
G-Protein Coupled Receptors (GPCRs) in Insects—A Potential Target for New Insecticide Development



1
Department of Entomology and Plant Pathology, Auburn University, Auburn, AL 36849, USA
2
College of Fisheries, Ocean University of China, Qingdao 266100, China
*
Author to whom correspondence should be addressed.
Molecules 2021, 26(10), 2993; https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26102993
Submission received: 8 April 2021
/
Revised: 10 May 2021
/
Accepted: 12 May 2021
/
Published: 18 May 2021
(This article belongs to the Special Issue G Protein-Coupled Receptors and Transporters in the CNS as Drug Targets)
Abstract
:G-protein coupled receptors (GPCRs) play important roles in cell biology and insects’ physiological processes, toxicological response and the development of insecticide resistance. New information on genome sequences, proteomic and transcriptome analysis and expression patterns of GPCRs in organs such as the central nervous system in different organisms has shown the importance of these signaling regulatory GPCRs and their impact on vital cell functions. Our growing understanding of the role played by GPCRs at the cellular, genome, transcriptome and tissue levels is now being utilized to develop new targets that will sidestep many of the problems currently hindering human disease control and insect pest management. This article reviews recent work on the expression and function of GPCRs in insects, focusing on the molecular complexes governing the insect physiology and development of insecticide resistance and examining the genome information for GPCRs in two medically important insects, mosquitoes and house flies, and their orthologs in the model insect species Drosophila melanogaster. The tissue specific distribution and expression of the insect GPCRs is discussed, along with fresh insights into practical aspects of insect physiology and toxicology that could be fundamental for efforts to develop new, more effective, strategies for pest control and resistance management.
1. Introduction
G-protein-coupled receptors (GPCRs), which are proteins that share a seven α-helical transmembrane structure, govern a number of biological and physiological processes in both the vertebrate and invertebrate kingdoms. The main function of GPCRs is to transduce extracellular and environmental signals and regulate intracellular second messengers through coupling to heterotrimeric G-proteins and their downstream effectors [1]. GPCRs are known to be involved in recognizing extracellular messengers, transducing signals to the cytosol, and mediating the cellular responses necessary for the normal physiological functions of organisms [2,3,4,5,6,7]. GPCR binding to a wide variety of molecules (ligands), including hormones, neurotransmitters, ions, photons, odorants, neuropeptides and other stimuli through coupling with G proteins and arrestins [1], results in interactions with G proteins that, in turn, activate the downstream effectors of GPCR pathways, including the adenylate cyclase/cyclic AMP (AC/cAMP), phosphatidylinositol/diacylglycerol/protein kinase C (PI/DAG/PKC) and voltage gated calcium channel (Ca2+ channels) pathways, ultimately playing an indispensable role in the organism’s growth, development, reproduction and other physiological activities [6,7,8,9,10]. These critical functions mean that a better understanding of the role played by GPCRs in cell physiology and biochemistry is crucial for efforts to develop new molecular-level human disease therapies [10,11,12,13,14,15,16].
In just a few short years, GPCR research in insects has progressed from the initial GPCR gene identification to comprehensive bioinformatics analyses, from single GPCR gene analysis to whole genome sequences of GPCRs and explorations of their pathways, and from traditional transcriptional analysis of the gene expression to gene functional characterization of the GPCR genes in insect physiology and cellular biology. The incredible progress being made in related fields includes a wide range of complementary technologies, including bioinformatics and quantitative expression analyses, with functional studies using RNA interferon revealing potential biological functions that significantly impact insect physiology [17,18], including reproduction [19,20,21,22,23,24], regulating growth and development [21,25,26,27,28,29,30,31,32,33,34,35], the stress response [36,37,38,39,40,41,42,43,44,45,46,47], feeding [21,35,48,49,50,51,52,53,54,55,56], general behaviors [20,57,58,59,60,61,62] and other physiological processes [39,63,64,65,66,67,68,69,70]. In addition, the overexpression of GPCRs in insecticide resistance has been reported in both mosquitoes [42,43,44], and house flies [71,72]. The functions of GPCR regulatory pathways and GPCR downstream intracellular cascades have been explored in the development of insecticide resistance in Culex quinquefasciatus [42,43,44], providing a good understanding of the complex molecular processes that govern the development of insecticide resistance through the GPCR signally transduction pathways. The outcomes of these physiological and toxicological studies not only provide us with a clear global picture that is enabling us to develop a clearer understanding of the highly complex mechanisms, genes and pathways involved in these insect physiological and toxicological processes, but are also providing a strong foundation that will allow us to develop tightly GPCR targeted new insecticides and/or environmentally sound insecticides for better vector control that circumvent the problems associated with resistance, representing a highly practical application of scientific research in this area. The findings reported in the most recent studies of GPCR functions in insect are opening up promising new avenues that will undoubtedly revolutionize future research on insect pest management.
This review article examines our current knowledge of the genomic information of the GPCRs in several important insect species and explores the general relationships between and functions of GPCRs in insect physiology. This genome sequencing and annotating information is essential for efforts to build a strong foundation that will enable us to pinpoint the molecular mechanisms and functions of GPCRs in insect physiology and molecular biology in general. We also discuss the distribution and location of GPCRs in various insect tissues where GPCRs are thought to play critical roles in the regulation of insect physiological processes. Since GPCRs are being considered as potential new targets for novel insecticides, understanding the GPCR expression locations and functions could provide valuable insights for those working on developing new insecticides for more effective insect pest control, especially in resistant populations, by identifying possible targets for new chemical and/or insecticide approaches.
2. Whole Genome Sequencing and Transcriptome Analysis—Sequence Comparison and GPCR Characterization in Insects
Recent developments in high-throughput sequencing (HTS) technologies have created powerful analytic tools that enable researchers to study the complex gene interactions in individual organisms and the diverse relationships between organisms. Whole-genome analysis and transcriptome gene expression have revolutionized efforts to identify and annotate insect GPCRs in many different insect species, with comparative genomics of insect populations being used to identify novel targets for insect pest control. Given the growing problem of the development of resistance to the insecticides that are conventionally used to control insects, especially those that are medically, agriculturally and economically important [73], there is an urgent need to shift from traditional chemical pesticides towards more targeted gene-centric approaches. Indeed, genome resequencing analysis followed by functional characterization has opened up many opportunities for discovering new targets, such as GPCRs, for new insecticides that not only control insect pests effectively but also overcome the problem of insecticide resistance. This section reviews GPCRs and their possible functions for several insect species at the whole genome level.
2.1. Classification Systems Used in Characterizing GPCRs
In general, two classification systems are used to characterize GPCRs in organisms, namely the A-F system [74,75] and GRAFS [76]. The A-F system classifies GPCRs primarily in terms of their sequences and functional similarities using six classes, labelled A, B, C, D, E and F. Class A, known as the “rhodopsin-like family”, is the largest group of GPCRs; Class B is the “secretin receptor family”; Class C contains the metabotropic glutamate family; Class D refers to the fungal mating pheromone receptors; Class E includes all the cAMP receptors; Class F are the frizzled/smoothened receptors [77]. The GRAFS system is based on the phylogenetic tree of the human GPCR sequences, with GPCRs being classified into five families: glutamate (G), rhodopsin (R), adhesion (A), frizzled/taste 2 (F) and secretin (S) [76]. In insects, the classification of GPCRs is based on the A-F system. Work on insect GPDRs was revolutionized by the publication of first insect genome sequence of Drosophila melanogaster, an important model organism [78]. To date, more than 100 insect genomes have been sequenced and published, including Anopheles gambiae [79], Aedes aegypti [80], Cx. quinquefasciatus [81], Musca domestica [82] and some other species [83]. Sequencing and annotating these insect genomes provide a strong foundation for future research and new insights that enrich our understanding of the potential functions of GPCRs in insect physiology and molecular biology.
As an important model species in the insect kingdom, D. melanogaster has 200 GPCR genes, over 70 of which belong to Class A (the rhodopsin-like family), 20 to Class B (the secretin-like family), 5 to Class C (the metabotropic glutamate-like family) and 5 to Class F (the frizzled/smoothened family); the remainder have not yet been clearly classified [84,85]. A homology-based bioinformatics analysis conducted on the genome of the mosquito An. gambiae revealed 276 GPCR genes, of which over 80 belong to Class A (the rhodopsin-like family), 20 to Class B (the secretin-like family), 8 to Class C (the metabotropic glutamate-like family), 8 belong to Class F (frizzled/smoothened) and the remainder were other GPCRs [86]. Several GPCRs were explored via reannotation in the mosquito Ae. aegypti, coding over 135 GPCR genes, with 89 belonging to Class A (the rhodopsin-like family), 24 to Class B (the secretin-like family), 8 to Class C (the metabotropic glutamate-like family), 11 belonging to Class F (frizzled/smoothened family) and the rest not been clearly classified [80]. Around 90 GPCRs from the silkworm Bombyx mori have also been annotated by bioinformatics analysis, with 69 being classified as Class A (the rhodopsin-like family), 8 as Class B (the secretin-like family), 9 as Class C (the metabotropic glutamate-like family), 3 as Class F (the frizzled/smoothened family) and others [87]. For Apis mellifera, around 50 genes have been coded for GPCR, of which 31 belong to Class A (the rhodopsin-like family) and 4 to Class B (the secretin-like family) [17]. For Cx. quinquefasciatus, around 115 GPCR genes have been identified, with 52 belonging to Class A (the rhodopsin-like family), 4 to Class B (the secretin-like family) and the rest not been classified [42]. For Musca domestica, around 94 genes have been identified as GPCR genes, 55 of which are classified as Class A (the rhodopsin-like family), 27 are classified as Class B (the secretin-like family), 4 are classified as Class C (the metabotropic glutamate-like family) and the rest are still unclear [72] (Table 1, Figure 1).
In addition to the insects above that have been extensively studied, GPCRs from several other insect species have also been identified and annotated. The genome of the blowfly, Lucilia cuprina, has also been characterized by typical genomic sequencing, RNA-seq and the assembly method. The subsequent genome sequence and gene/protein identification and annotation revealed 197 GPCRs, including 73 to be in Class A (the rhodopsin-like family), 18 in Class B (the secretin-like family) and 9 in Class C (the metabotropic glutamate receptor family), along with some unclassified proteins [88] (Figure 1, Table 1). In recent work, a transcriptome study on GPCRs in the stick insect, Carausius morosus, conducted via RNA-seq and de novo RNA assembly revealed 430 putative GPCR genes [89]; a transcriptome analysis predicted over 300 transcripts for coding GPCRs in fire ants [90] and 65 genes for putative neuropeptide GPCRs were identified in the spider mite [91].
Generally, Class A GPCRs are the predominant class of GPCRs in insects, comprising their neuropeptide and protein hormone receptors, muscarinic acetylcholine receptors, dopamine receptors, 5-hydroxytryptamine receptors, tyramine receptors, opsin receptors, vasopressin receptors and orphan receptors, among others [92,93]. These perform a variety of have different functions [94,95,96], including metamorphosis (e.g., dopamine receptors) [29,34,35,97,98], feeding modulation (e.g., allatostatin receptors) [99,100], behavioral changes (e.g., sex peptide receptors and SIF amide neuropeptide receptors) [101,102,103,104], metabolism regulation (e.g., CCHamide-2 receptors and adipokinetic hormone receptor) [51,53,62,105,106] and visual photoreceptors (e.g., rhodopsin proteins) [107], among many others [92,95,108,109,110]. The Class B GPCRs, which include diuretic hormone receptors, methuselah-type receptors and others [111,112], mainly focus on the insect’s water balance (e.g., corticotropin-releasing factor receptors) [113], body temperature modulation (e.g., diuretic hormone receptors) [114], stress response (e.g., methuselah GPCRs) [115,116] and other functions [117]. Their Class C receptors are mainly composed of metabotropic GABA receptors, which serve as neurotransmitters receptors and are involved in signal transportation [118]. The frizzled receptors and smoothened-type receptors, which are mainly Class F GPCRs, focus on Wnt signaling [119,120,121] and the hedgehog signaling pathway, respectively [122,123] (Figure 1). Both the Wnt signaling pathway and the hedgehog pathway are important for insect development [124,125].
2.2. Receptors of GPCR Involved in Insect Physiology or Insecticide Resistance That Are Potential Targets for Insecticide Development
Several GPCRs have been reported to be involved in different functions in insect physiology as described below (Table 2) and could thus be potential targets for new insecticides. The majority are Class A GPCRs, including the dopamine receptor, which mainly regulates sexual activity [60,126], morphogenesis [29,34,35,97], mushroom and locomotor activity [127] and ethanol-induced sedation [68]. Neuropeptide and hormone receptors such as the adipokinetic hormone receptor (AKHR) can receive signals from the adipokinetic hormone and regulate lipid mobilization [51,53,62,106], while the allatostatin receptor (AstR) regulates the juvenile hormone synthesis [26,128], the diapause hormone receptor (DHR) is involved in insect development [39,41,45] and the neuropeptide receptors focus on the regulation of insect feeding behavior [50,129] and ecdysone synthesis [33]. Orphan receptor, like BNGR in B. mori regulates the insect’s food intake and growth [130], and DLGR2 in D. melanogaster regulates the insect’s bursicon bioactivity [25]. Tyramine receptors are responsible for the development of antiparasitic compounds [131], while calcitonin receptors regulate hindgut contraction and primary urine secretion [66] and the 5-HT (5-hydroxytryptamine) receptors and muscarinie acetylcholine receptors are important for the insect’s neural processes [132,133,134]. Rhodopsin receptors perform vital functions in both the insect’s reproduction system [22] and its vision [107]. Class B GPCRs such as methuselah receptors are also involved in insect longevity [135], and oxidative stress resistance [136] and diuretic hormone receptors regulate the body temperature and homeostasis [114]; GPCRs in the Class C family, such as metabotropic GABA receptors, are important for the central nervous system (CNS) [118].
In addition to the vital functions they perform in the insect’s physiology, GPCRs are also involved in insecticide resistance. The methuselah-like receptor in L. dispar [135], the calcitonin receptor and opsin receptor in Cx. quinquefasciatus [42] and the arrestin gene in Culex pipiens [137] have all been shown to be involved in the upregulation of detoxified enzymes such as cytochrome P450. The leucokinin receptor, opsin receptor, methuselah-like receptors and dopamine receptors have also exhibited higher levels of expression in resistant housefly strains compared to susceptible strains [72]. Octopamine and tyramine receptors are also known to be involved in Amitraz resistance in ticks based on the results of an SNP analysis [138].
Chemicals or techniques that target these receptors and destroy their function could serve as very effective insecticides. For example, chemicals that act on dopamine receptors like amitriptyline and doxepin have been shown to cause the death of both Ae. aegypti [139,140] and Cx. quinquefasciatus [141], while an RNAi technique that targets the dopamine 1 receptor also led to the death of both larvae and adult in Ae. aegypti [142]. Many more chemicals could be investigated to target various GPCR receptors and improve insect management (Table 3).
3. Tissue Specific Expression Analysis of GPCRs in Insects
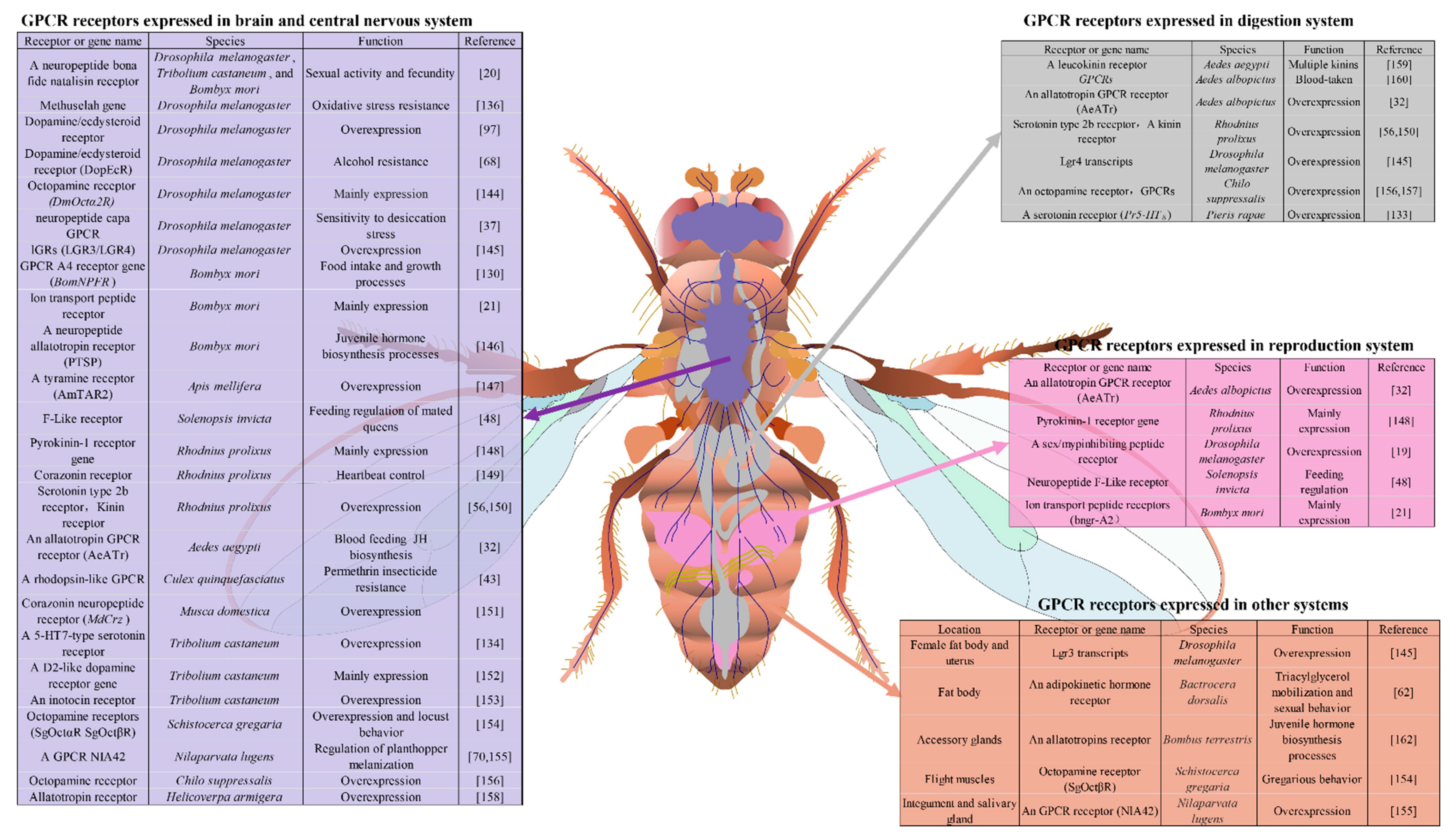
Tissue specific analyses of gene expression usually provide new insights into potential physiological mechanisms and correlates the functions of the genes with the functions of specific tissues. GPCRs are distributed in various insect tissues, where GPCRs and their ligands play critical roles in the regulation of insect physiological processes. Since the focus here is on considering GPCRs as new targets of novel insecticides, understanding the GPCR expression locations and functions could provide valuable insights and contributions for new insecticide development. Discovering and identifying the GPCR gene profiles in different tissues will help broaden our understanding of the functions of various types of tissue and gene functions, and the biological mechanisms involved [143]. The aberrant expression of tissue specific expressed GPCRs may also be implicated in various abnormal functions of the insects, and hence important for the control of insect pests. This section reviews recent discoveries related to the GPCR genes present in insect nervous, digestive and reproductive systems, and in others, such as fat bodies, muscles and salivary glands (Figure 2).
3.1. Brain Tissue and Central Nervous System
Several GPCRs have been identified as being predominately expressed in the brain and CNS, corresponding to neuropeptide expression in diverse insect species. A neuropeptide bona fide natalisin receptor found to be highly expressed in the CNS of D. melanogaster, T. castaneum and B. mori regulates sexual activity and fecundity in insects [20]. A mutant methuselah gene expressed in the brain of Drosophila, a model insect often used to investigate gene functions, plays a critical function in oxidative stress resistance [136]; a dopamine/ecdysteroid receptor that is a head-specific expression gene is also overexpressed in Drosophila adults and embryos [97]. Another study also found the dopamine/ecdysteroid receptor (DopEcR) to be highly expressed in Drosophila nervous system and brain tissues, and its mutant has been implicated in the insect’s resistance to alcohol [68]. A Drosophila sex/myp-inhibiting peptide receptor expressed in the CNS is highly conserved in insects [19], while an octopamine receptor DmOctα2R transcript present at much higher levels in Drosophila males than in females is mainly expressed in the head of adults [144] and a Drosophila neuropeptide capa GPCR expressed in larvae central nerve system is responsible for sensitivity to desiccation stress [37]. Of two lGRs (LGR3/LGR4) characterized in diverse Drosophila tissues, Lgr3 transcripts were predominately expressed in the CNS, while Lgr4 transcripts were highly expressed in the CNS of the wandering larval stage. In adults, the Lgr4 transcripts showed significantly high expression in the male thoracic-abdominal ganglion and brain tissues [145].
The silkworm, B. mori, which is classed as a beneficial insect, is another model species frequently used in gene functional studies. A neuropeptide GPCR A4 receptor gene (BomNPFR) amplified from its brain tissue was predicted to be involved in food intake and growth processes [130]. Three GPCR receptors responding to neuron ion transport peptides (ITPs) were identified in silkworm larvae using tissue specific expression, revealing that bngr-A2 was mainly expressed in the brain, with no expression in the CNS, and bngr-A34 was not present in nervous tissues [21]. A neuropeptide allatotropin receptor was significantly overexpressed in the corpora cardiaca and brain of B. mori, indicating the potential involvement of juvenile hormone (JH) biosynthesis processes [146]. In honeybees, an important beneficial insect that plays a critical role in pollination, an octopamine receptor was identified in the honeybee brain [63]. A tyramine receptor was later identified that was highly expressed in nurse bees and forager brain neuropils [147].
The red import fire ant, Solenopsis invicta, is an invasive urban insect species whose gene expression and functions have been widely studied over the last couple of decades. Ten GPCR gene expressions have exhibited significant differences in the brain tissue of workers, alate virgin queens, and mated queens [90]. One neuropeptide F-Like receptor was isolated from S. invicta and its differential expression levels in the brain suggest its potential function to be feeding regulation for mated queens [48]. In the medically important kissing bug, Rhodnius prolixus, an important Chagas disease vector, GPCRs have been found to be involved in multiple physiological pathways. Three variants of a pyrokinin-1 receptor gene characterized in different tissues of fifth instar larvae of R. prolixus revealed two variants that were mainly expressed in the CNS [148]. A corazonin receptor was found to be overexpressed in the brain and involved in R. prolixus heartbeat control [149] and a serotonin type 2b receptor transcript has also been shown to be upregulated in the CNS of R. prolixus [150]. To predict the potential function of a kinin receptor in R. prolixus blood post-feeding, a kinin receptor was characterized in various tissues and found to be overexpressed in the CNS of late instar larvae [56]. In mosquitoes, which are responsible for transmitting a number of human and animal diseases, GPCRs have been identified as being involved in mosquito blood feeding and insecticide resistance. The overexpression of an allatotropin GPCR receptor (AeATr) gene was characterized in the nervous system and corpora alata-corpara cardiac complex of Ae. aegypti. Blood feeding depressed the transcript level of AeATr, and was associated with JH biosynthesis in mosquitoes [32]. A rhodopsin-like GPCR overexpressed in the head of adult mosquitoes was found to play a critical role in the development of permethrin insecticide resistance in the mosquito, Cx. Quinquefasciatus [43]. Tissue specific studies conducted on this GPCR gene revealed its significantly high expression in the insect’s brain tissue along with a G-protein alpha subunit, two adenylyl cyclase and one protein kinase A gene, playing important roles in the GPCR-leading intracellular pathway in the insect’s neuron system and regulating its insecticide resistance [44]. A corazonin neuropeptide receptor (MdCrz) has been found to be overexpressed in the larval CNS of the house fly, another medically important pest, including in its ventral nerve cord, the protocerebral DL neurons of the brain lobes and its vCrz neurons. Although it was overexpressed in the heads of both male and female houseflies, there was no expression in other body tissues, indicating that MdCrz is a brain-specific expression gene [151].
The red flour beetle, T. castaneum, is a stored product pest that causes problem for communities around the world. The spatial expression of a 5-HT7-type serotonin receptor identified in T. castaneum showed the highest expression levels to be in the head of both male and female adults, predominantly expressed in the brain but accompanied by high levels of expression in the optic lobes, predicting the functional importance of this receptor in neural processes [134]. A D2-like dopamine receptor gene predominantly expressed in the head and CNS of T. castnaeum adults [152] and an inotocin receptor was found to be overexpressed in the head of the insect’s early larval stage [153]. In the desert locust, S. gregaria, an agricultural pest species, two novel octopamine receptors were characterized in adults, with SgOctαR being overexpressed in the CNS, including the brain, optic lobes, subesophageal ganglion and thoracic ganglions, while SgOctβR was overexpressed in the CNS. Interestingly, these receptors were highly expressed in the long-term gregarious locusts but not in the solitarious locusts, suggesting the receptors’ function in locust behavior [154]. GPCRs have also been widely studied in other agricultural pest species. The GPCR NIA42 was found to be highly expressed in the brain and abdominal integuments of the adult brown planthopper, N. lugens, where it was linked to the neuropeptide NI-elevenin that is involved in the regulation of planthopper melanization [70,155]. A novel octopamine receptor gene expression has been characterized in diverse tissues of the fifth-instar larva rice stem borer, Chilo suppressalis, showing a high level of expression in the nerve cord [156]. A large screen of GPCRs that was characterized in C. suppressalis identified 51 putative GPCR genes. The expression of these genes was examined in tissues of the insect’s CNS, fat body, gut and hemocytes, with most of the receptor genes being highly expressed in the CNS [157]. A serotonin receptor (Pr5-HT8) was identified as being highly expressed in the nerve cord of the larva small white butterfly, Pieris rapae. 5-hydroxytryptamine (5-HT) has been identified as a neurotransmitter that plays critical roles in the regulation of a number of physiological processes, which is consistent with the expression of Pr5-HT8 in the butterfly’s CNS [133]. An allatotropin receptor has also been found to be overexpressed in the brain, thoracic ganglion and abdomen ganglion of the bollworm, H. armigera [158].
3.2. Digestion and Reproduction Systems
In mosquito species, GPCRs are known to be expressed in the digestion system and involved in blood-feeding behavior. A leucokinin receptor has diverse functions, responding to multiple kinins in the mosquito, Ae. aegypti, and is expressed in the mosquito’s hindgut and Malpighian tubules [159]. Piermarini’s group also identified several GPCRs and metabolic genes upregulated or downregulated in the Malpighian tubules of blood-taken Aedes albopictus [160]. An allatotropin GPCR receptor (AeATr) gene has also been found to be overexpressed in the ovary of adult mosquitoes [32]. In another blood-feeding insect, R. prolixus, a serotonin type 2b receptor transcript was overexpressed in the Malpighian tubules, salivary glands and guts [150]. A kinin receptor has also been found to be highly overexpressed in the gut system of late instar larvae of R. prolixus [56]. Two variants of a pyrokinin-1 receptor gene were mainly expressed in the male testes and prothoracic glands of fifth instar larvae of R. prolixus [148]. In Drosophila, several GPCRs were identified as overexpressed in the digestion and reproduction systems. These Drosophila Lgr4 transcripts were highly expressed in the gut system of the wandering larval stage, and significantly highly expressed in the male midgut and crop [145]. Additionally, a sex/myp-inhibiting peptide receptor was highly expressed in the male Drosophila reproduction organs [19] and an octopamine receptor gene was highly expressed in the Malpighian tubules, with lesser amounts found in the midgut and hemocytes of the fifth-instar larva rice stem borer, C. suppressalis [156]. Several GPCRs in C. suppressalis were overexpressed in the gut system [157]. A neuropeptide F-Like receptor detected in the gut and reproduction systems of S. invicta suggests its potential function was involved in the feeding regulation of mated queens [48]. One ITP gene identified in B. mori, bngr-A2, was found to be mainly expressed in the reproduction system [21]. A serotonin receptor (Pr5-HT8) is known to be highly expressed in the Malpighian tubules, fat body and midgut of larvae of the small white butterfly, P. rapae [133].
3.3. Other Insect Organs
In addition to the GPCRs identified in the CNS, digestive and reproductive systems in insect species, many GPCRs have also been characterized in other organs. In Drosophila adults, Lgr3 transcripts were found to be highly expressed in the female fat body and uterus, and the male salivary glands [145]. An adipokinetic hormone receptor was predominately expressed in the fat body of the oriental fruit fly, B. dorsalis, and was also involved in its triacylglycerol mobilization and sexual behavior [62]. Forty six putative GPCR transcripts have been isolated from the foreleg tissues of the cattle tick, Phipicephalus australis, providing valuable information for GPCR studies of signal transduction, host preference and mating behavior in insects [161]. An allatotropin receptor has been identified that is overexpressed in the male bumblebee accessory glands, predicting its potential involvement in JH biosynthesis [162] and in B. mori a sex peptide receptor has been found to be highly expressed in the prothoracic gland, predicting the critical roles of receptor in regulating ecdysteroidogenesis [28]. A novel octopamine receptor, SgOctβR, was also overexpressed in the flight muscles of S. gregaria, suggesting its involvement in locust gregarious behavior [154], while a GPCR receptor, NlA42, was significantly highly expressed in the integument and salivary gland of the brown planthopper indicating its function in the insect’s melanization [155].
![Molecules 26 02993 g002]()
Figure 2.
Tissue specific expression of GPCR genes in insect species. Insect GPCRs are mainly expressed in the brain and central nervous system, highlighted in light blue; those in the digestive system are highlighted in gray; those in the reproductive system are highlighted win light purple; those in other organs are highlighted in orange. Twenty-six GPCRs that are highly expressed in the brain and central nervous system in 13 insect species are involved in sexual activity and fecundity, oxidative stress resistance, desiccation stress, food intake and growth, juvenile hormone biosynthesis, feeding regulation, heartbeat control, permethrin insecticide resistance, behavior and cuticle melanization.
Figure 2.
Tissue specific expression of GPCR genes in insect species. Insect GPCRs are mainly expressed in the brain and central nervous system, highlighted in light blue; those in the digestive system are highlighted in gray; those in the reproductive system are highlighted win light purple; those in other organs are highlighted in orange. Twenty-six GPCRs that are highly expressed in the brain and central nervous system in 13 insect species are involved in sexual activity and fecundity, oxidative stress resistance, desiccation stress, food intake and growth, juvenile hormone biosynthesis, feeding regulation, heartbeat control, permethrin insecticide resistance, behavior and cuticle melanization.

However, while the precise functions of the GPCRs in specific tissues are still in need of further characterization, the impact of the GPCRs on insect behaviors can now be predicted to some extent based on the functions of those specific tissues in insects. The importance of GPCRs in insects can be partially appreciated by considering their distribution, location and function within the cells of specific tissues. In humans, if any of the fundamental processes governed by GPCRs in specific tissues are dysfunctional, this will lead to acute or chronic diseases [163,164,165,166]. The physical location and expression of GPCRs may provide a direct mechanism for the transduction of extracellular messages into tissue responses and functions. Consequently, the identification of the GPCRs in brains and CNS of insects could shed new light on the important roles they play in neuronal firing, the regulation of ion transportation across cell membranes and the modulation of neuron membrane potentials in other species.
4. Conclusions
The biological and medical importance of GPCRs has been well characterized and is now widely recognized, making GPCR an important focus for drug discovery. Yet, while none or few insecticides have been developed that specifically target GPCRs in insects, recent progress on identifying the GPCR genome sequences in insects, the specific expression of the GPCRs in different tissues and the crucial role of GPCRs for insect physiologic processes and toxicology promise to provide fresh insights into GPCRs’ biochemical functions in insects and assist in the development of new insect-specific insecticides. The value of the findings summarized in this review of the current status of research into GPCR in insects, including information on their sequences, functions in physiology and toxicology and the tissues and organs that they are involved in, are fundamental for understanding how these GPCR systems function to modulate a broad spectrum of cellular activities. A recent review on insect GPCRs uncovered GPCR signaling pathways, functions in insect physiology and toxicology and the latest exciting technological advances and new techniques for gene expression and functional of the GPCRs in insects [167]. Together, these reviews on insect GPCRs will help researchers in the field develop new insect-specific insecticides that will help control insect pests in the future.
Author Contributions
N.L., T.L., Y.W. and S.L. wrote the paper and reviewed the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by AAES Hatch/Multistate Grants ALA08-045 and ALA015-1-10026, and ALA015-1-16009 to N.L.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hilger, D.; Masureel, M.; Kobilka, B.K. Structure and dynamics of GPCR signaling complexes. Nat. Struct. Mol. Biol. 2018, 25, 4–12. [Google Scholar] [CrossRef]
- Bockaert, J.; Pin, J.P. Molecular tinkering of G protein-coupled receptors: An evolutionary success. EMBO J. 1999, 18, 1723–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maudsley, S.; Martin, B.; Luttrell, L.M. The Origins of Diversity and Specificity in G Protein-Coupled Receptor Signaling. J. Pharmacol. Exp. Ther. 2005, 314, 485–494. [Google Scholar] [CrossRef] [Green Version]
- Lagerström, M.C.; Schiöth, H.B. Structural diversity of G protein-coupled receptors and significance for drug discovery. Nat. Rev. Drug Discov. 2008, 7, 339–357. [Google Scholar] [CrossRef]
- Spehr, M.; Munger, S.D. Olfactory receptors: G protein-coupled receptors and beyond. J. Neurochem. 2009, 109, 1570–1583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millar, R.P.; Newton, C.L. The Year in G Protein-Coupled Receptor Research. Mol. Endocrinol. 2010, 24, 261–274. [Google Scholar] [CrossRef]
- Eglen, R.M.; Reisine, T. GPCRs Revisited: New Insights Lead to Novel Drugs. Pharmaceuticals 2011, 4, 244–272. [Google Scholar] [CrossRef] [Green Version]
- Gether, U. Uncovering Molecular Mechanisms Involved in Activation of G Protein-Coupled Receptors. Endocr. Rev. 2000, 21, 90–113. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, Z.G.; Dhanasekaran, D.N. G Protein regulation of MAPK networks. Oncogene 2007, 26, 3122–3142. [Google Scholar] [CrossRef] [Green Version]
- Gerald, W.Z. Calcium channel signaling complexes with receptors and channels. Curr. Mol. Pharmacol. 2015, 8, 8–11. [Google Scholar]
- Flower, D.R. Modelling G-protein-coupled receptors for drug design. Biochim. Biophys. Acta (BBA)-Rev. Biomembr. 1999, 1422, 207–234. [Google Scholar] [CrossRef]
- Hopkins, A.L.; Groom, C.R. The druggable genome. Nat. Rev. Drug Discov. 2002, 1, 727–730. [Google Scholar] [CrossRef] [PubMed]
- Robas, N.; O’Reilly, M.; Katugampola, S.; Fidock, M. Maximizing serendipity: Strategies for identifying ligands for orphan G-protein-coupled receptors. Curr. Opin. Pharmacol. 2003, 3, 121–126. [Google Scholar] [CrossRef]
- Jacoby, E.; Bouhelal, R.; Gerspacher, M.; Seuwen, K. The 7 TM G-Protein-Coupled Receptor Target Family. ChemMedChem 2006, 1, 760–782. [Google Scholar] [CrossRef] [PubMed]
- Hauser, A.S.; Attwood, M.M.; Rask-Andersen, M.; Schiöth, H.B.; Gloriam, D.E. Trends in GPCR drug discovery: New agents, targets and indications. Nat. Rev. Drug Discov. 2017, 16, 829–842. [Google Scholar] [CrossRef] [PubMed]
- Congreve, M.; de Graaf, C.; Swain, N.A.; Tate, C.G. Impact of GPCR Structures on Drug Discovery. Cell 2020, 181, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Hauser, F.; Cazzamali, G.; Williamson, M.; Blenau, W.; Grimmelikhuijzen, C.J. A review of neurohormone GPCRs present in the fruitfly Drosophila melanogaster and the honey bee Apis mellifera. Prog. Neurobiol. 2006, 80, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Hauser, F.; Cazzamali, G.; Williamson, M.; Park, Y.; Li, B.; Tanaka, Y.; Predel, R.; Neupert, S.; Schachtner, J.; Verleyen, P.; et al. A genome-wide inventory of neurohormone GPCRs in the red flour beetle Tribolium castaneum. Front. Neuroendocr. 2008, 29, 142–165. [Google Scholar] [CrossRef]
- Poels, J.; Van Loy, T.; Vandersmissen, H.P.; Van Hiel, B.; Van Soest, S.; Nachman, R.J.; Broeck, J.V. Myoinhibiting peptides are the ancestral ligands of the promiscuous Drosophila sex peptide receptor. Cell. Mol. Life Sci. 2010, 67, 3511–3522. [Google Scholar] [CrossRef]
- Jiang, H.; Lkhagva, A.; Park, Y.; Kim, Y.-J.; Daubnerová, I.; Chae, H.-S.; Šimo, L.; Jung, S.-H.; Yoon, Y.-K.; Lee, N.-R.; et al. Natalisin, a tachykinin-like signaling system, regulates sexual activity and fecundity in insects. Proc. Natl. Acad. Sci. USA 2013, 110, E3526–E3534. [Google Scholar] [CrossRef] [Green Version]
- Nagai, C.; Mabashi-Asazuma, H.; Nagasawa, H.; Nagata, S. Identification and Characterization of Receptors for Ion Transport Peptide (ITP) and ITP-like (ITPL) in the Silkworm Bombyx mori. J. Biol. Chem. 2014, 289, 32166–32177. [Google Scholar] [CrossRef] [Green Version]
- Bai, H.; Palli, S.R. Identification of G protein-coupled receptors required for vitellogenin uptake into the oocytes of the red flour beetle, Tribolium castaneum. Sci. Rep. 2016, 6, 27648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, Y.-P.; An, H.; Zhang, S.; Wang, N.; Zhou, S. Protein kinase C mediates juvenile hormone–dependent phosphorylation of Na+/K+-ATPase to induce ovarian follicular patency for yolk protein uptake. J. Biol. Chem. 2018, 293, 20112–20122. [Google Scholar] [CrossRef] [Green Version]
- Marciniak, P.; Urbański, A.; Lubawy, J.; Szymczak, M.; Pacholska-Bogalska, J.; Chowański, S.; Kuczer, M.; Rosiński, G. Short neuropeptide F signaling regulates functioning of male reproductive system in Tenebrio molitor beetle. J. Comp. Physiol. B 2020, 190, 521–534. [Google Scholar] [CrossRef] [PubMed]
- Mendive, F.M.; Van Loy, T.; Claeysen, S.; Poels, J.; Williamson, M.; Hauser, F.; Grimmelikhuijzen, C.J.; Vassart, G.; Broeck, J.V. Drosophila molting neurohormone bursicon is a heterodimer and the natural agonist of the orphan receptor DLGR2. FEBS Lett. 2005, 579, 2171–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.-J.; Zitnan, D.; Cho, K.-H.; Schooley, D.A.; Mizoguchi, A.; Adams, M.E. Central peptidergic ensembles associated with organization of an innate behavior. Proc. Natl. Acad. Sci. USA 2006, 103, 14211–14216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Žitňan, D.; Kim, Y.-J.; Žitňanová, I.; Roller, L.; Adams, M. Complex steroid–peptide–receptor cascade controls insect ecdysis. Gen. Comp. Endocrinol. 2007, 153, 88–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamanaka, N.; Hua, Y.-J.; Roller, L.; Spalovská-Valachová, I.; Mizoguchi, A.; Kataoka, H.; Tanaka, Y. Bombyx prothoracicostatic peptides activate the sex peptide receptor to regulate ecdysteroid biosynthesis. Proc. Natl. Acad. Sci. USA 2010, 107, 2060–2065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, H.; Zhu, F.; Shah, K.; Palli, S.R. Large-scale RNAi screen of G protein-coupled receptors involved in larval growth, molting and metamorphosis in the red flour beetle. BMC Genom. 2011, 12, 388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Beeman, R.W.; Park, Y. Functions of duplicated genes encoding CCAP receptors in the red flour beetle, Tribolium castaneum. J. Insect Physiol. 2011, 57, 1190–1197. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Jia, Q.; Li, S. Juvenile hormone signaling—A mini review. Insect Sci. 2019, 26, 600–606. [Google Scholar] [CrossRef]
- Nouzova, M.; Brockhoff, A.; Mayoral, J.G.; Goodwin, M.; Meyerhof, W.; Noriega, F.G. Functional characterization of an allatotropin receptor expressed in the corpora allata of Mosquitoes. Peptides 2012, 34, 201–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iga, M.; Nakaoka, T.; Suzuki, Y.; Kataoka, H. Pigment Dispersing Factor Regulates Ecdysone Biosynthesis via Bombyx Neuropeptide G Protein Coupled Receptor-B2 in the Prothoracic Glands of Bombyx mori. PLoS ONE 2014, 9, e103239. [Google Scholar] [CrossRef] [PubMed]
- Regna, K.; Kurshan, P.T.; Harwood, B.N.; Jenkins, A.M.; Lai, C.-Q.; Muskavitch, M.A.; Kopin, A.S.; Draper, I. A critical role for the Drosophila dopamine D1-like receptor Dop1R2 at the onset of metamorphosis. BMC Dev. Biol. 2016, 16, 15. [Google Scholar] [CrossRef] [Green Version]
- Kang, X.-L.; Zhang, J.-Y.; Wang, D.; Zhao, Y.-M.; Han, X.-L.; Wang, J.-X.; Zhao, X.-F. The steroid hormone 20-hydroxyecdysone binds to dopamine receptor to repress lepidopteran insect feeding and promote pupation. PLoS Genet. 2019, 15, e1008331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homma, T.; Watanabe, K.; Tsurumaru, S.; Kataoka, H.; Imai, K.; Kamba, M.; Niimi, T.; Yamashita, O.; Yaginuma, T. G protein-coupled receptor for diapause hormone, an inducer of Bombyx embryonic diapause. Biochem. Biophys. Res. Commun. 2006, 344, 386–393. [Google Scholar] [CrossRef]
- Terhzaz, S.; Cabrero, P.; Robben, J.H.; Radford, J.C.; Hudson, B.D.; Milligan, G.; Dow, J.A.T.; Davies, S.-A. Mechanism and Function of Drosophila capa GPCR: A Desiccation Stress-Responsive Receptor with Functional Homology to Human NeuromedinU Receptor. PLoS ONE 2012, 7, e29897. [Google Scholar] [CrossRef] [Green Version]
- Bryon, A.; Wybouw, N.; Dermauw, W.; Tirry, L.; Van Leeuwen, T. Genome wide gene-expression analysis of facultative reproductive diapause in the two-spotted spider mite Tetranychus urticae. BMC Genom. 2013, 14, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Choi, M.-Y.; Estep, A.; Sanscrainte, N.; Becnel, J.; Meer, R.K.V. Identification and expression of PBAN/diapause hormone and GPCRs from Aedes aegypti. Mol. Cell. Endocrinol. 2013, 375, 113–120. [Google Scholar] [CrossRef]
- Devambez, I.; Agha, M.A.; Mitri, C.; Bockaert, J.; Parmentier, M.-L.; Marion-Poll, F.; Grau, Y.; Soustelle, L. Gαo Is Required for L-Canavanine Detection in Drosophila. PLoS ONE 2013, 8, e63484. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Wei, Z.; Nachman, R.J.; Park, Y. Molecular cloning and functional characterization of the diapause hormone receptor in the corn earworm Helicoverpa zea. Peptides 2014, 53, 243–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Liu, L.; Zhang, L.; Liu, N. Role of G-protein-coupled Receptor-related Genes in Insecticide Resistance of the Mosquito, Culex quinquefasciatus. Sci. Rep. 2015, 4, 6474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Cao, C.; Yang, T.; Zhang, L.; He, L.; Xi, Z.; Bian, G.; Liu, N. A G-protein-coupled receptor regulation pathway in cytochrome P450-mediated permethrin-resistance in Mosquitoes, Culex quinquefasciatus. Sci. Rep. 2015, 5, 17772. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Liu, N. Regulation of P450-mediated permethrin resistance in Culex quinquefasciatus by the GPCR/Gαs/AC/cAMP/PKA signaling cascade. Biochem. Biophys. Rep. 2017, 12, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Jiang, X.; Yan, L.; Chen, Y.; Wang, W.; Shi, Y.; Shi, L.; Liu, D.; Zhou, N. Structural basis for the interaction of diapause hormone with its receptor in the silkworm, Bombyx mori. FASEB J. 2018, 32, 1338–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-H.; Di, Y.-Q.; Shen, Q.-Y.; Wang, J.-X.; Zhao, X.-F. The steroid hormone 20-hydroxyecdysone induces phosphorylation and aggregation of stromal interacting molecule 1 for store-operated calcium entry. J. Biol. Chem. 2019, 294, 14922–14936. [Google Scholar] [CrossRef]
- Petruccelli, E.; Lark, A.; Mrkvicka, J.A.; Kitamoto, T. Significance of DopEcR, a G-protein coupled dopamine/ecdysteroid receptor, in physiological and behavioral response to stressors. J. Neurogenet. 2020, 34, 55–68. [Google Scholar] [CrossRef]
- Chen, M.-E.; Pietrantonio, P.V. The short neuropeptide F-like receptor from the red imported fire ant, Solenopsis invicta Buren (Hymenoptera: Formicidae). Arch. Insect Biochem. Physiol. 2006, 61, 195–208. [Google Scholar] [CrossRef]
- Kersch, C.N.; Pietrantonio, P.V. MosquitoAedes aegypti (L.) leucokinin receptor is critical forin vivofluid excretion post blood feeding. FEBS Lett. 2011, 585, 3507–3512. [Google Scholar] [CrossRef] [Green Version]
- Dillen, S.; Zels, S.; Verlinden, H.; Spit, J.; Van Wielendaele, P.; Broeck, J.V. Functional Characterization of the Short Neuropeptide F Receptor in the Desert Locust, Schistocerca gregaria. PLoS ONE 2013, 8, e53604. [Google Scholar] [CrossRef] [Green Version]
- Baumbach, J.; Xu, Y.; Hehlert, P.; Kühnlein, R.P. Gαq, Gγ1 and Plc21C Control Drosophila Body Fat Storage. J. Genet. Genom. 2014, 41, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Lin, F.; Hossain, M.A.; Post, S.; Karashchuk, G.; Tatar, M.; De Meyts, P.; Wade, J.D. Total Solid-Phase Synthesis of Biologically Active Drosophila Insulin-Like Peptide 2 (DILP2). Aust. J. Chem. 2017, 70, 208–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, K.; Zhang, X.; Chen, X.; Li, Y.; Li, W.; Cheng, Y.; Zhou, J.; You, K.; Zhou, Q. Adipokinetic Hormone Receptor Mediates Lipid Mobilization to Regulate Starvation Resistance in the Brown Planthopper, Nilaparvata lugens. Front. Physiol. 2018, 9, 1730. [Google Scholar] [CrossRef] [Green Version]
- Marchal, E.; Schellens, S.; Monjon, E.; Bruyninckx, E.; Marco, H.G.; Gäde, G.; Broeck, J.V.; Verlinden, H. Analysis of Peptide Ligand Specificity of Different Insect Adipokinetic Hormone Receptors. Int. J. Mol. Sci. 2018, 19, 542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Huang, J.; Liu, Y.; Li, J.; Luo, S.; Wu, J. Prediction of the post-translational modifications of adipokinetic hormone receptors from solitary to eusocial bees. Sociobiology 2018, 65, 271–279. [Google Scholar] [CrossRef]
- Sangha, V.; Lange, A.B.; Orchard, I. Identification and cloning of the kinin receptor in the Chagas disease vector, Rhodnius prolixus. Gen. Comp. Endocrinol. 2020, 289, 113380. [Google Scholar] [CrossRef]
- Bainton, R.J.; Tsai, L.T.-Y.; Schwabe, T.; DeSalvo, M.; Gaul, U.; Heberlein, U. moody Encodes Two GPCRs that Regulate Cocaine Behaviors and Blood-Brain Barrier Permeability in Drosophila. Cell 2005, 123, 145–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyun, S.; Lee, Y.; Hong, S.-T.; Bang, S.; Paik, D.; Kang, J.; Shin, J.; Lee, J.; Jeon, K.; Hwang, S.; et al. Drosophila GPCR Han Is a Receptor for the Circadian Clock Neuropeptide PDF. Neuron 2005, 48, 267–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thamm, M.; Balfanz, S.; Scheiner, R.; Baumann, A.; Blenau, W. Characterization of the 5-HT1A receptor of the honeybee (Apis mellifera) and involvement of serotonin in phototactic behavior. Cell. Mol. Life Sci. 2010, 67, 2467–2479. [Google Scholar] [CrossRef]
- Abrieux, A.; Debernard, S.; Maria, A.; Gaertner, C.; Anton, S.; Gadenne, C.; Duportets, L. Involvement of the G-Protein-Coupled Dopamine/Ecdysteroid Receptor DopEcR in the Behavioral Response to Sex Pheromone in an Insect. PLoS ONE 2013, 8, e72785. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.; Agha, M.A.; Smith, R.C.; Nachman, R.J.; Marion-Poll, F.; Pietrantonio, P.V. Leucokinin mimetic elicits aversive behavior in Mosquito Aedes aegypti (L.) and inhibits the sugar taste neuron. Proc. Natl. Acad. Sci. USA 2016, 113, 6880–6885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Q.-L.; Chen, E.-H.; Jiang, H.-B.; Wei, D.-D.; Gui, S.-H.; Wang, J.-J.; Smagghe, G. Adipokinetic hormone receptor gene identification and its role in triacylglycerol mobilization and sexual behavior in the oriental fruit fly (Bactrocera dorsalis). Insect Biochem. Mol. Biol. 2017, 90, 1–13. [Google Scholar] [CrossRef]
- Grohmann, L.; Blenau, W.; Erber, J.; Ebert, P.R.; Strünker, T.; Baumann, A. Molecular and functional characterization of an octopamine receptor from honeybee (Apis mellifera) brain. J. Neurochem. 2003, 86, 725–735. [Google Scholar] [CrossRef] [Green Version]
- Johnson, E.C.; Bohn, L.M.; Taghert, P.H. Drosophila CG8422 encodes a functional diuretic hormone receptor. J. Exp. Biol. 2004, 207, 743–748. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.; Lu, H.-L.; Longnecker, M.T.; Pietrantonio, P.V. Role in Diuresis of a Calcitonin Receptor (GPRCAL1) Expressed in a Distal-Proximal Gradient in Renal Organs of the Mosquito Aedes aegypti (L.). PLoS ONE 2012, 7, e50374. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.; Pietrantonio, P.V. Calcitonin receptor 1 (AedaeGPCRCAL1) hindgut expression and direct role in myotropic action in females of the Mosquito Aedes aegypti (L.). Insect Biochem. Mol. Biol. 2013, 43, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Broeck, J.V.; Lange, A.B. Identification and Expression of the CCAP Receptor in the Chagas’ Disease Vector, Rhodnius prolixus, and Its Involvement in Cardiac Control. PLoS ONE 2013, 8, e68897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petruccelli, E.; Li, Q.; Rao, Y.; Kitamoto, T. The Unique Dopamine/Ecdysteroid Receptor Modulates Ethanol-Induced Sedation in Drosophila. J. Neurosci. 2016, 36, 4647–4657. [Google Scholar] [CrossRef] [PubMed]
- Pietrantonio, P.V.; Xiong, C.; Nachman, R.J.; Shen, Y. G protein-coupled receptors in arthropod vectors: Omics and pharmacological approaches to elucidate ligand-receptor interactions and novel organismal functions. Curr. Opin. Insect Sci. 2018, 29, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, H.; Maehara, S.; Ohta, H.; Seki, T.; Tanaka, Y. Elevenin regulates the body color through a G protein-coupled receptor NlA42 in the brown planthopper Nilaparvata lugens. Gen. Comp. Endocrinol. 2018, 258, 33–38. [Google Scholar] [CrossRef]
- Li, M.; Reid, W.R.; Zhang, L.; Scott, J.G.; Gao, X.; Kristensen, M.; Liu, N. A whole transcriptomal linkage analysis of gene co-regulation in insecticide resistant house flies, Musca domestica. BMC Genom. 2013, 14, 803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.; Zhang, Y.; You, C.; Zeng, X.; Gao, X. The role of G protein-coupled receptor-related genes in cytochrome P450-mediated resistance of the house fly, Musca domestica (Diptera: Muscidae), to imidacloprid. Insect Mol. Biol. 2019, 29, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Liu, N. Insecticide Resistance in Mosquitoes: Impact, Mechanisms, and Research Directions. Annu. Rev. Ѐntomol. 2015, 60, 537–559. [Google Scholar] [CrossRef]
- Attwood, T.K.; Findlay, J.B.C. Fingerprinting G-protein-coupled receptors. Protein Eng. Des. Sel. 1994, 7, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Kolakowski, L.F., Jr. GCRDb: A G-protein-coupled receptor database. Recept. Channels 1994, 2, 1–7. [Google Scholar] [PubMed]
- Schiöth, H.B.; Fredriksson, R. The GRAFS classification system of G-protein coupled receptors in comparative perspective. Gen. Comp. Endocrinol. 2005, 142, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.-M.; Mai, T.-L.; Chen, C.-M. Visualizing the GPCR Network: Classification and Evolution. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef]
- Adams, M.D.; Celniker, S.E.; Holt, R.A.; Evans, C.A.; Gocayne, J.D.; Amanatides, P.G.; Scherer, S.E.; Li, P.W.; Hoskins, R.A.; Galle, R.F.; et al. The Genome Sequence of Drosophila melanogaster. Science 2000, 287, 2185–2195. [Google Scholar] [CrossRef] [Green Version]
- Holt, R.A.; Subramanian, G.M.; Halpern, A.; Sutton, G.G.; Charlab, R.; Nusskern, D.R.; Wincker, P.; Clark, A.G.; Ribeiro, J.C.; Wides, R.; et al. The Genome Sequence of the Malaria Mosquito Anopheles gambiae. Science 2002, 298, 129–149. [Google Scholar] [CrossRef] [PubMed]
- Nene, V.; Wortman, J.R.; Lawson, D.; Haas, B.; Kodira, C.; Tu, Z.; Loftus, B.; Xi, Z.; Megy, K.; Grabherr, M.; et al. Genome Sequence of Aedes aegypti, a Major Arbovirus Vector. Science 2007, 316, 1718–1723. [Google Scholar] [CrossRef] [Green Version]
- Arensburger, P.; Megy, K.; Waterhouse, R.M.; Abrudan, J.; Amedeo, P.; Antelo, B.; Bartholomay, L.; Bidwell, S.; Caler, E.; Camara, F.; et al. Sequencing of Culex quinquefasciatus Establishes a Platform for Mosquito Comparative Genomics. Science 2010, 330, 86–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, J.G.; Warren, W.C.; Beukeboom, L.W.; Bopp, D.; Clark, A.G.; Giers, S.D.; Hediger, M.; Jones, A.K.; Kasai, S.; A Leichter, C.; et al. Genome of the house fly, Musca domestica L., a global vector of diseases with adaptations to a septic environment. Genome Biol. 2014, 15, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Zhao, X.; Li, M.; He, K.; Huang, C.; Zhou, Y.; Li, Z.; Walters, J.R. Insect genomes: Progress and challenges. Insect Mol. Biol. 2019, 28, 739–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brody, T.; Cravchik, A. Drosophila melanogaster G Protein–Coupled Receptors. J. Cell Biol. 2000, 150, F83–F88. [Google Scholar] [CrossRef]
- Hanlon, C.D.; Andrew, D.J. Outside-in signaling–A brief review of GPCR signaling with a focus on the Drosophila GPCR family. J. Cell Sci. 2015, 128, 3533–3542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, C.A.; Fox, A.N.; Pitts, R.J.; Kent, L.B.; Tan, P.L.; Chrystal, M.A.; Cravchik, A.; Collins, F.H.; Robertson, H.M.; Zwiebel, L.J. G Protein-Coupled Receptors inAnopheles gambiae. Science 2002, 298, 176–178. [Google Scholar] [CrossRef]
- Fan, Y.; Sun, P.; Wang, Y.; He, X.; Deng, X.; Chen, X.; Zhang, G.; Chen, X.; Zhou, N. The G protein-coupled receptors in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2010, 40, 581–591. [Google Scholar] [CrossRef]
- Anstead, C.A.; Korhonen, P.K.; Young, N.D.; Hall, R.S.; Jex, A.R.; Murali, S.C.; Hughes, D.S.; Lee, S.F.; Perry, T.; Stroehlein, A.J.; et al. Lucilia cuprina genome unlocks parasitic fly biology to underpin future interventions. Nat. Commun. 2015, 6, 7344. [Google Scholar] [CrossRef] [Green Version]
- Şahbaz, B.D. Prediction and expression analysis of G protein-coupled receptors in the laboratory stick insect, Carausius morosus. Turk. J. Boil. 2019, 43, 77–88. [Google Scholar] [CrossRef]
- Calkins, T.L.; Tamborindeguy, C.; Pietrantonio, P.V. GPCR annotation, G proteins, and transcriptomics of fire ant (Solenopsis invicta) queen and worker brain: An improved view of signaling in an invasive superorganism. Gen. Comp. Endocrinol. 2019, 278, 89–103. [Google Scholar] [CrossRef]
- Veenstra, J.A.; Rombauts, S.; Grbić, M. In silico cloning of genes encoding neuropeptides, neurohormones and their putative G-protein coupled receptors in a spider mite. Insect Biochem. Mol. Biol. 2012, 42, 277–295. [Google Scholar] [CrossRef] [Green Version]
- Aikins, M.J.; Schooley, D.A.; Begum, K.; Detheux, M.; Beeman, R.W.; Park, Y. Vasopressin-like peptide and its receptor function in an indirect diuretic signaling pathway in the red flour beetle. Insect Biochem. Mol. Biol. 2008, 38, 740–748. [Google Scholar] [CrossRef] [PubMed]
- Caers, J.; Verlinden, H.; Zels, S.; Vandersmissen, H.P.; Vuerinckx, K.; Schoofs, L. More than two decades of research on insect neuropeptide GPCRs: An overview. Front. Endocrinol. 2012, 3, 151. [Google Scholar] [CrossRef] [Green Version]
- Garczynski, S.F.; Brown, M.R.; Shen, P.; Murray, T.F.; Crim, J.W. Characterization of a functional neuropeptide F receptor from Drosophila melanogaster. Peptides 2002, 23, 773–780. [Google Scholar] [CrossRef]
- Vogel, K.J.; Brown, M.R.; Strand, M.R. Phylogenetic Investigation of Peptide Hormone and Growth Factor Receptors in Five Dipteran Genomes. Front. Endocrinol. 2013, 4, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, R.; Li, M.; Wu, Y.; Qi, Y.; Ye, G.; Huang, J. A new family of insect muscarinic acetylcholine receptors. Insect Mol. Biol. 2016, 25, 362–369. [Google Scholar] [CrossRef]
- Srivastava, D.P.; Yu, E.J.; Kennedy, K.; Chatwin, H.; Reale, V.; Hamon, M.; Smith, T.; Evans, P.D. Rapid, Nongenomic Responses to Ecdysteroids and Catecholamines Mediated by a Novel Drosophila G-Protein-Coupled Receptor. J. Neurosci. 2005, 25, 6145–6155. [Google Scholar] [CrossRef]
- Deveci, D.; Martin, F.A.; Leopold, P.; Romero, N.M. AstA Signaling Functions as an Evolutionary Conserved Mechanism Timing Juvenile to Adult Transition. Curr. Biol. 2019, 29, 813–822. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Reiher, W.; Hermann-Luibl, C.; Sellami, A.; Cognigni, P.; Kondo, S.; Helfrich-Förster, C.; Veenstra, J.A.; Wegener, C. Allatostatin A Signalling in Drosophila Regulates Feeding and Sleep and Is Modulated by PDF. PLoS Genet. 2016, 12, e1006346. [Google Scholar] [CrossRef] [Green Version]
- Zandawala, M.; Yurgel, M.E.; Texada, M.J.; Liao, S.; Rewitz, K.F.; Keene, A.C.; Nässel, D.R. Modulation of Drosophila post-feeding physiology and behavior by the neuropeptide leucokinin. PLoS Genet. 2018, 14, e1007767. [Google Scholar] [CrossRef] [Green Version]
- Yapici, N.; Kim, Y.-J.; Ribeiro, C.; Dickson, B.J. A receptor that mediates the post-mating switch in Drosophila reproductive behaviour. Nat. Cell Biol. 2007, 451, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Sellami, A.; Veenstra, J.A. SIFamide acts on fruitless neurons to modulate sexual behavior in Drosophila melanogaster. Peptides 2015, 74, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Yurgel, M.E.; Kakad, P.; Zandawala, M.; Nässel, D.R.; Godenschwege, T.A.; Keene, A.C. A single pair of leucokinin neurons are modulated by feeding state and regulate sleep–metabolism interactions. PLoS Biol. 2019, 17, e2006409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, F.; Deng, B.; Xiao, N.; Wang, T.; Li, Y.; Wang, R.; Shi, K.; Luo, D.-G.; Rao, Y.; Zhou, C. A neuropeptide regulates fighting behavior in Drosophila melanogaster. eLife 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Sano, H.; Nakamura, A.; Texada, M.J.; Truman, J.W.; Ishimoto, H.; Kamikouchi, A.; Nibu, Y.; Kume, K.; Ida, T.; Kojima, M. The Nutrient-Responsive Hormone CCHamide-2 Controls Growth by Regulating Insulin-like Peptides in the Brain of Drosophila melanogaster. PLoS Genet. 2015, 11, e1005209. [Google Scholar] [CrossRef] [Green Version]
- Iyison, N.B.; Sinmaz, M.G.; Sahbaz, B.D.; Shahraki, A.; Aksoydan, B.; Durdagi, S. In silico characterization of adipokinetic hormone receptor and screening for pesticide candidates against stick insect, Carausius morosus. J. Mol. Graph. Model. 2020, 101, 107720. [Google Scholar] [CrossRef]
- Posnien, N.; Hopfen, C.; Hilbrant, M.; Ramos-Womack, M.; Murat, S.; Schönauer, A.; Herbert, S.L.; Nunes, M.D.S.; Arif, S.; Breuker, C.J.; et al. Evolution of Eye Morphology and Rhodopsin Expression in the Drosophila melanogaster Species Subgroup. PLoS ONE 2012, 7, e37346. [Google Scholar] [CrossRef]
- Egerod, K.; Reynisson, E.; Hauser, F.; Cazzamali, G.; Williamson, M.; Grimmelikhuijzen, C.J.P. Molecular cloning and functional expression of the first two specific insect myosuppressin receptors. Proc. Natl. Acad. Sci. USA 2003, 100, 9808–9813. [Google Scholar] [CrossRef] [Green Version]
- Hauser, F.; Williamson, M.; Cazzamali, G.; Grimmelikhuijzen, C.J.P. Identifying neuropeptide and protein hormone receptors in Drosophila melanogaster by exploiting genomic data. Briefings Funct. Genom. Proteom. 2006, 4, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Bayliss, A.; Roselli, G.; Evans, P.D. A comparison of the signalling properties of two tyramine receptors from Drosophila. J. Neurochem. 2013, 125, 37–48. [Google Scholar] [CrossRef]
- Li, C.; Chen, M.; Sang, M.; Liu, X.; Wu, W.; Li, B. Comparative genomic analysis and evolution of family-B G protein-coupled receptors from six model insect species. Gene 2013, 519, 1–12. [Google Scholar] [CrossRef] [PubMed]
- De Mendoza, A.; Jones, J.W.; Friedrich, M. Methuselah/Methuselah-like G protein-coupled receptors constitute an ancient metazoan gene family. Sci. Rep. 2016, 6, 21801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hector, C.E.; Bretz, C.A.; Zhao, Y.; Johnson, E.C. Functional differences between two CRF-related diuretic hormone receptors in Drosophila. J. Exp. Biol. 2009, 212, 3142–3147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goda, T.; Doi, M.; Umezaki, Y.; Murai, I.; Shimatani, H.; Chu, M.L.; Nguyen, V.H.; Okamura, H.; Hamada, F.N. Calcitonin receptors are ancient modulators for rhythms of preferential temperature in insects and body temperature in mammals. Genes Dev. 2018, 32, 140–155. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-J.; Seroude, L.; Benzer, S. Extended Life-Span and Stress Resistance in the Drosophila Mutant methuselah. Science 1998, 282, 943–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cvejic, S.; Zhu, Z.; Felice, S.J.; Berman, Y.; Huang, X.-Y. The endogenous ligand Stunted of the GPCR Methuselah extends lifespan in Drosophila. Nat. Cell Biol. 2004, 6, 540–546. [Google Scholar] [CrossRef]
- Harmar, A.J. Family-B G-protein-coupled receptors. Genome Biol. 2001, 2, 1–3013. [Google Scholar] [CrossRef] [Green Version]
- Mezler, M.; Müller, T.; Raming, K. Cloning and functional expression of GABABreceptors from Drosophila. Eur. J. Neurosci. 2001, 13, 477–486. [Google Scholar] [CrossRef]
- Adler, P.N.; Vinson, C.; Park, W.J.; Conover, S.; Klein, L. Molecular structure of frizzled, a Drosophila tissue polarity gene. Genetics 1990, 126, 401–416. [Google Scholar] [CrossRef]
- I Strutt, D. Asymmetric Localization of Frizzled and the Establishment of Cell Polarity in the Drosophila Wing. Mol. Cell 2001, 7, 367–375. [Google Scholar] [CrossRef]
- Povelones, M.; Nusse, R. The role of the cysteine-rich domain of Frizzled in Wingless-Armadillo signaling. EMBO J. 2005, 24, 3493–3503. [Google Scholar] [CrossRef] [Green Version]
- Heuvel, M.V.D.; Ingham, P.W. smoothened encodes a receptor-like serpentine protein required for hedgehog signalling. Nat. Cell Biol. 1996, 382, 547–551. [Google Scholar] [CrossRef]
- Fan, J.; Liu, Y.; Jia, J. Hh-induced Smoothened conformational switch is mediated by differential phosphorylation at its C-terminal tail in a dose- and position-dependent manner. Dev. Biol. 2012, 366, 172–184. [Google Scholar] [CrossRef] [Green Version]
- Martin, B.L.; Kimelman, D. Wnt Signaling and the Evolution of Embryonic Posterior Development. Curr. Biol. 2009, 19, R215–R219. [Google Scholar] [CrossRef] [Green Version]
- Villarreal, C.M.; Darakananda, K.; Wang, V.R.; Jayaprakash, P.M.; Suzuki, Y. Hedgehog signaling regulates imaginal cell differentiation in a basally branching holometabolous insect. Dev. Biol. 2015, 404, 125–135. [Google Scholar] [CrossRef] [Green Version]
- Abrieux, A.; Duportets, L.; Debernard, S.; Gadenne, C.; Anton, S. The GPCR membrane receptor, DopEcR, mediates the actions of both dopamine and ecdysone to control sex pheromone perception in an insect. Front. Behav. Neurosci. 2014, 8, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lark, A.; Kitamoto, T.; Martin, J.-R. Modulation of neuronal activity in the Drosophila mushroom body by DopEcR, a unique dual receptor for ecdysone and dopamine. Biochim. Biophys. Acta (BBA)-Bioenerg. 2017, 1864, 1578–1588. [Google Scholar] [CrossRef] [PubMed]
- Lungchukiet, P.; Zhang, J.; Tobe, S.S.; Bendena, W.G. Quantification of allatostatin receptor mRNA levels in the cockroach, Diploptera punctata, using real-time PCR. J. Insect Physiol. 2008, 54, 981–987. [Google Scholar] [CrossRef] [PubMed]
- Ida, T.; Takahashi, T.; Tominaga, H.; Sato, T.; Kume, K.; Ozaki, M.; Hiraguchi, T.; Maeda, T.; Shiotani, H.; Terajima, S.; et al. Identification of the novel bioactive peptides dRYamide-1 and dRYamide-2, ligands for a neuropeptide Y-like receptor in Drosophila. Biochem. Biophys. Res. Commun. 2011, 410, 872–877. [Google Scholar] [CrossRef]
- Deng, X.; Yang, H.; He, X.; Liao, Y.; Zheng, C.; Zhou, Q.; Zhu, C.; Zhang, G.; Gao, J.; Zhou, N. Activation of Bombyx neuropeptide G protein-coupled receptor A4 via a Gαi-dependent signaling pathway by direct interaction with neuropeptide F from silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2014, 45, 77–88. [Google Scholar] [CrossRef]
- Gross, A.D.; Temeyer, K.B.; Day, T.A.; de León, A.A.P.; Kimber, M.J.; Coats, J.R. Pharmacological characterization of a tyramine receptor from the southern cattle tick, Rhipicephalus (Boophilus) microplus. Insect Biochem. Mol. Biol. 2015, 63, 47–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collin, C.; Hauser, F.; Krogh-Meyer, P.; Hansen, K.K.; De Valdivia, E.G.; Williamson, M.; Grimmelikhuijzen, C.J. Identification of the Drosophila and Tribolium receptors for the recently discovered insect RYamide neuropeptides. Biochem. Biophys. Res. Commun. 2011, 412, 578–583. [Google Scholar] [CrossRef]
- Qi, Y.-X.; Xia, R.-Y.; Wu, Y.-S.; Stanley, D.; Huang, J.; Ye, G.-Y. Larvae of the small white butterfly, Pieris rapae, express a novel serotonin receptor. J. Neurochem. 2014, 131, 767–777. [Google Scholar] [CrossRef]
- Vleugels, R.; Lenaerts, C.; Broeck, J.V.; Verlinden, H. Signalling properties and pharmacology of a 5-HT7-type serotonin receptor fromTribolium castaneum. Insect Mol. Biol. 2013, 23, 230–243. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Sun, L.; Du, H.; Moural, T.W.; Bai, H.; Liu, P.; Zhu, F. Physiological functions of a methuselah-like G protein coupled receptor in Lymantria dispar Linnaeus. Pestic. Biochem. Physiol. 2019, 160, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Khatoon, R.; Saini, S.; Vimal, D.; Patel, D.K.; Narayan, G.; Chowdhuri, D.K. Efficacy of methuselah gene mutation toward tolerance of dichlorvos exposure in Drosophila melanogaster. Free. Radic. Biol. Med. 2015, 83, 54–65. [Google Scholar] [CrossRef]
- Sun, Y.; Zou, P.; Yu, X.-Y.; Chen, C.; Yu, J.; Shi, L.-N.; Hong, S.-C.; Zhou, D.; Chang, X.-L.; Wang, W.-J.; et al. Functional characterization of an arrestin gene on insecticide resistance of Culex pipiens pallens. Parasites Vectors 2012, 5, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baron, S.; Van Der Merwe, N.A.; Madder, M.; Maritz-Olivier, C. SNP Analysis Infers that Recombination Is Involved in the Evolution of Amitraz Resistance in Rhipicephalus microplus. PLoS ONE 2015, 10, e0131341. [Google Scholar] [CrossRef] [Green Version]
- Meyer, J.M.; Ejendal, K.F.K.; Avramova, L.V.; Garland-Kuntz, E.E.; Giraldo-Calderón, G.I.; Brust, T.F.; Watts, V.J.; Hill, C.A. A “Genome-to-Lead” Approach for Insecticide Discovery: Pharmacological Characterization and Screening of Aedes aegypti D1-like Dopamine Receptors. PLoS Negl. Trop. Dis. 2012, 6, e1478. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.A.; Meyer, J.M.; Ejendal, K.F.; Echeverry, D.F.; Lang, E.G.; Avramova, L.V.; Conley, J.M.; Watts, V.J. Re-invigorating the insecticide discovery pipeline for vector control: GPCRs as targets for the identification of next gen insecticides. Pestic. Biochem. Physiol. 2013, 106, 141–148. [Google Scholar] [CrossRef]
- Nuss, A.B.; Ejendal, K.F.K.; Doyle, T.B.; Meyer, J.M.; Lang, E.G.; Watts, V.J.; Hill, C.A. Dopamine Receptor Antagonists as New Mode-of-Action Insecticide Leads for Control of Aedes and Culex Mosquito Vectors. PLoS Negl. Trop. Dis. 2015, 9, e0003515. [Google Scholar] [CrossRef]
- Hapairai, L.K.; Mysore, K.; Sun, L.; Li, P.; Wang, C.-W.; Scheel, N.D.; Lesnik, A.; Scheel, M.P.; Igiede, J.; Wei, N.; et al. Characterization of an adulticidal and larvicidal interfering RNA pesticide that targets a conserved sequence in Mosquito G protein-coupled dopamine 1 receptor genes. Insect Biochem. Mol. Biol. 2020, 120, 103359. [Google Scholar] [CrossRef] [PubMed]
- Tobin, A.B.; Butcher, A.J.; Kong, K.C. Location, location, location…site-specific GPCR phosphorylation offers a mechanism for cell-type-specific signalling. Trends Pharmacol. Sci. 2008, 29, 413–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Y.-X.; Xu, G.; Gu, G.-X.; Mao, F.; Ye, G.-Y.; Liu, W.; Huang, J. A new Drosophila octopamine receptor responds to serotonin. Insect Biochem. Mol. Biol. 2017, 90, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Van Hiel, M.B.; Vandersmissen, H.P.; Proost, P.; Broeck, J.V. Cloning, constitutive activity and expression profiling of two receptors related to relaxin receptors in Drosophila melanogaster. Peptides 2015, 68, 83–90. [Google Scholar] [CrossRef]
- Yamanaka, N.; Yamamoto, S.; Žitňan, D.; Watanabe, K.; Kawada, T.; Satake, H.; Kaneko, Y.; Hiruma, K.; Tanaka, Y.; Shinoda, T.; et al. Neuropeptide Receptor Transcriptome Reveals Unidentified Neuroendocrine Pathways. PLoS ONE 2008, 3, e3048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reim, T.; Balfanz, S.; Baumann, A.; Blenau, W.; Thamm, M.; Scheiner, R. AmTAR2: Functional characterization of a honeybee tyramine receptor stimulating adenylyl cyclase activity. Insect Biochem. Mol. Biol. 2017, 80, 91–100. [Google Scholar] [CrossRef]
- Paluzzi, J.-P.; O’Donnell, M.J. Identification, spatial expression analysis and functional characterization of a pyrokinin-1 receptor in the Chagas’ disease vector, Rhodnius prolixus. Mol. Cell. Endocrinol. 2012, 363, 36–45. [Google Scholar] [CrossRef]
- Hamoudi, Z.; Lange, A.B.; Orchard, I. Identification and Characterization of the Corazonin Receptor and Possible Physiological Roles of the Corazonin-Signaling Pathway in Rhodnius prolixus. Front. Neurosci. 2016, 10, 357. [Google Scholar] [CrossRef]
- Paluzzi, J.-P.V.; Ebhatt, G.; Wang, C.-H.J.; Ezandawala, M.; Lange, A.B.; Eorchard, I. Identification, functional characterization, and pharmacological profile of a serotonin type-2b receptor in the medically important insect, Rhodnius prolixus. Front. Neurosci. 2015, 9, 175. [Google Scholar] [CrossRef] [Green Version]
- Sha, K.; Conner, W.C.; Choi, D.Y.; Park, J.H. Characterization, expression, and evolutionary aspects of Corazonin neuropeptide and its receptor from the House Fly, Musca domestica (Diptera: Muscidae). Gene 2012, 497, 191–199. [Google Scholar] [CrossRef]
- Verlinden, H.; Vleugels, R.; Verdonck, R.; Urlacher, E.; Broeck, J.V.; Mercer, A. Pharmacological and signalling properties of a D2-like dopamine receptor (Dop3) in Tribolium castaneum. Insect Biochem. Mol. Biol. 2015, 56, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Stafflinger, E.; Hansen, K.K.; Hauser, F.; Schneider, M.; Cazzamali, G.; Williamson, M.; Grimmelikhuijzen, C.J.P. Cloning and identification of an oxytocin/vasopressin-like receptor and its ligand from insects. Proc. Natl. Acad. Sci. USA 2008, 105, 3262–3267. [Google Scholar] [CrossRef] [Green Version]
- Verlinden, H.; Vleugels, R.; Marchal, E.; Badisco, L.; Tobback, J.; Pflüger, H.-J.; Blenau, W.; Broeck, J.V. The cloning, phylogenetic relationship and distribution pattern of two new putative GPCR-type octopamine receptors in the desert locust (Schistocerca gregaria). J. Insect Physiol. 2010, 56, 868–875. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, W.; Ma, Q.; Shen, Z.; Zhang, M.; Zhou, N.; Zhang, C. Elevenin signaling modulates body color through the tyrosine-mediated cuticle melanism pathway. FASEB J. 2019, 33, 9731–9741. [Google Scholar] [CrossRef]
- Wu, S.-F.; Xu, G.; Qi, Y.-X.; Xia, R.-Y.; Huang, J.; Ye, G.-Y. Two splicing variants of a novel family of octopamine receptors with different signaling properties. J. Neurochem. 2014, 129, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Gu, G.-X.; Teng, Z.-W.; Wu, S.-F.; Huang, J.; Song, Q.-S.; Ye, G.-Y.; Fang, Q. Identification and expression profiles of neuropeptides and their G protein-coupled receptors in the rice stem borer Chilo suppressalis. Sci. Rep. 2016, 6, 28976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Wang, J.; Thakur, K.; Hu, F.; Zhang, J.-G.; Jiang, X.-F.; An, S.-H.; Jiang, H.; Jiang, L.; Wei, Z.-J. Isolation functional characterization of allatotropin receptor from the cotton bollworm, Helicoverpa armigera. Peptides 2019, 122, 169874. [Google Scholar] [CrossRef] [PubMed]
- Pietrantonio, P.V.; Jagge, C.; Taneja-Bageshwar, S.; Nachman, R.J.; Barhoumi, R. The Mosquito Aedes aegypti (L.) leucokinin receptor is a multiligand receptor for the three Aedes kinins. Insect Mol. Biol. 2005, 14, 55–67. [Google Scholar] [CrossRef]
- Esquivel, C.J.; Cassone, B.J.; Piermarini, P.M. Ade novotranscriptome of the Malpighian tubules in non-blood-fed and blood-fed Asian tiger Mosquitoes Aedes albopictus: Insights into diuresis, detoxification, and blood meal processing. PeerJ 2016, 4, e1784. [Google Scholar] [CrossRef] [Green Version]
- Munoz, S.; Guerrero, F.D.; Kellogg, A.; Heekin, A.M.; Leung, M.-Y. Bioinformatic prediction of G protein-coupled receptor encoding sequences from the transcriptome of the foreleg, including the Haller’s organ, of the cattle tick, Rhipicephalus australis. PLoS ONE 2017, 12, e0172326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verlinden, H.; Lismont, E.; Bil, M.; Urlacher, E.; Mercer, A.; Broeck, J.V.; Huybrechts, R. Characterisation of a functional allatotropin receptor in the bumblebee, Bombus terrestris (Hymenoptera, Apidae). Gen. Comp. Endocrinol. 2013, 193, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, N.; Miyoshi, K.; Horio, S.; Fukui, H. β2-Adrenergic Receptor-Mediated Histamine H1 Receptor Down-Regulation: Another Possible Advantage of β2 Agonists in Asthmatic Therapy. J. Pharmacol. Sci. 2004, 94, 449–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drake, M.T.; Shenoy, S.K.; Lefkowitz, R.J. Trafficking of G Protein–Coupled Receptors. Circ. Res. 2006, 99, 570–582. [Google Scholar] [CrossRef]
- Chen, J.-F.; Sonsalla, P.K.; Pedata, F.; Melani, A.; Domenici, M.R.; Popoli, P.; Geiger, J.; Lopes, L.V.; de Mendonça, A. Adenosine A2A receptors and brain injury: Broad spectrum of neuroprotection, multifaceted actions and “fine tuning” modulation. Prog. Neurobiol. 2007, 83, 310–331. [Google Scholar] [CrossRef] [PubMed]
- Duan, W.; Gui, L.; Zhou, Z.; Liu, Y.; Tian, H.; Chen, J.-F.; Zheng, J. Adenosine A2A receptor deficiency exacerbates white matter lesions and cognitive deficits induced by chronic cerebral hypoperfusion in mice. J. Neurol. Sci. 2009, 285, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Wang, Y.; Li, T.; Feng, X. G-protein coupled receptors (GPCRs): Signaling pathways, characterization and functions in insect physiology and toxicology. Int. J. Mol. Sci. 2021, 22, 5260. [Google Scholar] [CrossRef]
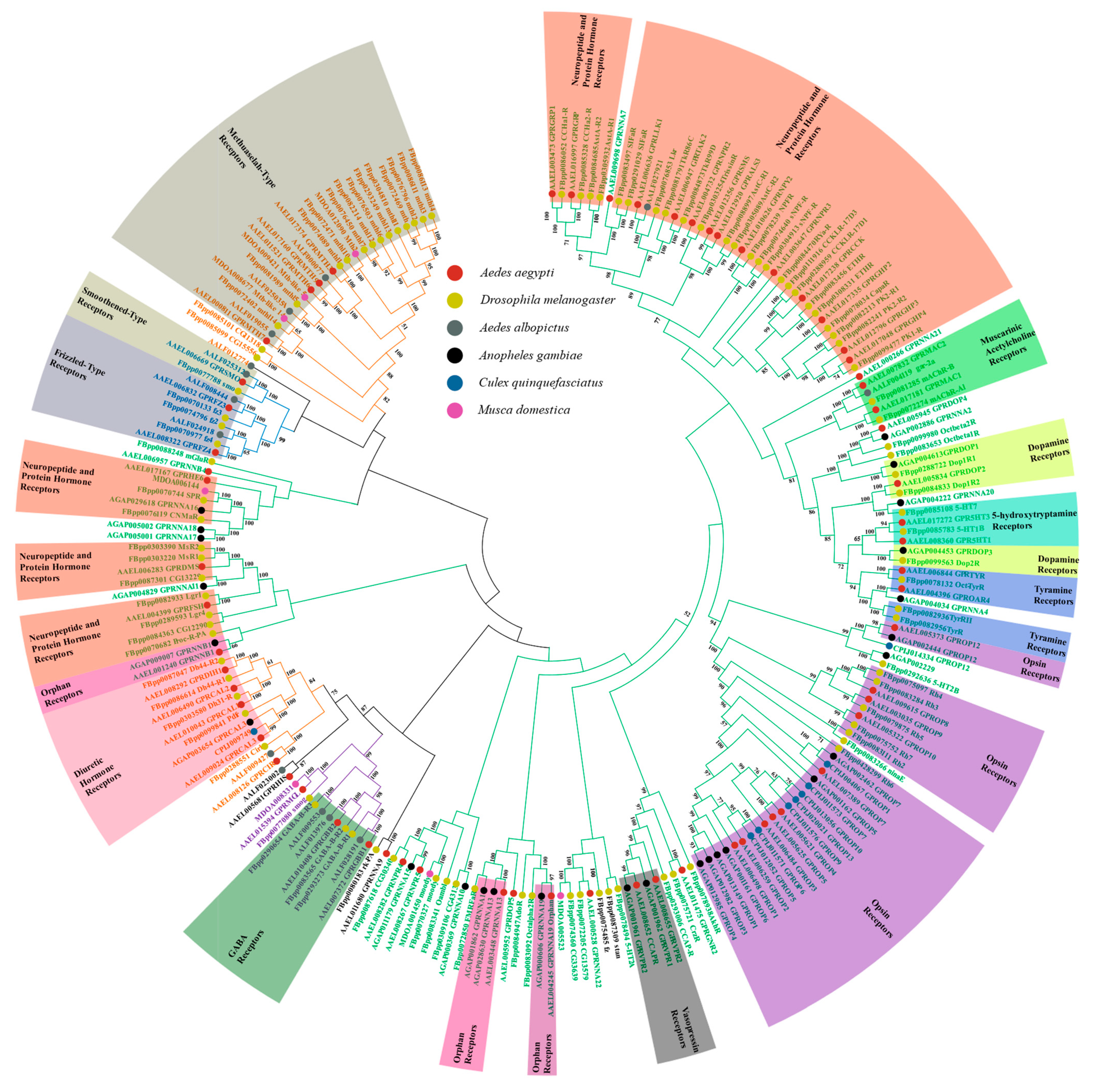
Figure 1.
Sequence comparison of GPCRs in insects. The phylogenetic tree includes 64 Ae. aegypti GPCRs, 98 D. melanogaster GPCRs, 14 Aedes albopictus GPCRs, 26 An. Gambiae GPCRs, 9 Cx. Quinquefasciatus GPCRs and 7 M. domestica GPCRs. Different colored circles represent different species. Except for the genes listed for D. melanogaster, which are taken from Flybase (https://flybase.org, accessed on 7 May 2021), all the genes are from Vectorbase (https://vectorbase.org, accessed on 7 May 2021). Sequence alignment was conducted in MEGAX and the phylogenetic tree was developed using a neighbor-joining method by MEGAX with 2000 bootstrap replicates. The evolutionary distances were computed using the p-distance method. Different classes of GPCRs are represented by colored branches—green for the Class A (the rhodopsin-like family), yellow for the Class B (the secretin receptor family), purple for the Class C (the metabotropic glutamate family) and blue for the Class F (the frizzled and smoothened receptor family). The major types of GPCRs in each of classes are labeled.
Figure 1.
Sequence comparison of GPCRs in insects. The phylogenetic tree includes 64 Ae. aegypti GPCRs, 98 D. melanogaster GPCRs, 14 Aedes albopictus GPCRs, 26 An. Gambiae GPCRs, 9 Cx. Quinquefasciatus GPCRs and 7 M. domestica GPCRs. Different colored circles represent different species. Except for the genes listed for D. melanogaster, which are taken from Flybase (https://flybase.org, accessed on 7 May 2021), all the genes are from Vectorbase (https://vectorbase.org, accessed on 7 May 2021). Sequence alignment was conducted in MEGAX and the phylogenetic tree was developed using a neighbor-joining method by MEGAX with 2000 bootstrap replicates. The evolutionary distances were computed using the p-distance method. Different classes of GPCRs are represented by colored branches—green for the Class A (the rhodopsin-like family), yellow for the Class B (the secretin receptor family), purple for the Class C (the metabotropic glutamate family) and blue for the Class F (the frizzled and smoothened receptor family). The major types of GPCRs in each of classes are labeled.


{kind=link}
{kind=link}
Table 1.
The information of GPCR genes in insect species.
Table 2.
The functions of GPCR receptors in insect physiology.
| Receptor Group | Receptor Name | Classes | Species | Function | Reference |
|---|---|---|---|---|---|
| 5-HT receptors | Trica5-HT7 R | Class A | Tribolium castaneum | Insect’s neural processes | [134] |
| Adipokinetic hormone receptor | Akh receptor | Class A | Bactrocera dorsalis | Lipid mobilization | [62] |
| Adipokinetic hormone receptor | Akh receptor | Class A | D. melanogaster | Lipid mobilization | [51] |
| Adipokinetic hormone receptor | Akh receptor | Class A | Nilaparvata lugens | Lipid mobilization | [53] |
| Allatostatin receptor | AstAR1 | Class A | D. melanogaster | Metamorphosis | [98] |
| Allatostatin receptor | DAR-1/DAR-2 | Class A | D. melanogaster | Feeding modulation | [99] |
| Allatostatin receptor | Dippu-AstR | Class A | Diploptera punctata | Juvenile hormone synthesis | [128] |
| Arginine vasopressin-like receptor | AVPL receptor | Class A | T. castaneum | Diuretic signaling pathway | [92] |
| Calcitonin receptors | GPCRCAL1 | Class A | Ae. aegypti | primary urine secretion | [66] |
| CCHa2 receptor | CCHa2-R | Class A | D. melanogaster | Insulin production | [105] |
| Diapause hormone receptor | DH-R | Class A | Ae. aegypti | Development | [39] |
| Diapause hormone receptor | Bommo-DHR | Class A | B. mori | Development | [45] |
| Diapause hormone receptor | HzDHr | Class A | Helicoverpa zea | Development | [41] |
| Dopamine receptor | Dop1R2, DmDopEcR | Class A | D. melanogaster | Morphogenesis | [34,97] |
| Dopamine receptor | DopEcR | Class A | D. melanogaster | Mushroom and locomotor activity | [127] |
| Dopamine receptor | DopEcR | Class A | D. melanogaster | Ethanol-induced sedation | [68] |
| Dopamine receptor | AipsDopEcR | Class A | Agrotis ipsilon | Sexual activity regulation | [60,126] |
| Dopamine receptor | DopEcR | Class A | Helicoverpa armigera | Morphogenesis | [35] |
| Dopamine receptor | D2R | Class A | T. castaneum | Morphogenesis | [29] |
| Leucokinin receptor | LKr | Class A | D. melanogaster | Feeding modulation | [100,103] |
| Myosuppressin receptors | CG8985/CG13803 | Class A | D. melanogaster | visceral muscles inhibition | [108] |
| Neuropeptide receptors | GPCR-B2 | Class A | B. mori | Ecdysone synthesis | [33] |
| Neuropeptide receptors | Schgr-sNPFR | Class A | Schistocerca gregaria | Feeding behavior | [50] |
| Neuropeptide Drosulfakinin receptor | CCKLR-17D1 | Class A | D. melanogaster | Fighting behavior | [104] |
| Orphan receptor | DLGR2 | Class A | D. melanogaster | Bursicon bioactivity | [25] |
| Orphan receptor | BNGR-A4 receptor | Class A | B. mori | Food intake and growth | [130] |
| Rhodopsin receptors | Rh2 | Class A | T. castaneum | Reproduction | [22] |
| Sex peptide receptor | SPR | Class A | D. melanogaster | Reproductive behavior | [101] |
| SIFamide receptor | SIFaR | Class A | D. melanogaster | Reproductive behavior | [102] |
| Tyramine receptor | TAR1 | Class A | Rhipicephalus (Boophilus) microplus | Development of antiparasitic | [131] |
| Corticotropin releasing factor receptor | CG12370 | Class B | D. melanogaster | Water balance | [113] |
| Diuretic hormone receptors | DH31R | Class B | D. melanogaster | temperature regulation and homeostasis | [114] |
| Methuselah receptor | mth | Class B | D. melanogaster | Oxidative stress resistance | [136] |
| Methuselah receptor | Ldmthl1 | Class B | Lymantria dispar | Insect longevity | [135] |
| Metabotropic GABA receptors | D-GABABR1, R2 and R3 | Class C | D. melanogaster | Central nervous system | [118] |
Table 3.
The GPCR genes that been reported in insecticide resistance.
| Receptor Name | Gene | Class | Species | Insecticide | References |
|---|---|---|---|---|---|
| Calcitonin receptor | CPIJ014419 | Class A | Cx. quinquefasciatus | Permethrin | [42] |
| Pteropsin | CPIJ014334 | Class A | Cx. quinquefasciatus | Permethrin | [42] |
| Conserved hypothetical protein | CPIJ019111 | Not clear yet | Cx. quinquefasciatus | Permethrin | [42] |
| Leucokinin receptor | LOC101891982 | Class A | M. domestica | Imidacloprid | [72] |
| Opsin receptor | LOC101900880, LOC101900148 | Class A | M. domestica | Imidacloprid | [72] |
| Methuselah-like receptor | LOC101889292, LOC101899380, LOC105262457, LOC101894839 | Class B | M. domestica | Imidacloprid | [72] |
| Dopamine receptor | LOC101896361 | Class A | M. domestica | Imidacloprid | [72] |
| Crustacean cardioactive peptide receptor | LOC101898141 | Class A | M. domestica | Imidacloprid | [72] |
| Methuselah-like GPCR | Ldmthl1 | Class B | L. dispar | Deltamethrin | [135] |
| Arrestin | HQ833831 | Cx. pipiens | Deltamethrin | [137] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, N.; Li, T.; Wang, Y.; Liu, S. G-Protein Coupled Receptors (GPCRs) in Insects—A Potential Target for New Insecticide Development. Molecules 2021, 26, 2993. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26102993
AMA Style
Liu N, Li T, Wang Y, Liu S. G-Protein Coupled Receptors (GPCRs) in Insects—A Potential Target for New Insecticide Development. Molecules. 2021; 26(10):2993. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26102993
Chicago/Turabian StyleLiu, Nannan, Ting Li, Yifan Wang, and Shikai Liu. 2021. "G-Protein Coupled Receptors (GPCRs) in Insects—A Potential Target for New Insecticide Development" Molecules 26, no. 10: 2993. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26102993