
Fermentation of Jamaican Cherries Juice Using Lactobacillus plantarum Elevates Antioxidant Potential and Inhibitory Activity against Type II Diabetes-Related Enzymes

 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
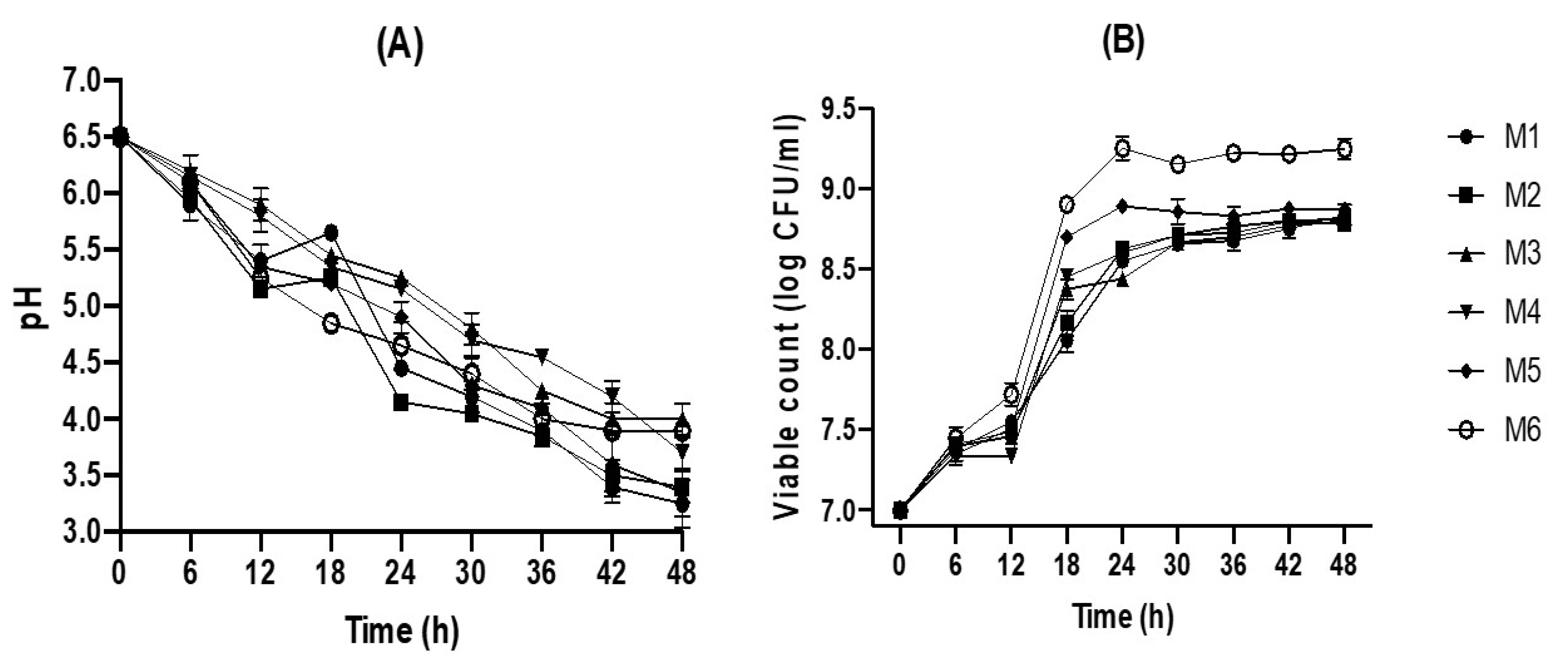
2.1. Fermentation of Jamaican Juice by L. plantarum
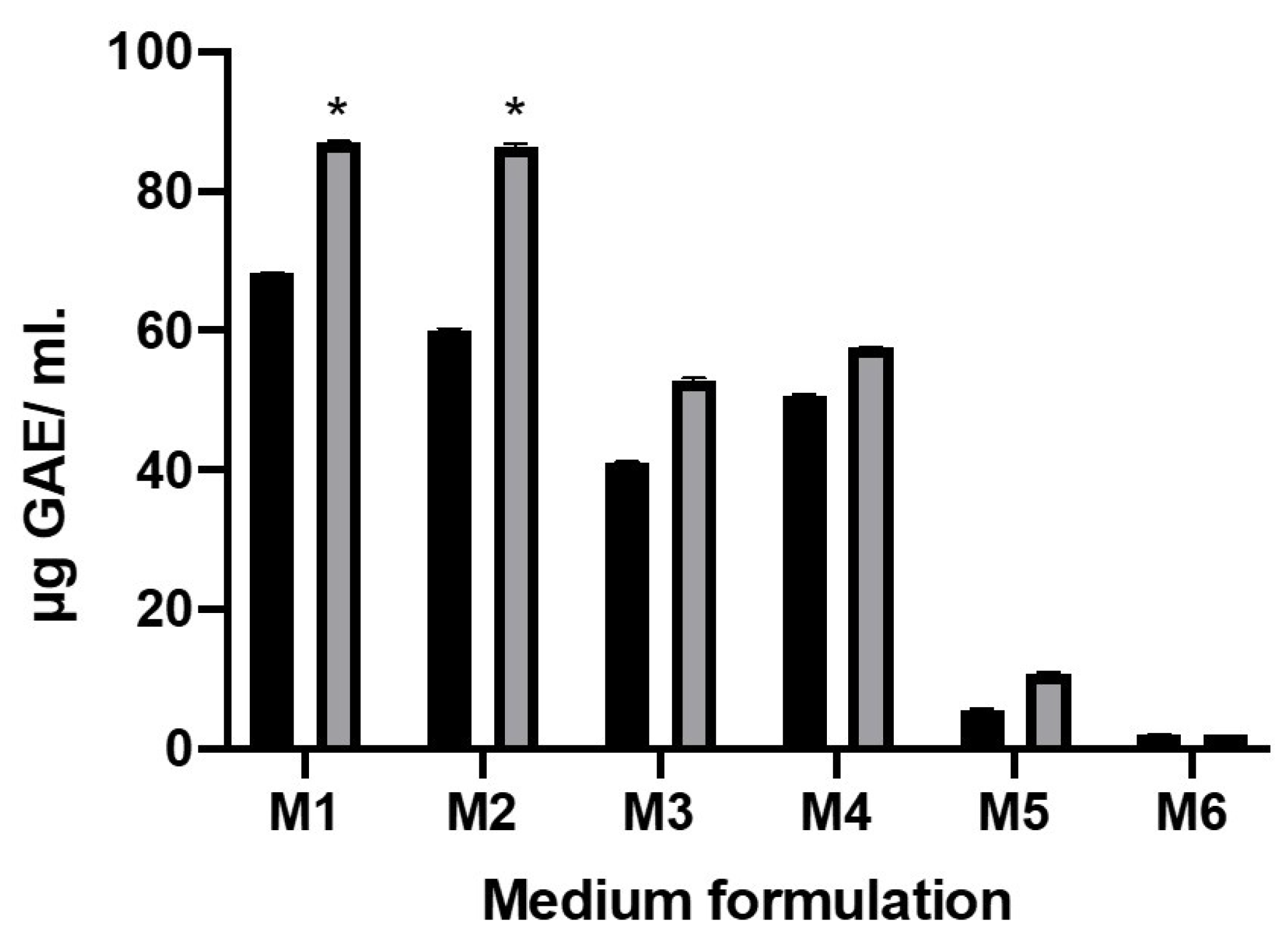
2.2. Total Phenolic Content after Fermentation
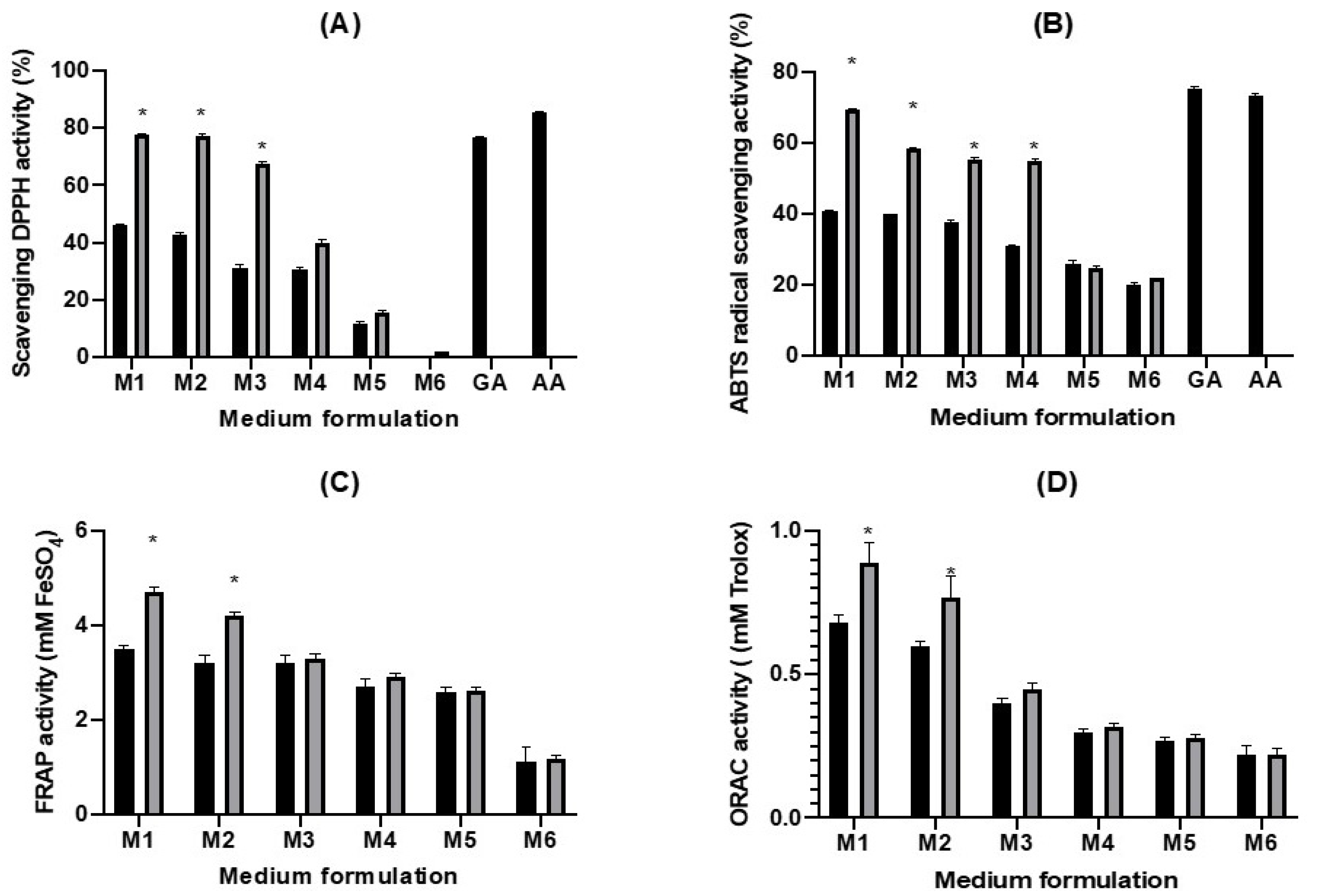
2.3. Antioxidative Activity of the Fermented Juices
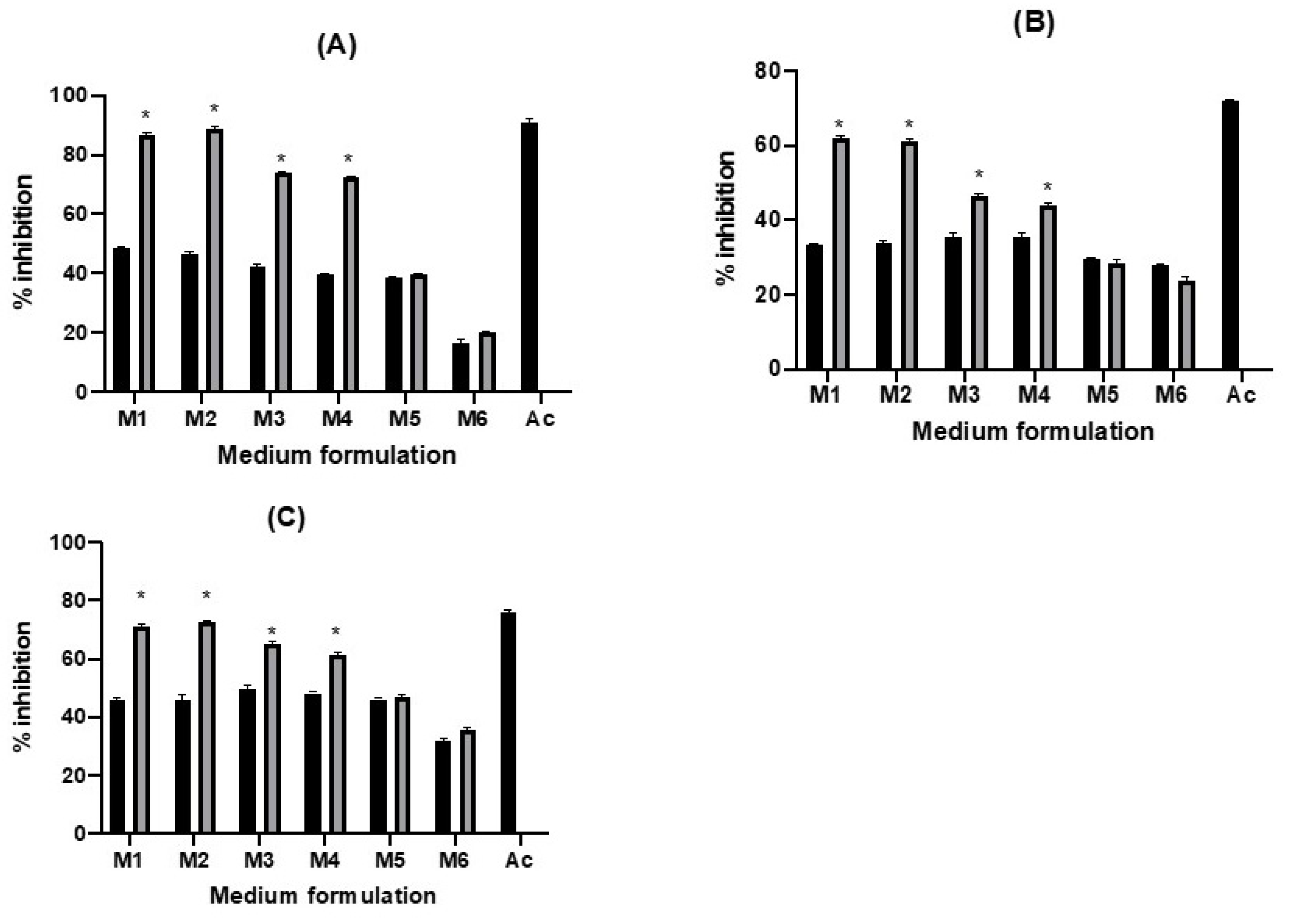
2.4. Enzyme Inhibition Ability
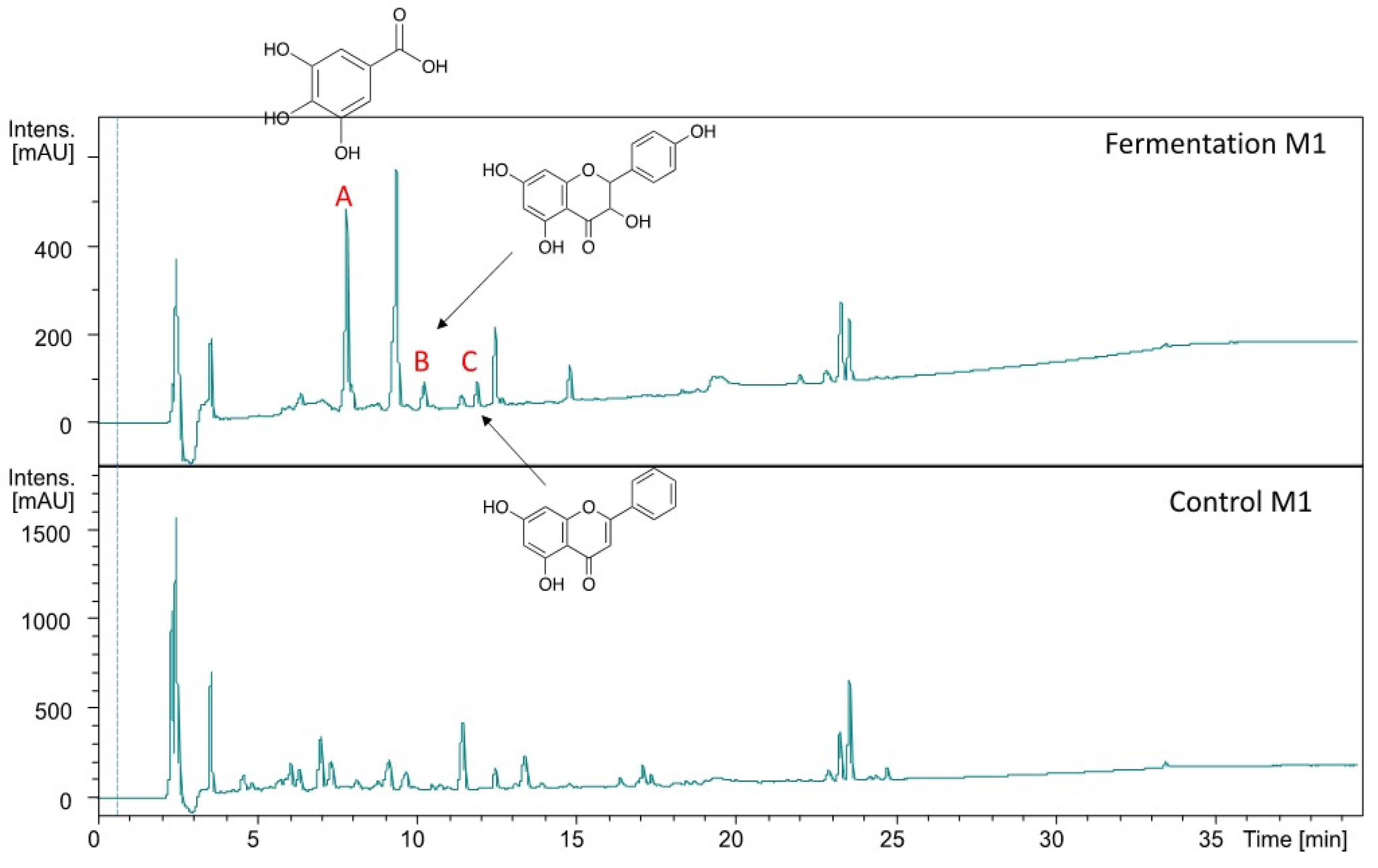
2.5. Mass Spectrometry Analysis of M1 Substrate
3. Materials and Methods
3.1. Chemical and Reagents
3.2. Bacterial Strain and Fruit Material
3.3. Preparation of Inoculant
3.4. Fermentation Procedure
3.5. Bacterial Cell Separation
3.6. Bacterial Viable Count
3.7. pH Measurement
3.8. Total Phenolic Content Measurement
3.9. 2,2-Diphenyl-1-picrylhydrazyl (DPPH) Radical Scavenging Activity
3.10. 2,2′-Azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) Assay
3.11. Ferric Reducing-Antioxidant Power (FRAP) Assay
3.12. Oxygen Radical Absorbance Capacity (ORAC) Assay
3.13. In Vitro Inhibiting Activity of α-Glucosidase
3.14. In Vitro Inhibiting Activity of α-Amylase
3.15. In Vitro Inhibiting Activity of Amyloglucosidase
3.16. UPLC-HRMS-MS Analysis
3.17. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Mahmood, N.; Nasir, N.; Rofiee, M.; Tohid, S.M.; Ching, S.; Teh, L.; Salleh, M.; Zakaria, Z. Muntingia calabura: A review of its traditional uses, chemical properties, and pharmacological observations. Pharm. Biol. 2014, 52, 1598–1623. [Google Scholar] [CrossRef]
- Harshini, V.; Gayathri, H.S.; Padmaja, A. Development of Muntingia calabura Fruit Based Squash. Asian J. Dairy Food Res. 2020, 9, 256–260. [Google Scholar]
- Buhian, W.P.C.; Rubio, R.O.; Valle, D.L., Jr.; Martin-Puzon, J.J. Bioactive metabolite profiles and antimicrobial activity of ethanolic extracts from Muntingia calabura L. leaves and stems. Asian Pac. J. Trop. Biomed. 2016, 6, 682–685. [Google Scholar] [CrossRef]
- Zolkeflee, N.K.Z.; Isamail, N.A.; Maulidiani, M.; Abdul Hamid, N.A.; Ramli, N.S.; Azlan, A.; Abas, F. Metabolite variations and antioxidant activity of Muntingia calabura leaves in response to different drying methods and ethanol ratios elucidated by NMR-based metabolomics. Phytochem. Anal. 2021, 32, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Sufian, A.S.; Ramasamy, K.; Ahmat, N.; Zakaria, Z.A.; Yusof, M.I.M. Isolation and identification of antibacterial and cytotoxic compounds from the leaves of Muntingia calabura L. J. Ethnopharmacol. 2013, 146, 198–204. [Google Scholar] [CrossRef]
- Zakaria, Z.A.; Mahmood, N.D.; Mamat, S.S.; Nasir, N.; Omar, M.H. Endogenous Antioxidant and LOX-Mediated Systems Contribute to the Hepatoprotective Activity of Aqueous Partition of Methanol Extract of Muntingia calabura L. Leaves against Paracetamol Intoxication. Front. Pharmacol. 2018, 8, 982. [Google Scholar] [CrossRef] [Green Version]
- Shih, C.-D.; Chen, J.-J.; Lee, H.-H. Activation of nitric oxide signaling pathway mediates hypotensive effect of Muntingia calabura L. (Tiliaceae) leaf extract. Am. J. Chin. Med. 2006, 34, 857–872. [Google Scholar] [CrossRef] [Green Version]
- Pereira, G.A.; Arruda, H.S.; de Morais, D.R.; Eberlin, M.N.; Pastore, G.M. Carbohydrates, volatile and phenolic compounds composition, and antioxidant activity of calabura (Muntingia calabura L.) fruit. Food Res. Int. 2018, 108, 264–273. [Google Scholar] [CrossRef]
- Recuenco, M.C.; Lacsamana, M.S.; Hurtada, W.A.; Sabularse, V.C. Total phenolic and total flavonoid contents of selected fruits in the Philippines. Philipp. J. Sci. 2016, 145, 275–281. [Google Scholar]
- Frediansyah, A.; Nurhayati, R.; Sholihah, J. Lactobacillus pentosus isolated from Muntingia calabura shows inhibition activity toward alpha-glucosidase and alpha-amylase in intra and extracellular level. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2019; p. 012045. [Google Scholar]
- Zielińska, D.; Kolożyn-Krajewska, D. Food-origin lactic acid bacteria may exhibit probiotic properties. Biomed. Res. Int. 2018, 2018. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Lu, Y.; Yu, H.; Chen, Z.; Tian, H. Influence of 4 lactic acid bacteria on the flavor profile of fermented apple juice. Food Biosci. 2019, 27, 30–36. [Google Scholar] [CrossRef]
- Parada, J.L.; Caron, C.R.; Medeiros, A.B.P.; Soccol, C.R. Bacteriocins from lactic acid bacteria: Purification, properties and use as biopreservatives. Braz. Arch. Biol. Technol. 2007, 50, 512–542. [Google Scholar] [CrossRef] [Green Version]
- Sharma, N.; Angural, S.; Rana, M.; Puri, N.; Kondepudi, K.K.; Gupta, N. Phytase producing lactic acid bacteria: Cell factories for enhancing micronutrient bioavailability of phytate rich foods. Trends Food Sci. Technol. 2020, 96, 1–12. [Google Scholar] [CrossRef]
- Szutowska, J. Functional properties of lactic acid bacteria in fermented fruit and vegetable juices: A systematic literature review. Eur. Food Res. Technol. 2020, 246, 357–372. [Google Scholar] [CrossRef]
- Frediansyah, A.; Nurhayati, R.; Romadhoni, F. Enhancement of antioxidant activity, α-glucosidase and α-amylase inhibitory activities by spontaneous and bacterial monoculture fermentation of Indonesian black grape juices. In AIP Conference Proceedings; AIP Publishing LLC: Melville, NY, USA, 2017; p. 020022. [Google Scholar]
- Bergqvist, S.W.; Andlid, T.; Sandberg, A.-S. Lactic acid fermentation stimulated iron absorption by Caco-2 cells is associated with increased soluble iron content in carrot juice. Br. J. Nutr. 2006, 96, 705–711. [Google Scholar]
- Du, X.; Myracle, A.D. Fermentation alters the bioaccessible phenolic compounds and increases the alpha-glucosidase inhibitory effects of aronia juice in a dairy matrix following in vitro digestion. Food Funct. 2018, 9, 2998–3007. [Google Scholar] [CrossRef] [PubMed]
- Siro, I.; Kápolna, E.; Kápolna, B.; Lugasi, A. Functional food. Product development, marketing and consumer acceptance—A review. Appetite 2008, 51, 456–467. [Google Scholar] [CrossRef]
- Østlie, H.M.; Helland, M.H.; Narvhus, J.A. Growth and metabolism of selected strains of probiotic bacteria in milk. Int. J. Food Microbiol. 2003, 87, 17–27. [Google Scholar] [CrossRef]
- McFeeters, R. Fermentation microorganisms and flavor changes in fermented foods. J. Food Sci. 2004, 69, FMS35–FMS37. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, W.; Wei, Z.; Yin, B.; Man, C.; Jiang, Y. Enhancement of functional characteristics of blueberry juice fermented by Lactobacillus plantarum. LWT 2021, 139, 110590. [Google Scholar] [CrossRef]
- Kwaw, E.; Ma, Y.; Tchabo, W.; Apaliya, M.T.; Wu, M.; Sackey, A.S.; Xiao, L.; Tahir, H.E. Effect of lactobacillus strains on phenolic profile, color attributes and antioxidant activities of lactic-acid-fermented mulberry juice. Food Chem. 2018, 250, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, R.; Zhang, Y.; Yang, Y.; Sun, X.; Zhang, Q.; Yang, N. Biotransformation of phenolics and metabolites and the change in antioxidant activity in kiwifruit induced by Lactobacillus plantarum fermentation. J. Sci. Food Agric. 2020, 100, 3283–3290. [Google Scholar] [CrossRef]
- Liao, X.-Y.; Guo, L.-Q.; Ye, Z.-W.; Qiu, L.-Y.; Gu, F.-W.; Lin, J.-F. Use of autochthonous lactic acid bacteria starters to ferment mango juice for promoting its probiotic roles. Prep. Biochem. Biotechnol. 2016, 46, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Pisoschi, A.M.; Pop, A.; Cimpeanu, C.; Predoi, G. Antioxidant capacity determination in plants and plant-derived products: A review. Oxidative Med. Cell. Longev. 2016, 2016. [Google Scholar] [CrossRef] [Green Version]
- Santos-Sánchez, N.F.; Salas-Coronado, R.; Villanueva-Cañongo, C.; Hernández-Carlos, B. Antioxidant Compounds and Their Antioxidant Mechanism; IntechOpen: London, UK, 2019. [Google Scholar]
- Preethi, K.; Vijayalakshmi, N.; Shamna, R.; Sasikumar, J. In vitro antioxidant activity of extracts from fruits of Muntingia calabura Linn from India. Pharmacogn. J. 2010, 2, 11–18. [Google Scholar] [CrossRef]
- Muhialdin, B.J.; Kadum, H.; Zarei, M.; Hussin, A.S.M. Effects of metabolite changes during lacto-fermentation on the biological activity and consumer acceptability for dragon fruit juice. LWT 2020, 121, 108992. [Google Scholar] [CrossRef]
- Wu, C.; Li, T.; Qi, J.; Jiang, T.; Xu, H.; Lei, H. Effects of lactic acid fermentation-based biotransformation on phenolic profiles, antioxidant capacity and flavor volatiles of apple juice. LWT 2020, 122, 109064. [Google Scholar] [CrossRef]
- Hashemi, S.M.B.; Jafarpour, D. Fermentation of bergamot juice with Lactobacillus plantarum strains in pure and mixed fermentations: Chemical composition, antioxidant activity and sensorial properties. LWT 2020, 131, 109803. [Google Scholar] [CrossRef]
- Hur, S.J.; Lee, S.Y.; Kim, Y.-C.; Choi, I.; Kim, G.-B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef]
- Friedman, M.; Jürgens, H.S. Effect of pH on the stability of plant phenolic compounds. J. Agric. Food Chem. 2000, 48, 2101–2110. [Google Scholar] [CrossRef]
- Nurhayati, R.; Frediansyah, A.; Rachmah, D.L. Lactic Acid Bacteria Producing Inhibitor of Alpha Glucosidase Isolated from Ganyong (Canna Edulis) and Kimpul (Xanthosoma sagittifolium). In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2017; p. 012009. [Google Scholar]
- Wang, Z.; Hwang, S.H.; Lee, S.Y.; Lim, S.S. Fermentation of purple Jerusalem artichoke extract to improve the α-glucosidase inhibitory effect in vitro and ameliorate blood glucose in db/db mice. Nutr. Res. Pract. 2016, 10, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Wang, N.; Yin, B.; Fang, D.; Zhao, J.; Zhang, H.; Wang, G.; Chen, W. Lactobacillus plantarum X1 with α-glucosidase inhibitory activity ameliorates type 2 diabetes in mice. RSC Adv. 2016, 6, 63536–63547. [Google Scholar] [CrossRef]
- Mazlan, F.A.; Annuar, M.S.M.; Sharifuddin, Y. Biotransformation of Momordica charantia fresh juice by Lactobacillus plantarum BET003 and its putative antidiabetic potential. PeerJ 2015, 3, e1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar-Zarate, P.; Cruz, M.A.; Montañez, J.; Rodriguez-Herrera, R.; Wong-Paz, J.E.; Belmares, R.E.; Aguilar, C.N. Gallic acid production under anaerobic submerged fermentation by two bacilli strains. Microb. Cell Factories 2015, 14, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnaveni, M.; Dhanalakshmi, R. Qualitative and quantitative study of phytochemicals in Muntingia calabura L. leaf and fruit. World J. Pharm. Res. 2014, 3, 1687–1696. [Google Scholar]
- Degrain, A.; Manhivi, V.; Remize, F.; Garcia, C.; Sivakumar, D. Effect of lactic acid fermentation on color, phenolic compounds and antioxidant activity in African nightshade. Microorganisms 2020, 8, 1324. [Google Scholar] [CrossRef] [PubMed]
- Zha, J.; Koffas, M.A. Production of anthocyanins in metabolically engineered microorganisms: Current status and perspectives. Synth. Syst. Biotechnol. 2017, 24, 259–266. [Google Scholar] [CrossRef]
- Rodríguez, H.; Landete, J.M.; de las Rivas, B.; Muñoz, R.J.F.C. Metabolism of food phenolic acids by Lactobacillus plantarum CECT 748T. Food Chem. 2008, 107, 1393–1398. [Google Scholar] [CrossRef] [Green Version]
- Zha, J.; Wu, X.; Gong, G.; Koffas, M.A. Pathway enzyme engineering for flavonoid production in recombinant microbes. Metab. Eng. Commun. 2019, 9, e00104. [Google Scholar] [CrossRef]
- Adefegha, S.A.; Oboh, G.; Ejakpovi, I.I.; Oyeleye, S.I. Antioxidant and antidiabetic effects of gallic and protocatechuic acids: A structure–function perspective. Comp. Clin. Pathol. 2015, 24, 1579–1585. [Google Scholar] [CrossRef]
- Al-Salih, R.M. Clinical experimental evidence: Synergistic effect of Gallic acid and tannic acid as Antidiabetic and antioxidant agents. Thi-Qar Med. J. 2010, 4, 109–119. [Google Scholar]
- Punithavathi, V.R.; Prince, P.S.M.; Kumar, R.; Selvakumari, J. Antihyperglycaemic, antilipid peroxidative and antioxidant effects of gallic acid on streptozotocin induced diabetic Wistar rats. Eur. J. Pharmacol. 2011, 650, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Motevaseli, E.; Shirzad, M.; Raoofian, R.; Hasheminasab, S.-M.; Hatami, M.; Dianatpour, M.; Modarressi, M.-H. Differences in vaginal lactobacilli composition of Iranian healthy and bacterial vaginosis infected women: A comparative analysis of their cytotoxic effects with commercial vaginal probiotics. Iran. Red Crescent Med. J. 2013, 15, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosem, N.; Han, Y.-H.; Moongkarndi, P. Antioxidant and cytoprotective activities of methanolic extract from Garcinia mangostana hulls. Sci. Asia 2007, 33, 283–292. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Cecchini, S.; Fazio, F. Assessment of Total Antioxidant Capacity in Serum of Heathy and Stressed Hens. Animals 2020, 10, 2019. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Zulueta, A.; Esteve, M.J.; Frígola, A. ORAC and TEAC assays comparison to measure the antioxidant capacity of food products. Food Chem. 2009, 114, 310–316. [Google Scholar] [CrossRef]
- Telagari, M.; Hullatti, K. In-Vitro α-amylase and α-glucosidase inhibitory activity of Adiantum caudatum Linn and Celosia argentea Linn extracts and fractions. Indian J. Pharmacol. 2015, 47, 425. [Google Scholar]
- Saul, R.; Molyneux, R.; Elbein, A. Studies on the mechanism of castanospermine inhibition of α-and β-glucosidases. Arch. Biochem. Biophys. 1984, 230, 668–675. [Google Scholar] [CrossRef]
- Warren, F.J.; Zhang, B.; Waltzer, G.; Gidley, M.J.; Dhital, S. The interplay of α-amylase and amyloglucosidase activities on the digestion of starch in in vitro enzymic systems. Carbohydr. Polym. 2015, 117, 192–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Putative Compound | Experiment (m/z) | Theoretical (m/z) | Adduct | Error (ppm) | rdb |
|---|---|---|---|---|---|---|
| A | gallic acid | 169.0141 | 169.0142 | [M-H]− | 0.7 | 5 |
| B | dihydrokaempferol | 287.0561 | 287.0561 | [M-H]− | −0.1 | 10 |
| C | 5,7-dihydroxyflavone | 253.0505 | 23.0506 | [M-H]− | 0.5 | 11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frediansyah, A.; Romadhoni, F.; Suryani; Nurhayati, R.; Wibowo, A.T. Fermentation of Jamaican Cherries Juice Using Lactobacillus plantarum Elevates Antioxidant Potential and Inhibitory Activity against Type II Diabetes-Related Enzymes. Molecules 2021, 26, 2868. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26102868
Frediansyah A, Romadhoni F, Suryani, Nurhayati R, Wibowo AT. Fermentation of Jamaican Cherries Juice Using Lactobacillus plantarum Elevates Antioxidant Potential and Inhibitory Activity against Type II Diabetes-Related Enzymes. Molecules. 2021; 26(10):2868. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26102868
Chicago/Turabian StyleFrediansyah, Andri, Fitrio Romadhoni, Suryani, Rifa Nurhayati, and Anjar Tri Wibowo. 2021. "Fermentation of Jamaican Cherries Juice Using Lactobacillus plantarum Elevates Antioxidant Potential and Inhibitory Activity against Type II Diabetes-Related Enzymes" Molecules 26, no. 10: 2868. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26102868