
Chemokine Receptor 5 Antagonism Causes Reduction in Joint Inflammation in a Collagen-Induced Arthritis Mouse Model

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
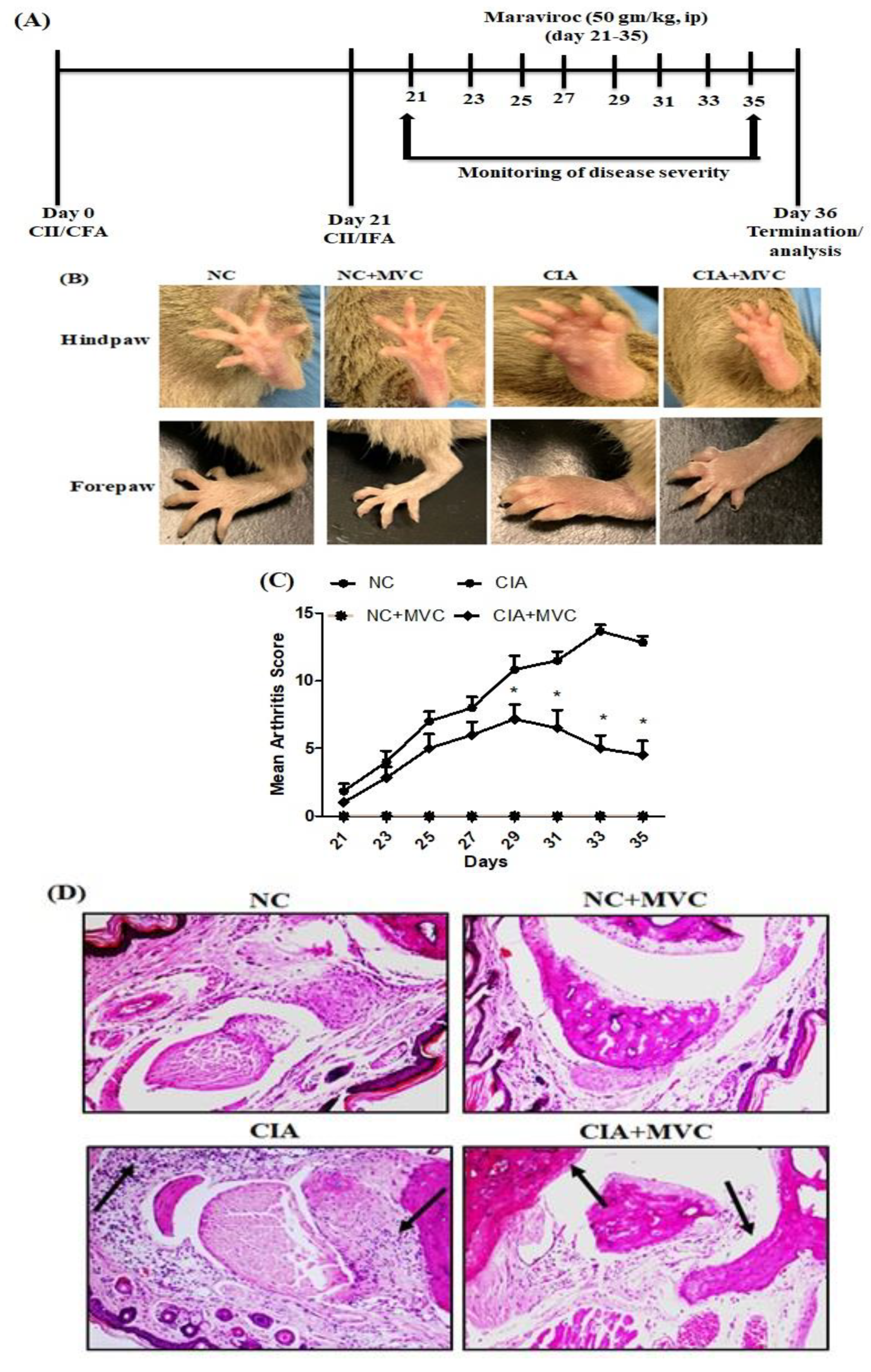
2.1. MVC Exerts Therapeutic Effects in CIA Mice
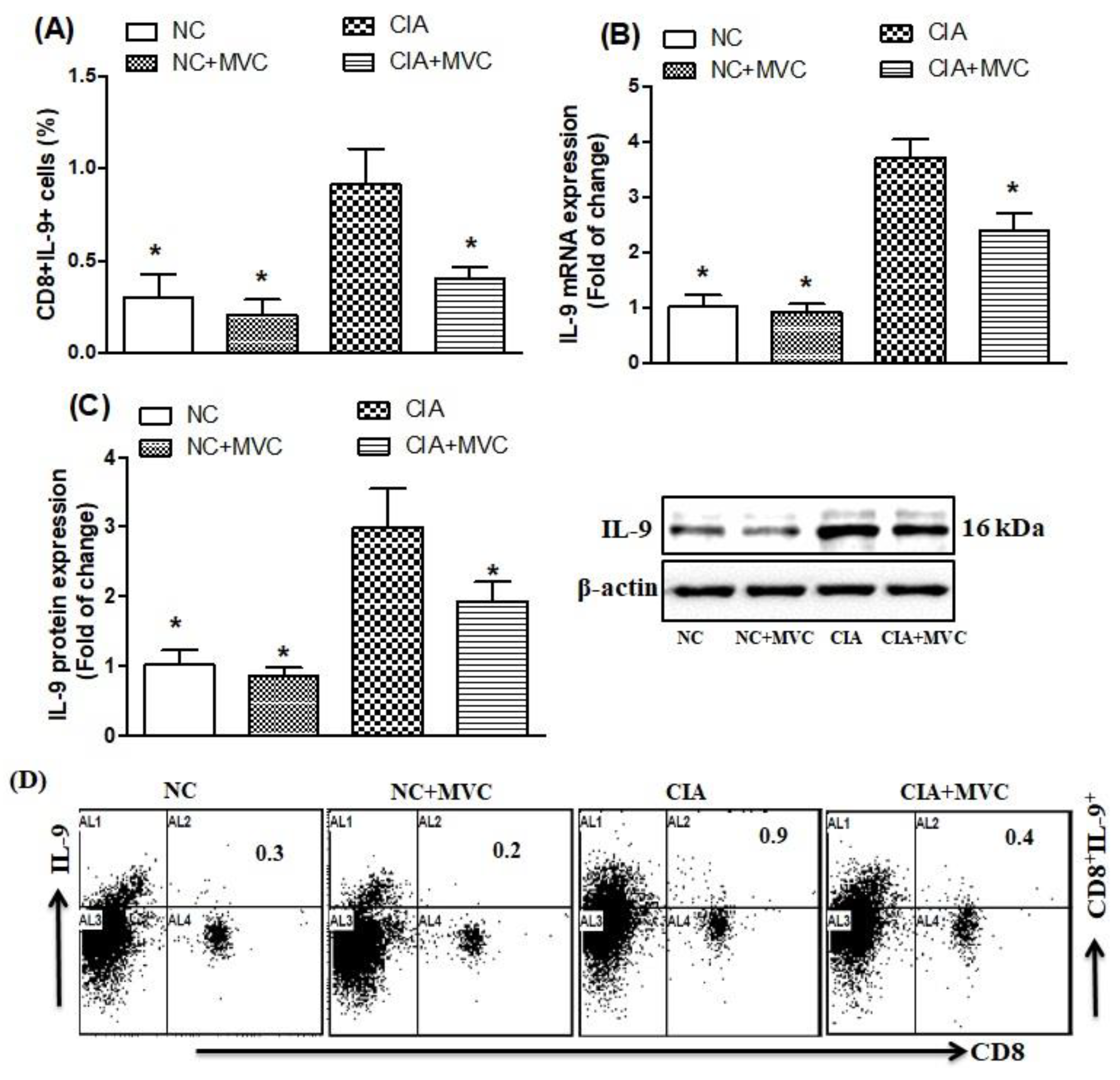
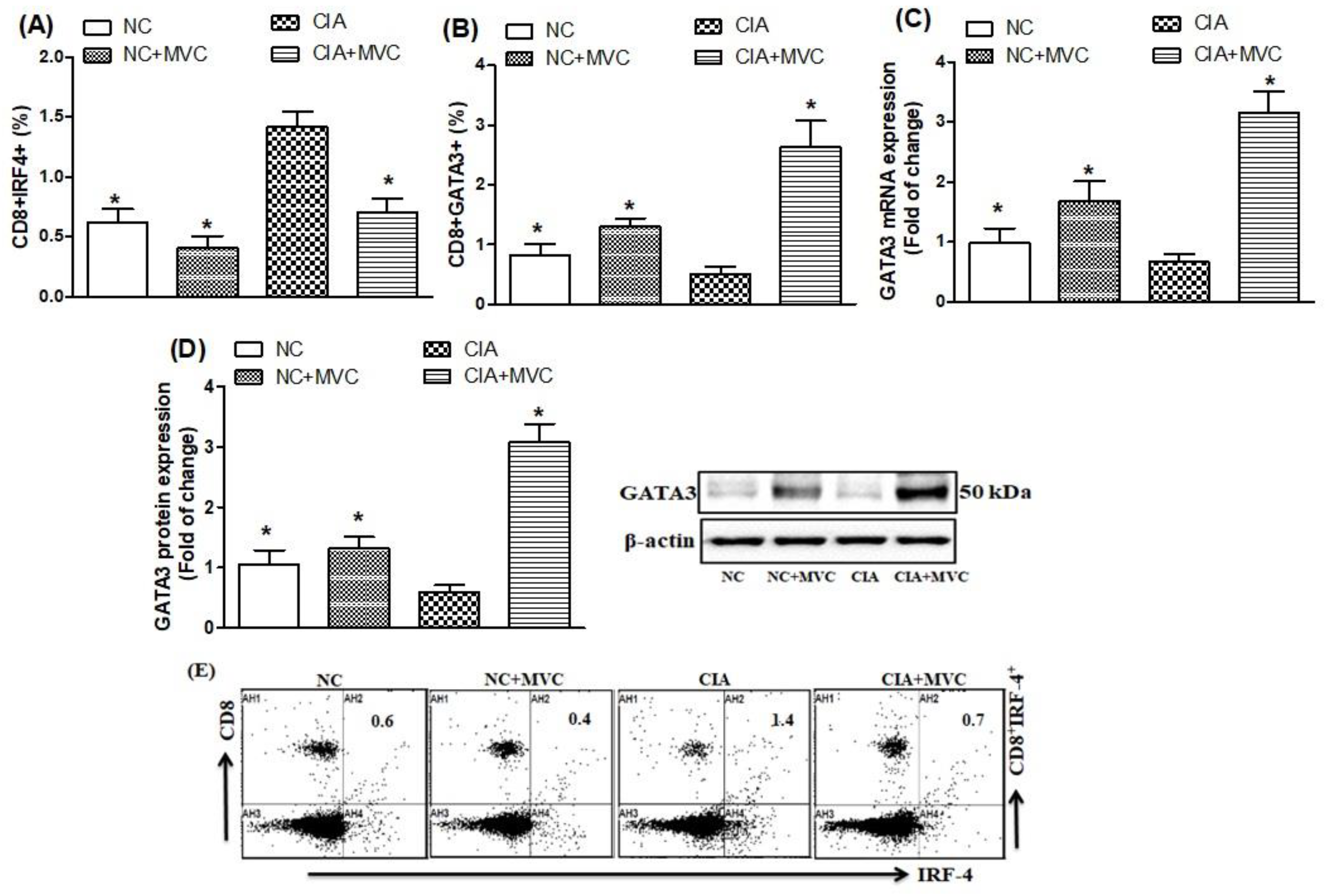
2.2. Effect of MVC on Th9-Related Transcription Factors
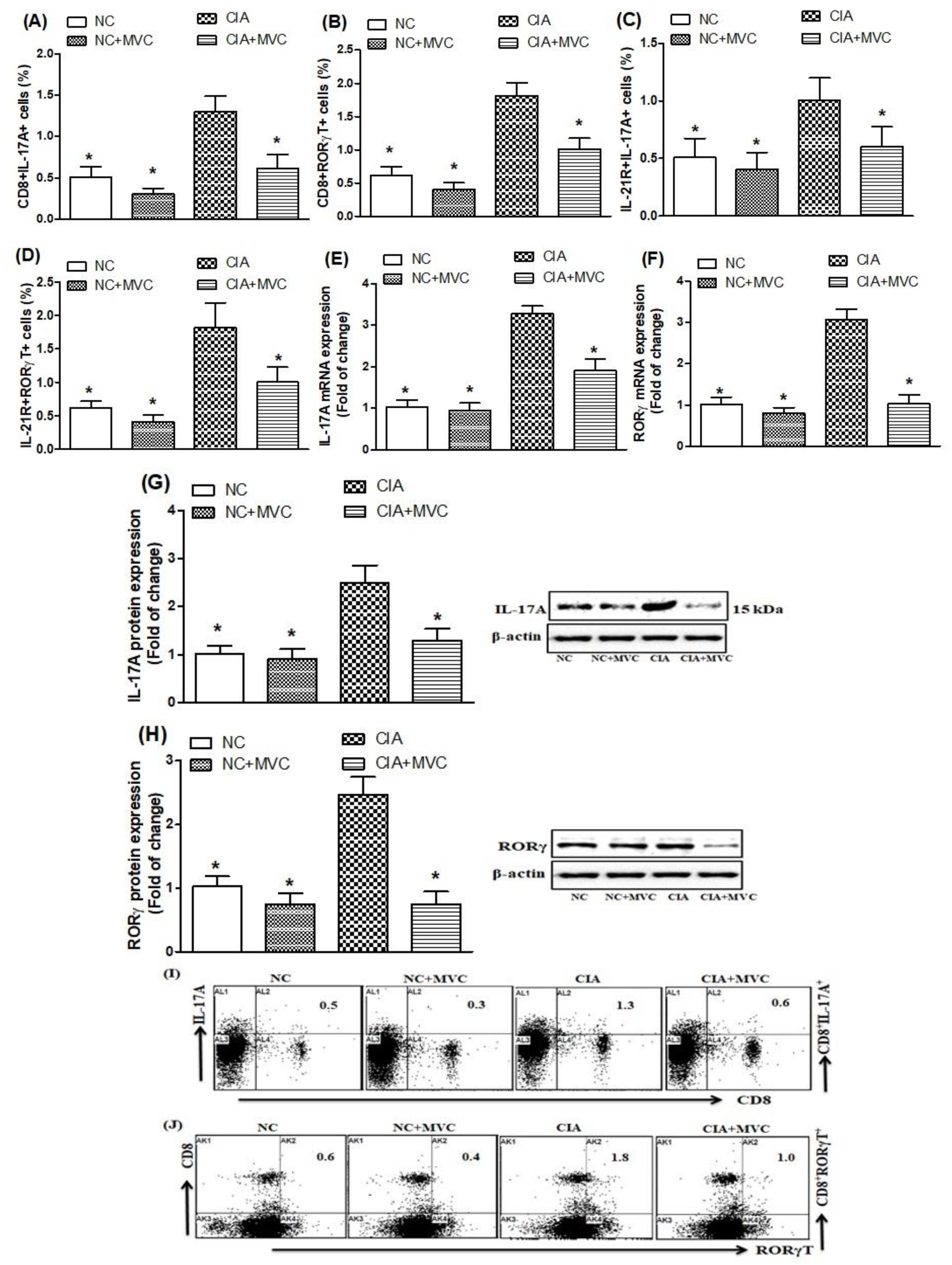
2.3. MVC Treatment Inhibits Th17 Cell Related Signaling
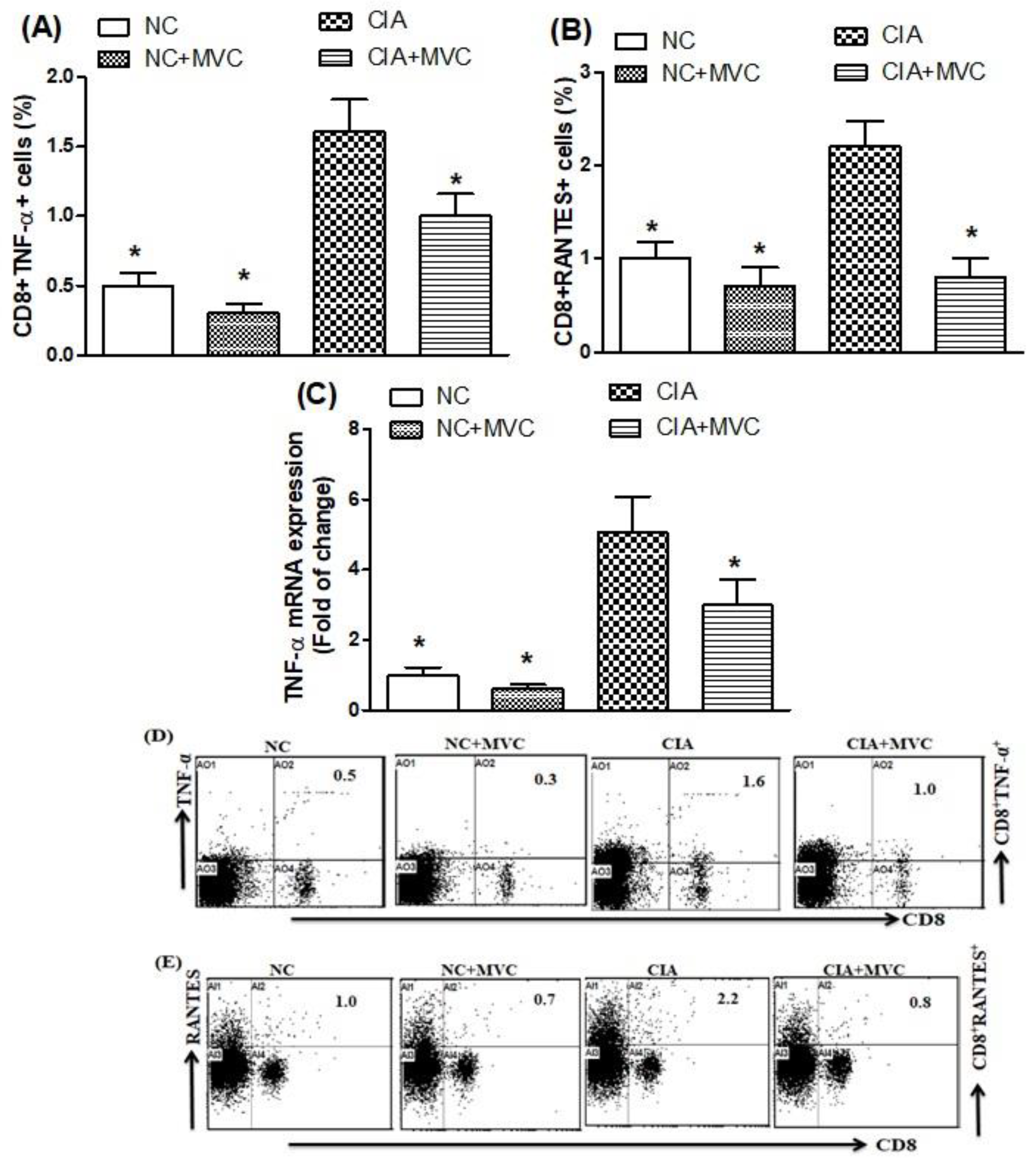
2.4. MVC Treatment Inhibits TNF-α and RANTES Production in CIA Mice
3. Discussion
4. Material Methods
4.1. Animals
4.2. Induction of Experimental CIA and MVC Administration
4.3. Clinical Assessment of Arthritis
4.4. Histological Assessment
4.5. Flow Cytometric Analysis
4.6. Real-Time RT-PCR
4.7. Western Blot Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Iwamoto, T.; Okamoto, H.; Toyama, Y.; Momohara, S. Molecular aspects of rheumatoid arthritis: Chemokines in the joints of pa-tients. FEBS J. 2008, 275, 4448–4455. [Google Scholar] [CrossRef]
- Firestein, G.S. Evolving concepts of rheumatoid arthritis. Nat. Cell Biol. 2003, 423, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Scott, D.L.; Wolfe, F.; Huizinga, T.W. Rheumatoid arthritis. Lancet 2010, 376, 1094–1108. [Google Scholar] [CrossRef]
- Park, J.S.; Moon, S.J.; Lim, M.; Byun, J.K.; Hwang, S.H.; Yang, S.; Kim, E.K.; Kim, S.-M.; Lee, J.; Kwok, S.; et al. Retinoic Acid Receptor-Related Receptor Alpha Ameliorates Autoimmune Arthritis via Inhibiting of Th17 Cells and Osteoclastogenesis. Front. Immunol. 2019, 10, 2270. [Google Scholar] [CrossRef] [PubMed]
- Angelotti, F.; Parma, A.; Cafaro, G.; Capecchi, R.; Alunno, A.; Puxeddu, I. One year in review 2017: Pathogenesis of rheumatoid arthritis. Clin. Exp. Rheumatol. 2017, 35, 368–378. [Google Scholar] [PubMed]
- Deng, Y.; Wang, Z.; Chang, C.; Lu, L.; Lau, C.S.; Lu, Q. Th9 cells and IL-9 in autoimmune disorders: Pathogenesis and therapeutic potentials. Hum. Immunol. 2017, 78, 120–128. [Google Scholar] [CrossRef]
- Yanaba, K.; Yoshizaki, A.; Asano, Y.; Kadono, T.; Sato, S. Serum Interleukin 9 Levels Are Increased in Patients with Systemic Sclerosis: Association with Lower Frequency and Severity of Pulmonary Fibrosis. J. Rheumatol. 2011, 38, 2193–2197. [Google Scholar] [CrossRef]
- Ciccia, F.; Guggino, G.; Rizzo, A.; Manzo, A.; Vitolo, B.; La Manna, M.P.; Giardina, G.; Sireci, G.; Dieli, F.; Montecucco, C.M.; et al. Potential involvement of IL-9 and Th9 cells in the pathogenesis of rheumatoid arthritis. Rheumatol. 2015, 54, 2264–2272. [Google Scholar] [CrossRef] [Green Version]
- Kundu-Raychaudhuri, S.; Abria, C.; Raychaudhuri, S.P. IL-9, a local growth factor for synovial T cells in inflammatory arthritis. Cytokine 2016, 79, 45–51. [Google Scholar] [CrossRef]
- Biswas, P.S.; Gupta, S.; Chang, E.; Song, L.; Stirzaker, R.A.; Liao, J.K.; Bhagat, G.; Pernis, A.B. Phosphorylation of IRF4 by ROCK2 regulates IL-17 and IL-21 production and the development of autoimmunity in mice. J. Clin. Investig. 2010, 120, 3280–3295. [Google Scholar] [CrossRef] [Green Version]
- van Hamburg, J.P.; Mus, A.M.; de Bruijn, M.J.; de Vogel, L. GATA-3 protects against severe joint inflammation and bone erosion and reduces differentiation of Th17 cells during experimental arthritis. Arthritis Rheum. 2009, 60, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Hueber, A.J.; Asquith, D.L.; Miller, A.M.; Reilly, J.H.; Kerr, S.; Leipe, J.; Melendez, A.J.; McInnes, I.B. Cutting Edge: Mast Cells Express IL-17A in Rheumatoid Arthritis Synovium. J. Immunol. 2010, 184, 3336–3340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, H.; Goodall, J.C.; Gaston, J.S.H. Frequency and phenotype of peripheral blood Th17 cells in ankylosing spondylitis and rheumatoid arthritis. Arthritis Rheum. 2009, 60, 1647–1656. [Google Scholar] [CrossRef] [PubMed]
- Kotake, S.; Udagawa, N.; Takahashi, N.; Matsuzaki, K.; Itoh, K.; Ishiyama, S.; Saito, S.; Inoue, K.; Kamatani, N.; Gillespie, M.T.; et al. IL-17 in synovial fluids from patients with rheumatoid arthritis is a potent stimulator of osteoclastogenesis. J. Clin. Investig. 1999, 103, 1345–1352. [Google Scholar] [CrossRef]
- Ivanov, I.I.; McKenzie, B.S.; Zhou, L.; Tadokoro, C.E. The orphan nuclear receptor RORgammat directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell 2006, 126, 1121–1133. [Google Scholar] [CrossRef] [Green Version]
- Young, D.A.; Hegen, M.; Ma, H.L.M.; Whitters, M.J.; Albert, L.M.; Lowe, L.; Senices, M.; Wu, P.W.; Sibley, B.; Leathurby, Y.; et al. Blockade of the interleukin-21/interleukin-21 receptor pathway ameliorates disease in animal models of rheumatoid arthritis. Arthritis Rheum. 2007, 56, 1152–1163. [Google Scholar] [CrossRef]
- Mu¨ssener, Å.; Litton, M.J.; Lindroos, E.; Klareskog, L. Cytokine production in synovial tissue of mice with collagen-induced arthritis (CIA). Clin. Exp. Immunol. 1997, 107, 485–493. [Google Scholar] [CrossRef]
- Patel, D.D.; Zachariah, J.P.; Whichard, L.P. CXCR3 and CCR5 Ligands in Rheumatoid Arthritis Synovium. Clin. Immunol. 2001, 98, 39–45. [Google Scholar] [CrossRef]
- Lagane, B.; García-Pérez, J.; Kellenberger, E. Modeling the allosteric modulation of CCR5 function by Maraviroc. Drug Discov. Today Technol. 2013, 10, e297–e305. [Google Scholar] [CrossRef]
- Tan, Q.; Zhu, Y.; Li, J.; Chen, Z.; Han, G.W.; Kufareva, I.; Li, T.; Ma, L.; Fenalti, G.; Zhang, W.; et al. Structure of the CCR5 Chemokine Receptor-HIV Entry Inhibitor Maraviroc Complex. Science 2013, 341, 1387–1390. [Google Scholar] [CrossRef] [Green Version]
- Ermocida, A.; Mengoni, F.; Sauzullo, I.; Rossi, R. Extravirologic modulation of immune response by an NRTI-sparingantiretroviral regimen including darunavir and maraviroc. New Microbiol. 2014, 37, 225–229. [Google Scholar] [PubMed]
- Reshef, R.; Luger, S.M.; Hexner, E.O.; Loren, A.W.; Frey, N.V.; Nasta, S.D.; Goldstein, S.C.; Stadtmauer, E.A.; Smith, J.; Bailey, S.; et al. Blockade of Lymphocyte Chemotaxis in Visceral Graft-versus-Host Disease. New Engl. J. Med. 2012, 367, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Tang, B.; Ren, H.; Liu, H.; Shi, Y.; Dong, Y.; Yin, Y.; Miao, S. CCR5 blockade combined with cyclosporine A attenuates liver GVHD by impairing T cells function. Inflamm. Res. 2016, 65, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Mencarelli, A.; Cipriani, S.; Francisci, D.; Santucci, L.; Baldelli, F.; Distrutti, E.; Fiorucci, S. Highly specific blockade of CCR5 inhibits leukocyte trafficking and reduces mucosal inflammation in murine colitis. Sci. Rep. 2016, 6, 30802. [Google Scholar] [CrossRef] [Green Version]
- Kiely, P.D.; O’Brien, D.; Oliveira, D.B. Anti-CD8 treatment reduces the severity of inflammatory arthritis, but not vasculitis, in mercuric chloride-induced autoimmunity. Clin. Exp. Immunol. 1996, 106, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, E.; Tomita, T.; Toyosaki-Maeda, T.; Kaneko, M.; Takano, H.; Hashimoto, H.; Sugamoto, K.; Suzuki, R.; Ochi, T. Establishment and characterization of nurse cell-like stromal cell lines from synovial tissues of patients with rheumatoid arthritis. Arthritis Rheum. 1999, 42, 221–228. [Google Scholar] [CrossRef]
- Berner, B.; Akça, D.; Jung, T.; Muller, G.A.; Reuss-Borst, M.A. Analysis of Th1 and Th2 cytokines expressing CD4+ and CD8+ T cells in rheumatoid arthritis by flow cytometry. J. Rheumatol. 2000, 27, 1128–1135. [Google Scholar]
- Zapico, I.; Coto, E.; Rodriguez, A.; Alvarez, C.; Torre, J.C.; Álvarez, V. CCR5 (chemokine receptor-5) DNA-polymorphism influences the severity of rheumatoid arthritis. Genes Immun. 2000, 1, 288–289. [Google Scholar] [CrossRef] [Green Version]
- Koch, A.E.; Kunkel, S.L.; Harlow, L.A.; Mazarakis, D.D.; Haines, G.K.; Burdick, M.D.; Pope, R.M.; Strieter, R.M. Macrophage inflammatory protein-1 alpha. A novel chemotactic cytokine for macrophages in rheumatoid arthritis. J. Clin. Investig. 1994, 93, 921–928. [Google Scholar] [CrossRef]
- Norii, M.; Yamamura, M.; Iwahashi, M.; Ueno, A.; Yamana, J.; Makino, H. Selective recruitment of CXCR3+ and CCR5+ CCR4+ T cells into synovial tissue in patients with rheumatoid arthritis. Acta Med. Okayama 2006, 60, 149–157. [Google Scholar]
- Chowdhury, K.; Kumar, U.; Das, S.; Chaudhuri, J.; Kumar, P.; Kanjilal, M.; Ghosh, P.; Sircar, G.; Basyal, R.K.; Kanga, U.; et al. Synovial IL-9 facilitates neutrophil survival, function and differentiation of Th17 cells in rheumatoid arthritis. Arthritis Res. 2018, 20, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Jäger, A.; Dardalhon, V.; Sobel, R.A.; Bettelli, E.; Kuchroo, V.K. Th1, Th17, and Th9 Effector Cells Induce Experimental Autoimmune Encephalomyelitis with Different Pathological Phenotypes. J. Immunol. 2009, 183, 7169–7177. [Google Scholar] [CrossRef]
- Nowak, E.C.; Weaver, C.T.; Turner, H.; Begum-Haque, S.; Becher, B.; Schreiner, B.; Coyle, A.J.; Kasper, L.H.; Noelle, R.J. IL-9 as a mediator of Th17-driven inflammatory disease. J. Exp. Med. 2009, 206, 1653–1660. [Google Scholar] [CrossRef] [Green Version]
- Cole, S.; Walsh, A.; Yin, X.; Wechalekar, M.D.; Smith, M.D.; Proudman, S.M.; Veale, D.J.; Fearon, U.; Pitzalis, C.; Humby, F.; et al. Integrative analysis reveals CD38 as a therapeutic target for plasma cell-rich pre-disease and established rheumatoid arthritis and systemic lupus erythematosus. Arthritis Res. 2018, 20, 1–14. [Google Scholar] [CrossRef]
- Zhu, W.; Li, X.; Fang, S.; Zhang, X.; Wang, Y.; Zhang, T.; Li, Z.; Xu, Y.; Qu, S.; Liu, C.; et al. Anti-Citrullinated Protein Antibodies Induce Macrophage Subset Disequilibrium in RA Patients. Inflammation 2015, 38, 2067–2075. [Google Scholar] [CrossRef]
- Indrevær, R.L.; Moskaug, J.Ø.; Paur, I.; Bøhn, S.K.; Jørgensen, S.F.; Blomhoff, R.; Aukrust, P.; Fevang, B.; Blomhoff, H.K. IRF4 Is a Critical Gene in Retinoic Acid–Mediated Plasma Cell Formation and Is Deregulated in Common Variable Immunodeficiency–Derived B Cells. J. Immunol. 2015, 195, 2601–2611. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Liu, H.; Wang, Y.; Chen, S.; Liu, J.; Li, W.; Dou, H.; Hou, W.; Meng, M. Study of the adoptive immunotherapy on rheumatoid arthritis with Thymus-derived invariant natural killer T cells. Int. Immunopharmacol. 2019, 67, 427–440. [Google Scholar] [CrossRef] [PubMed]
- Rosu, A.; Margaritescu, C.; Stepan, A.; Musetescu, A.; Ene, M. IL-17 patterns in synovium, serum and synovial fluid from treat-ment-naive, early rheumatoid arthritis patients. Rom. J. Morphol. Embryol. 2012, 53, 73–80. [Google Scholar] [PubMed]
- Nakae, S.; Nambu, A.; Sudo, K.; Iwakura, Y. Suppression of Immune Induction of Collagen-Induced Arthritis in IL-17-Deficient Mice. J. Immunol. 2003, 171, 6173–6177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katz, Y.; Nadiv, O.; Beer, Y. Interleukin-17 enhances tumor necrosis factor alpha-induced synthesis of interleukins 1,6, and 8 in skin and synovial fibroblasts: A possible role as a "fine-tuning cytokine" in inflammation processes. Arthritis Rheum. 2001, 44, 2176–2184. [Google Scholar] [CrossRef]
- Gaffen, S.L. The role of interleukin-17 in the pathogenesis of rheumatoid arthritis. Curr. Rheumatol. Rep. 2009, 11, 365–370. [Google Scholar] [CrossRef]
- Kondo, Y.; Yao, Z.; Tahara, M.; Iizuka, M.; Yokosawa, M.; Kaneko, S.; Segawa, S.; Tsuboi, H.; Yoh, K.; Takahashi, S.; et al. Involvement of RORγt-overexpressing T cells in the development of autoimmune arthritis in mice. Arthritis Res. Ther. 2015, 17, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jüngel, A.; Distler, J.H.W.; Kurowska-Stolarska, M.; Seemayer, C.A.; Seibl, R.; Forster, A.; Michel, B.A.; Gay, R.E.; Emmrich, F.; Gay, S.; et al. Expression of interleukin-21 receptor, but not interleukin-21, in synovial fibroblasts and synovial macrophages of patients with rheumatoid arthritis. Arthritis Rheum. 2004, 50, 1468–1476. [Google Scholar] [CrossRef] [PubMed]
- Marijnissen, R.J.; Roeleveld, D.M.; Young, D.; Nickerson-Nutter, C.; Abdollahi-Roodsaz, S.; De Aquino, S.G.; Van De Loo, F.A.J.; Van Spriel, A.B.; Boots, A.M.H.; Berg, W.B.V.D.; et al. Interleukin-21 Receptor Deficiency Increases the Initial Toll-like Receptor 2 Response but Protects Against Joint Pathology by Reducing Th1 and Th17 Cells During Streptococcal Cell Wall Arthritis. Arthritis Rheumatol. 2013, 66, 886–895. [Google Scholar] [CrossRef] [PubMed]
- Parrish-Novak, J.; Dillon, S.R.; Nelson, A.; Hammond, A. Interleukin 21 and its receptor are involved in NK cell expansion and regulation of lymphocyte function. Nature 2000, 408, 57–63. [Google Scholar] [CrossRef]
- Osta, B.; Roux, J.P.; Lavocat, F.; Pierre, M. Differential Effects of IL-17A and TNF-α on Osteoblastic Differentiation of Isolated Synoviocytes and on Bone Explants from Arthritis Patients. Front Immunol. 2015, 6, 151. [Google Scholar] [CrossRef]
- Jing, W.; Sun, W.; Zhang, N.; Zhao, C.; Yan, X. The protective effects of the GPR39 agonist TC-G 1008 against TNF-α-induced inflammation in human fibroblast-like synoviocytes (FLSs). Eur. J. Pharmacol. 2019, 865, 172663. [Google Scholar] [CrossRef]
- Qadir, M.M.F.; Bhatti, A.; Ashraf, M.U.; Sandhu, M.A.; Anjum, S.; John, P. Immunomodulatory and therapeutic role of Cinnamomum verum extracts in collagen-induced arthritic BALB/c mice. Inflammopharmacology 2018, 26, 157–170. [Google Scholar] [CrossRef]
- Meroni, P.-L.; Valesini, G. Tumour Necrosis Factor α Antagonists in the Treatment of Rheumatoid Arthritis: An Immunological Perspective. BioDrugs 2014, 28, 5–13. [Google Scholar] [CrossRef]
- Agere, S.A.; Akhtar, N.; Watson, J.M.; Ahmed, S. RANTES/CCL5 Induces Collagen Degradation by Activating MMP-1 and MMP-13 Expression in Human Rheumatoid Arthritis Synovial Fibroblasts. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Ellingsen, T.; Buus, A.; Møller, B.K.; Stengaard-Pedersen, K. In vitro migration of mononuclear cells towards synovial fluid and plasma from rheumatoid arthritis patients correlates to RANTES synovial fluid levels and to clinical pain parameters. Scand. J. Rheumatol. 2000, 29, 216–221. [Google Scholar] [PubMed]
- Fleishaker, D.L.; Garcia Meijide, J.A.; Petrov, A.; Kohen, M.D. Maraviroc, a chemokine receptor-5 antagonist, fails to demon-strate efficacy in the treatment of patients with rheumatoid arthritis in a randomized, double-blind placebo-controlled trial. Arthritis Res Ther. 2012, 14, R11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arberas, H.; Guardo, A.C.; Bargalló, M.E.; Maleno, M.J.; Calvo, M.; Blanco, J.L. In vitro effects of the CCR5 inhibitor maraviroc on human T cell function. J. Antimicrob. Chemother. 2013, 68, 577–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanmogne, G.D.; Woollard, S.M. Maraviroc: A review of its use in HIV infection and beyond. Drug Des. Dev. Ther. 2015, 9, 5447–5468. [Google Scholar] [CrossRef] [Green Version]
- Cipriani, S.; Francisci, D.; Mencarelli, A.; Renga, B.; Schiaroli, E.; D’Amore, C.; Baldelli, F.; Fiorucci, S. Efficacy of the CCR5 Antagonist Maraviroc in Reducing Early, Ritonavir-Induced Atherogenesis and Advanced Plaque Progression in Mice. Circulation 2013, 127, 2114–2124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karampoor, S.; Zahednasab, H.; Amini, R.; Esghaei, M.; Sholeh, M.; Keyvani, H. Maraviroc attenuates the pathogenesis of ex-perimental autoimmune encephalitis. Int. Immunopharmacol. 2020, 80, 106138. [Google Scholar] [CrossRef]
- Lo, C.K.C.; Lam, Q.L.K.; Sun, L.; Wang, S.; Ko, K.-H.; Xu, H.; Wu, C.-Y.; Zheng, B.-J.; Lu, L. Natural killer cell degeneration exacerbates experimental arthritis in mice via enhanced interleukin-17 production. Arthritis Rheum. 2008, 58, 2700–2711. [Google Scholar] [CrossRef] [Green Version]
- Bakheet, S.A.; Ansari, M.A.; Nadeem, A.; Attia, S.M.; Alhoshani, A.R.; Gul, G.; Al-Qahtani, Q.; AlBekairi, N.A.; Ibrahim, K.E.; Ahmad, S.F. CXCR3 antagonist AMG487 suppresses rheumatoid arthritis pathogenesis and progression by shifting the Th17/Treg cell balance. Cell. Signal. 2019, 64, 109395. [Google Scholar] [CrossRef]
- Ahmad, S.F.; Ansari, M.A.; Nadeem, A.; Zoheir, K.M.; Bakheet, S.A.; Al-Shabanah, O.A.; Al Rikabi, A.C.; Attia, S.M. The tyrosine kinase inhibitor tyrphostin AG126 reduces activation of inflammatory cells and increases Foxp3+ regulatory T cells during pathogenesis of rheumatoid arthritis. Mol. Immunol. 2016, 78, 65–78. [Google Scholar] [CrossRef]
- Abd-Allah, A.R.A.; Ahmad, S.F.; AlRashidi, I.; Abdel-Hamied, H.E.; Zoheir, K.M.A.; Ashour, A.E.; Bakheet, S.A.; Attia, S.M. Involvement of histamine 4 receptor in the pathogenesis and progression of rheumatoid arthritis. Int. Immunol. 2014, 26, 325–340. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Lee, S.; Baek, S.; Lee, C.; Lee, W.; Rhim, B.; Hong, K.; Kim, C. Suppression of PU.1-linked TLR4 expression by cilostazol with decrease of cytokine production in macrophages from patients with rheumatoid arthritis. Br. J. Pharmacol. 2012, 168, 1401–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability:Samples of the compounds for this experiment are available upon request by contact with the corresponding author. |





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ansari, M.A.; Nadeem, A.; Bakheet, S.A.; Attia, S.M.; Shahid, M.; Alyousef, F.S.; Alswailem, M.A.; Alqinyah, M.; Ahmad, S.F. Chemokine Receptor 5 Antagonism Causes Reduction in Joint Inflammation in a Collagen-Induced Arthritis Mouse Model. Molecules 2021, 26, 1839. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26071839
Ansari MA, Nadeem A, Bakheet SA, Attia SM, Shahid M, Alyousef FS, Alswailem MA, Alqinyah M, Ahmad SF. Chemokine Receptor 5 Antagonism Causes Reduction in Joint Inflammation in a Collagen-Induced Arthritis Mouse Model. Molecules. 2021; 26(7):1839. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26071839
Chicago/Turabian StyleAnsari, Mushtaq A., Ahmed Nadeem, Saleh A. Bakheet, Sabry M. Attia, Mudassar Shahid, Faris S. Alyousef, Mohammed A. Alswailem, Mohammed Alqinyah, and Sheikh F. Ahmad. 2021. "Chemokine Receptor 5 Antagonism Causes Reduction in Joint Inflammation in a Collagen-Induced Arthritis Mouse Model" Molecules 26, no. 7: 1839. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26071839