Unveiling Cultivable and Uncultivable Halophilic Bacteria Inhabiting Marakkanam Saltpan, India and Their Potential for Biotechnological Applications. Geomicrobiol. J. 2020, 37, 691–701. [Google Scholar] [CrossRef]
- Fernández, A.B.; Vera-Gargallo, B.; Sánchez-Porro, C.; Ghai, R.; Papke, R.T.; Rodriguez-Valera, F.; Ventosa, A. Comparison of Prokaryotic Community Structure from Mediterranean and Atlantic Saltern Concentrator Ponds by a Metagenomic Approach. Front. Microbiol. 2014, 5, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Villegas, P.; Vigara, J.; León, R. Characterization of the Microbial Population Inhabiting a Solar Saltern Pond of the Odiel Marshlands (SW Spain). Mar. Drugs. 2018, 16, 332. [Google Scholar] [CrossRef] [Green Version]
- Mora-Ruiz, M.D.R.; Cifuentes, A.; Font-Verdera, F.; Pérez-Fernández, C.; Farias, M.E.; González, B.; Orfila, A.; Rosselló-Móra, R. Biogeographical Patterns of Bacterial and Archaeal Communities from Distant Hypersaline Environments. Syst. Appl. Microbiol. 2018, 41, 139–150. [Google Scholar] [CrossRef]
- Mani, K.; Taib, N.; Hugoni, M.; Bronner, G.; Bragança, J.M.; Debroas, D. Transient Dynamics of Archaea and Bacteria in Sediments and Brine Across a Salinity Gradient in a Solar Saltern of Goa, India. Front. Microbiol. 2020, 11, 1891. [Google Scholar] [CrossRef]
- Alfinito, S.; Iberite, M.; Fumanti, B. The Algal Microflora of the Salt Works of Tarquinia (Italy). Hydrobiology 1990, 203, 137–146. [Google Scholar] [CrossRef]
- Frondoni, R.; Iberite, M. The Halophile Vegetation of the Sedimentary Coast of Lazio (Central Tyrrhenian District, Italy). Plant. Biosyst. 2002, 136, 49–67. [Google Scholar] [CrossRef]
- Angeletti, D.; Cimmaruta, R.; Nascetti, G. Genetic Diversity of the Killifish Aphanius fasciatus Paralleling the Environmental Changes of Tarquinia Salterns Habitat. Genetica 2010, 138, 1011–1021. [Google Scholar] [CrossRef]
- Angeletti, D.; Cimmaruta, R.; Sebbio, C.; Bellisario, B.; Carere, C.; Nascetti, G. Environmental Heterogeneity Promotes Microgeographic Genetic Divergence in the Mediterranean Killifish Aphanius fasciatus (Cyprinodontidae). Ethol. Ecol. Evol. 2017, 29, 367–386. [Google Scholar] [CrossRef]
- Pasqualetti, M.; Giovannini, V.; Barghini, P.; Gorrasi, S.; Fenice, M. Diversity and Ecology of Culturable Marine Fungi Associated with Posidonia ceanica Leaves and Their Epiphytic Algae Dictyota Dichotoma and Sphaerococcus Coronopifolius. Fungal Ecol. 2020, 44, 100906. [Google Scholar] [CrossRef]
- Bellisario, B.; Cerfolli, F.; Nascetti, G. Spatial Network Structure and Robustness of Detritus-based Communities in a Patchy Environment. Ecol. Res. 2010, 25, 813–821. [Google Scholar] [CrossRef]
- Bellisario, B.; Cerfolli, F.; Nascetti, G. The Interplay Between Network Structure and Functioning of Detritus-based Communities in Patchy Aquatic Environment. Aquat. Ecol. 2012, 46, 431–441. [Google Scholar] [CrossRef]
- Bellisario, B.; Carere, C.; Cerfolli, F.; Angeletti, D.; Nascetti, G.; Cimmaruta, R. Infaunal Macrobenthic Community Dynamics in a Manipulated Hyperhaline Ecosystem: A Long-term Study. Aquat. Biosyst. 2013, 9, 20. [Google Scholar] [CrossRef] [Green Version]
- Cerfolli, F.; Bellisario, B.; Battisti, C. Detritus-based Assemblage Responses Under Salinity Stress Conditions in a Disused Aquatic Artificial Ecosystem. Aquat. Biosyst. 2013, 9, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasqualetti, M.; Bernini, R.; Carletti, L.; Crisante, F.; Tempesta, S. Salinity and Nitrate Concentration on the Growth and Carotenoids Accumulation in a Strain of Dunaliella salina (Chlorophyta) Cultivated Under Laboratory Conditions. Transit. Water. Bull. 2010, 4, 94–104. [Google Scholar] [CrossRef]
- Tempesta, S.; Paoletti, M.; Pasqualetti, M. Morphological and Molecular Identification of a Strain of the Unicellular Green Alga Dunaliella sp. Isolated from Tarquinia Salterns. Transit. Water. Bull. 2010, 4, 60–70. [Google Scholar] [CrossRef]
- Gorrasi, S.; Pasqualetti, M.; Franzetti, A.; Pittino, F.; Fenice, M. Vibrio Communities Along a Salinity Gradient Within a Marine Salterns Hypersaline Environment (Saline di Tarquinia, Italy). Environ. Microbiol. 2020, 22, 4356–4366. [Google Scholar] [CrossRef]
- Rodriguez-Brito, B.; Li, L.; Wegley, L.; Furlan, M.; Angly, F.; Breitbart, M.; Buchanan, J.; Desnues, C.; Dinsdale, E.; Edwards, R.; et al. Viral and Microbial Community Dynamics in Four Aquatic Environments. ISME J. 2010, 4, 739. [Google Scholar] [CrossRef]
- Kimbrel, J.A.; Ballor, N.; Wu, Y.W.; David, M.M.; Hazen, T.C.; Simmons, B.A.; Singer, S.W.; Jansson, J.K. Microbial Community Structure and Functional Potential Along a Hypersaline Gradient. Front. Microbiol. 2018, 9, 1492. [Google Scholar] [CrossRef] [PubMed]
- Pašić, L.; Rodriguez-Mueller, B.; Martin-Cuadrado, A.B.; Mira, A.; Rohwer, F.; Rodriguez-Valera, F. Metagenomic Islands of Hyperhalophiles: The Case of Salinibacter ruber. BMC Genom. 2009, 10, 570. [Google Scholar] [CrossRef] [Green Version]
- Zhaxybayeva, O.; Stepanauskas, R.; Mohan, N.R.; Papke, R.T. Cell Sorting Analysis of Geographically Separated Hypersaline Environments. Extremophiles 2013, 17, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Couto-Rodríguez, R.L.; Montalvo-Rodríguez, R. Temporal Analysis of the Microbial Community from the Crystallizer Ponds in Cabo Rojo, Puerto Rico, Using Metagenomics. Genes 2019, 10, 422. [Google Scholar] [CrossRef] [Green Version]
- Ramos-Barbero, M.D.; Martin-Cuadrado, A.B.; Viver, T.; Santos, F.; Martinez-Garcia, M.; Antón, J. Recovering Microbial Genomes from Metagenomes in Hypersaline Environments: The Good, the Bad and the Ugly. Syst. Appl. Microbiol. 2019, 42, 30–40. [Google Scholar] [CrossRef]
- Jang, G.I.; Cho, Y.; Cho, B.C. Pontimonas salivibrio gen. nov., sp. nov., A New Member of the Family Microbacteriaceae Isolated from a Seawater Reservoir of a Solar Saltern. Int. J. Syst. Evol. Microbiol. 2013, 63, 2124–2131. [Google Scholar] [CrossRef]
- Cho, B.C.; Jang, G.I.; Hwang, C.Y. Pontimonas. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.E.B., Ed.; John Wiley & Sons: New York, NY, USA, 2015; pp. 1–5. ISBN 9781118960608. [Google Scholar]
- Cho, B.C.; Hardies, S.C.; Jang, G.I.; Hwang, C.Y. Complete Genome of Streamlined Marine Actinobacterium Pontimonas salivibrio Strain CL-TW6 T Adapted to Coastal Planktonic Lifestyle. BMC Genom. 2018, 19, 625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selivanova, E.A.; Poshvina, D.V.; Khlopko, Y.A.; Gogoleva, N.E.; Plotnikov, A.O. Diversity of Prokaryotes in Planktonic Communities of Saline Sol-Iletsk Lakes (Orenburg Oblast, Russia). Microbiology 2018, 87, 569–582. [Google Scholar] [CrossRef]
- Shurigin, V.; Hakobyan, A.; Panosyan, H.; Egamberdieva, D.; Davranov, K.; Birkeland, N.K. A Glimpse of the Prokaryotic Diversity of the Large Aral Sea Reveals Novel Extremophilic Bacterial and Archaeal Groups. Microbiol. Open 2019, 8, e00850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanden, G.N.; Weyant, R.S. Bordetella. In Bergey’s Manual of Systematics of Archaea and Bacteria; Whitman, W.E.B., Ed.; John Wiley & Sons: New York, NY, USA, 2015; pp. 1–20. ISBN 9781118960608. [Google Scholar]
- Soumana, I.H.; Linz, B.; Harvill, E.T. Environmental Origin of the Genus Bordetella. Front. Microbiol. 2017, 8, 28. [Google Scholar] [CrossRef]
- Badamchi, A.; Papizadeh, M. Comparative Phylogeny of the Genus Bordetella Using Sequence Analysis of 16S rRNA and ompA Genes. J. Med. Bacteriol. 2017, 6, 1–13. [Google Scholar]
- Linz, B.; Ma, L.; Rivera, I.; Harvill, E.T. Genotypic and Phenotypic Adaptation of Pathogens: Lesson from the Genus: Bordetella. Curr. Opin. Infect. Dis. 2019, 32, 223–230. [Google Scholar] [CrossRef]
- Taylor-Mulneix, D.L.; Soumana, I.H.; Linz, B.; Harvill, E.T. Evolution of Bordetellae from Environmental Microbes to Human Respiratory Pathogens: Amoebae as a Missing Link. Front. Cell. Infect. Microbiol. 2017, 7, 510. [Google Scholar] [CrossRef]
- Kersters, K.; Hinz, K.H.; Hertle, A.; Segers, P.; Lievens, A.; Siegmann, O.; De Ley, J. Bordetella avium sp. nov., Isolated from the Respiratory Tracts of Turkeys and Other Birds. Int. J. Syst. Evol. Microbiol. 1984, 34, 56–70. [Google Scholar] [CrossRef] [Green Version]
- Raffel, T.R.; Register, K.B.; Marks, S.A.; Temple, L. Prevalence of Bordetella avium Infection in Selected Wild and Domesticated Birds in the Eastern USA. J. Wildl. Dis. 2002, 38, 40–46. [Google Scholar] [CrossRef] [Green Version]
- Stenzel, T.; Pestka, D.; Tykałowski, B.; Śmiałek, M.; Koncicki, A.; Bancerz-Kisiel, A. Detection of Bordetella avium by TaqMan Real-time PCR in Tracheal Swabs from Wildlife birds. Pol. J. Vet. Sci. 2017, 20, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Lanzuisi, E. Waterbirds of a Mediterranean Wetland, Tarquinia Salinas. Regional and Local Environmental Variables and Management Prescriptions. Ph.D. Thesis, Università degli Studi della Tuscia, Viterbo, Italy, 2009. [Google Scholar]
- Biondi, M. Check-list Degli Uccelli della RNPA Saline di Tarquinia (VT) Aggiornata al Maggio. In La Riserva Naturale Statale “Saline di Tarquinia”, 1st ed.; Colletti, L., Ed.; Corpo Forestale dello Stato, Ufficio territoriale per la Biodiversità di Roma: Roma, Italy, 2014; pp. 119–132. ISBN 978-88-909200-7-3. [Google Scholar]
- López-Pérez, M.; Ghai, R.; Leon, M.J.; Rodríguez-Olmos, Á.; Copa-Patiño, J.L.; Soliveri, J.; Sanchez-Porro, C.; Ventosa, A.; Rodriguez-Valera, F. Genomes of “Spiribacter”, a Streamlined, Successful Halophilic Bacterium. BMC Genom. 2013, 14, 787. [Google Scholar] [CrossRef] [Green Version]
- León, M.J.; Fernández, A.B.; Ghai, R.; Sánchez-Porro, C.; Rodriguez-Valera, F.; Ventosa, A. From Metagenomics to Pure Culture: Isolation and Characterization of the Moderately Halophilic Bacterium Spiribacter salinus gen. nov., sp. nov. Appl. Environ. Microbiol. 2014, 80, 3850–3857. [Google Scholar] [CrossRef] [Green Version]
- Antón, J.; Rosselló-Mora, R.; Rodríguez-Valera, F.; Amann, R. Extremely Halophilic Bacteria in Crystallizer Ponds from Solar Salterns. Appl. Environ. Microbiol. 2000, 66, 3052–3057. [Google Scholar] [CrossRef] [Green Version]
- Antón, J.; Oren, A.; Benlloch, S.; Rodríguez-Valera, F.; Amann, R.; Rosselló-Mora, R. Salinibacter ruber gen. nov., sp. nov., A Novel, Extremely Halophilic Member of the Bacteria from Saltern Crystallizer Ponds. Int. J. Syst. Evol. Microbiol. 2002, 52, 485–491. [Google Scholar] [CrossRef]
- León, M.J.; Aldeguer-Riquelme, B.; Antón, J.; Sánchez-Porro, C.; Ventosa, A. Spiribacter aquaticus sp. nov., A Novel Member of the Genus Spiribacter Isolated from a Saltern. Int. J. Syst. Evol. Microbiol. 2017, 67, 2947–2952. [Google Scholar] [CrossRef]
- Gorrasi, S.; Pesciaroli, C.; Barghini, P.; Pasqualetti, M.; Giovannini, V.; Fenice, M. Metagenetic Profiling of the Bacterial Communities of an Intertidal Pool in Kandalaksha Bay (White Sea, Russia). J. Environ. Prot. Ecol. 2019, 20, 1317–1324. [Google Scholar]
- Rice, E.W.; Baird, R.B.; Eaton, A.D.; Clesceri, L.S. Standard Methods for the Examination of Water and Wastewater, 22nd ed.; American Public Health Association: Washington, DC, USA, 2012; ISBN 9780875530130. [Google Scholar]
- Centro Funzionale Regionale, Regione Lazio, Annali. Available online: http://www.idrografico.regione.lazio.it/annali/index.htm (accessed on 31 August 2020).
- Pesciaroli, C.; Rodelas, B.; Juarez-Jiménez, B.; Barghini, P.; Fenice, M. Bacterial Community Structure of a Coastal Area in Kandalaksha Bay, White Sea, Russia: Possible Relation to Tidal Hydrodynamics. Ann. Microbiol. 2015, 65, 443–453. [Google Scholar] [CrossRef]
- Gorrasi, S.; Pesciaroli, C.; Barghini, P.; Pasqualetti, M.; Fenice, M. Structure and Diversity of the Bacterial Community of an Arctic Estuarine System (Kandalaksha Bay) Subject to Intense Tidal Currents. J. Mar. Syst. 2019, 196, 77–85. [Google Scholar] [CrossRef]
- Gandolfi, I.; Canedoli, C.; Imperato, V.; Tagliaferri, I.; Gkorezis, P.; Vangronsveld, J.; Padoa Schioppa, E.; Papacchini, M.; Bestetti, G.; Franzetti, A. Diversity and Hydrocarbon-degrading Potential of Epiphytic Microbial Communities on Platanus x acerifolia Leaves in an Urban Area. Environ. Pollut. 2017, 220, 650–658. [Google Scholar] [CrossRef] [PubMed]
- Pittino, F.; Maglio, M.; Gandolfi, I.; Azzoni, R.S.; Diolaiuti, G.; Ambrosini, R.; Franzetti, A. Bacterial Communities of Cryoconite Holes of a Temperate Alpine Glacier Show Both Seasonal Trends and Year-to-year Variability. Ann. Glaciol. 2018, 59, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Huber, J.A.; Welch, D.B.M.; Morrison, H.G.; Huse, S.M.; Neal, P.R.; Butterfield, D.A.; Sogin, M.L. Microbial Population Structures in the Deep Marine Biosphere. Science 2007, 318, 97–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Qian, P.Y. Conservative Fragments in Bacterial 16S rRNA Genes and Primer Design for 16S Ribosomal DNA Amplicons in Metagenomic Studies. PLoS ONE 2009, 4, e7401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. UPARSE: Highly Accurate OTU Sequences from Microbial Amplicon Reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Claesson, M.J.; O’Sullivan, O.; Wang, O.; Nikkilä, J.; Marchesi, J.R.; Smidt, H.; de Vos, W.M.; Ross, R.P.; O’Toole, P.W. Comparative Analysis of Pyrosequencing and a Phylogenetic Microarray for Exploring Microbial Community Structures in the Human Distal Intestine. PLoS ONE 2009, 4, e6669. [Google Scholar] [CrossRef] [Green Version]
- Good, I.J. The Population Frequencies of Species and the Estimation of Population Parameters. Biometrika 1953, 40, 237–264. [Google Scholar] [CrossRef]
- Shannon, C.E. A Mathematical Theory of Communication. Bell. Sys. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Gini, C. Variabilità e Mutabilità: Contributo allo Studio delle Distribuzioni e delle Relazioni Statistiche; Tipografia di Paolo Cuppini: Bologna, Italy, 1912. [Google Scholar]
- Wittebolle, L.; Marzorati, M.; Clement, L.; Balloi, A.; Daffonchio, D.; Heylen, K.; De Vos, P.; Verstraete, W.; Boon, N. Initial Community Evenness Favours Functionality under Selective Stress. Nature 2009, 458, 623–626. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. Waste Not, Want Not: Why Rarefying Microbiome Data is Inadmissible. PLoS Comput. Biol. 2014, 10, e1003531. [Google Scholar] [CrossRef] [Green Version]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012; ISBN 9780444538680. [Google Scholar]
- Zuur, A.F.; Hilbe, J.M.; Ieno, E.N. A Beginner’s Guide to GLM and GLMM with R: A Frequentist and Bayesian Perspective for Ecologists, 2nd ed.; Highland Statistics L.t.d.: Newburgh, UK, 2015; ISBN 9780957174139. [Google Scholar]
- Benjamini, Y.; Yekutieli, D. The Control of the False Discovery Rate in Multiple Testing under Dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | df | Variance | F | p |
|---|---|---|---|---|
| sin(Month) × cos(Month) | 1 | 0.126 | 2.1049 | 0.001 |
| Year | 2 | 0.313 | 2.6093 | 0.001 |
| Salinity | 1 | 0.098 | 1.6318 | 0.027 |
| Chlorophyll pigments | 1 | 0.0711 | 1.1830 | 0.255 |
| sin(Month) × Sampling site | 3 | 0.3489 | 1.936 | 0.001 |
| cos(Month) × Sampling site | 3 | 0.3479 | 1.9209 | 0.001 |
| Residuals | 79 | 4.745 | ||
| F10,79 = 3.254, p = 0.001, adjusted R2 = 0.275 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gorrasi, S.; Franzetti, A.; Ambrosini, R.; Pittino, F.; Pasqualetti, M.; Fenice, M. Spatio-Temporal Variation of the Bacterial Communities along a Salinity Gradient within a Thalassohaline Environment (Saline di Tarquinia Salterns, Italy). Molecules 2021, 26, 1338. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051338
Gorrasi S, Franzetti A, Ambrosini R, Pittino F, Pasqualetti M, Fenice M. Spatio-Temporal Variation of the Bacterial Communities along a Salinity Gradient within a Thalassohaline Environment (Saline di Tarquinia Salterns, Italy). Molecules. 2021; 26(5):1338. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051338
Chicago/Turabian StyleGorrasi, Susanna, Andrea Franzetti, Roberto Ambrosini, Francesca Pittino, Marcella Pasqualetti, and Massimiliano Fenice. 2021. "Spatio-Temporal Variation of the Bacterial Communities along a Salinity Gradient within a Thalassohaline Environment (Saline di Tarquinia Salterns, Italy)" Molecules 26, no. 5: 1338. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051338