
Three Small Molecule Entities (MPK18, MPK334 and YAK308) with Activity against Haemonchus contortus In Vitro

 , , , , , ,
, , , , , ,  and
and
Abstract
:1. Introduction
2. Results and Discussion
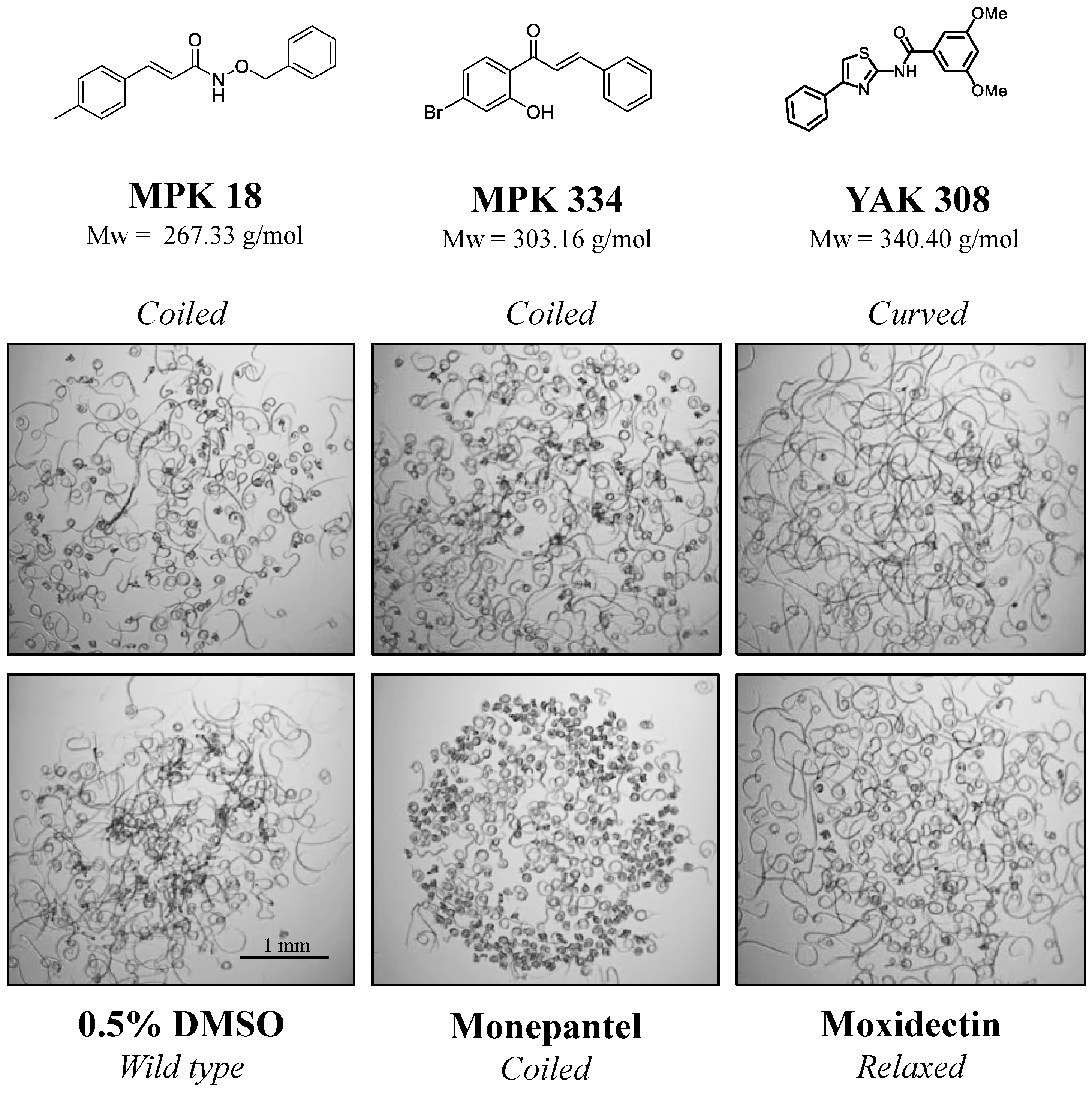
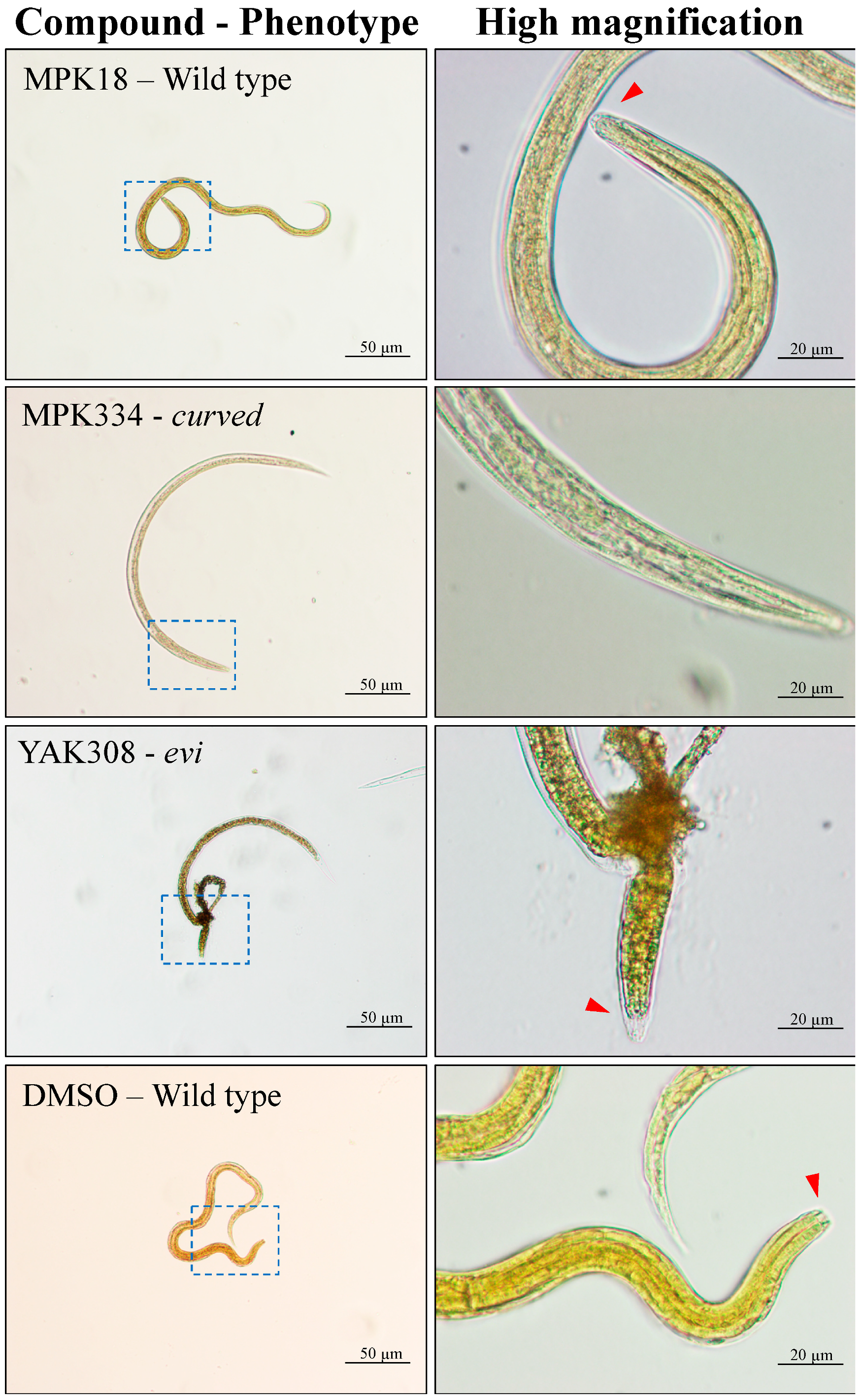
2.1. Three Compounds Induced a Phenotypic Alteration in the Primary Screen
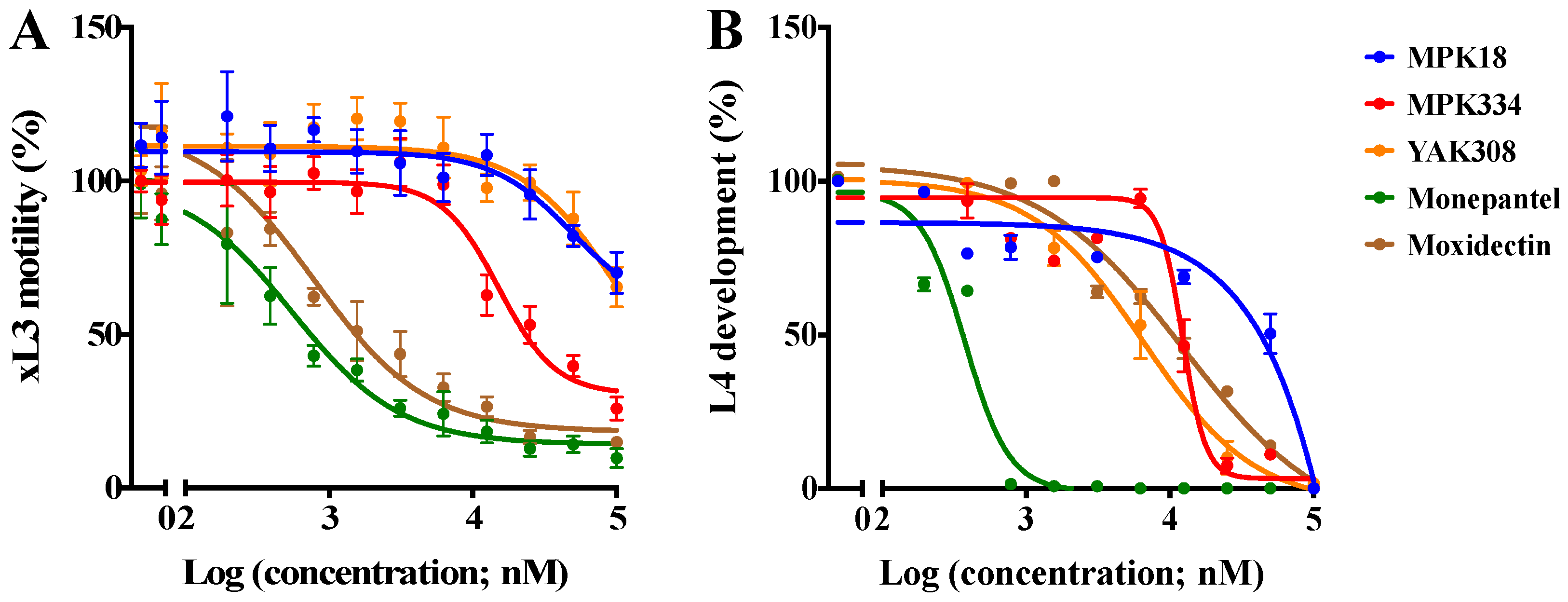
2.2. Inhibitory Effects of Active Compounds on Larval Motility and Development
3. Materials and Methods
3.1. Compound Preparation for Screening
3.2. Procurement and Preparation of Parasite Larvae
3.3. Screening of Compounds on H. contortus xL3s
3.4. Assessing Inhibitory Activity on Larval Motility
3.5. Evaluating Inhibitory Activity on Larval Development
3.6. Evaluation of Toxicity of Active Compounds on HepG2 Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Roeber, F.; Jex, A.R.; Gasser, R.B. Impact of gastrointestinal parasitic nematodes of sheep, and the role of advanced molecular tools for exploring epidemiology and drug resistance—An Australian perspective. Parasit. Vectors 2013, 6, 153. [Google Scholar] [CrossRef] [Green Version]
- Lane, J.; Jubb, T.; Shepherd, R.; Webb-Ware, J.; Fordyce, G. Priority List of Endemic Diseases for the Red Meat Industries. Final Report (B.AHE.0010); Meat & Livestock Australia Ltd.: North Sydney, New South Wales, Australia, 2015; ISBN -9781741918946. [Google Scholar]
- Gasser, R.B.; von Samson-Himmelstjerna, G. Haemonchus contortus and Haemonchosis—Past, Present and Future Trends. In Advances in Parasitology; Academic Press: London, UK, 2016; Volume 111, ISBN 978-0-128-10395-1. [Google Scholar]
- Besier, R.B.; Kahn, L.P.; Sargison, N.D.; Van Wyk, J.A. Diagnosis, treatment and management of Haemonchus contortus in small ruminants. Adv. Parasitol. 2016, 93, 181–238. [Google Scholar]
- Sepúlveda-Crespo, D.; Reguera, R.M.; Rojo-Vázquez, F.; Balaña-Fouce, R.; Martínez-Valladares, M. Drug discovery technologies: Caenorhabditis elegans as a model for anthelmintic therapeutics. Med. Res. Rev. 2020, 40, 1715–1753. [Google Scholar] [CrossRef]
- Kotze, A.C.; Prichard, R.K. Anthelmintic resistance in Haemonchus contortus: History, mechanisms and diagnosis. Adv. Parasitol. 2016, 93, 397–428. [Google Scholar]
- Nisbet, A.J.; Meeusen, E.N.; González, J.F.; Piedrafita, D.M. Immunity to Haemonchus contortus and vaccine development. Adv. Parasitol. 2016, 93, 353–396. [Google Scholar] [PubMed]
- Kaminsky, R.; Gauvry, N.; Weber, S.S.; Skripsky, T.; Bouvier, J.; Wenger, A.; Schroeder, F.; Desaules, Y.; Hotz, R.; Goebel, T.; et al. Identification of the amino-acetonitrile derivative monepantel (AAD 1566) as a new anthelmintic drug development candidate. Parasitol. Res. 2008, 103, 931–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prichard, R.K.; Geary, T.G. Fresh hope to can the worms. Nature 2008, 452, 157–158. [Google Scholar] [CrossRef] [PubMed]
- Little, P.; Hodge, A.; Watson, T.; Seed, J.; Maeder, S. Field efficacy and safety of an oral formulation of the novel combination anthelmintic, derquantel-abamectin, in sheep in New Zealand. N. Z. Vet. J. 2010, 58, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Preston, S.; Gasser, R.B. Working towards new drugs against parasitic worms in a public-development partnership. Trends Parasitol. 2018, 34, 4–6. [Google Scholar] [CrossRef]
- Jiao, Y.; Preston, S.; Hofmann, A.; Taki, A.; Baell, J.; Chang, B.C.H.; Jabbar, A.; Gasser, R.B. A perspective on the discovery of selected compounds with anthelmintic activity against the barber’s pole worm-Where to from here? Adv. Parasitol. 2020, 108, 1–45. [Google Scholar] [PubMed]
- Herath, H.M.P.D.; Taki, A.C.; Sleebs, B.E.; Nguyen, N.; Hofmann, A.; Jabbar, A.; Davis, D.A.; Gasser, R.B. Advances in the discovery and development of anthelmintics by harnessing natural product scaffolds. Adv. Parasitol. 2021, 111, 203–251. [Google Scholar]
- Preston, S.; Jabbar, A.; Nowell, C.; Joachim, A.; Ruttkowski, B.; Baell, J.; Cardno, T.; Korhonen, P.K.; Piedrafita, D.; Ansell, B.R.E.; et al. Low cost whole-organism screening of compounds for anthelmintic activity. Int. J. Parasitol. 2015, 45, 333–343. [Google Scholar] [CrossRef] [Green Version]
- Asfaha, Y.; Schrenk, C.; Avelar, L.A.A.; Lange, F.; Wang, C.; Bandolik, J.J.; Hamacher, A.; Kassack, M.U.; Kurz, T. Novel alkoxyamide-based histone deacetylase inhibitors reverse cisplatin resistance in chemoresistant cancer cells. Bioorg. Med. Chem. 2019, 28, 115108. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-A.; Park, S.-M.; Chu, K.-B.; Quan, F.-S.; Kurz, T.; Pflieger, M.; Moon, E.-K. Application of histone deacetylase inhibitors MPK472 and KSK64 as a potential treatment option for Acanthamoeba keratitis. Antimicrobiol. Agents 2020, 64, e01506-20. [Google Scholar] [CrossRef]
- Jiao, Y.; Preston, S.; Koehler, A.V.; Stroehlein, A.J.; Chang, B.C.H.; Simpson, K.J.; Cowley, K.J.; Palmer, M.J.; Laleu, B.; Wells, T.N.C.; et al. Screening of the ‘Stasis Box’ identifies two kinase inhibitors under pharmaceutical development with activity against Haemonchus contortus. Parasit. Vectors 2017, 10, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preston, S.; Jiao, Y.; Baell, J.B.; Keiser, J.; Crawford, S.; Koehler, A.V.; Wang, T.; Simpson, M.M.; Kaplan, R.M.; Cowley, K.J.; et al. Screening of the ‘Open Scaffolds’ collection from Compounds Australia identifies a new chemical entity with anthelmintic activities against different developmental stages of the barber’s pole worm and other parasitic nematodes. Int. J. Parasitol. Drugs Drug Resist. 2017, 7, 286–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.T.; Kurz, T.; Preston, S.; Brueckmann, H.; Lungerich, B.; Herath, H.M.P.D.; Koehler, A.V.; Wang, T.; Skálová, L.; Jabbar, A.; et al. Phenotypic screening of the ‘Kurz-box’ of chemicals identifies two compounds (BLK127 and HBK4) with anthelmintic activity in vitro against parasitic larval stages of Haemonchus contortus. Parasit. Vectors 2019, 12, 191. [Google Scholar] [CrossRef] [PubMed]
- de Castro, C.C.B.; Costa, P.S.; Laktin, G.T.; de Carvalho, P.H.D.; Geraldo, R.B.; de Moraes, J.; Pinto, P.L.S.; Couri, M.R.C.; Pinto, P. de F.; Da Silva Filho, A.A. Cardamonin, a schistosomicidal chalcone from Piper aduncum L. (Piperaceae) that inhibits Schistosoma mansoni ATP diphosphohydrolase. Phytomedicine 2015, 22, 921–928. [Google Scholar] [CrossRef] [Green Version]
- Caboni, P.; Aissani, N.; Demurtas, M.; Ntalli, N.; Onnis, V. Nematicidal activity of acetophenones and chalcones against Meloidogyne incognita and structure–activity considerations. Pest. Manag. Sci. 2016, 72, 125–130. [Google Scholar] [CrossRef]
- Silva, F.J.; Campos, V.P.; Oliveira, D.F.; Gomes, V.A.; Barros, A.F.; Din, Z.U.; Rodrigues-Filho, E. Chalcone analogues: Synthesis, activity against Meloidogyne incognita, and in silico interaction with cytochrome P450. J. Phytopathol. 2019, 167, 197–208. [Google Scholar] [CrossRef]
- Sinha, S.; Batovska, D.I.; Medhi, B.; Radotra, B.D.; Bhalla, A.; Markova, N.; Sehgal, R. In vitro anti-malarial efficacy of chalcones: Cytotoxicity profile, mechanism of action and their effect on erythrocytes. Malaria J. 2019, 18, 421. [Google Scholar] [CrossRef]
- Ochoa, R.; García, E.; Robledo, S.M.; G., W.C. Virtual and experimental screening of phenylfuranchalcones as potential anti-Leishmania candidates. J. Mol. Graph. Model. 2019, 91, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Chacon-Vargas, K.F.; Domínguez-Méndez, V.G.; Nogueda-Torres, B.; Chávez-Flores, D.; Camacho-Dávila, A.A.; Sánchez-Torres, L.E.; Espinoza-Hicks, J.C. O-geranylchalcones: Synthesis and metabolic inhibition against Leishmania mexicana and Trypanosoma cruzi. Med. Chem. Res. 2020, 29, 156–165. [Google Scholar] [CrossRef]
- Cáceres-Castillo, D.; Carballo, R.M.; Quijano-Quiñones, R.; Mirón-López, G.; Graniel-Sabido, M.; Moo-Puc, R.E.; Mena-Rejón, G.J. Synthesis, in vitro antigiardial activity, SAR analysis and docking study of substituted chalcones. Med. Chem. Res. 2020, 29, 431–441. [Google Scholar] [CrossRef]
- Vázquez-Bravo, J.; Aguilar-Marcelino, L.; Castañeda-Ramírez, G.S.; De los Santos-Pérez, I.; Arroyo-Carmona, R.E.; Bernès, S.; Hernández-Pareja, U.; Gómez-Rodríguez, O.; Rosas-Saito, G.H. In vitro nematicidal activity of two ferrocenyl chalcones against larvae of Haemonchus contortus (L3) and Nacobbus aberrans (J2). J. Helminthol. 2020, 94, e190. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, C.; Zhang, W.; Sheng, C.; Zhang, W.; Xing, C.; Miao, Z. Chalcone: A privileged structure in medicinal chemistry. Chem. Rev. 2017, 117, 7762–7810. [Google Scholar] [CrossRef] [PubMed]
- Gaonkar, S.L.; Vignesh, U.N. Synthesis and pharmacological properties of chalcones: A review. Res. Chem. Intermed. 2017, 43, 6043–6077. [Google Scholar] [CrossRef]
- Gomes, M.N.; Muratov, E.N.; Pereira, M.; Peixoto, J.C.; Rosseto, L.P.; Cravo, P.V.L.; Andrade, C.H.; Neves, B.J. Chalcone derivatives: Promising starting points for drug design. Molecules 2017, 22, 1210. [Google Scholar] [CrossRef] [Green Version]
- Page, A.P.; Stepek, G.; Winter, A.D.; Pertab, D. Enzymology of the nematode cuticle: A potential drug target? Int. J. Parasitol. Drugs Drug Resist. 2014, 4, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Preston, S.; Garcia-Bustos, J.F.; Baell, J.B.; Ventura, S.; Le, T.; McNamara, N.; Nguyen, N.; Botteon, A.; Skinner, C.; et al. Tetrahydroquinoxalines induce a lethal evisceration phenotype in Haemonchus contortus in vitro. Int. J. Parasitol. Drugs Drug Resist. 2018, 9, 59–71. [Google Scholar] [CrossRef]
- Pereira, A.S.A.; Silveira, G.O.; Amaral, M.S.; Almeida, S.M.V.; Oliveira, J.F.; Lima, M.C.A.; Verjovski-Almeida, S. In vitro activity of aryl-thiazole derivatives against Schistosoma mansoni schistosomula and adult worms. PLoS ONE 2019, 14, e0225425. [Google Scholar] [CrossRef] [PubMed]
- Ouattara, M.; Sissouma, D.; Koné, M.; Menan, H.; Touré, S.; Ouattara, L. Synthesis and anthelmintic activity of some hybrid benzimidazolyl-chalcone derivatives. Trop. J. Pharm. Res. 2011, 10, 767–775. [Google Scholar] [CrossRef]
- Maghraby, M.T.-E.; Abou-Ghadir, O.M.F.; Abdel-Moty, S.G.; Ali, A.Y.; Salem, O.I.A. Novel class of benzimidazole-thiazole hybrids: The privileged scaffolds of potent anti-inflammatory activity with dual inhibition of cyclooxygenase and 15-lipoxygenase enzymes. Bioorg. Med. Chem. 2020, 28, 115403. [Google Scholar] [CrossRef]
- Schwarz, E.M.; Korhonen, P.K.; Campbell, B.E.; Young, N.D.; Jex, A.R.; Jabbar, A.; Hall, R.S.; Mondal, A.; Howe, A.C.; Pell, J.; et al. The genome and developmental transcriptome of the strongylid nematode Haemonchus contortus. Genome Biol. 2013, 14, R89. [Google Scholar] [CrossRef] [Green Version]
- Sommerville, R.I. The development of Haemonchus contortus to the fourth stage in vitro. J. Parasitol. 1966, 52, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Śliwka, L.; Wiktorska, K.; Suchocki, P.; Milczarek, M.; Mielczarek, S.; Lubelska, K.; Cierpiał, T.; Łyżwa, P.; Kiełbasiński, P.; Jaromin, A. The comparison of MTT and CVS assays for the assessment of anticancer agent interactions. PLoS ONE 2016, 11, e0155772. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Compound | xL3 Motility (72 h) IC50 ± SEM (µM) | xL3 Motility (72 h) Inhibition (%) | L4 Development (7 days) IC50 ± SEM (µM) | HepG2 Cells (48 h) CC50 (µM) | SI xL3 Motility/L4 Development |
|---|---|---|---|---|---|
| MPK18 | 45.2 ± 4.6 | 30.0 | nd | >100 | >1.0/nd |
| MPK334 | 17.1 ± 4.2 | 74.2 | 12.3 ± 2.3 | >100 | 5.8/8.1 |
| YAK308 | 52.7 ± 6.7 | 34.6 | 6.5 ± 1.6 | >100 | >1.0/16.1 |
| Monepantel | 0.6 ± 0.1 | 90.3 | 0.4 ± 0.05 | >100 | 166/250 |
| Moxidectin | 0.8 ± 0.2 | 85.1 | 12.4 ± 0.1 | >100 | 125/8.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taki, A.C.; Jabbar, A.; Kurz, T.; Lungerich, B.; Ma, G.; Byrne, J.J.; Pflieger, M.; Asfaha, Y.; Fischer, F.; Chang, B.C.H.; et al. Three Small Molecule Entities (MPK18, MPK334 and YAK308) with Activity against Haemonchus contortus In Vitro. Molecules 2021, 26, 2819. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092819
Taki AC, Jabbar A, Kurz T, Lungerich B, Ma G, Byrne JJ, Pflieger M, Asfaha Y, Fischer F, Chang BCH, et al. Three Small Molecule Entities (MPK18, MPK334 and YAK308) with Activity against Haemonchus contortus In Vitro. Molecules. 2021; 26(9):2819. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092819
Chicago/Turabian StyleTaki, Aya C., Abdul Jabbar, Thomas Kurz, Beate Lungerich, Guangxu Ma, Joseph J. Byrne, Marc Pflieger, Yodita Asfaha, Fabian Fischer, Bill C. H. Chang, and et al. 2021. "Three Small Molecule Entities (MPK18, MPK334 and YAK308) with Activity against Haemonchus contortus In Vitro" Molecules 26, no. 9: 2819. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26092819