
Identification and Quantification of Coumarins by UHPLC-MS in Arabidopsis thaliana Natural Populations

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
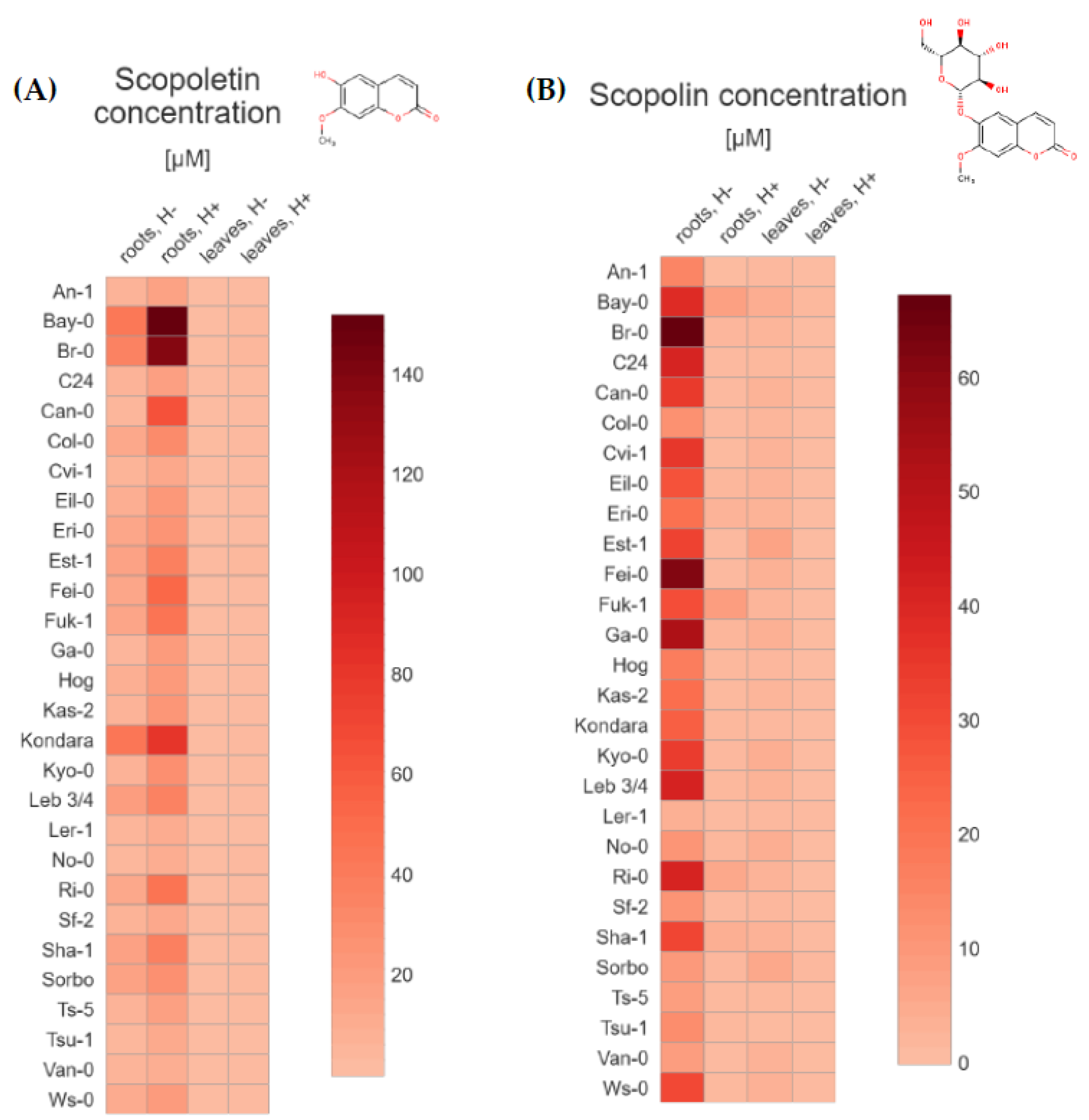
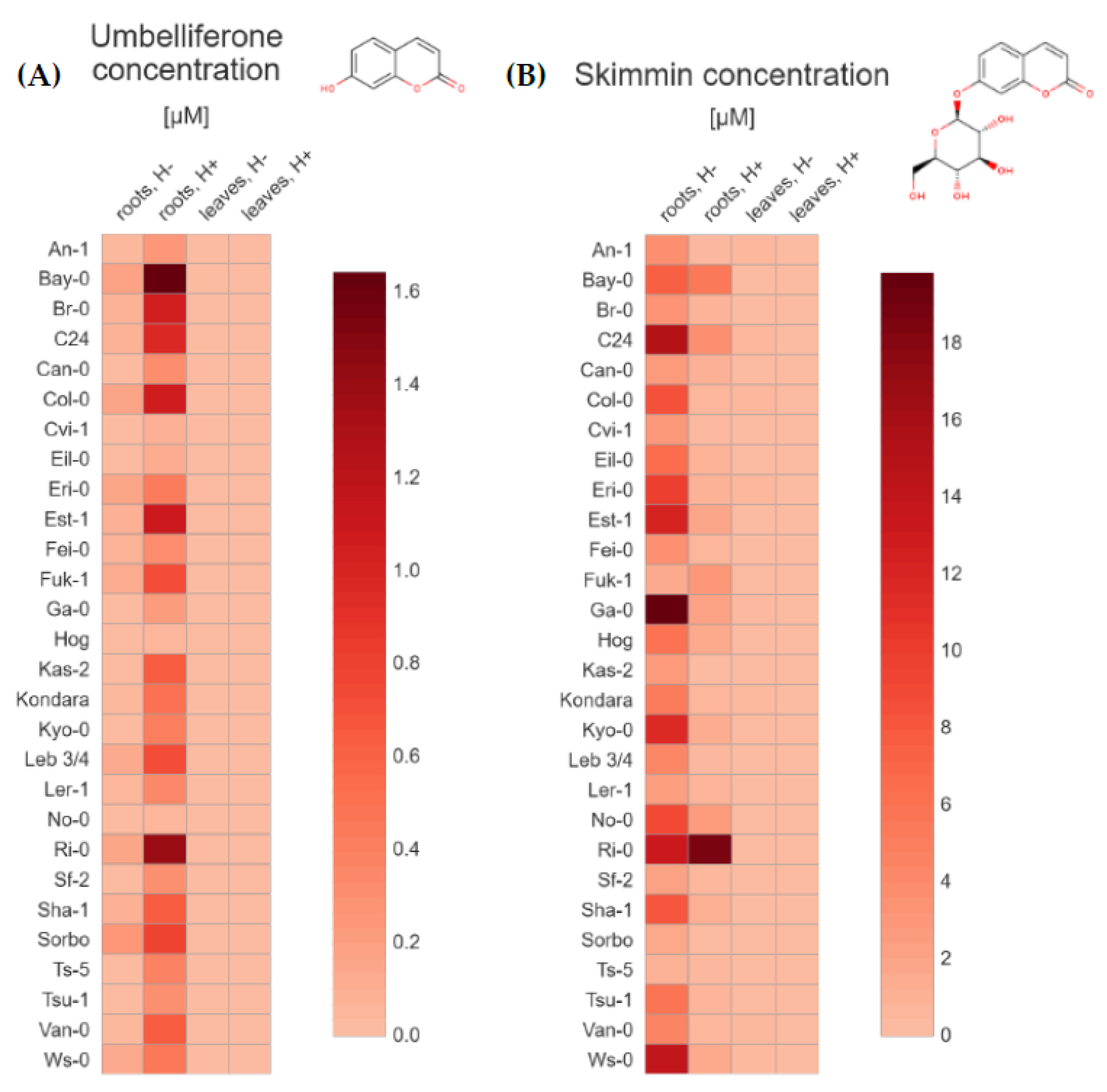
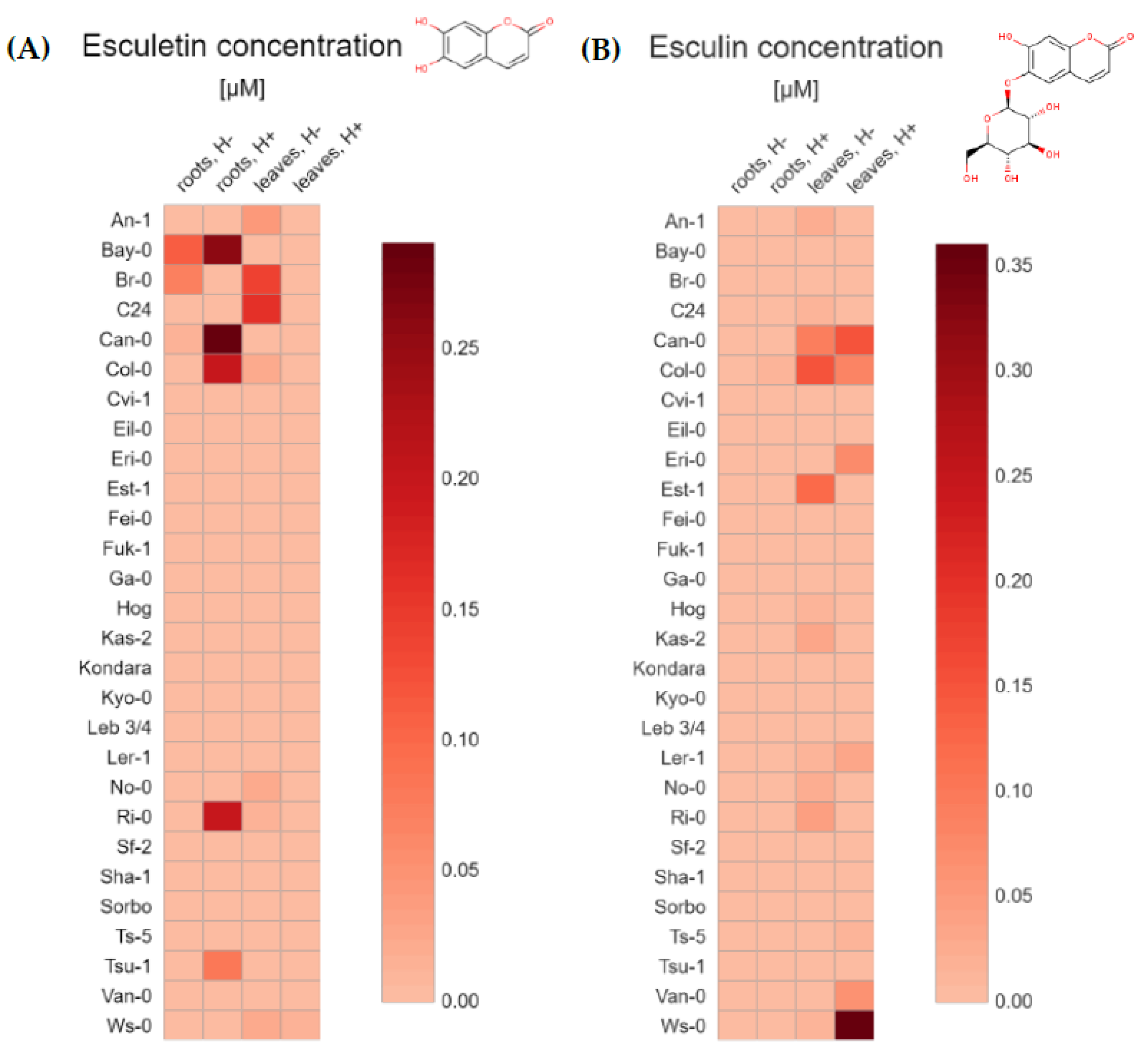
2.1. UHPLC-MS Targeted Metabolite Profiling of Root and Leaf Tissues Reveals Distinct Metabolic Phenotypes for 28 Arabidopsis Accessions
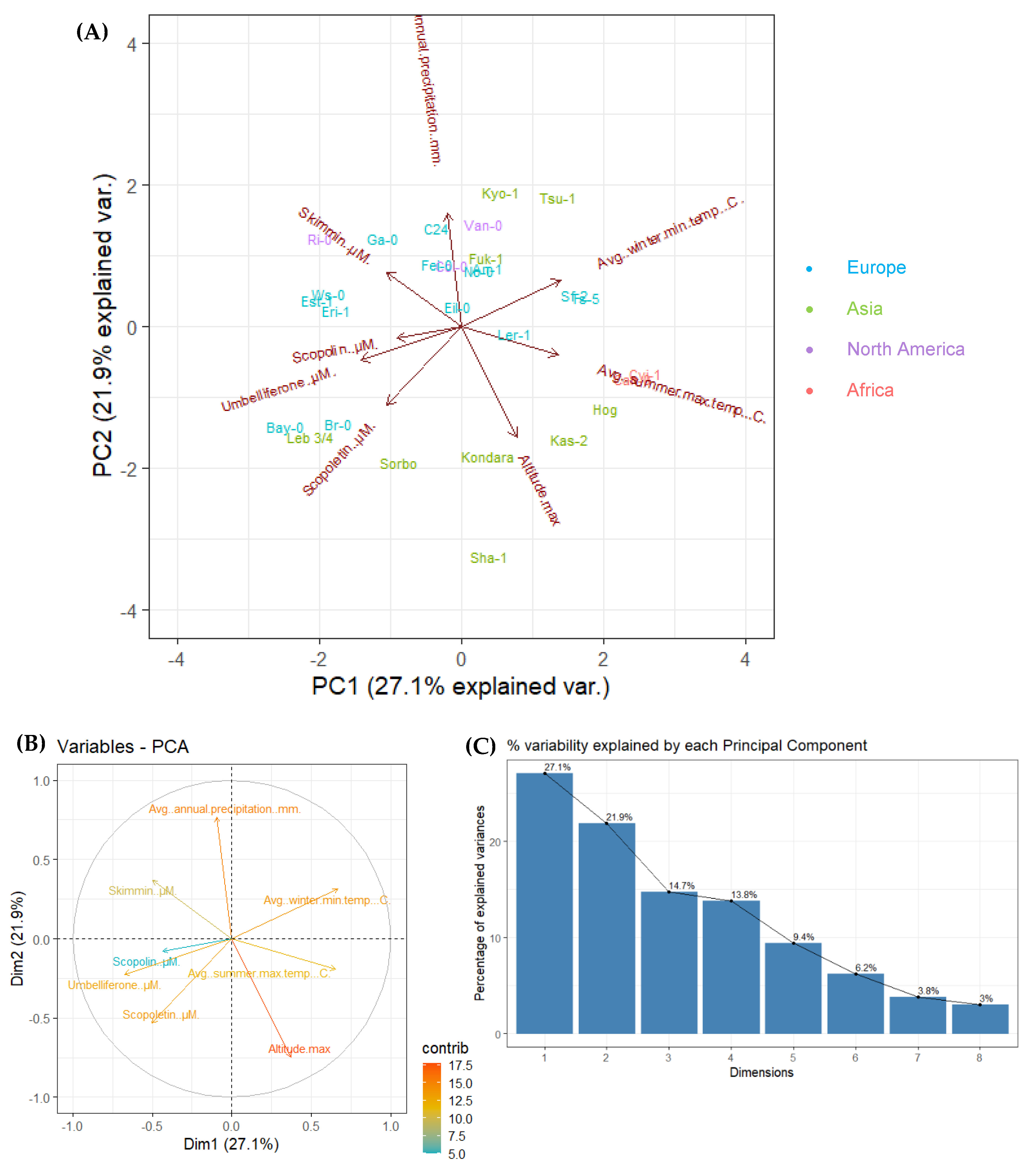
2.2. Principal Component Analysis (PCA) for 28 Arabidopsis Accessions Using Coumarin Quantification by UHPLC-MS in Selected Geographic and under Diverse Climatic Factors
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Plant Material
4.3. In Vitro Plant Culture
4.4. Metabolites Extraction
4.5. Enzymatic Hydrolysis
4.6. UHPLC Separation
4.7. MS Detection
4.8. Peak Identification and Quantitation
4.9. Principal Component Analysis (PCA)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Matos, M.J.; Santana, L.; Uriarte, E.; Abreu, O.A.; Molina, E.; Yordi, E.G. Coumarins—An Important Class of Phytochemicals. Phytochem. Isol. Charact. Role Hum. Health 2015, 113–140. [Google Scholar] [CrossRef] [Green Version]
- Riveiro, M.E.; De Kimpe, N.; Moglioni, A.; Vazquez, R.; Monczor, F.; Shayo, C.; Davio, C. Coumarins: Old Compounds with Novel Promising Therapeutic Perspectives. Curr. Med. Chem. 2010, 17, 1325–1338. [Google Scholar] [CrossRef]
- Rajniak, J.; Giehl, R.F.H.; Chang, E.; Murgia, I.; Von Wirén, N.; Sattely, E.S. Biosynthesis of redox-active metabolites in response to iron deficiency in plants. Nat. Chem. Biol. 2018, 14, 442–450. [Google Scholar] [CrossRef]
- Reen, F.J.; Gutiérrez-Barranquero, J.A.; Parages, M.L.; O´gara, F. Coumarin: A novel player in microbial quorum sensing and biofilm formation inhibition. Appl. Microbiol. Biotechnol. 2018, 102, 2063–2073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robe, K.; Conejero, G.; Gao, F.; Lefebvre-Legendre, L.; Sylvestre-Gonon, E.; Rofidal, V.; Hem, S.; Rouhier, N.; Barberon, M.; Hecker, A.; et al. Coumarin accumulation and trafficking in Arabidopsis thaliana: A complex and dynamic process. New Phytol. 2021, 229, 2062–2079. [Google Scholar] [CrossRef]
- Siwinska, J.; Siatkowska, K.; Olry, A.; Grosjean, J.; Hehn, A.; Bourgaud, F.; A Meharg, A.; Carey, M.; Lojkowska, E.; Ihnatowicz, A. Scopoletin 8-hydroxylase: A novel enzyme involved in coumarin biosynthesis and iron-deficiency responses in Arabidopsis. J. Exp. Bot. 2018, 69, 1735–1748. [Google Scholar] [CrossRef] [Green Version]
- Stassen, M.J.; Hsu, S.-H.; Pieterse, C.M.; Stringlis, I.A. Coumarin Communication Along the Microbiome–Root–Shoot Axis. Trends Plant Sci. 2021, 26, 169–183. [Google Scholar] [CrossRef] [PubMed]
- A Stringlis, I.; De Jonge, R. The Age of Coumarins in Plant–Microbe Interactions. Plant Cell Physiol. 2019, 60, 1405–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegler, J.; Schmidt, S.; Chutia, R.; Müller, J.; Böttcher, C.; Strehmel, N.; Scheel, D.; Abel, S. Non-targeted profiling of semi-polar metabolites in Arabidopsis root exudates uncovers a role for coumarin secretion and lignification during the local response to phosphate limitation. J. Exp. Bot. 2016, 67, 1421–1432. [Google Scholar] [CrossRef] [PubMed]
- Badri, D.V.; Loyola-Vargas, V.M.; Broeckling, C.D.; Vivanco, J.M. Root Secretion of Phytochemicals in Arabidopsis Is Predominantly Not Influenced by Diurnal Rhythms. Mol. Plant 2010, 3, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Chutia, R.; Abel, S.; Ziegler, J. Iron and Phosphate Deficiency Regulators Concertedly Control Coumarin Profiles in Arabidopsis thaliana Roots During Iron, Phosphate, and Combined Deficiencies. Front. Plant Sci. 2019, 10, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fourcroy, P.; Sisó-Terraza, P.; Sudre, D.; Savirón, M.; Reyt, G.; Gaymard, F.; Abadía, A.; Abadia, J.; Álvarez-Fernández, A.; Briat, J.-F. Involvement of the ABCG37 transporter in secretion of scopoletin and derivatives by Arabidopsis roots in response to iron deficiency. New Phytol. 2014, 201, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Hildreth, S.B.; Foley, E.E.; Muday, G.K.; Helm, R.F.; Winkel, B.S.J. The dynamic response of the Arabidopsis root metabolome to auxin and ethylene is not predicted by changes in the transcriptome. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Mönchgesang, S.; Strehmel, N.; Schmidt, S.; Westphal, L.; Taruttis, F.; Müller, E.; Herklotz, S.; Neumann, S.; Scheel, D. Natural variation of root exudates in Arabidopsis thaliana-linking metabolomic and genomic data. Sci. Rep. 2016, 6, 29033. [Google Scholar] [CrossRef]
- Mönchgesang, S.; Strehmel, N.; Trutschel, D.; Westphal, L.; Neumann, S.; Scheel, D. Plant-to-Plant Variability in Root Metabolite Profiles of 19 Arabidopsis thaliana Accessions Is Substance-Class-Dependent. Int. J. Mol. Sci. 2016, 17, 1565. [Google Scholar] [CrossRef] [Green Version]
- Strehmel, N.; Böttcher, C.; Schmidt, S.; Scheel, D. Profiling of secondary metabolites in root exudates of Arabidopsis thaliana. Phytochemistry 2014, 108, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, J.; Schmidt, S.; Strehmel, N.; Scheel, D.; Abel, S. Arabidopsis Transporter ABCG37/PDR9 contributes primarily highly oxygenated Coumarins to Root Exudation. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clemens, S.; Weber, M. The essential role of coumarin secretion for Fe acquisition from alkaline soil. Plant Signal. Behav. 2016, 11, e1114197. [Google Scholar] [CrossRef] [Green Version]
- Mitchell-Olds, T.; Pedersen, D. The molecular basis of quantitative genetic variation in central and secondary metabolism in Arabidopsis. Genetics 1998, 149, 739–747. [Google Scholar] [PubMed]
- Hirai, M.Y.; Yano, M.; Goodenowe, D.B.; Kanaya, S.; Kimura, T.; Awazuhara, M.; Arita, M.; Fujiwara, T.; Saito, K. From the Cover: Integration of transcriptomics and metabolomics for understanding of global responses to nutritional stresses in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2004, 101, 10205–10210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Roepenack-Lahaye, E.; Degenkolb, T.; Zerjeski, M.; Franz, M.; Roth, U.; Wessjohann, L.; Schmidt, J.; Scheel, D.; Clemens, S. Profiling of Arabidopsis Secondary Metabolites by Capillary Liquid Chromatography Coupled to Electrospray Ionization Quadrupole Time-of-Flight Mass Spectrometry. Plant Physiol. 2004, 134, 548–559. [Google Scholar] [CrossRef] [Green Version]
- Keurentjes, J.J.; Fu, J.; De Vos, C.R.; Lommen, A.; Hall, R.D.; Bino, R.J.; Van Der Plas, L.H.; Jansen, R.C.; Vreugdenhil, D.; Koornneef, M. The genetics of plant metabolism. Nat. Genet. 2006, 38, 842–849. [Google Scholar] [CrossRef] [Green Version]
- Fiehn, O. Metabolite Profiling in Arabidopsis. In Arabidopsis Protocols; Springer Science and Business Media LLC: Berlin, Germany, 2006; Volume 323, pp. 439–448. [Google Scholar]
- Ebert, B.; Zöller, D.; Erban, A.; Fehrle, I.; Hartmann, J.; Niehl, A.; Kopka, J.; Fisahn, J. Metabolic profiling of Arabidopsis thaliana epidermal cells. J. Exp. Bot. 2010, 61, 1321–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannah, M.A.; Caldana, C.; Steinhauser, D.; Balbo, I.; Fernie, A.R.; Willmitzer, L. Combined Transcript and Metabolite Profiling of Arabidopsis Grown under Widely Variant Growth Conditions Facilitates the Identification of Novel Metabolite-Mediated Regulation of Gene Expression. Plant Physiol. 2010, 152, 2120–2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Routaboul, J.-M.; Dubos, C.; Beck, G.; Marquis, C.; Bidzinski, P.; Loudet, O.; Lepiniec, L. Metabolite profiling and quantitative genetics of natural variation for flavonoids in Arabidopsis. J. Exp. Bot. 2012, 63, 3749–3764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, M.; Balazadeh, S.; Tohge, T.; Erban, A.; Giavalisco, P.; Kopka, J.; Mueller-Roeber, B.; Fernie, A.R.; Hoefgen, R. Comprehensive Dissection of Spatiotemporal Metabolic Shifts in Primary, Secondary, and Lipid Metabolism during Developmental Senescence in Arabidopsis. Plant Physiol. 2013, 162, 1290–1310. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Alseekh, S.; Cuadros-Inostroza, Á.; Fusari, C.M.; Mutwil, M.; Kooke, R.; Keurentjes, J.B.; Fernie, A.R.; Willmitzer, L.; Brotman, Y. Combined Use of Genome-Wide Association Data and Correlation Networks Unravels Key Regulators of Primary Metabolism in Arabidopsis thaliana. PLoS Genet. 2016, 12, e1006363. [Google Scholar] [CrossRef]
- Orlita, A.; Sidwa-Gorycka, M.; Paszkiewicz, M.; Malinski, E.; Kumirska, J.; Siedlecka, E.M.; Łojkowska, E.; Stepnowski, P. Application of chitin and chitosan as elicitors of coumarins and furoquinolone alkaloids in Ruta graveolens L. (common rue). Biotechnol. Appl. Biochem. 2008, 51, 91–96. [Google Scholar] [CrossRef]
- Sidwa-Gorycka, M.; Krolicka, A.; Orlita, A.; Maliński, E.; Golebiowski, M.; Kumirska, J.; Chromik, A.; Biskup, E.; Stepnowski, P.; Lojkowska, E. Genetic transformation of Ruta graveolens L. by Agrobacterium rhizogenes: Hairy root cultures a promising approach for production of coumarins and furanocoumarins. Plant Cell Tissue Organ Cult. (PCTOC) 2009, 97, 59–69. [Google Scholar] [CrossRef]
- Sidwa-Gorycka, M.; Królicka, A.; Kozyra, M.; Głowniak, K.; Bourgaud, F.; Łojkowska, E. Establishment of a co-culture of Ammi majus L. and Ruta graveolens L. for the synthesis of furanocoumarins. Plant Sci. 2003, 165, 1315–1319. [Google Scholar] [CrossRef]
- Matsumoto, S.; Mizutani, M.; Sakata, K.; Shimizu, B.-I. Molecular cloning and functional analysis of the ortho-hydroxylases of p-coumaroyl coenzyme A/feruloyl coenzyme A involved in formation of umbelliferone and scopoletin in sweet potato, Ipomoea batatas (L.) Lam. Phytochemistry 2012, 74, 49–57. [Google Scholar] [CrossRef]
- Vialart, G.; Hehn, A.; Olry, A.; Ito, K.; Krieger, C.; Larbat, R.; Paris, C.; Shimizu, B.-I.; Sugimoto, Y.; Mizutani, M.; et al. A 2-oxoglutarate-dependent dioxygenase from Ruta graveolens L. exhibits p-coumaroyl CoA 2′-hydroxylase activity (C2′H): A missing step in the synthesis of umbelliferone in plants. Plant J. 2012, 70, 460–470. [Google Scholar] [CrossRef]
- Yang, X.; Wei, S.; Liu, B.; Guo, D.; Zheng, B.; Feng, L.; Liu, Y.; Tomás-Barberán, F.A.; Luo, L.; Huang, D. A novel integrated non-targeted metabolomic analysis reveals significant metabolite variations between different lettuce (Lactuca sativa. L.) varieties. Hortic. Res. 2018, 5, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micallef, S.A.; Shiaris, M.P.; Colón-Carmona, A. Influence of Arabidopsis thaliana accessions on rhizobacterial communities and natural variation in root exudates. J. Exp. Bot. 2009, 60, 1729–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siwinska, J.; Kadzinski, L.; Banasiuk, R.; Gwizdek-Wisniewska, A.; Olry, A.; Banecki, B.; Lojkowska, E.; Ihnatowicz, A. Identification of QTLs affecting scopolin and scopoletin biosynthesis in Arabidopsis thaliana. BMC Plant Biol. 2014, 14, 280. [Google Scholar] [CrossRef] [Green Version]
- Kai, K.; Shimizu, B.-I.; Mizutani, M.; Watanabe, K.; Sakata, K. Accumulation of coumarins in Arabidopsis thaliana. Phytochemistry 2006, 67, 379–386. [Google Scholar] [CrossRef]
- Kai, K.; Mizutani, M.; Kawamura, N.; Yamamoto, R.; Tamai, M.; Yamaguchi, H.; Sakata, K.; Shimizu, B.-I. Scopoletin is biosynthesized via ortho-hydroxylation of feruloyl CoA by a 2-oxoglutarate-dependent dioxygenase in Arabidopsis thaliana. Plant J. 2008, 55, 989–999. [Google Scholar] [CrossRef] [PubMed]
- Schmid, N.B.; Giehl, R.F.; Döll, S.; Mock, H.-P.; Strehmel, N.; Scheel, D.; Kong, X.; Hider, R.C.; Von Wirén, N. Feruloyl-CoA 6’-Hydroxylase1-Dependent Coumarins Mediate Iron Acquisition from Alkaline Substrates in Arabidopsis. Plant Physiol. 2014, 164, 160–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koornneef, M.; Alonso-Blanco, C.; Vreugdenhil, D. Naturally occurring genetic variation in Arabidopsis thaliana. Annu. Rev. Plant Biol. 2004, 55, 141–172. [Google Scholar] [CrossRef] [Green Version]
- Stringlis, I.A.; Yu, K.; Feussner, K.; De Jonge, R.; Van Bentum, S.; Van Verk, M.C.; Berendsen, R.L.; Bakker, P.A.H.M.; Feussner, I.; Pieterse, C.M.J. MYB72-dependent coumarin exudation shapes root microbiome assembly to promote plant health. Proc. Natl. Acad. Sci. USA 2018, 115, E5213–E5222. [Google Scholar] [CrossRef] [Green Version]
- Harbort, C.J.; Hashimoto, M.; Inoue, H.; Niu, Y.; Guan, R.; Rombolà, A.D.; Kopriva, S.; Voges, M.J.; Sattely, E.S.; Garrido-Oter, R.; et al. Root-Secreted Coumarins and the Microbiota Interact to Improve Iron Nutrition in Arabidopsis. Cell Host Microbe 2020, 28, 825–837.e6. [Google Scholar] [CrossRef] [PubMed]
- Voges, M.J.E.E.E.; Bai, Y.; Schulze-Lefert, P.; Sattely, E.S. Plant-derived coumarins shape the composition of an Arabidopsis synthetic root microbiome. Proc. Natl. Acad. Sci. USA 2019, 116, 12558–12565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacoby, R.P.; Koprivova, A.; Kopriva, S. Pinpointing secondary metabolites that shape the composition and function of the plant microbiome. J. Exp. Bot. 2021, 72, 57–69. [Google Scholar] [CrossRef]
- Schmidt, H.; Günther, C.; Weber, M.; Spörlein, C.; Loscher, S.; Böttcher, C.; Schobert, R.; Clemens, S. Metabolome Analysis of Arabidopsis thaliana Roots Identifies a Key Metabolic Pathway for Iron Acquisition. PLoS ONE 2014, 9, e102444. [Google Scholar] [CrossRef] [Green Version]
- Döll, S.; Kuhlmann, M.; Rutten, T.; Mette, M.F.; Scharfenberg, S.; Petridis, A.; Berreth, D.-C.; Mock, H.-P. Accumulation of the coumarin scopolin under abiotic stress conditions is mediated by the Arabidopsis thaliana THO/TREX complex. Plant J. 2018, 93, 431–444. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, C.; Bouque, V.; Bourgaud, F.; Guckert, A. Quantification of Daidzein and Furanocoumarin Conjugates of Psoralea cinerea L. (Leguminosae). Phytochem. Anal. 1997, 8, 27–31. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
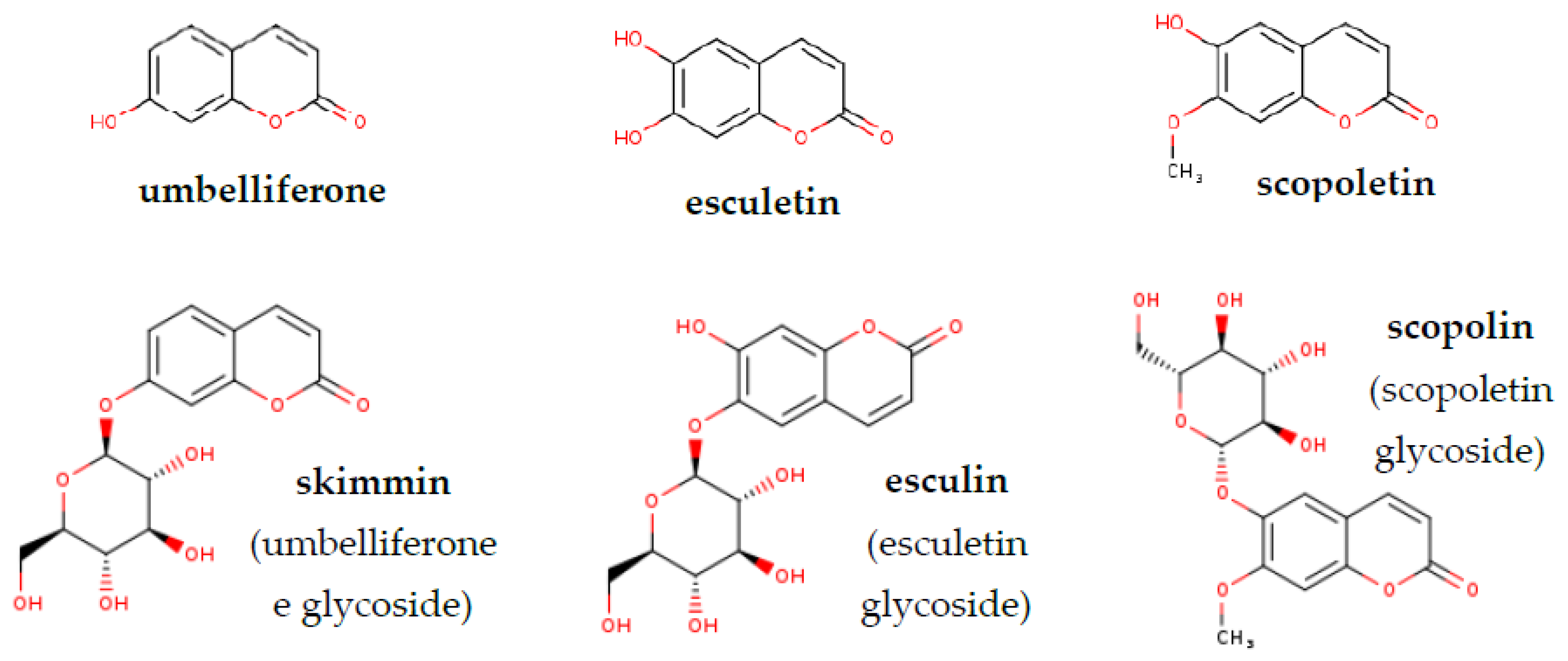
| Peak Number | Retention Time tR (min) | Compound | Mass (m/z Ratio) | LOQ |
|---|---|---|---|---|
| (1) | 14.5 | Umbelliferone | 163 (M+H+) | 0.2 µM |
| (2) | 11.8 | Esculetin | 179 (M+H+) | 0.5 µM |
| (3) | 14.8 | Scopoletin | 193 (M+H+) | 0.2 µM |
| (5) | 10.1 | Skimmin (glycosylated umbelliferone) | 325 (M+H+) | 0.1 µM |
| (5) | 11.8 | Esculin (glycosylated esculetin) | 341 (M+H+) | 0.1 µM |
| (6) | 11 | Scopolin (glycosylated scopoletin) | 355 (M+H+) | 0.1 µM |
| No. | Full Name | Abbreviation | Country of Origin |
|---|---|---|---|
| 1 | Antwerpen | An-1 | Belgium |
| 2 | Bayreuth | Bay-0 | Germany |
| 3 | Brunn | Br-0 | Czech Republic |
| 4 | Coimbra | C24 | Portugal |
| 5 | Canary Islands | Can-0 | Spain |
| 6 | Columbia | Col-0 | USA |
| 7 | Cape Verdi | Cvi-1 | Cape Verde Islands |
| 8 | Eilenburg | Eil-0 | Germany |
| 9 | Eringsboda | Eri-1 | Sweden |
| 10 | Estland | Est-1 | Russia |
| 11 | St. Maria d. Feiria | Fei-0 | Portugal |
| 12 | Fukuyama | Fuk-1 | Japan |
| 13 | Gabelstein | Ga-0 | Germany |
| 14 | Hodja-Obi-Garm | Hog | Tajikistan |
| 15 | Kashmir | Kas-2 | India |
| 16 | Kondara | Kondara | Tajikistan |
| 17 | Kyoto | Kyo-1 | Japan |
| 18 | Lebjasche | Leb 3/4 | Russia |
| 19 | Landsberg erecta | Ler-1 | Germany |
| 20 | Nossen | No-0 | Germany |
| 21 | Richmond | Ri-0 | Canada |
| 22 | San Feliu | Sf-2 | Spain |
| 23 | Shakdara | Sha-1 | Tajikistan |
| 24 | Sorbo | Sorbo | Tajikistan |
| 25 | Tossa del Mar | Ts-5 | Spain |
| 26 | Tsushima | Tsu-1 | Japan |
| 27 | Vancouver | Van-0 | Canada |
| 28 | Wassilewskija | Ws-0 | Belarus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perkowska, I.; Siwinska, J.; Olry, A.; Grosjean, J.; Hehn, A.; Bourgaud, F.; Lojkowska, E.; Ihnatowicz, A. Identification and Quantification of Coumarins by UHPLC-MS in Arabidopsis thaliana Natural Populations. Molecules 2021, 26, 1804. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26061804
Perkowska I, Siwinska J, Olry A, Grosjean J, Hehn A, Bourgaud F, Lojkowska E, Ihnatowicz A. Identification and Quantification of Coumarins by UHPLC-MS in Arabidopsis thaliana Natural Populations. Molecules. 2021; 26(6):1804. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26061804
Chicago/Turabian StylePerkowska, Izabela, Joanna Siwinska, Alexandre Olry, Jérémy Grosjean, Alain Hehn, Frédéric Bourgaud, Ewa Lojkowska, and Anna Ihnatowicz. 2021. "Identification and Quantification of Coumarins by UHPLC-MS in Arabidopsis thaliana Natural Populations" Molecules 26, no. 6: 1804. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26061804