
Evaluation of Carotenoids Accumulation and Biosynthesis in Two Genotypes of Pomelo (Citrus maxima) during Early Fruit Development

 , and
, and 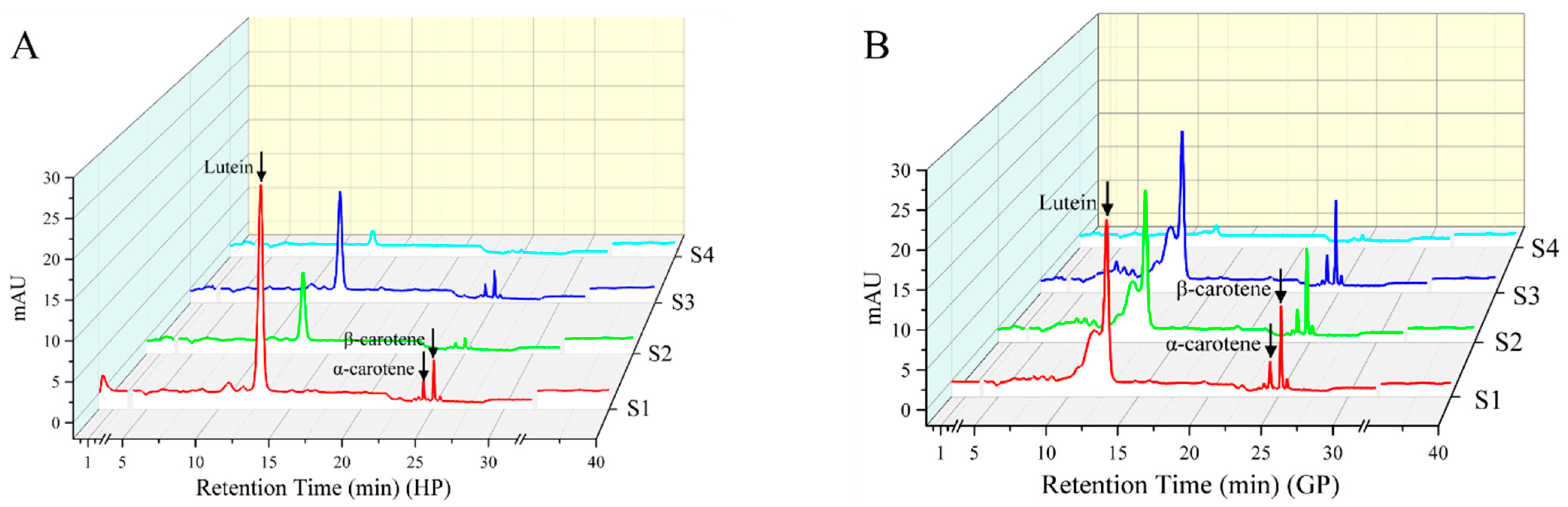{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
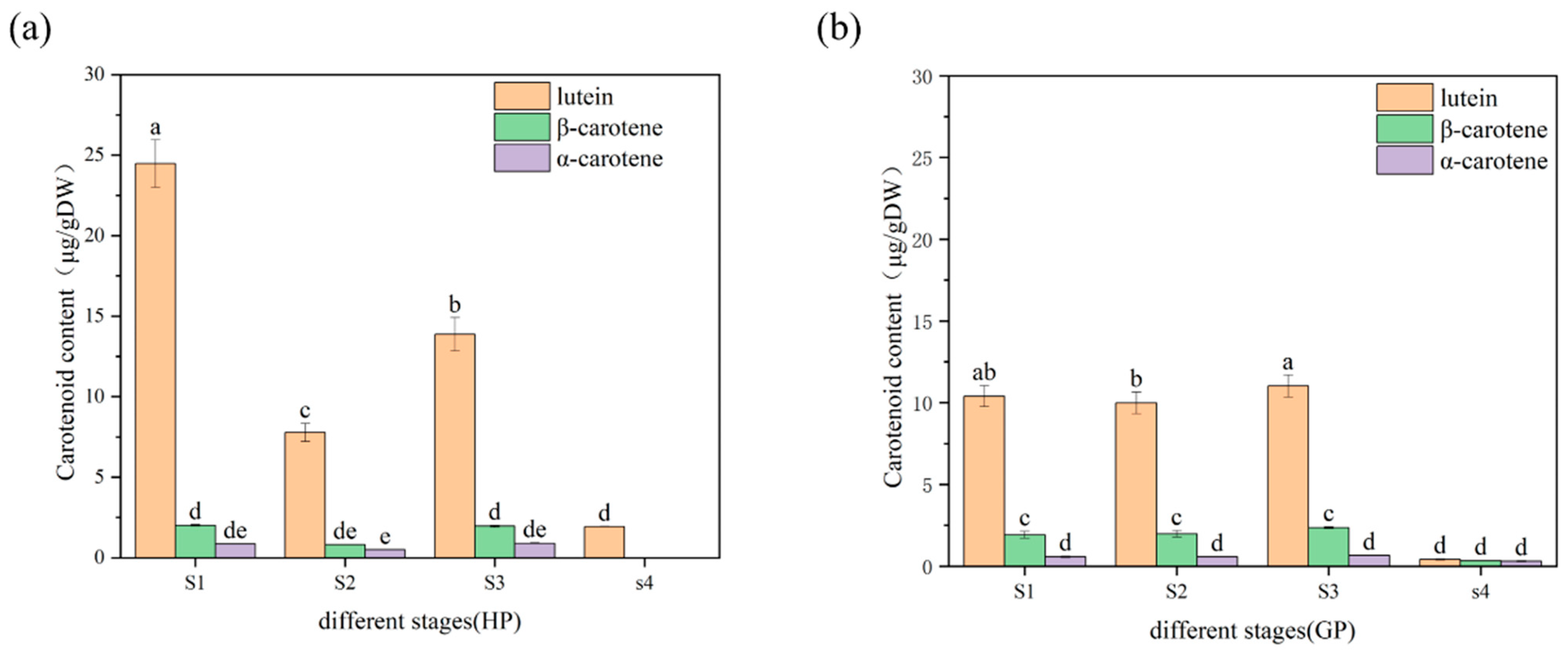
2.1. Changes of Carotenoid Profiles in Pomelo during Early Fruit Development
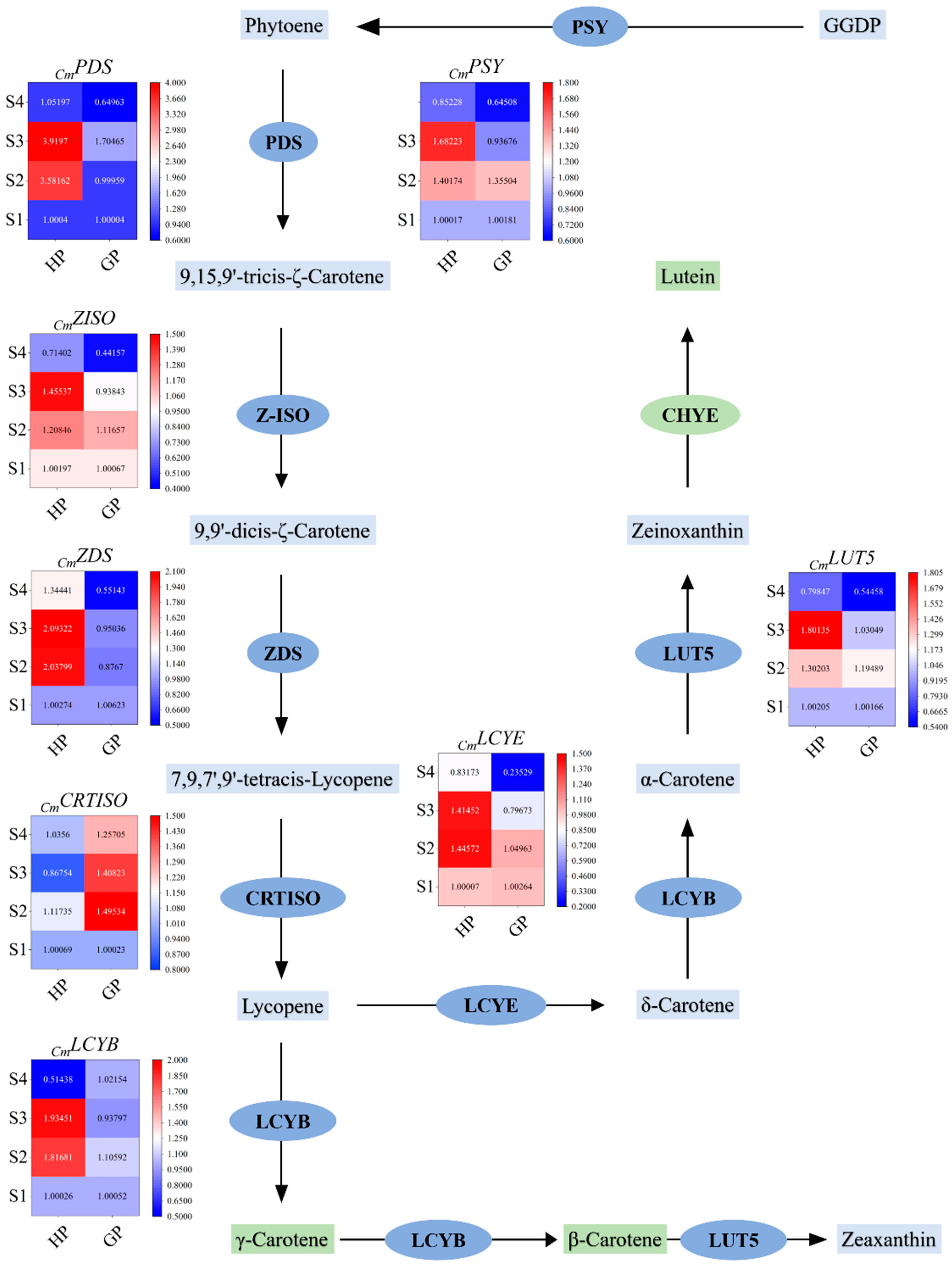
2.2. Differential Genes Expression in Carotenoid Biosynthesis Pathway
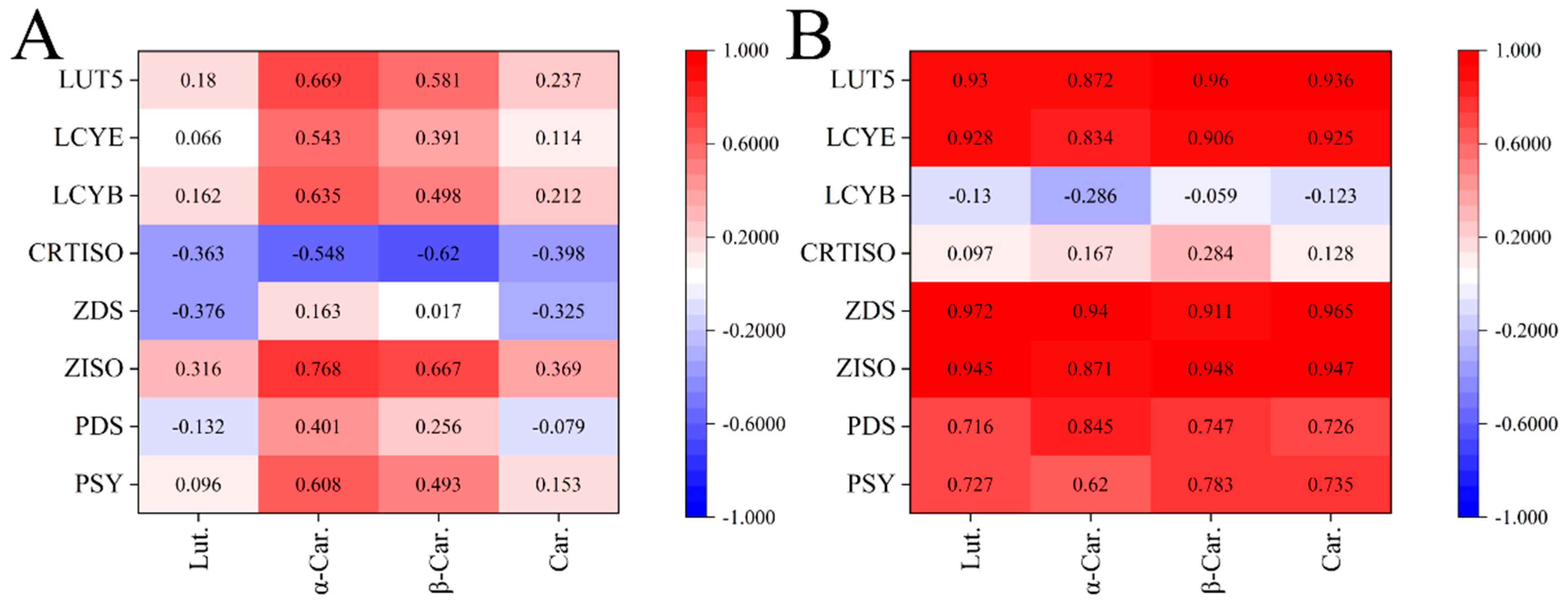
2.3. Correlations among Compositions and Gene Expressions
3. Discussion
3.1. Variations in Carotenoid Accumulation and Gene Expression
3.2. Alterations of Carotenoid Profiles and Contents during Fruit Development
4. Materials and Methods
4.1. Plant Materials and Sample Collection
4.2. Extraction of Carotenoids
4.3. Carotenoid Analysis
4.4. RT-qPCR Assay
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Huang, S.; Dong, T.; Xiong, B.; Qiu, X.; Sun, G.; Liao, L.; Fan, N.; Wang, X.; Deng, H.; He, S.; et al. Variation in the content and composition of limonoids in fruits of four pomelo varieties during fruit development: The natural debittering process in pomelo fruits. J. Food Compos. Anal. 2021, 100, 103928. [Google Scholar] [CrossRef]
- Xi, W.; Fang, B.; Zhao, Q.; Jiao, B.; Zhou, Z. Flavonoid composition and antioxidant activities of chinese local pummelo (Citrus grandis osbeck.) varieties. Food Chem. 2014, 161, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.L.; Yang, X.W.; Wu, B.F.; Shang, J.H.; Liu, Y.P.; Zhi, D.; Luo, X.D. Anti-inflammatory effect of pomelo peel and its bioactive coumarins. J. Agric. Food Chem. 2019, 67, 8810–8818. [Google Scholar] [CrossRef]
- Xu, C.J.; Fraser, P.D.; Wang, W.J.; Bramley, P.M. Differences in the carotenoid content of ordinary citrus and lycopene-accumulating mutants. J. Agric. Food Chem. 2006, 54, 5474–5481. [Google Scholar] [CrossRef]
- Li, L.; Ban, Z.; Li, X.; Wang, X.; Guan, J. Phytochemical and microbiological changes of honey pomelo (Citrus grandis L.) slices stored under super atmospheric oxygen, low-oxygen and passive modified atmospheres. Int. J. Food Sci. Technol. 2012, 47, 2205–2211. [Google Scholar] [CrossRef]
- Lü, Z.; Zhang, Z.; Wu, H.; Zhou, Z.; Yu, J. Phenolic composition and antioxidant capacities of chinese local pummelo cultivars’ peel. Hortic. Plant J. 2016, 2, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Han, R.M.; Zhang, J.P.; Skibsted, L.H. Reaction dynamics of flavonoids and carotenoids as antioxidants. Molecules 2012, 17, 2140–2160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid metabolism in plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef] [Green Version]
- Honda, M.; Kageyama, H.; Hibino, T.; Zhang, Y.; Diono, W.; Kanda, H.; Yamaguchi, R.; Takemura, R.; Fukaya, T.; Goto, M. Improved carotenoid processing with sustainable solvents utilizing z-isomerization-induced alteration in physicochemical properties: A review and future directions. Molecules 2019, 24, 2149. [Google Scholar] [CrossRef] [Green Version]
- Lado, J.; Zacarías, L.; Rodrigo, M.J. Regulation of Carotenoid Biosynthesis During Fruit Development. In Carotenoids in Nature. Subcellular Biochemistry; Stange, C., Ed.; Springer: Cham, Switzerland, 2016; Volume 79, pp. 161–198. [Google Scholar] [CrossRef]
- Lu, S.; Li, L. Carotenoid metabolism: Biosynthesis, regulation, and beyond. J. Integr. Plant Biol. 2008, 50, 778–785. [Google Scholar] [CrossRef]
- Cazzonelli, C.I.; Pogson, B.J. Source to sink: Regulation of carotenoid biosynthesis in plants. Trends Plant Sci. 2010, 15, 266–274. [Google Scholar] [CrossRef] [PubMed]
- Männistö, S.; Smith-Warner, S.A.; Spiegelman, D.; Albanes, D.; Anderson, K.; van den Brandt, P.A.; Cerhan, J.R.; Colditz, G.; Feskanich, D.; Freudenheim, J.L.; et al. Dietary carotenoids and risk of lung cancer in a pooled analysis of seven cohort studies. Cancer Epidemiol. Biomark. Prev. 2004, 13, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikoma, Y.; Matsumoto, H.; Kato, M. Diversity in the carotenoid profiles and the expression of genes related to carotenoid accumulation among citrus genotypes. Breed. Sci. 2016, 66, 139–147. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chung, S.-J.; McCullough, M.L.; Song, W.O.; Fernandez, M.L.; Koo, S.I.; Chun, O.K. Dietary carotenoids are associated with cardiovascular disease risk biomarkers mediated by serum carotenoid concentrations. J. Nutr. 2014, 144, 1067–1074. [Google Scholar] [CrossRef]
- Sugiyama, A.; Omura, M.; Matsumoto, H.; Shimada, T.; Fujii, H.; Endo, T.; Shimizu, T.; Nesumi, H.; Ikoma, Y. Quantitative trait loci (QTL) analysis of carotenoid content in Citrus fruit. J. Jpn. Soc. Hortic. Sci. 2011, 80, 136–144. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.-C.; Zhang, Y.-F.; Lin, Y.-J.; Chen, Y.; Lu, X.-K. Illumina® sequencing reveals candidate genes of carotenoid metabolism in three pummelo cultivars (Citrus maxima) with different pulp color. Int. J. Mol. Sci. 2019, 20, 2246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, N.F.A.; Shamsudin, R.; Ismail, A.; Shah, N.N.A.K.; Varith, J. Effects of drying methods on total phenolic contents and antioxidant capacity of the pomelo (Citrus grandis (L.) osbeck) peels. Innov. Food Sci. Emerg. Technol. 2018, 50, 217–225. [Google Scholar] [CrossRef]
- Lin, Z.; Li, J.; Fang, S.; Chen, T.; Chen, J. Analysis of volatile aroma components in pomelo peel tea by headspace-gas chromatography-mass spectrometry. Food Sci. 2012, 33, 256–259. [Google Scholar]
- Huang, R.; Cao, M.; Guo, H.; Qi, W.; Su, R.; He, Z. Enhanced ethanol production from pomelo peel waste by integrated hydrothermal treatment, multienzyme formulation, and fed-batch operation. J. Agric. Food Chem. 2014, 62, 4643–4651. [Google Scholar] [CrossRef]
- Chai, W.B.; Liu, X.Y.; Zou, J.C.; Zhang, X.Y.; Li, B.B.; Yin, T.T. Pomelo peel modified with acetic anhydride and styrene as new sorbents for removal of oil pollution. Carbohydr. Polym. 2015, 132, 245–251. [Google Scholar] [CrossRef]
- Xiao, L.; Ye, F.; Zhou, Y.; Zhao, G. Utilization of pomelo peels to manufacture value-added products: A review. Food Chem. 2021, 351, 129247. [Google Scholar] [CrossRef]
- Tocmo, R.; Pena-Fronteras, J.; Calumba, K.F.; Mendoza, M.; Johnson, J.J. Valorization of pomelo (Citrus grandis Osbeck) peel: A review of current utilization, phytochemistry, bioactivities, and mechanisms of action. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1969–2012. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G.; Ademosun, A.O. Shaddock peels (Citrus maxima) phenolic extracts inhibit α-amylase, α-glucosidase and angiotensin i-converting enzyme activities: A nutraceutical approach to diabetes management. Diabetes Metab. Syndr. Clin. Res. Rev. 2011, 5, 148–152. [Google Scholar] [CrossRef]
- Lan-Phi, N.; Vy, T.T. Chemical composition, antioxidant and antibacterial activities of peels’ essential oils of different pomelo varieties in the south of vietnam. Int. Food Res. J. 2015, 22, 2426–2431. [Google Scholar]
- Bramley, P.M. Carotenoid biosynthesis and chlorophyll degradation. In The Molecular Biology and Biochemistry of Fruit Ripening; Seymour, G.B., Poole, M., Giovannoni, J.J., Tucker, G.A., Eds.; Wiley: Hoboken, New Jersey, USA, 2013. [Google Scholar] [CrossRef]
- Cao, S.; Liang, M.; Shi, L.; Shao, J.; Song, C.; Bian, K.; Chen, W.; Yang, Z. Accumulation of carotenoids and expression of carotenogenic genes in peach fruit. Food Chem. 2017, 214, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Yungyuen, W.; Vo, T.T.; Uthairatanakij, A.; Ma, G.; Zhang, L.; Tatmala, N.; Kaewsuksaeng, S.; Jitareerat, P.; Kato, M. Carotenoid accumulation and the expression of carotenoid metabolic genes in mango during fruit development and ripening. Appl. Sci. 2021, 11, 4249. [Google Scholar] [CrossRef]
- Obrero, Á.; González-Verdejo, C.I.; Die, J.V.; Gómez, P.; Del Río-Celestino, M.; Román, B. Carotenogenic gene expression and carotenoid accumulation in three varieties of cucurbita pepo during fruit development. J. Agric. Food Chem. 2013, 61, 6393–6403. [Google Scholar] [CrossRef]
- Karppinen, K.; Zoratti, L.; Sarala, M.; Carvalho, E.; Hirsimäki, J.; Mentula, H.; Martens, S.; Häggman, H.; Jaakola, L. Carotenoid metabolism during bilberry (Vaccinium myrtillus L.) fruit development under different light conditions is regulated by biosynthesis and degradation. BMC Plant Biol. 2016, 16, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Mao, J.; Yan, S.; Yu, Y.; Xie, L.; Hu, J.G.; Li, T.; Abbasi, A.M.; Guo, X.; Liu, R.H. Evaluation of carotenoid biosynthesis, accumulation and antioxidant activities in sweetcorn (Zea mays L.) during kernel development. Int. J. Food Sci. Technol. 2018, 53, 381–388. [Google Scholar] [CrossRef]
- Morris, W.L.; Ducreux, L.; Griffiths, D.W.; Stewart, D.; Davies, H.V.; Taylor, M.A. Carotenogenesis during tuber development and storage in potato. J. Exp. Bot. 2004, 55, 975–982. [Google Scholar] [CrossRef] [Green Version]
- Lv, P.; Li, N.; Liu, H.; Gu, H.; Zhao, W.-E. Changes in carotenoid profiles and in the expression pattern of the genes in carotenoid metabolisms during fruit development and ripening in four watermelon cultivars. Food Chem. 2015, 174, 52–59. [Google Scholar] [CrossRef]
- Lin, C.; Wang, Z.; Mao, H. A study on the extraction and stability of the pigment from pomelo peel. Food Ferment. Ind. 1996, 4, 50–53. [Google Scholar]
- Promkaew, P.; Pongprasert, N.; Wongs-Aree, C.; Kaewsuksaeng, S.; Opio, P.; Kondo, S.; Srilaong, V. Carotenoids accumulation and carotenoids biosynthesis gene expression during fruit development in pulp of tubtim-siam pummelo fruit. Sci. Hortic. 2020, 260, 108870. [Google Scholar] [CrossRef]
- Kodentsova, V.M.; Vrzhesinskaia, O.A. About recommended daily intake values of vitamins A, E, C and B2. Vopr. Pitan. 2009, 78, 51–56. [Google Scholar]
- Zhang, L.; Zhang, Z.; Zheng, T.; Wei, W.; Zhu, Y.; Gao, Y.; Yang, X.; Lin, S. Characterization of carotenoid accumulation and carotenogenic gene expression during fruit development in yellow and white loquat fruit. Hortic. Plant J. 2016, 2, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Xiang, N.; Li, C.; Li, G.; Yu, Y.; Hu, J.; Guo, X. Comparative evaluation on vitamin e and carotenoid accumulation in sweet corn (Zea mays L.) seedlings under temperature stress. J. Agric. Food Chem. 2019, 67, 9772–9781. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Yang, X.; Hu, Y.; Gu, Q.; Chen, W.; Li, J.; Guo, X.; Liu, Y. Evaluation of Carotenoids Accumulation and Biosynthesis in Two Genotypes of Pomelo (Citrus maxima) during Early Fruit Development. Molecules 2021, 26, 5054. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26165054
Zhao Y, Yang X, Hu Y, Gu Q, Chen W, Li J, Guo X, Liu Y. Evaluation of Carotenoids Accumulation and Biosynthesis in Two Genotypes of Pomelo (Citrus maxima) during Early Fruit Development. Molecules. 2021; 26(16):5054. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26165054
Chicago/Turabian StyleZhao, Yihan, Xufeng Yang, Yuwei Hu, Qiuming Gu, Weiling Chen, Jiaqi Li, Xinbo Guo, and Yutao Liu. 2021. "Evaluation of Carotenoids Accumulation and Biosynthesis in Two Genotypes of Pomelo (Citrus maxima) during Early Fruit Development" Molecules 26, no. 16: 5054. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26165054