
Broussochalcone A Is a Novel Inhibitor of the Orphan Nuclear Receptor NR4A1 and Induces Apoptosis in Pancreatic Cancer Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
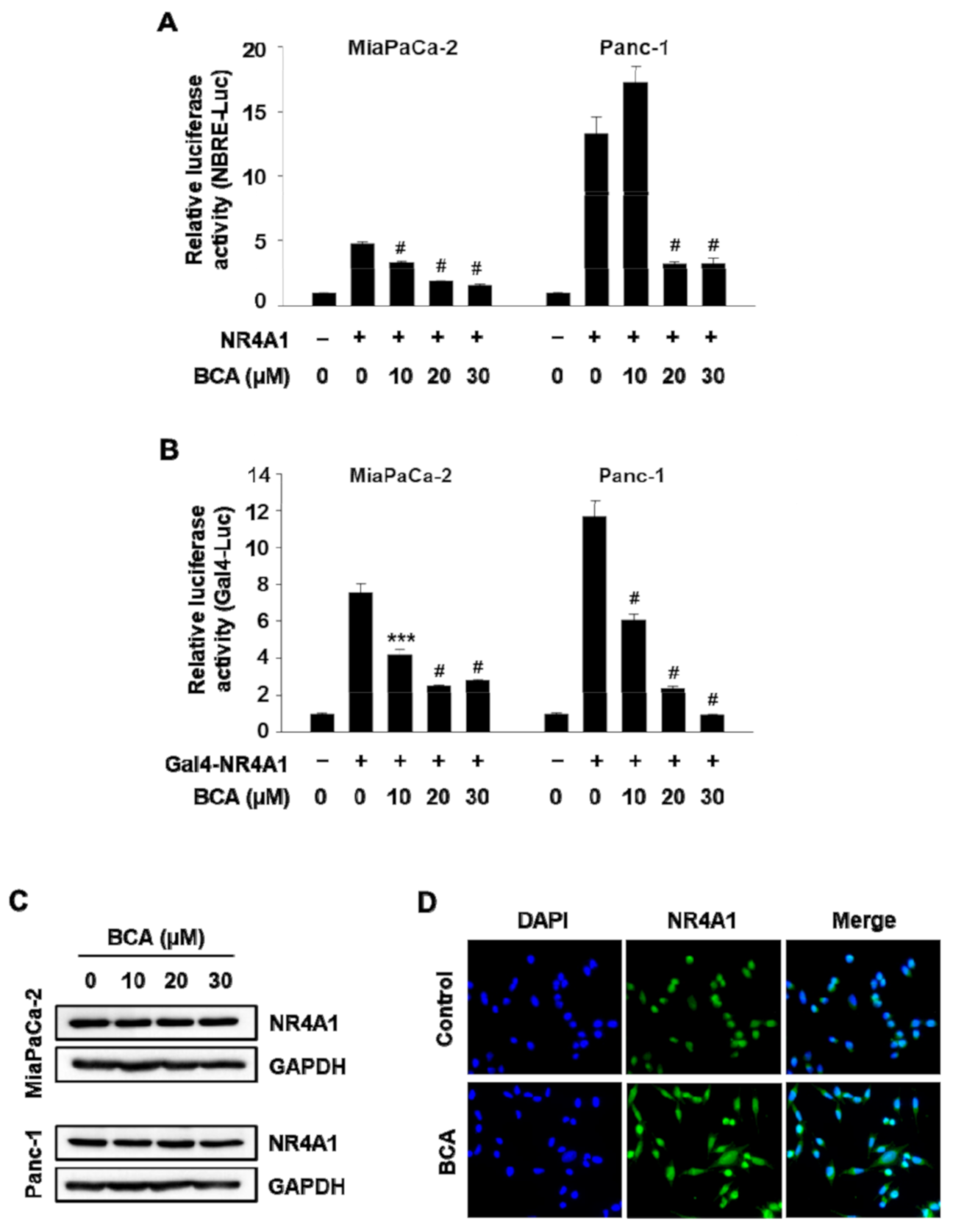
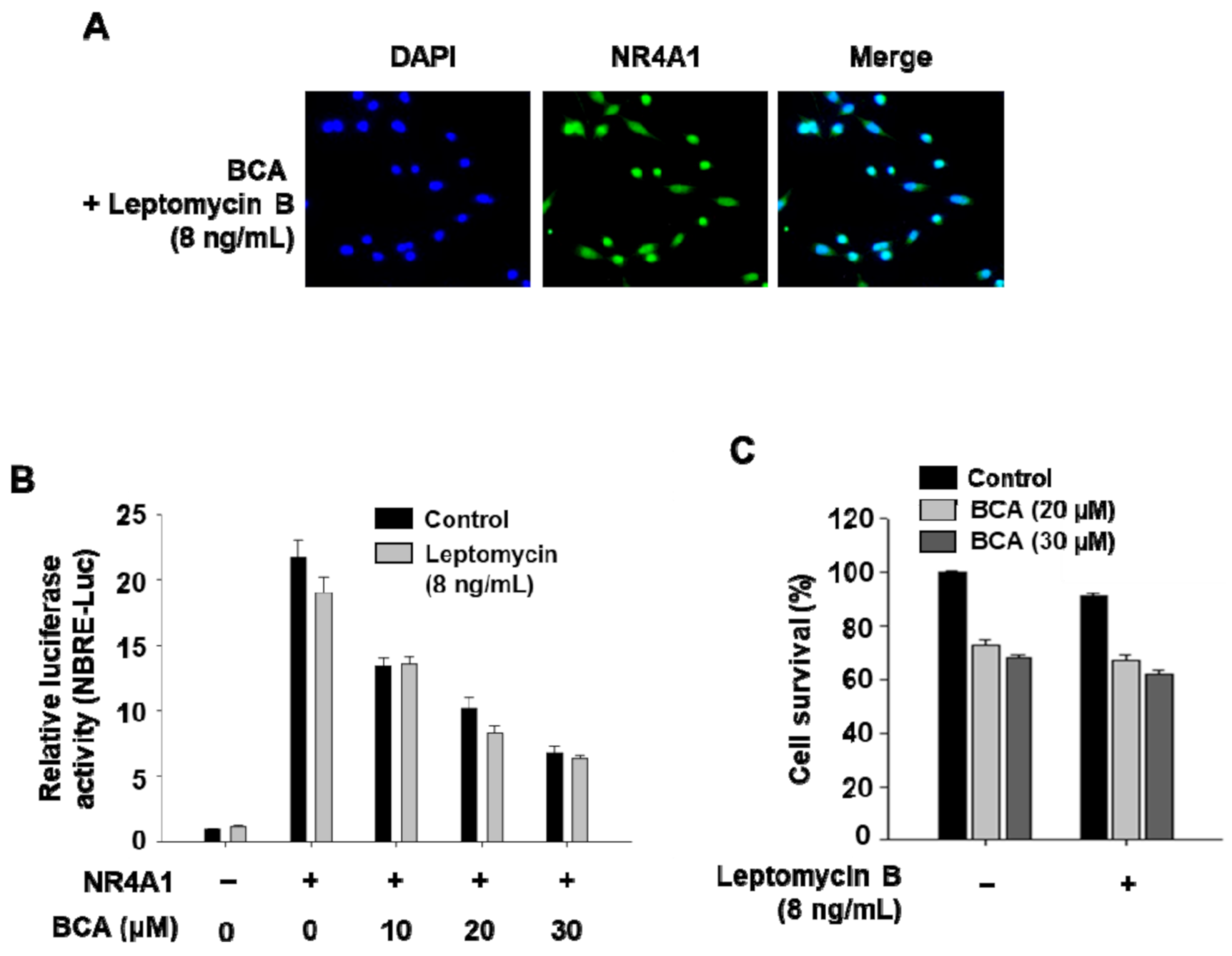
2.1. BCA Inhibits NR4A1-Mediated Transactivation in Pancreatic Cancer Cells
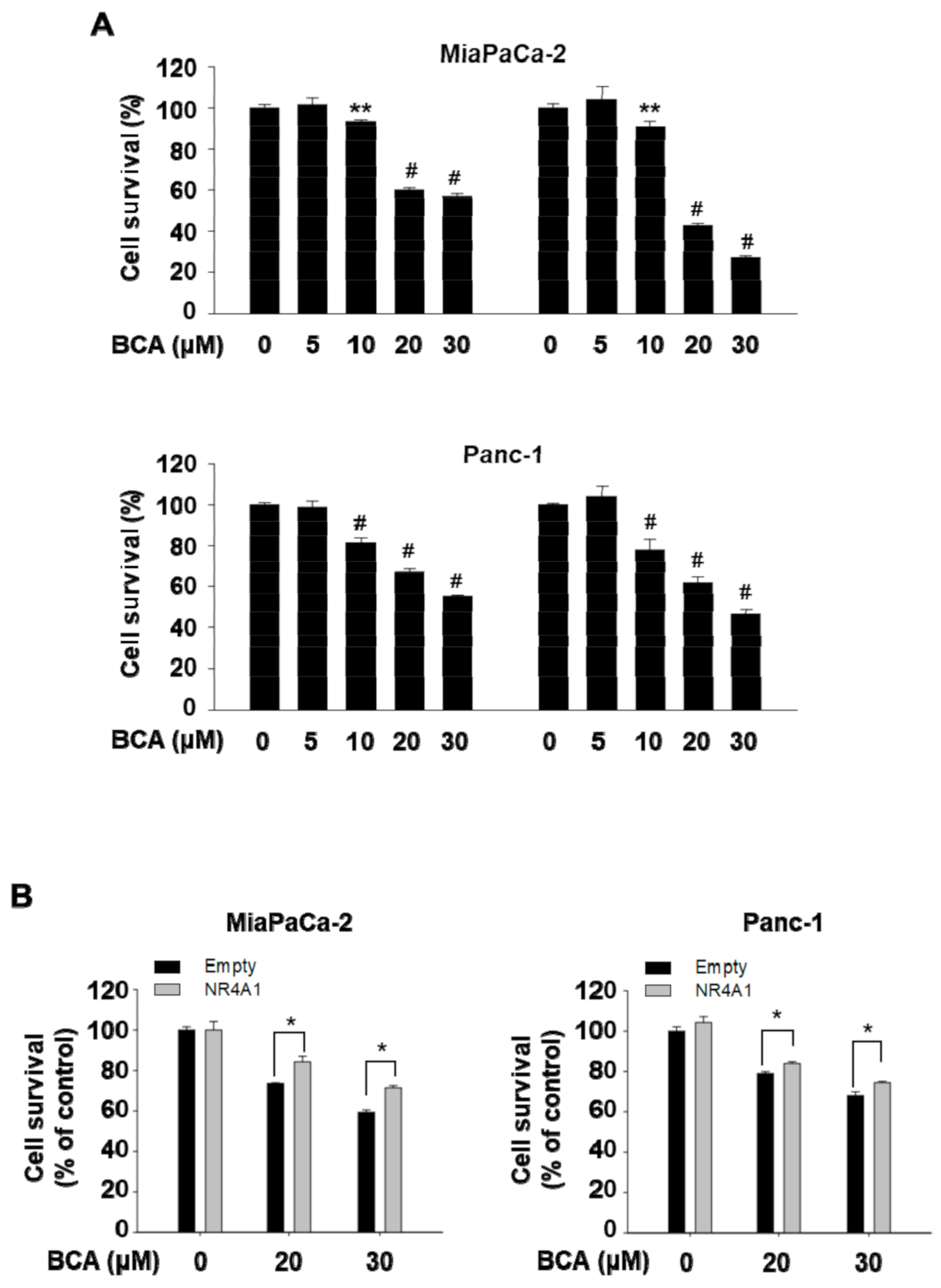
2.2. Inactivation of NR4A1 Contributes to Growth Inhibition by BCA in Pancreatic Cancer Cells
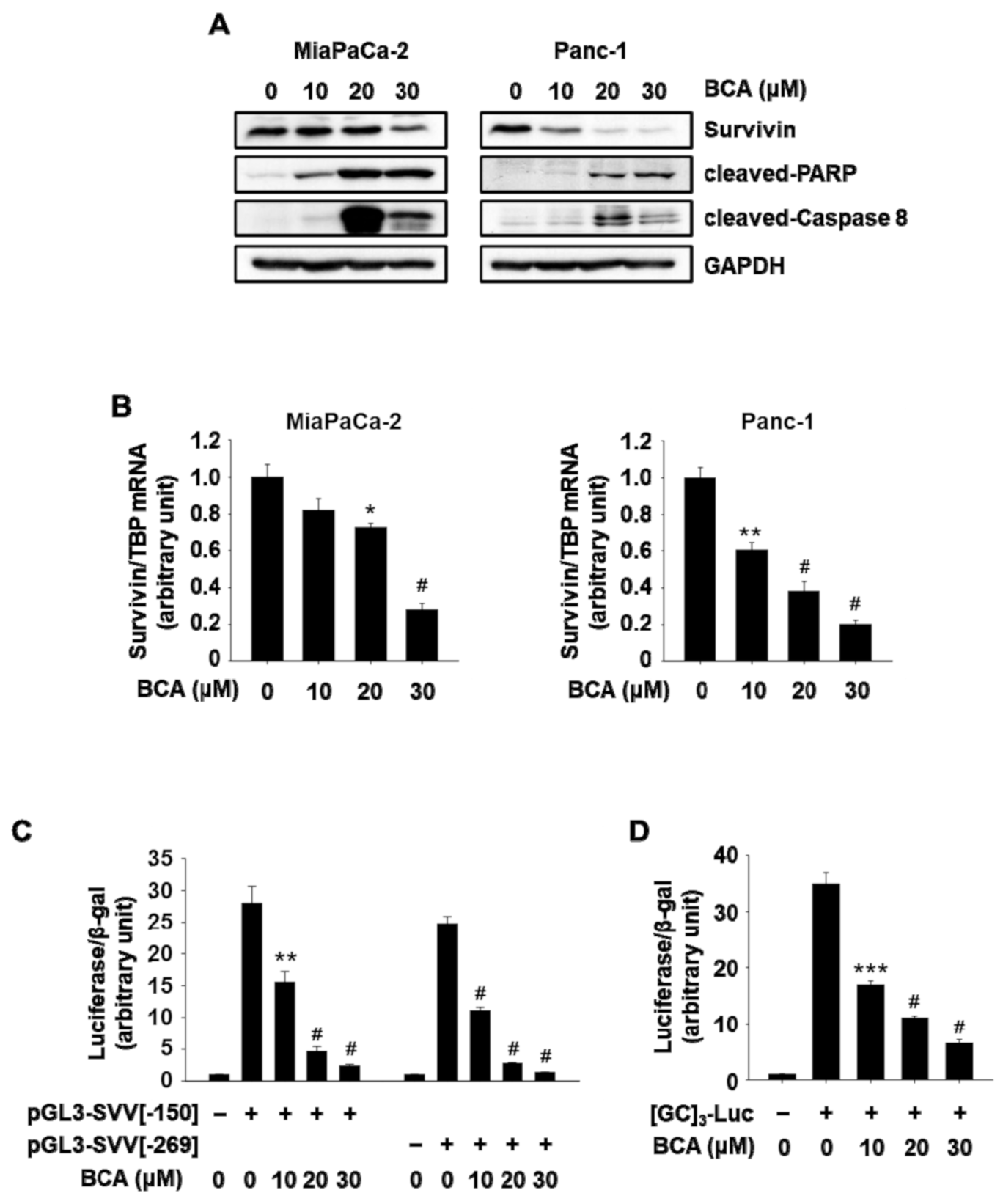
2.3. BCA Induces NR4A1-Dependent Apoptosis in Pancreatic Cancer Cells: Sp1/Survivin-Mediated Apoptosis
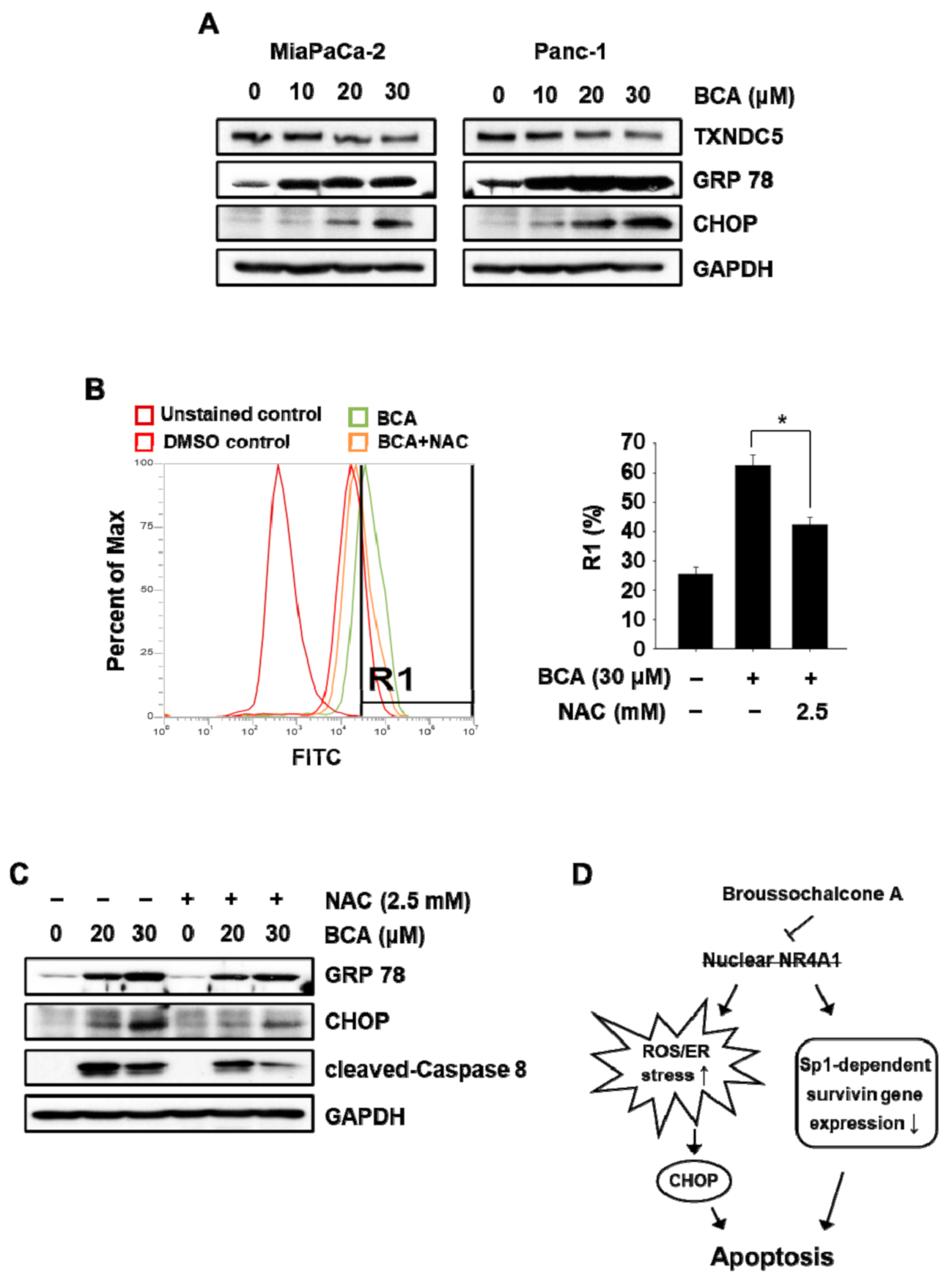
2.4. BCA Induces NR4A1-Dependent Apoptosis in Pancreatic Cancer Cells: ROS/ER Stress-Mediated Apoptosis
3. Materials and Methods
3.1. Isolation of BCA from Broussonetia Papyrifera Extract
3.2. Cell Lines and Plasmids
3.3. Antibodies, Reagents, Quantitative Real-Time PCR, and Western Blot Analysis
3.4. Cell Proliferation Assay, Transfection, and Reporter Gene Assay
3.5. Subcellular Localization Assays
3.6. Measurement of Intracellular Level of ROS
3.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hedrick, E.; Lee, S.O.; Doddapaneni, R.; Singh, M.; Safe, S. NR4A1 antagonists inhibit beta1-integrin-dependent breast cancer cell migration. Mol. Cell. Biol. 2016, 36, 1383–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.O.; Abdelrahim, M.; Yoon, K.; Chintharlapalli, S.; Papineni, S.; Kim, K.; Wang, H.; Safe, S. Inactivation of the orphan nuclear receptor TR3/Nur77 inhibits pancreatic cancer cell and tumor growth. Cancer Res. 2010, 70, 6824–6836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.O.; Andey, T.; Jin, U.H.; Kim, K.; Singh, M.; Safe, S. The nuclear receptor TR3 regulates mTORC1 signaling in lung cancer cells expressing wild-type p53. Oncogene 2012, 31, 3265–3276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.O.; Jin, U.H.; Kang, J.H.; Kim, S.B.; Guthrie, A.S.; Sreevalsan, S.; Lee, J.S.; Safe, S. The orphan nuclear receptor NR4A1 (Nur77) regulates oxidative and endoplasmic reticulum stress in pancreatic cancer cells. Mol. Cancer Res. 2014, 12, 527–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, B.; Kolluri, S.K.; Lin, F.; Liu, W.; Han, Y.H.; Cao, X.; Dawson, M.I.; Reed, J.C.; Zhang, X.K. Conversion of Bcl-2 from protector to killer by interaction with nuclear orphan receptor Nur77/TR3. Cell 2004, 116, 527–540. [Google Scholar] [CrossRef] [Green Version]
- Hedrick, E.; Lee, S.O.; Kim, G.; Abdelrahim, M.; Jin, U.H.; Safe, S.; Abudayyeh, A. Nuclear receptor 4A1 (NR4A1) as a drug target for renal cell adenocarcinoma. PLoS ONE 2015, 10, e0128308. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Safe, S.; Lee, S.O. Inactivation of the orphan nuclear receptor NR4A1 contributes to apoptosis induction by fanchinoline in pancreatic cancer cells. Toxicol. Appl. Pharmacol. 2017, 332, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, P.; Loh, W.M.; Gopinath, S.C.B.; Bonam, S.R.; Fareez, I.M.; Guad, R.M.; Sim, M.S.; Wu, Y.S. Selective phytochemicals targeting pancreatic stellate cells as new anti-fibrotic agents for chronic pancreatitis and pancreatic cancer. Acta Pharm. Sin. B 2020, 10, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Sultana, F.; Manasa, K.L.; Shaik, S.P.; Bonam, S.R.; Kamal, A. Zinc dependent histone deacetylase inhibitors in cancer therapeutics: Recent update. Curr. Med. Chem. 2019, 26, 7212–7280. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.R.; Lee, S.O. Novel hepatoprotective peptides derived from protein hydrolysates of mealworm (Tenebrio molitor). Food Res. Int. 2020, 33, 109194. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.D.; Yoon, K.; Chintharlapalli, S.; Abdelrahim, M.; Lei, P.; Hamilton, S.; Khan, S.; Ramaiah, S.K.; Safe, S. Nur77 agonists induce proapoptotic genes and responses in colon cancer cells through nuclear receptor-dependent and nuclear receptor-independent pathways. Cancer Res. 2007, 67, 674–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safe, S.; Kim, K.; Li, X.; Lee, S.O. NR4A orphan receptors and cancer. Nucl. Recept. Signal. 2011, 9, e002. [Google Scholar] [CrossRef]
- Bhanot:, U.; Heydrich, R.; Moller, P.; Hasel, C. Survivin expression in pancreatic intraepithelial neoplasia (PanIN): Steady increase along the developmental stages of pancreatic ductal adenocarcinoma. Am. J. Surg. Pathol. 2006, 30, 754–759. [Google Scholar] [CrossRef] [PubMed]
- Stauber, R.H.; Mann, W.; Knauer, S.K. Nuclear and cytoplasmic survivin: Molecular mechanism, prognostic, and therapeutic potential. Cancer Res. 2007, 67, 5999–6002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Z.P.; Cheng, K.W.; Chao, J.; Wu, J.; Wang, M. Tyrosinase inhibitors from paper mulberry (Broussonetia papyrifera). Food Chem. 2008, 106, 529–535. [Google Scholar] [CrossRef]
- Lin, C.N.; Lu, C.M.; Lin, H.C.; Fang, S.C.; Shieh, B.J.; Hsu, M.F.; Wang, J.P.; Ko, F.N.; Teng, C.M. Novel antiplatelet constituents from Formosan moraceous plants. J. Nat. Prod. 1996, 59, 834–838. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, R.; Chakraborty, A.; Biswas, A.; Chowdhuri, S. Identification of polyphenols from Broussonetia papyrifera as SARS CoV-2 main protease inhibitors using in silico docking and molecular dynamics simulation approaches. J. Biomol. Struct. Dyn. 2020, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Lin, C.; Hwang, T.; Teng, C. Broussochalcone A, a potent antioxidant and effective suppressor of inducible nitric oxide synthase in lipopolysaccharide-activated macrophages. Biochem. Pharmacol. 2001, 61, 939–946. [Google Scholar] [CrossRef]
- Park, S.H.; Lee, J.; Shon, J.C.; Phuc, N.M.; Jee, J.G.; Liu, K.H. The inhibitory potential of Broussochalcone A for the human cytochrome P450 2J2 isoform and its anti-cancer effects via FOXO3 activation. Phytomedicine 2018, 15, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Son, Y.; Liu, K.H.; Kang, W.; Oh, S. Cytotoxic activity of broussochalcone a against colon and liver cancer cells by promoting destruction complex-independent β-catenin degradation. Food Chem. Toxicol. 2019, 131, 110550. [Google Scholar] [CrossRef] [PubMed]





Sample Availability: Samples of the compound are available from the authors (Soo Hyun Kim and Bo Mi Kim). |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-S.; Kim, S.-H.; Kim, B.-M.; Safe, S.; Lee, S.-O. Broussochalcone A Is a Novel Inhibitor of the Orphan Nuclear Receptor NR4A1 and Induces Apoptosis in Pancreatic Cancer Cells. Molecules 2021, 26, 2316. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26082316
Lee H-S, Kim S-H, Kim B-M, Safe S, Lee S-O. Broussochalcone A Is a Novel Inhibitor of the Orphan Nuclear Receptor NR4A1 and Induces Apoptosis in Pancreatic Cancer Cells. Molecules. 2021; 26(8):2316. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26082316
Chicago/Turabian StyleLee, Hyo-Seon, Soo-Hyun Kim, Bo-Mi Kim, Stephen Safe, and Syng-Ook Lee. 2021. "Broussochalcone A Is a Novel Inhibitor of the Orphan Nuclear Receptor NR4A1 and Induces Apoptosis in Pancreatic Cancer Cells" Molecules 26, no. 8: 2316. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26082316